Introduction
Chronic lymphocytic leukemia (CLL) is the most
commonly observed hematological malignancy in Western countries
(1–3).
It represents 0.8% of all the neoplasms and almost 30% of all the
leukemia cases documented worldwide (1–3). CLL
affects particularly the elderly, since the median age at diagnosis
is 72 years, although increased morbidity among younger patients
has been recently reported (1–3). The
incidence of CLL is 2–6 cases/100,000 patients/year, and it
increases with age (1–3). The risk of malignancy development is
double in men compared with women (1–3). CLL is a
lymphoproliferative disorder that may be characterized by the
accumulation of small, homogeneous, mature cluster of
differentiation (CD)5+ CD19+ B-lymphocytes in
the peripheral blood, bone marrow and secondary lymphoid organs
(3,4).
The clinical course of CLL is diverse. Certain patients display a
favorable course of disease, with stable or slowly increasing
lymphocytosis, long survival and no requirement for treatment,
whereas other patients experience an aggressive, progressive course
of CLL that requires immediate intensive therapy (3,4). The
clinical heterogeneity observed in the course of CLL is due to
genetic and epigenetic abnormalities (2–5). The
microenvironment is additionally known to be involved in
leukemogenesis (2–5). However, the pathological mechanism of
CLL remains to be elucidated (2–5). Several
prognostic factors have been described that are useful for risk
stratification in CLL, including cytogenetic abnormalities
(deletion of the chromosomes 11q and 17p), expression of CD38 and
zeta-chain-associated protein kinase 70, and mutation status in
immunoglobulin heavy chain variable region genes (2–5). Recent
improvements in CLL treatment have led to an increased percentage
of complete remissions. However, due to the advanced age of the
patients and/or the presence of negative markers, relapse is
expected in the majority of cases (2–5). Hence,
the requirement for the development of novel therapies for the
treatment of CLL remains.
Medicinal mushrooms have been extensively used in
oriental medicine as a remedy against various disorders, including
malignancies (6). To the best of our
knowledge, of all the known species of mushrooms, 650 possess
documented medicinal properties, and ~20 are currently in clinical
use (6). Genera of mushrooms
displaying antineoplastic activity include Pleurotus, Phellinus,
Ganoderma, Agaricus, Antrodia, Clitocybe, Cordyceps, Trametes,
Calvatia, Xerocomus, Suillus, Schizophyllum, Flammulina, Funlia,
Inonotus, Lactarius, Inocybe, Albatrellus, Fomes and
Russula (6–8). Extracts from these mushrooms contain
bioactive compounds, including proteins, polysaccharides,
glycosides, fats, volatile oils, alkaloids, phenols, tocopherols,
folates, carotenoids, flavonoids, organic acids and ascorbic acid
enzymes (6–8). These extracts are able to inhibit
mitosis and angiogenesis, induce apoptosis and restrain
proliferation of neoplastic cells (6–8).
Laccase [benzenediol:oxygen oxidoreductase, enzyme
commission number, 1.10.3.2 (http://www.kegg.jp/dbget-bin/www_bget?ec:1.10.3.2);
LAC] is part of the largest subgroup of blue multicopper oxidases,
and exhibits the distinctive redox ability of copper ions, since it
is capable of catalyzing the oxidation of an extensive range of
aromatic substrates concomitantly with the reduction of molecular
oxygen to water (9,10). The distribution of LAC is widespread
among plants, fungi and bacteria (7).
In particular, white-rot fungi have been identified to be the most
efficient LAC producers (7,11–13).
Cerrena unicolor has been established as the most effective
fungal source of extracellular (ex)-LAC, with the highest activity
reported to be 60,000 nkat/l (14).
C. unicolor ex-LAC has been utilized in biodegradation,
bioremediation, delignification and decolorization, although no
data regarding its anticancer activity have been published to date
(15).
The present study aimed to investigate the in
vitro cytotoxicity of C. unicolor ex-LAC against
leukemic cells. CLL cells were used as a model of disease in order
to examine novel therapeutic agents, since they consist of two
compartments: i) an accumulation compartment in the peripheral
blood, followed by the spleen and liver; and ii) a proliferation
compartment in the lymph nodes and bone marrow (4). No transgenic model or cell line of CLL
currently exists (4). Therefore,
several hematological cell lines were used in the present study, in
addition to primary CLL cells, to evaluate the cytotoxic activity
of C. unicolor ex-LAC against leukemic cells.
Materials and methods
Strain, medium, growth processing and
preliminary separation of ex-LAC
C. unicolor (Bull.ex.Fr.) Murr, No. 139, was
acquired from the Regensburg Culture Collection, Archaea Centre,
University of Regensburg (Regensburg, Germany) and deposited in the
fungal collection at the Department of Biochemistry of Maria
Curie-Skłodowska University (Lublin, Poland) under the strain no.
139 (internal transcribed spacer sequence deposited in the GenBank
database under the accession no. DQ056858) (16). Fermenter scale cultivation was
performed at 28°C in a BioFlo® 310 fermenter (New Brunswick
Scientific; Eppendorf, Hamburg, Germany) containing 2 l Lindenberg
and Holm medium (Sigma-Aldrich, St. Louis, MO, USA) sterilized at
121°C for 30 min (14). The fermenter
was inoculated with crumbled fungal mats (10% of total volume),
aerated at 1 l air/min and stirred at 100 rpm. Antifoam B emulsion
(Sigma-Aldrich) was occasionally added to the fermenter cultures in
order to disperse any foam formation. Cultures (10-day-old) were
filtered through Miracloth (Calbiochem; EMD Millipore, Billerica,
MA, USA) and utilized for subsequent assays. The beginning of the
idiophase was determined according to the protocol recommended by
Betina (17). The culture liquid
obtained following mycelium separation was centrifuged (Sigma
4–16KS; SciQuip Ltd., Shropshire, UK) at 10,000 × g for 15 min. The
supernatant was subdivided into two fractions via ultrafiltration,
using Ultracel-10 membranes (EMD Millipore) incorporated into
Pellicon 2 Mini cassettes holders (EMD Millipore). The fraction
containing substances of molecular weight >10 kDa was used as
the source of crude ex-LAC, and subsequently purified according to
modified methods described by Rogalski and Janusz (16).
Preparation of fungal ex-LAC
Chromatography was performed using a chromatographic
fast protein liquid chromatography system (BioLogic™ Low-Pressure
Liquid Chromatography System; Bio-Rad Laboratories, Inc., Hercules,
CA, USA). The supernatant of culture fluid that was concentrated on
the aforementioned Pellicon 2 Mini ultrafiltration system was next
loaded on a diethylaminoethyl cellulose (DEAE)-Sepharose column of
2.5×15.0 cm (GE Healthcare Life Sciences, Chalfont, UK), which was
pre-equilibrated with 20 mM Tris-HCl buffer (Sigma-Aldrich), pH
6.5. Proteins were eluted using a 0.1–0.5 M linear gradient of NaCl
(Sigma-Aldrich) at a flow rate of 1 ml/min. The fraction containing
LAC activity was collected and desalted on a Sephadex G-50 column
of 5.0×20.0 cm (GE Healthcare Life Sciences). The purification
processes were performed at 4°C. The semi-pure ex-LAC solution was
next lyophilized using FreeZone 12 Liter Console Freeze Dry System
(Labconco Corporation, Kansas City, MO, USA).
Cell lines
Human chronic myeloid leukemia in blast crisis
(K562) and human multiple myeloma (RPMI 8226) cell lines were
acquired from the German Collection of Microorganisms and Cell
Cultures, Leibniz-Institute DSMZ (Brunswick, Germany). Human acute
promyelocytic leukemia (HL-60) and human T cell leukemia (Jurkat)
cell lines were purchased from the American Type Culture Collection
(Manassas, VA, USA). All the cell lines were cultured in RPMI 1640
medium (Biochrom; Merck Millipore, Darmstadt, Germany) supplemented
with 10% fetal bovine serum (Biochrom; Merck Millipore) and 1%
Penicillin-Streptomycin-Neomycin Solution (Sigma-Aldrich), and
maintained in a humidified atmosphere with 5% CO2 at
37°C (New Brunswick™ Galaxy® 170R CO2 Incubator;
Eppendorf).
Peripheral blood samples
Upon obtaining written informed consent, peripheral
blood was extracted from nine patients with CLL, whose clinical
characteristics are summarized in Table
I. This study was approved by the Ethics Committee of the
Medical University of Lublin (Lublin, Poland) (no.
