Introduction
The cell nucleolus is a dynamic, non-membrane-bound
subnuclear structure, which has a classically well-established role
in ribosome biogenesis, including the synthesis and processing of
the ribosomal RNA (rRNA) precursor molecules (pre-rRNAs) and the
assembly of pre-rRNAs with particular ribosomal and nonribosomal
proteins to form preribosomal particles (1,2). Mammalian
nuclei typically contain one nucleolus, whereas tumor cells tend to
contain several nucleoli (3). The
typical tripartite organization of the nucleolus in mammals, which
accounts for ribosomal gene transcription and pre-rRNA processing
efficiency, is composed of three distinct components: The fibrillar
center (FC), the dense fibrillar component (DFC) and the granular
component (GC) (4–7). The early accumulation and processing of
rRNA takes place in the DFC, whereas late rRNA processing and
building of the ribosomal components occurs in the GC (2). As well as small nucleolar RNAs, a number
of proteins, including fibrillarin and nucleolar phosphoprotein B23
(B23), are located in the nucleolus (6). With regard to the unique nulceolar
localization in the cells at interphase, fibrillarin and B23 are
also widely used as markers for the DFC and GC, respectively
(8).
Previous studies have indicated that the nucleolus
is plurifunctional (9,10). In addition to the typically recognized
function of ribosome biosynthesis, the nucleolus supports the
production of additional gene expression components, including
transfer RNA and translational apparatus, the signal recognition
particle, ribonucleoprotein enzyme involved in maintenance of
telomerase integrity and U6 RNA, a catalytic RNA of the spliceosome
(9).
Small G protein ras homolog family member A (RhoA),
with a molecular weight of 21 kDa, is the most extensively studied
member of the Rho guanosine triphosphate (GTP)ase family and is
part of the Ras super family of small G proteins (11). RhoA has been reported to regulate
numerous biological activities, including gene transcription
(12) and tumor progression (13). Recent research on the intracellular
localization of RhoA has shown that it is located not only in the
cytosol and cell membrane, but also in the cell nucleus (14–16).
Notably, previous studies have demonstrated that the nuclear
localization of RhoA with predominant concentration in the cell
nucleolus is a common feature in human cancer tissues and cell
lines (17,18). Although our previous study has
demonstrated that the nuclear translocation of RhoA is via active
transport, a process involving importin α in a nuclear
factor-κB1-dependent manner, the mechanism, biological function and
pathological meaning of the nucleolar residency of RhoA remain to
be elucidated (18).
The antitumor antibiotic actinomycin D is a
polypeptide antibiotic that is widely used as inhibitor of RNA
synthesis. Actinomycin D is able to bind to duplex DNA and inhibit
the progression of DNA-dependent RNA polymerase (19,20).
Studies on the subcellular distribution of B23 have demonstrated
consistently that it is sensitive to actinomycin D treatment and
translocates from the cell nucleoli to alternative regions of the
nucleus (21–24). By contrast, the results of studies
regarding the subcellular distribution of fibrillarin with
actinomycin D treatment are diverse (25–27). A
previous study demonstrated that nucleolar fibrillarin also
redistributed to nucleoplasmic entities in response to actinomycin
D treatment in HEp-2 cells (28).
By utilizing an RNA synthesis inhibitor, the present
study aimed to investigate the association between RNA synthesis
and the nucleolar concentration of RhoA in human HEp-2 cells, as
well as the potential molecular mechanisms underlying the nucleolar
residency of RhoA.
Materials and methods
Cell lines and cell culture
Human carcinoma HEp-2 cells, which were originally
derived from an epidermoid carcinoma of the larynx, purchased from
the American Type Culture Collection (Manassas, VA, USA) were
maintained in complete RPMI-1640 that contained 10% fetal bovine
serum (FBS) and was supplemented with L-glutamine (2 mmol/l), salt
pyruvate (1 mmol/l), 1% non-essential amino acids and streptomycin
(10 mg/l), at 37°C in a humidified atmosphere of 5% CO2.
The cell culture medium was replaced every two days and the cells
were maintained at subconfluence.
Reagents
RPMI-1640 culture medium and FBS were obtained from
Gibco (Thermo Fisher Scientific, Inc., Waltham, MA, USA).
L-glutamine, salt pyruvate, non-essential amino acids,
streptomycin, phosphate-buffered saline (PBS) and nuclear
fluorochrome Hoechst 33342 were obtained from Sigma-Aldrich (St.
Louis, MO, USA). Primary antibodies against RhoA (monoclonal mouse;
#sc-418; dilution, 1:200) and fibrillarin (mouse monoclonal;
#sc-166000; dilution, 1:100) were obtained from Santa Cruz
Biotechnology, Inc. (Dallas, TX, USA), whereas monoclonal mouse
antibody against B23 (#B0556; dilution, 1:100) was purchased from
Sigma-Aldrich. Actinomycin D was additionally obtained from
Sigma-Aldrich. Mouse monoclonal antibody against
glyceraldehyde-3-phosphate dehydrogenase (GAPDH; #KC-5G4; dilution,
1:5,000) was obtained from KangCheng Bio-tech Inc. (Shanghai,
China). Goat anti-mouse immunoglobulin (Ig)G fluorescein
isothiocyanate-conjugated (FITC; #115-097-003; dilution, 1:1,000)
and goat anti-mouse IgG horseradish peroxidase (HRP)-conjugated
(#115-035-209; dilution, 1:5,000) secondary antibodies were
obtained from Jackson ImmunoResearch Laboratories, Inc. (West
Grove, PA, USA). Enhanced chemiluminescence (ECL) reagents were
purchased from GE Healthcare Life Sciences (Chalfont, UK).
Actinomycin D treatment
A stock solution of actinomycin D (2.5 g/l) was
prepared in ethanol (Changzhou Yabang Chemical Co.,Ltd., Changzhou,
China). It was diluted at least 2,500-fold and freshly prepared
prior to addition to the cells. For the reversibility assay, HEp-2
cells grown on cover slips in a 6-well plate (Genetimes Biological
Mall, Shanghai, China) were treated with actinomycin D. Following
treatment, cover slips were washed 3 times (1 min each wash) in
prewarmed PBS, followed by 1 brief wash with prewarmed fresh cell
culture medium without actinomycin D. Subsequent to treatment with
actinomycin D at the concentrations of 0.05 mg/l (for 4 h) and 1
mg/l (for 1 h), cells were cultivated in fresh culture medium
without actinomycin D for an additional incubation of 24 or 27 h,
which was dependent on the re-accumulation of RhoA in the cell
nucleolus.
Immunofluorescence microscopy
HEp-2 cells grown on cover slips were fixed with
freshly prepared paraformaldehyde (Nantong Jiangtian Chemical Co.,
Ltd., Nangtong, China) (40 g/l in PBS) for 30 min. Subsequent to
being penetrated with 30 ml/l Triton X-100 (Beijing Solarbio
Science & Technology Co., Ltd., Beijing, China) for 8 min and
blocked with 30 g/l bovine serum albumin (Genetimes Biological
Mall) for 