KE-0254/116/2012). Peripheral blood mononuclear cells (PBMCs) were
isolated using Bicoll Density Centrifugation Media (Biochrom,
Berlin, Germany). The viability of cells was >95%, as determined
by trypan blue (Sigma-Aldrich) staining and quantification in a
Neubauer chamber (Zeiss AG, Oberkochen, Germany). Immediately
following isolation, cells were utilized in a
2,3-bis-(2-methoxy-4-nitro-5-sulfophenyl)-2H-tetrazolium-5-carboxanilide
(XTT; Sigma-Aldrich) assay.
 | Table I.Clinical characteristics of patients
with chronic lymphocytic leukemia. |
Table I.
Clinical characteristics of patients
with chronic lymphocytic leukemia.
| Clinical
characteristic | Total patients, n
(%) |
|---|
| Median age, years
(range) | 65 (47–82) |
| Gender |
|
|
Female | 6 (66.67%) |
|
Male | 3 (33.33%) |
| Rai stage |
|
| 0 | 4 (44.44%) |
| I | 4 (44.44%) |
| II | 1 (11.11%) |
| ZAP-70, cut-off
20% |
|
|
Positive | 2 (22.22%) |
|
Negative | 2 (22.22%) |
| Not
available | 5 (55.56%) |
| CD38, cut-off
30% |
|
|
Positive | 4 (44.44%) |
|
Negative | 5 (55.56%) |
| IGHV
status |
|
|
Mutated | 3 (33.33%) |
| Not
mutated | 4 (44.44%) |
| Not
available | 2 (22.22%) |
| Cytogenetics |
|
|
Del(13q) | 3 (33.33%) |
|
Del(17p) | 1 (11.11%) |
| No
changes | 6 (66.67%) |
Ex-LAC activity assay
Ex-LAC activity in the culture supernatant was
measured spectrophotometrically at 525 nm with UV-160A
spectrophotometer (Shimadzu Corporation, Tokyo, Japan), using
syringaldazine (Sigma-Aldrich) as a substrate (18). Enzyme and substrate blanks were
included in the assay. A unit (nkat) of ex-LAC activity was defined
as the amount of enzyme catalyzing the production of 1 nmol/sec of
colored product (quinone, εM = 65,000/M/cm) at 25°C and
pH 7.4. The enzymatic activity of ex-LAC was expressed as nkat/mg
of protein.
Protein concentration
Protein concentrations were determined using the
Bradford reagent (Sigma-Aldrich), which is based on the color
change of Coomassie brilliant blue G-250 dye in response to various
concentrations of protein (the dye binds to primarily basic,
especially arginine and aromatic amino acid residues), and bovine
serum albumin (Sigma-Aldrich) as a standard (19). Protein concentration was determined
following the addition of an acidic dye to the protein solution,
and absorbance was measured at a wavelength of 595 nm using a
UV-160A spectrophotometer (Shimadzu Corporation).
XTT assay
The cytotoxic effect of ex-LAC was measured using
the In Vitro Toxicology Assay Kit, XTT based
(Sigma-Aldrich). All cells were suspended in X–VIVO™ medium with or
without phenol red (Lonza Group Ltd., Basel, Switzerland) and
gentamicin (Lonza Group Ltd.). Cell lines were seeded onto 96-well
plates (GenoPlast Biochemicals, Rokocin, Poland) at a concentration
of 5×104 cells/100 µl/well. PBMCs obtained from patients
with CLL were dispensed at a concentration of 5×105
cells/100 µl/well. Cells were exposed to different concentrations
of ex-LAC ranging from 666.667 to 0.007 µg/ml. As a negative
control, live cells were used, while as a positive control, cells
treated with 0.1% Triton X-100 (Sigma-Aldrich) were used. XTT (25
µl) was added to all samples, prior to be incubated for 48 h in a
humidified atmosphere with 5% CO2 at 37°C. Optical
densities (OD) were measured at 450 nm, using a background
wavelength of 690 nm. OD measurements were performed every 24 h.
Each sample was assayed in triplicate, and the entire experiment
was performed three times. The results were expressed as half
maximal inhibitory concentration (IC50), and the
percentage of cytotoxicity was calculated as follows:
Cytotoxicity =
[1-(ODs-ODb)/(ODc-ODb)]
× 100%,
where ODs is the OD value of the assayed
sample, ODb is the OD value of the positive control and
ODc is the OD value of the live cells used as negative
control.
Apoptosis analysis
For apoptosis analysis, Jurkat cells were treated
with different concentrations of ex-LAC, and 0.1% Triton X-100 was
added as a positive control. Following 48 h of incubation, all
cells were washed using phosphate-buffered saline (PBS; Biochrom;
Merck Millipore), resuspended in a binding buffer (Sigma-Aldrich)
and stained with 5 µl Annexin V-fluorescein isothiocyanate (FITC)
and 10 µl propidium iodide (PI), according to the manufacturer's
protocol of the Annexin V-FITC Apoptosis Detection Kit
(Sigma-Aldrich). Cells were incubated for 10 min in the dark, and
immediately analyzed with FACSCalibur™ (BD Biosciences, San Jose,
CA, USA). Live cells served as a negative control.
Visualization of Jurkat and RPMI 8226
cells using fluorescence and scanning electron microscopy
(SEM)
Jurkat and RPMI 8226 cells were incubated with
ex-LAC for 48 h. Following incubation, all cells were washed with
PBS, resuspended in a binding buffer and stained with 5 µl Annexin
V-FITC and 10 µl PI, which were provided in the Annexin V-FITC
Apoptosis Detection Kit. Cells subjected to SEM (VEGA3 LM; Tescan,
Brno, Czech Republic) were pre-treated with 2.5% (v/v)
glutaraldehyde (Sigma-Aldrich) for 2 h and 1% osmium tetroxide
(Sigma-Aldrich) for 30 min. The osmium-fixed cells were
subsequently dehydrated using a series of graded ethanol solutions
(Sigma-Aldrich) ranging from 10% to absolute ethanol, followed by
air drying at room temperature and coating with gold
(Sigma-Aldrich). Magnification ×5,000 was used to observe the
cells.
Results
C. unicolor ex-LAC preparation
demonstrates enzymatic activity
Ex-LAC was isolated and partially purified from the
idiophasic cultures of C. unicolor according to the method
previously published by Rogalski and Janusz (16). Purification of C. unicolor ex-LAC was
performed using ion exchange chromatography on DEAE-Sepharose and
Sephadex G-50 columns. The enzyme isoforms Ia1, Ia2, Ib and IIa1
were recovered with a 65–92-fold increase in specific activity and
a yield of 6.7, 27.5, 9.7 and 21.0%, respectively. The isoelectric
points were in the range of 4.7–4.2 and the carbohydrate content in
the purified enzymes was between 1.6 and 3.5% (16). A total of 1 mg lyophilized ex-LAC
isoform mixture dissolved in 1 ml Milli-Q water (EMD Millipore)
possessed an activity of 1,150,110 nkat and a protein concentration
of 329 µg/ml.
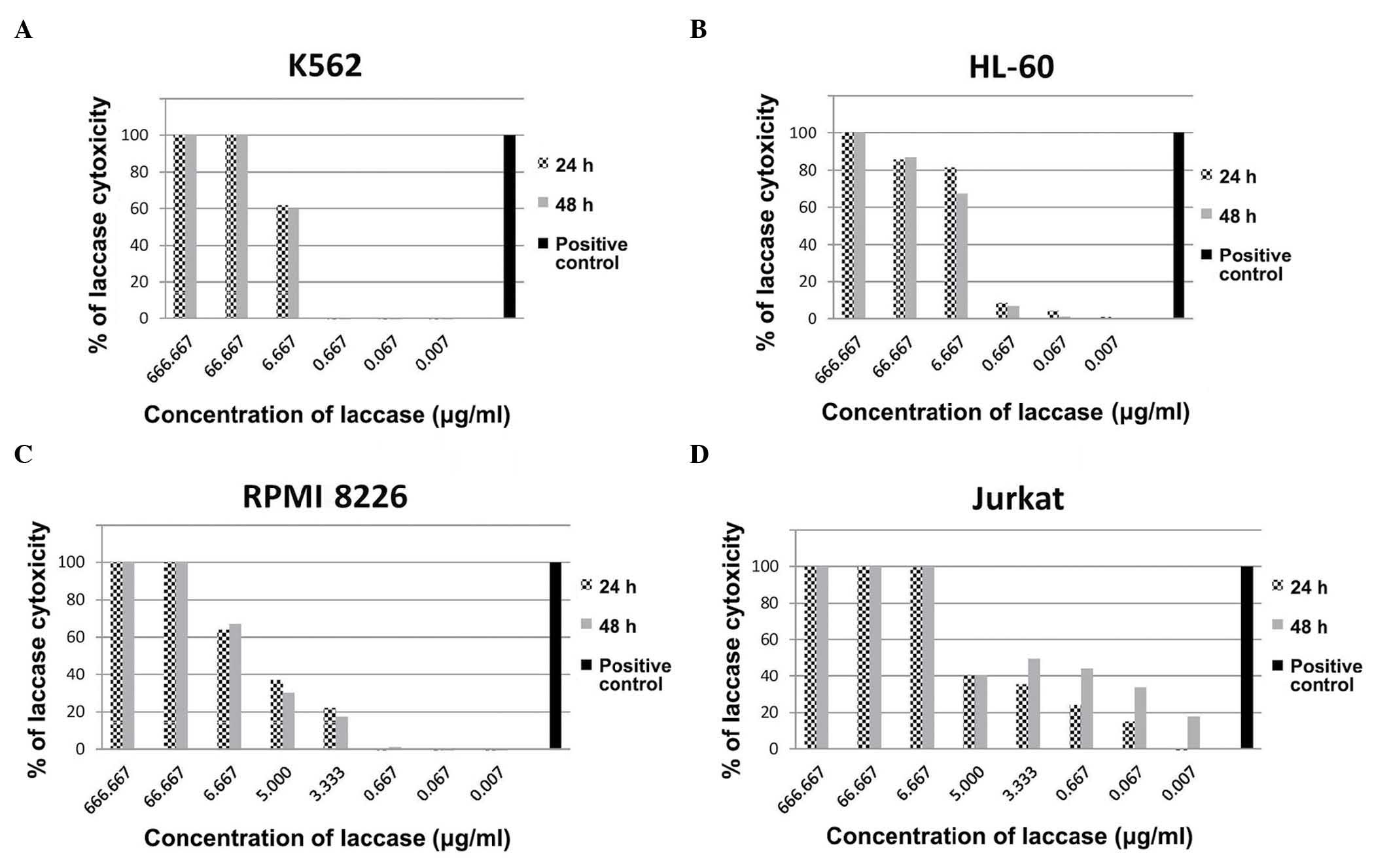
Cytotoxicity of C. unicolor ex-LAC on
the K562 cell line increases with concentration
To evaluate the cytotoxic effect of ex-LAC on cell
lines, cells were treated with a range of concentrations of ex-LAC
(666.667, 66.667, 6.667, 0.667, 0.067 and 0.007 µg/ml) for 24 and
48 h, and XTT assays were subsequently performed. The results
indicated that the cytotoxicity of ex-LAC on K562 cells increased
with concentration (Fig. 1A).
Cytotoxicity of 100.00% against K562 cells was observed following
24 and 48 h of incubation with 666.667 and 66.667 µg/ml ex-LAC. At
a concentration of 6.667 µg/ml, the cytotoxic effect of ex-LAC on
K562 cells following 24 h of incubation was 61.78%, and following
48 h of incubation the cytotoxic effect was 59.80%. No cytotoxic
effect was detected on K562 cells following 24 and 48 h of
incubation with 0.667, 0.067 and 0.007 µg/ml C. unicolor ex-LAC.
The IC50 values of ex-LAC on K562 cells following 24 and
48 h of treatment were 0.8 and 1.0 µg/ml, respectively, as
determined by XTT assay.
Concentration-dependent cytotoxic
activity of C. unicolor ex-LAC is observed in the HL-60 cell
line
The cytotoxicity of various concentrations of ex-LAC
on HL-60 cells was established using an XTT assay. Following 24 h
of incubation of HL-60 cells with 666.667, 66.667, 6.667, 0.667,
0.067 and 0.007 µg/ml ex-LAC, the percentage of cytotoxicity
observed was 100.00, 85.63, 81.55, 8.53, 4.39 and 1.11%,
respectively (Fig. 1B). The
IC50 value corresponding to 24-h treatment was 0.5
µg/ml. Similar results were obtained following 48 h of HL-60
incubation with ex-LAC. A total of 100.00% cytotoxicity was
observed following incubation with 666.667 µg/ml ex-LAC for 48 h,
87.00% following incubation with 66.667 µg/ml and 67.51% following
incubation with 6.667 µg/ml ex-LAC. Cytotoxicity of 6.94, 1.34 and
0.32% was observed following 48 h of incubation with 0.667, 0.067
and 0.007 µg/ml ex-LAC, respectively (Fig. 1B). Following 48 h of treatment, the
IC50 value of ex-LAC on HL-60 cells was 0.9 µg/ml.
C. unicolor ex-LAC decreases viability
of the RPMI 8226 cell line
The RPMI 8226 cell line was incubated with a series
of dilutions of ex-LAC, and the cytotoxic effect caused by ex-LAC
in these cells was also observed to be dose-dependent (Fig. 1C). Following 24- and 48-h treatment of
cells with 666.667 and 66.667 µg/ml ex-LAC, 100.00% cytotoxicity
was observed, while 6.667 µg/ml ex-LAC achieved 63.84 and 66.96%
cytotoxicity following 24 and 48 h of incubation, respectively.
Cytotoxicity of 36.77 and 30.20% was observed following treatment
with 5.000 µg/ml ex-LAC for 24 and 48 h, respectively. Following 24
and 48 h of incubation with 3.333 µg/ml ex-LAC, the observed
cytotoxicity on RPMI 8226 cells was 21.97 and 17.59%, respectively.
Cytotoxicity of 1.03% was noted following 48 h of treatment with
0.667 µg/ml ex-LAC. By contrast, no effect was observed following
24 h of incubation with 0.667 µg/ml ex-LAC. Similarly, no decrease
in RPMI 8226 cell viability was observed following 24 and 48 h of
incubation with 0.067 and 0.007 µg/ml ex-LAC. The IC50
values obtained following 24- and 48-h treatment with ex-LAC were
0.9 and 1.1 µg/ml, respectively. The IC50 values
following 24- and 48-h treatment with ex-LAC were similar for all
cell lines used (K562, 0.8 and 1.0 µg/ml; HL-60, 0.5 and 0.9 µg/ml;
RPMI8226, 0.9 and 1.1 µg/ml; and Jurkat, 0.8 and 0.4 µg/ml). The
differences in IC50 values observed result from the fact
that each cell line originates from a different neoplasm and
different cell lineages. Jurkat cells are derived from a human T
cell leukemia line and T cells are considered the most treatment
resistant cell type (20).
Cytotoxic activity of C. unicolor
ex-LAC is observed in the Jurkat cell line
The cytotoxic activity of ex-LAC was additionally
investigated in the Jurkat cell line via XTT assay (Fig. 1D). Following 24 h of incubation with
666.667 and 66.667 µg/ml ex-LAC, 100.00% cytotoxicity was observed
in Jurkat cells. Treatment with 6.667 µg/ml ex-LAC resulted in
99.28% cytotoxicity, while 24-h treatment with 5.000 and 3.333
µg/ml ex-LAC resulted in 40.39 and 35.22% cytotoxicity,
respectively. Cytotoxicity of 24.00 and 15.14% was observed
following 24-h incubation with 0.667 and 0.067 µg/ml ex-LAC,
respectively. No effect on Jurkat cell viability was observed with
a concentration of ex-LAC of 0.007 µg/ml. By contrast, incubation
with 666.667 µg/ml ex-LAC for 48 h resulted in 100.00%
cytotoxicity. Similarly, concentrations of 66.667 and 6.667 µg/ml
had a cytotoxic effect on Jurkat cells of 99.85 and 100.00%,
respectively. Similar results were obtained following 48 h of
incubation with 5.000, 3.333 and 0.667 µg/ml ex-LAC, which achieved
cytotoxicities of 40.35, 49.46 and 44.36%, respectively. Jurkat
cells treated with 0.067 and 0.007 µg/ml ex-LAC for 48 h exhibited
a percentage of cell death of 33.80 and 17.91%, respectively. The
IC50 values for 24- and 48-h incubation with ex-LAC were
0.8 and 0.4 µg/ml, respectively.
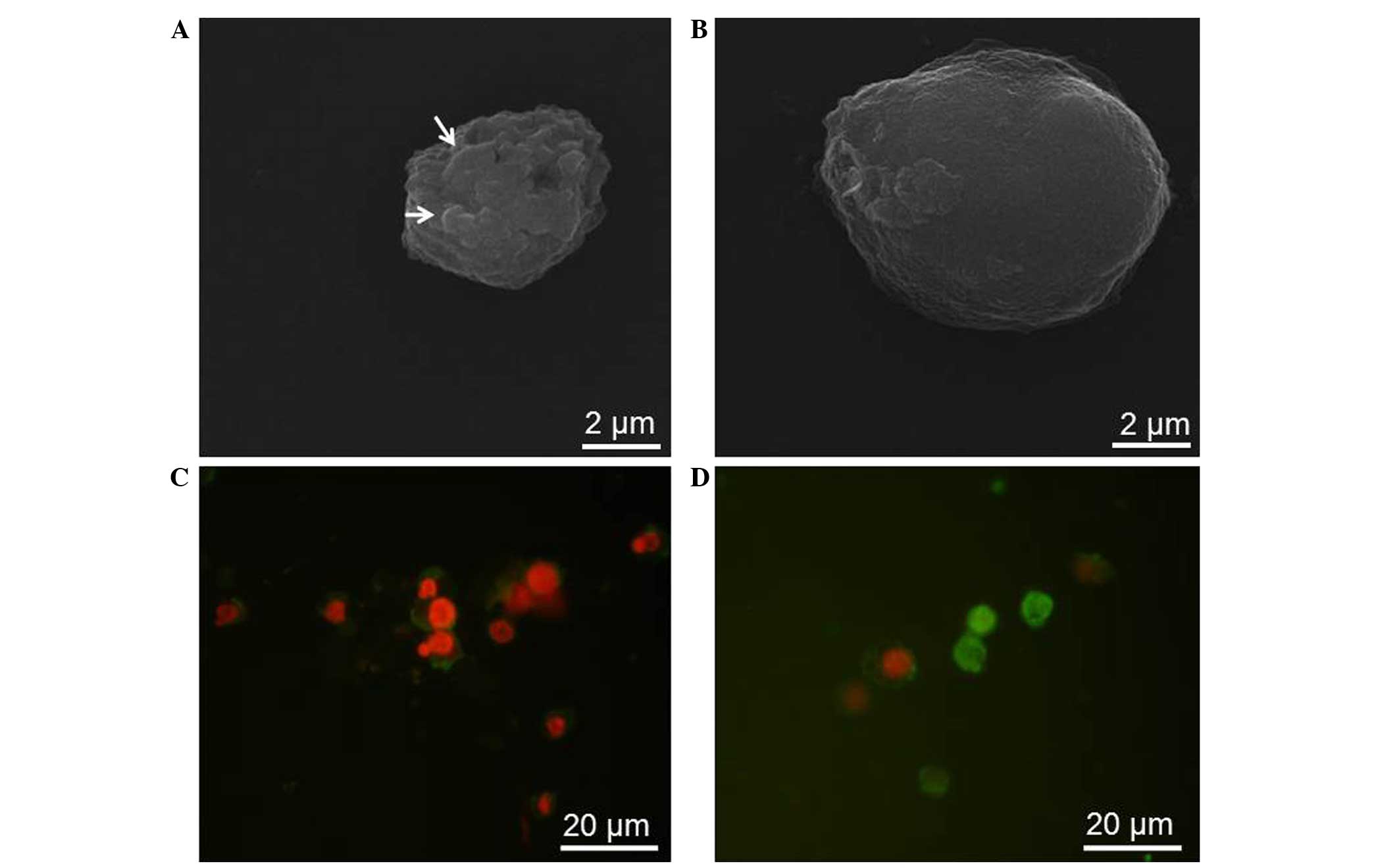
C. unicolor ex-LAC induces
morphological changes in the RPMI 8226 cell line
Morphological changes in RPMI 8226 cells following
48 h of treatment with various concentrations of ex-LAC were
observed under fluorescence microscopy and SEM (Fig. 2). Contrarily to untreated RPMI 8226
cells, which possessed a regular, oval shape (Fig. 2A), SEM revealed cell volume shrinkage,
membrane blebbing and apoptotic body formation in RPMI 8226 cells
treated with ex-LAC (Fig. 2B). For
apoptosis detection, ex-LAC-treated and untreated cells were
stained with Annexin V and PI, and visualized under a fluorescence
microscope (E-800; Nikon Corporation, Tokyo, Japan). In comparison
with untreated cells (Fig. 2C), cells
treated with ex-LAC displayed apoptotic-like changes, including
condensation and fragmentation of nuclei, in addition to cytoplasm
condensation (Fig. 2D).
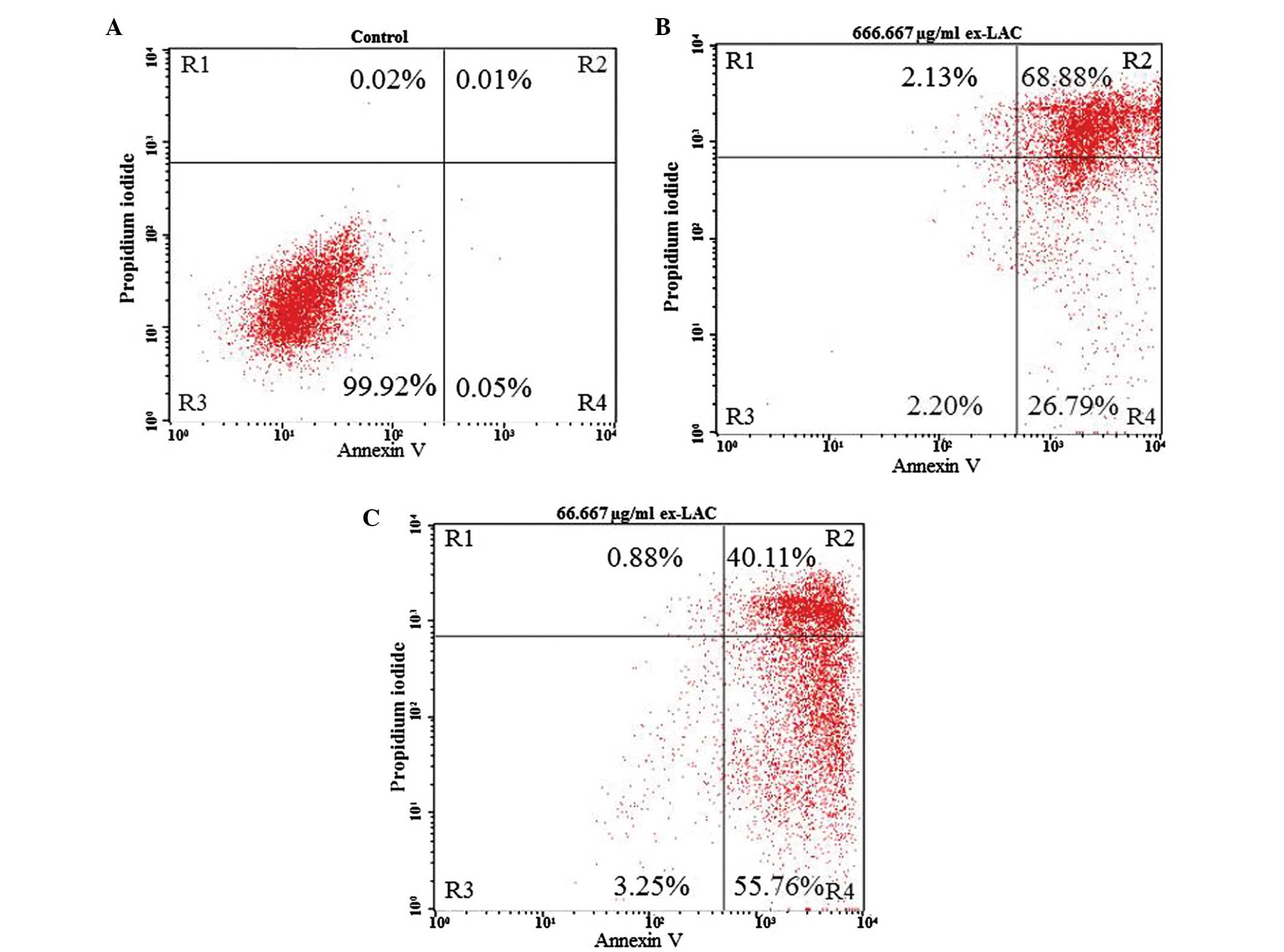
Apoptotic changes in the Jurkat cell
line are induced by ex-LAC
In order to confirm the results obtained by XTT
assay, Jurkat cells were analyzed using Annexin V/PI staining and
flow cytometry following 48 h of incubation with ex-LAC (Table II and Fig.
3). Compared with the control, the percentage of live cells
(R4) was decreased, and the percentage of apoptotic cells (R3+R5)
was increased for all the concentrations of enzyme tested (Table II). The frequency of live and
apoptotic cells in the untreated control was 99.93 and 0.05%,
respectively (Fig. 3A). At a
concentration of 666.667 µg/ml, the percentage of live vs.
apoptotic cells was 2.20 vs. 95.67% (Fig.
3B), while for a concentration of 66.667 µg/ml these
percentages were 3.25 and 95.87%, respectively (Fig. 3C). At a concentration of ex-LAC of
6.667 µg/ml the percentage of live vs. apoptotic cells was 0.82 vs.
98.00%, while it was 1.24 vs. 97.05% at 5.000 µg/ml ex-LAC. At a
concentration of 3.333 µg/ml, the percentage of live vs. apoptotic
cells were 1.65 vs. 96.76%, while these percentages were 0.68 vs.
98.90%, 7.95 vs. 90.24% and 8.86 vs. 88.98% for concentrations of
ex-LAC of 0.667, 0.067 and 0.007 µg/ml, respectively (Table II).
| Table II.Percentage of live, apoptotic and
necrotic Jurkat cells following 48-h treatment with Cerrena
unicolor ex-LAC, as determined by flow cytometry using Annexin
V and propidium iodide staining. |
Table II.
Percentage of live, apoptotic and
necrotic Jurkat cells following 48-h treatment with Cerrena
unicolor ex-LAC, as determined by flow cytometry using Annexin
V and propidium iodide staining.
| Concentration of
ex-LAC, µg/ml | Live cells, % | Apoptotic cells,
% | Necrotic cells,
% |
|---|
| 666.667 | 2.20 | 95.67 | 2.13 |
| 66.667 | 3.25 | 95.87 | 0.88 |
| 6.667 | 0.82 | 98.00 | 1.18 |
| 5.000 | 1.24 | 97.05 | 1.71 |
| 3.333 | 1.65 | 96.76 | 1.59 |
| 0.667 | 0.68 | 98.90 | 0.42 |
| 0.067 | 7.95 | 90.24 | 1.81 |
| 0.007 | 8.86 | 88.98 | 2.16 |
| Control | 99.93 |
0.05 | 0.02 |
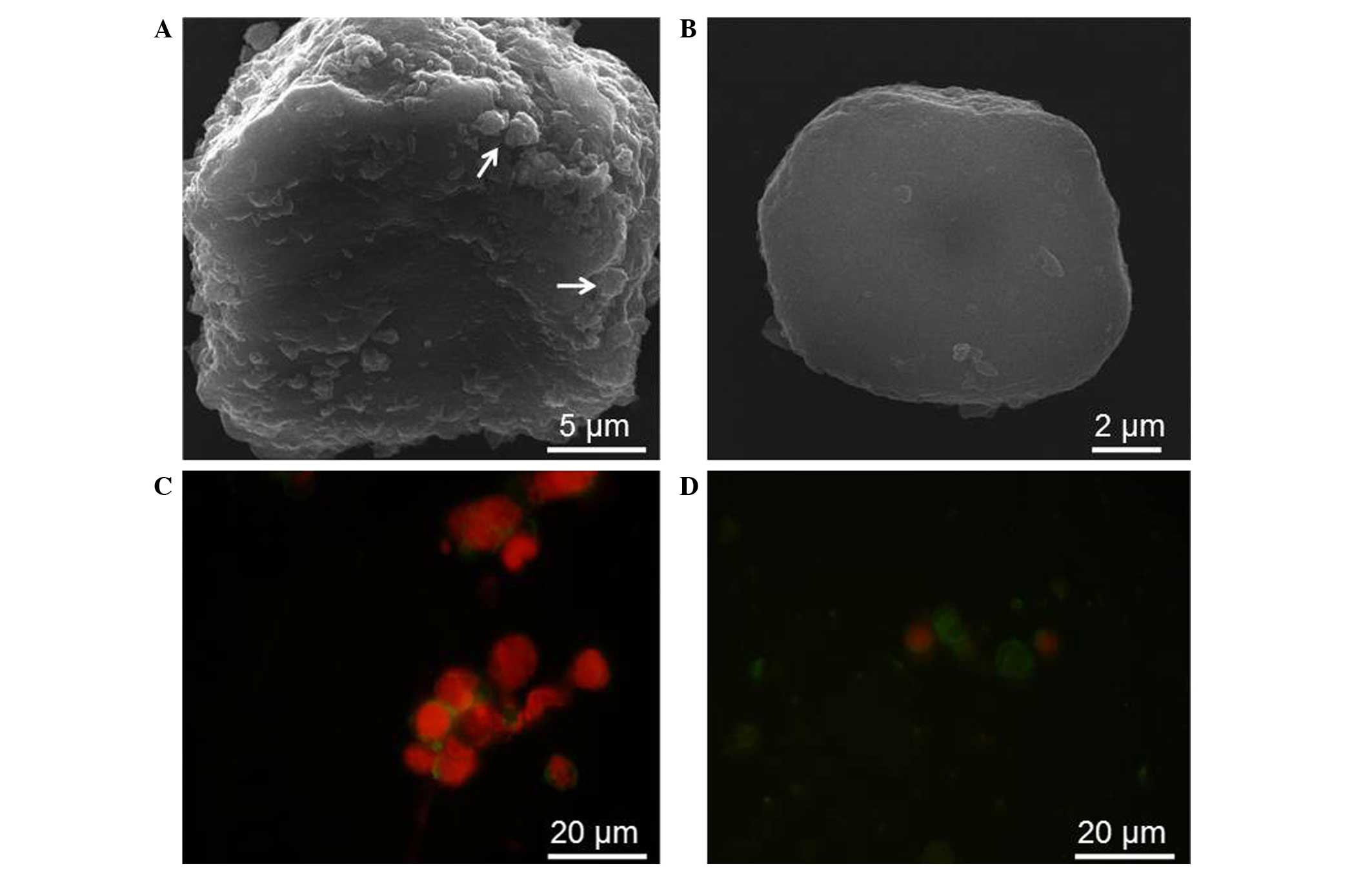
SEM and fluorescence microscopy were used to observe
the morphology of control cells and Jurkat cells undergoing
treatment with various concentrations of ex-LAC (Fig. 4). Whereas control cells possessed an
oval, regular shape (Fig. 4A), SEM
observation of Jurkat cells following 48 h of incubation with
ex-LAC revealed several characteristics of apoptosis, including
volume shrinkage and apoptotic body formation (Fig. 4B). Ex-LAC-treated and untreated Jurkat
cells were stained with Annexin V and PI prior to be observed under
fluorescence microscope (E-800; Nikon Corporation). Contrarily to
Jurkat control cells (Fig. 4C), cells
incubated with ex-LAC for 48 h displayed nucleus shrinkage and
fragmentation, as well as significant cytoplasm condensation
(Fig. 4D).
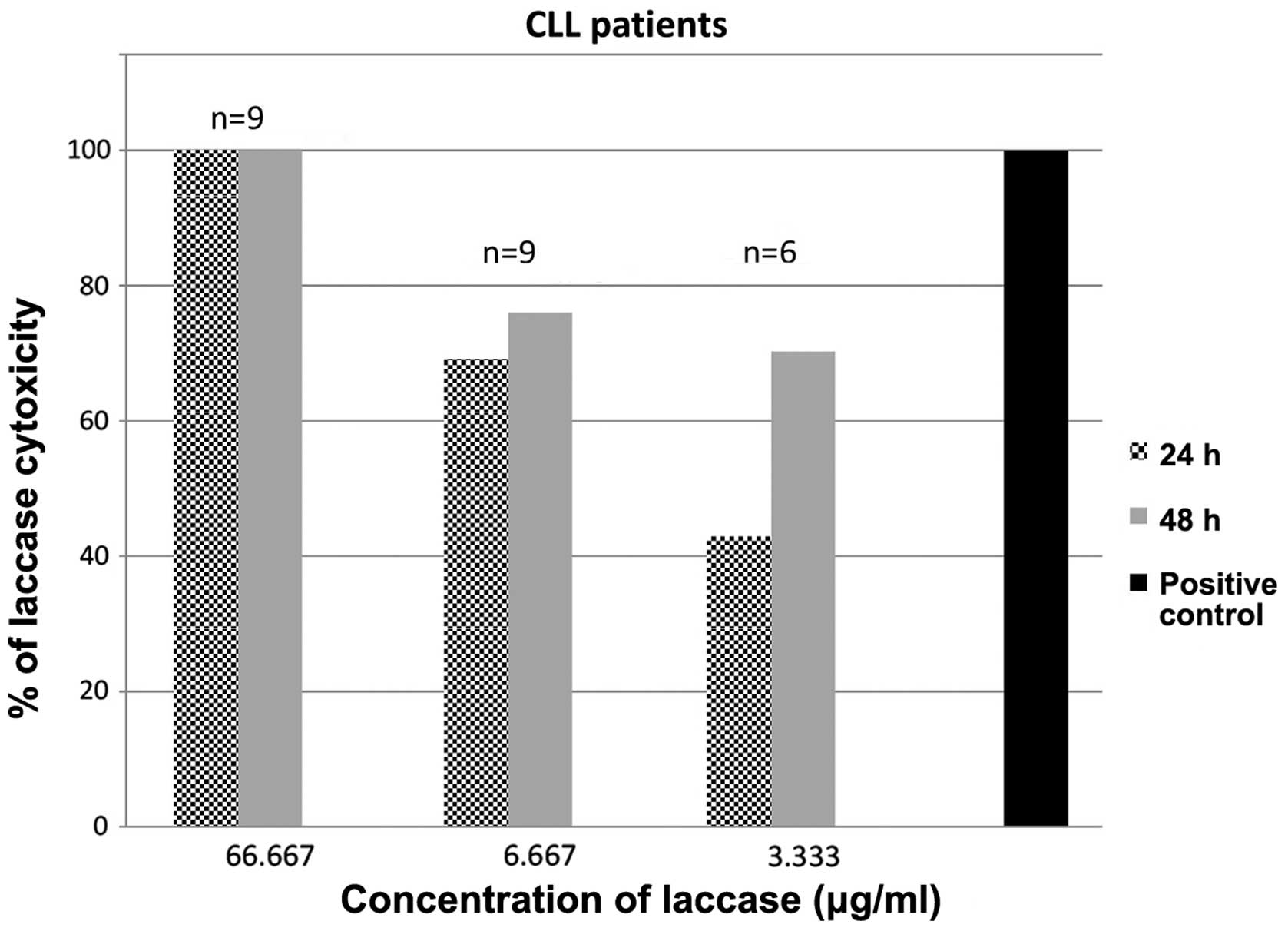
Cytotoxicity of ex-LAC is additionally
exerted against primary CLL cells
The cytotoxic activity of C. unicolor ex-LAC was
also assessed in PBMCs derived from nine patients with CLL via XTT
assay (Fig. 5). Based on the results
obtained in the established cell lines, three concentrations of
enzyme were selected for 24- and 48-h incubation with primary CLL
cells. Following 24 h of incubation with 66.667, 6.667 and 3.333
µg/ml ex-LAC, the median cytotoxic effect observed was 100.00,
69.06 and 42.85%, respectively. Following 48 h of incubation with
66.667, 6.667 and 3.333 µg/ml ex-LAC, a median cytotoxicity of
100.00, 75.99 and 70.26% was observed, respectively. The
IC50 values of ex-LAC on CLL cells were 0.7 and 0.9
µg/ml following 24 and 48 h of incubation, respectively.
Discussion
It has been reported that C. unicolor may be
a source of active ex-LAC (21).
Thus, we hypothesize that C. unicolor may be exploited to
produce high amounts of this biologically active substance with
pharmacological potential. The characterization of C.
unicolor ex-LAC isoforms has been previously described
(22). In the present study, the
enzyme isoforms Ia1, Ia2, Ib and IIa1 were recovered with a
65–92-fold increase in specific activity and a yield of 6.7, 27.5,
9.7 and 21.0%, respectively. The isoelectric points were in the
range of 4.7–4.2, and the carbohydrate content in the purified
enzymes was 1.6–3.5%. The specific activity of this isoform mixture
was 3,495.4 nkat/mg of protein. In comparison with a number of
alternative sources of ex-LAC, C. unicolor produces the
enzyme with high activity without requiring additional
supplementation such as aromatic compounds (21,23).
C. unicolor ex-LAC is currently utilized in
biodegradation, bioremediation, delignification and decolorization
processes (15). However, to the best
of our knowledge, there are no reports to date regarding its
anti-leukemic activity. In the present study, it was identified
that ex-LAC purified from C. unicolor possessed cytotoxic
activity against several hematological malignancies and primary CLL
cells using an XTT assay, and these findings were confirmed by
apoptosis analysis using flow cytometry, and additionally
visualized under SEM and fluorescence microscopy.
Anticancer properties have been reported for several
fungal extracts, including Funalia trogii (7,8). Unyayar
et al (8) measured the
inhibition of growth of the HeLa human cervical cancer cell line
and normal fibroblasts following 96 h of treatment with F.
trogii. The observed growth inhibition on HeLa cells was 27.2%
for 1 µl F. trogii extract, 39.7% for 3 µl, 56.1% following
treatment with 5 µl, 59.5% in the presence of 7 µl extract and
71.5% following incubation with 10 µl extract, as evaluated by
methyl thiazolyl tetrazolium assay (8). F. trogii extract contains LAC,
glutathione reductase and peroxidase, which have been documented to
be actively involved in cytotoxicity (8). Rashid et al (7) additionally confirmed the antitumor
activity of F. trogii extract against the HMEC-1 human
microvascular endothelial cell line using a trypan blue exclusion
assay (7). The highest toxicity
observed in the assay corresponded to 96-h treatment with 2.5 mg/ml
extract, while no toxicity was observed towards fibroblasts and
non-proliferating cells under those conditions. The results of the
XTT assay performed in the present study indicated that C.
unicolor ex-LAC induced cytotoxicity on HL-60, K562, RPMI 8226,
Jurkat and primary CLL cells in a dose-dependent manner. The
antitumor properties of LAC and peroxidase identified in F.
trogii are due to the presence of natural quinone substances
produced by the action of these enzymes on a lignin substrate
(8). Since all tumor cells are rich
in quinones and quinone-like molecules, extracts containing LACs
and/or peroxidases are able to selectively convert these molecules
into toxic substances that cause apoptosis of cells (8). The pro-oxidative and antibacterial
properties of ex-LAC from C. unicolor have been previously
described (24). In that study, the
potential of ex-LAC for production of reactive oxygen species was
investigated chemiluminometrically, and a marked pro-oxidative
action of the enzyme was identified (24). Estimation of ex-LAC toxicity using a
Microtox® detection system demonstrated that the exposure of the
marine bacterium Vibrio fischeri to ex-LAC caused 38 and 51%
cell damage following 5 and 15 min of incubation, respectively
(24). Ex-LAC has also been reported
to be effective against Escherichia coli (24). The apoptosis analysis of ex-LAC
extracted from C. unicolor conducted in the present study
additionally demonstrated apoptosis of Jurkat cells in the presence
of various concentrations of ex-LAC, compared with untreated
control cells. Apoptotic changes in Jurkat and RMPI 8226 cells
caused by C. unicolor ex-LAC were also observed under SEM
and fluorescence microscopy, thus confirming the above
findings.
Lau et al (25)
and Unyayar et al (8) have
independently documented the cytotoxic activity of Coriolus
versicolor extracts. Lau et al (25) observed significant dose-dependent
inhibitory effects on the proliferation of the Raji human Burkitt's
lymphoma B-cell line and the NB-4 and HL-60 human acute
promyelocytic leukemia cell lines treated with C. versicolor
extract. In that study, >90% inhibition was detected following
72 h of treatment. For the Raji lymphoma cell line, the
IC50 value of C. versicolor extract was 253.8
µg/ml, while for the NB-4 and HL-60 cell lines, the IC50
values were 269.3 and 147.3 µg/ml, respectively. The results of the
present study revealed that C. unicolor ex-LAC was able to
inhibit proliferation of human leukemic cell lines in a
dose-dependent manner at lower dosages than those documented by Lau
et al (25), since the
IC50 values following 48 h of treatment were 0.9, 0.4,
1.1 and 1.0 µg/ml for HL-60, Jurkat, RPMI 8226 and K562 cells,
respectively. Furthermore, the inhibitory effect of C.
versicolor extracts on the HeLa cell line demonstrated by
Unyayar et al (8) was lower
than the activity of C. unicolor ex-LAC identified in the
present study, since 1 µl C. versicolor extract caused 27.5%
growth inhibition in HeLa cells, and the maximum inhibition
observed was 45.5% following treatment with 10 µl extract (8). The results of the present study
indicated an inhibitory effect of C. unicolor ex-LAC of
~100.00% at a concentration of 666.667 µg/ml on all the cell lines
tested.
Anti-leukemic activity of agaritine, a β-glucan
isolated from Agaricus blazei, was reported by Endo et
al (26). In that study, the
2-(2-methoxy-4-nitrophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfophenyl)-2H-tetrazolium
assay was used to evaluate the inhibitory effect of agaritine from
A. blazei on the U937 human leukemic monocyte lymphoma,
MOLT-4 human acute lymphoblastic leukemia, HL-60 and K652 cell
lines following 48 h of treatment. The authors observed that the
viability of all the cell lines tested decreased with increasing
concentrations of agaritine. Agaritine suppressed cell growth in
U937, MOLT-4, HL-60 and K562 cells with an IC50 value of
2.7, 9.4, 13.0 and 16.0 µg/ml, respectively (26). By contrast, in the present study,
following 48 h of incubation of HL-60 and K562 cells with C.
unicolor ex-LAC, the IC50 values measured (0.9 and
1.0 µg/ml, respectively) were lower than those reported by Endo
et al (26), indicating
increased activity of C. unicolor ex-LAC compared with A.
blazei agaritine.
Chen et al (27) investigated the antitumor and
immunomodulatory effects of PCP-3A, a non-lectin glycoprotein
extracted from the mushroom Pleurotus citrinopileatus
(27). Trypan blue exclusion assay
revealed 37–64% growth inhibition of U937 cells following 72 h of
incubation with 25 µg/ml PCP-3A. In addition, P.
citrinopileatus PCP-3A was able to stimulate the secretion of
tumor necrosis factor α, interleukin-2 and interferon-γ by
CD4+ T cells, which indirectly suppressed the growth of
U937 cells, indicating that PCP-3A possessed antitumor and
immunomodulatory activities (27). By
contrast, C. unicolor ex-LAC inhibited the growth of K562,
Jurkat and RPMI 8226 cells by 100.00% at a concentration of 66.6667
µg/ml, while 87.00% cytotoxicity was observed in the HL-60 cell
line at a concentration of 66.667 µg/ml, as assessed by XTT assay
in the present study. Furthermore, 6.667 µg/ml C. unicolor
ex-LAC exhibited a higher cytotoxic rate against the tested cell
lines than 25 µg/ml P. citrinopileatus PCP-3A, with
cytotoxic rates of 67.51, 59.80, 100.00 and 66.96% for HL-60, K562,
Jurkat and RPMI 8226 cells, respectively. Additionally, C.
unicolor ex-LAC was demonstrated to induce apoptosis, which was
further confirmed in Jurkat and RPMI 8226 cell lines using flow
cytometry and microscopic techniques in the present study.
Tsai et al (28) reported a novel non-lectin glycoprotein
(HM-3A) purified from Hypsizygus marmoreus with
anti-leukemic activity against the U937 cell line (28). The antiproliferative effect of HM-3A
increased with concentration. As demonstrated by trypan blue
exclusion assay, HM-3A induced cytotoxicity against U937 cells at
concentrations ranged between 12.5 and 100.0 µg/ml, following
treatment for 24–72 h. At 100.0 µg/ml, HM-3A led to 96.2% growth
inhibition of U937 cells within 72 h (28). Similar cytotoxic rates were observed
in the current study, as 100.00% cytotoxicity was achieved at
concentrations of 666.667 and 66.667 µg/ml C. unicolor
ex-LAC in K562, Jurkat and RPMI 8226 cell lines. In HL-60 cells,
growth inhibition at 666.667 and 66.667 µg/ml C. unicolor
ex-LAC was 100.00 and 87.00%, respectively. Therefore, the results
of the present study indicated that C. unicolor ex-LAC
possessed similar anti-leukemic activity to H. marmoreus
HM-3A.
PNAP, a novel protein with antitumor activity
towards HeLa cells and the MCF7 breast cancer cell line, was
isolated from Pholiota nameko by Zhang et al
(29). The results of a trypan blue
exclusion assay conducted by these authors revealed dose-dependent
antiproliferative effects of PNAP following 24 h of treatment, with
an IC50 value of 9.97 µM for MCF7 cells and 12.11 µM for
HeLa cells. The authors observed that the cytotoxic effect of PNAP
on cancer cell lines was significantly increased, compared with the
effect on normal cells (29). In the
present study, the IC50 values observed following 24 h
of incubation with C. unicolor ex-LAC were 0.5, 0.8, 0.9,
0.8 and 0.7 µg/ml for HL-60, Jurkat, RPMI 8226, K562 and CLL
primary cells, respectively. Flow cytometric analysis of apoptosis
performed by Zhang et al (29)
following 48 h of incubation of MCF7 cells with 5, 10 and 15 µM
PNAP revealed the presence of 5.29, 10.05 and 22.88% apoptotic
cells, respectively, whilst the apoptotic rate in non-treated cells
was 3.23%. By contrast, in the present study, apoptotic rates
ranging from 88.98 to 98.90% were observed in Jurkat cells
following 48 h of ex-LAC treatment, depending on the concentration
of ex-LAC used. Furthermore, Zhang et al (29) measured the accumulation of
mitochondrial cytochrome c release into the cytosol, and
observed that the cytosol from untreated cells contained low
amounts of cytochrome c, in contrast to MCF7 cells incubated
with PNAP, where cytochrome c was significantly accumulated.
Similarly, reduced levels of cytochrome c in the
mitochondrial fraction were also detected. Release of cytochrome
c resulted in the activation of caspase-mediated apoptosis
(29). In the present study, SEM and
fluorescence microscopy images of ex-LAC-treated Jurkat and RPMI
8226 cells also indicated apoptosis involving cell volume and
nucleus shrinkage, apoptotic body formation and fragmentation, as
well as significant cytoplasm condensation.
To the best of our knowledge, a limited number of
LACs of fungal origin with anti-malignant properties have been
described thus far. Zhang et al (29) reported antiproliferative activity of
Clitocybe maxima LAC against HepG2 and MCF7 hepatocellular
carcinoma cells, and inhibitory activity towards human
immunodeficiency virus (HIV)-1 reverse transcriptase (30). At concentrations of 2.5, 5.0, 10.0 and
20.0 µM, purified LAC from C. maxima inhibited the
proliferation of HepG2 cells by 9.1, 20.4, 43.0 and 80.5%,
respectively, and inhibited cell growth in MCF7 cells by 40.2,
75.3, 90.2 and 95.4%, respectively. The IC50 values
against HepG2 and MCF7 cells were 12.3 and 3.0 µM, respectively.
The authors observed that C. maxima LAC reduced the activity
of HIV-1 reverse transcriptase with an IC50 value of
14.4 µM, and the percentage of inhibition of HIV-1 reverse
transcriptase activity at 5, 10 and 20 µM C. maxima LAC was
13.7, 35.1 and 70.4%, respectively. Zhang et al (31) purified LAC from the white-rot fungus
Abortiporus biennis and proved its antitumor activity
against HepG2 and MCF7 cells, as well as observing its inhibitory
activity against HIV-1 reverse transcriptase, achieving
IC50 values of 12.5, 6.7 and 9.2 µM, respectively
(31). Hu et al (32) demonstrated that LAC extracted from the
fruiting bodies of Agrocybe cylindracea species possessed
antiproliferative activity against MCF7 and HepG2 cell lines
(32). The authors noted that A.
cylindracea LAC additionally possessed HIV-1 reverse
transcriptase inhibitory activity, with percentages of inhibition
at 3.2, 8.0 and 20.0 µM of 15.7, 40.2 and 62.5%, respectively, and
a calculated IC50 value of 12.7 µM. The cytotoxicity of
A. cylindracea LAC towards HepG2 cells was 7.8, 30.2, 46.4
and 78.5% at concentrations of 1.2, 2.5, 5.0 and 10.0 µM,
respectively, with an IC50 value of 5.6 µM. In the case
of MCF7 cells, the IC50 value was 6.5 µM, and the
percentage of inhibition was 7.2, 22.7, 41.3 and 70.6% at
concentrations of 1.2, 2.5, 5.0 and 10.0 µM, respectively (32). In the present study, ex-LAC isolated
from C. unicolor induced a higher cytotoxic effect towards
HL-60, Jurkat, RPMI 8226 and K562 cells than A. cylindracea
LAC did in the above previous study, since C. unicolor LAC
demonstrated IC50 values of 0.5 (0.01 µM), 0.8 (0.014
µM) and 0.9 (0.016 µM) µg/ml for HL-60, Jurkat, RPMI 8226 and K562
cells, respectively.
To the best of our knowledge, the present study
represents the first report to investigate the anti-leukemic
activity of ex-LAC isolated and partially purified from idiophasic
cultures of C. unicolor. The present study provided novel
data concerning the isolation and chemical characterization of
bioactive compounds of the white-rot fungus C. unicolor. The
cytotoxic effect of ex-LAC extracted from this fungus was
demonstrated on HL-60, Jurkat, RPMI 8226 and K562 cell lines, as
well as CLL primary cells, using XTT assay. Additional analysis of
Jurkat and RPMI 8226 cells revealed that C. unicolor ex-LAC
was able to induce apoptosis of leukemic cells, even at low
concentrations. Compared with other compounds of fungal origin, the
IC50 values of C. unicolor ex-LAC were reduced,
indicating high anti-malignant activity of this enzyme. In
conclusion, the results of the present study suggest that C.
unicolor ex-LAC should be considered as a novel therapeutic
agent for the treatment of hematological malignancies.
Acknowledgements
The present study was supported by a grant from the
Medical University of Lublin (Lublin, Poland) (grant no. DS
462).
References
|
1
|
Hoehn D, Medeiros LJ and Konoplev S:
Molecular pathology of chronic lymphocytic leukemia.
Hematopathology: Genomic mechanisms of neoplastic diseases. Crisan
D: (New York). Humana Press. 255–291. 2010.
|
|
2
|
Xu Z, Zhang J, Wu S, Zheng Z, Chen Z and
Zhan R: Younger patients with chronic lymphocytic leukemia benefit
from rituximab treatment: A single center study in China. Oncol
Lett. 5:1266–1272. 2013.PubMed/NCBI
|
|
3
|
Ghia P, Ferreri AM and Caligaris-Cappio F:
Chronic lymphocytic leukemia. Crit Rev Oncol Hematol. 64:234–246.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Giannopoulos K: Biology and prognosis in
chronic lymphocytic leukemia. Acta Haematol Pol. 41:433–440.
2010.(In Polish).
|
|
5
|
Giannopoulos K, Dmoszynska A, Kowal M,
Wasik-Szczepanek E, Bojarska-Junak A, Rolinski J, Döhner H,
Stilgenbauer S and Bullinger L: Thalidomide exerts distinct
molecular antileukemic effects and combined thalidomide/fludarabine
therapy is clinically effective in high-risk chronic lymphocytic
leukemia. Leukemia. 23:1771–1778. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Patel S and Goyal A: Recent developments
in mushrooms as anti-cancer therapeutics: A review. 3 Biotech.
2:1–15. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Rashid S, Unyayar A, Mazmanci MA, McKeown
SR, Banat IM and Worthington J: A study of anti-cancer effects of
Funalia trogii in vitro and in vivo. Food Chem
Toxicol. 49:1477–1483. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Unyayar A, Demirbilek M, Turkoglu M, Celik
A, Mazmanci MA, Erkurt EA, Unyayar S, Cekic O and Atacag H:
Evaluation of cytotoxic and mutagenic effects of Coriolus
versicolor and Funalia trogii extracts on mammalian
cells. Drug Chem Toxicol. 29:69–83. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Leatham GF and Stahmann MA: Studies on the
laccase of Lentinus edodes: Specificity, localization and
association with the development of fruiting bodies. J Gen
Microbiol. 125:147–157. 1981.
|
|
10
|
Solomon EI, Sundaram UM and Machonkin TE:
Multicopper oxidases and oxygenases. Chem Rev. 96:2563–2606. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Baldrian P: Fungal laccases - occurrence
and properties. FEMS Microbiol Rev. 30:215–242. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Abadulla E, Tzanov T, Costa S, Robra KH,
Cavaco-Paulo A and Gübitz GM: Decolorization and detoxification of
textile dyes with a laccase from Trametes hirsuta. Appl
Environ Microbiol. 66:3357–3362. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Telke AA, Ghodake GS, Kalyani DC, Dhanve
RS and Govindwar SP: Biochemical characteristics of a textile dye
degrading extracellular laccase from a Bacillussp. ADR.
Bioresour Technol. 102:1752–1756. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Janusz G, Rogalski J and Szczodrak J:
Increased production of laccase by Cerrena unicolor in
submerged liquid cultures. World J Microbiol Biotechnol.
23:1459–1464. 2007. View Article : Google Scholar
|
|
15
|
Kucharzyk KH, Janusz G, Karczmarczyk I and
Rogalski J: Chemical modifications of laccase from white-rot
basidiomycete Cerrena unicolor. Appl Biochem Biotechnol.
168:1989–2003. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Rogalski J and Janusz G: Purification of
extracellular laccase from Cerrena unicolor. Prep Biochem
Biotechnol. 40:242–255. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Betina V: Differentiation and secondary
metabolism in some prokaryotes and fungi. Folia Microbiol (Praha).
40:51–67. 1995. View Article : Google Scholar
|
|
18
|
Leonowicz A and Grzywnowicz K:
Quantitative estimation of laccase forms in some white rot fungi
using syringaldazine as a substrate. Enzyme Microb Technol.
3:55–58. 1981. View Article : Google Scholar
|
|
19
|
Bradford MM: A rapid and sensitive method
for the quantitation of microgram quantities of protein utilizing
the principle of protein dye binding. Anal Biochem. 72:248–254.
1976. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jabbour E, O'Brien S, Konopleva M and
Kantarjian H: New insights into the pathophysiology and therapy of
adult acute lymphoblastic leukemia. Cancer. 121:2517–2528. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Leonowicz A, Gianfreda L, Rogalski J,
Jaszek M, Luterek J, Wasilewska MW, Malarczyk E, Dawidowicz A,
Fink-Boots M, Ginalska G, et al: Appearance of laccase in
wood-rotting fungi and its inducibility. J Korean Wood Sci Technol.
25:29–36. 1997.
|
|
22
|
Janusz G, Mazur A, Checinska A, Malek W,
Rogalski J and Ohga S: Cloning and characterization of a laccase
gene from biotechnologically important basidiomycete Cerrena
unicolor. J Fac Agr Kyushu U. 57:41–49. 2012.
|
|
23
|
Majeau JA, Brar SK and Tyagi RD: Laccases
for removal of recalcitrant and emerging pollutants. Bioresour
Technol. 101:2331–2350. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Jaszek M, Osińska-Jaroszuk M, Janusz G,
Matuszewska A, Stefaniuk D, Sulej J, Polak J, Ruminowicz M,
Grzywnowicz K and Jarosz-Wilkołazka A: New bioactive fungal
molecules with high antioxidant and antimicrobial capacity isolated
from Cerrena unicolor idiophasic cultures. BioMed Res Int.
2013:4974922013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lau CB, Ho CY, Kim CF, Leung KN, Fung KP,
Tse TF, Chan HH and Chow MS: Cytotoxic activities of Coriolus
versicolor (Yunzhi) extract on human leukemia and lymphoma
cells by induction of apoptosis. Life Sci. 75:797–808. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Endo M, Beppu H, Akiyama H, Wakamatsu K,
Ito S, Kawamoto Y, Shimpo K, Sumiya T, Koike T and Matsui T:
Agaritine purified from Agaricus blazei Murrill exerts
anti-tumor activity against leukemic cells. Biochim Biophys Acta.
1800:669–673. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chen JN, Ma CY, Tsai PF, Wang YT and Wu
JS: In vitro antitumor and immunomodulatory effects of the
protein PCP-3A from mushroom Pleurotus citrinopileatus. J
Agric Food Chem. 58:12117–12122. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tsai PF, Ma CY and Wu JS: A novel
glycoprotein from mushroom Hypsizygus marmoreus (Peck)
Bigelow with growth inhibitory effect against human leukemic U937
cells. Food Chem. 141:1252–1258. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang Y, Liu Z, Ng TB, Chen Z, Qiao W and
Liu F: Purification and characterization of a novel antitumor
protein with antioxidant and deoxyribonuclease activity from edible
mushroom Pholiota nameko. Biochimie. 99:28–37. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang GQ, Wang YF, Zhang XQ, Ng TB and
Wang HX: Purification and characterization of a novel laccase from
the edible mushroom Clitocybe maxima. Process Biochem.
45:627–633. 2010. View Article : Google Scholar
|
|
31
|
Zhang GQ, Tian T, Liu YP, Wang HX and Chen
QJ: A laccase with anti-proliferative activity against tumor cells
from a white root fungus Abortiporus biennis. Process
Biochem. 46:2336–2340. 2011. View Article : Google Scholar
|
|
32
|
Hu DD, Zhang RY, Zhang GQ, Wang HX and Ng
TB: A laccase with antiproliferative activity against tumor cells
from an edible mushroom, white common Agrocybe cylindracea.
Phytomedicine. 18:374–379. 2011. View Article : Google Scholar : PubMed/NCBI
|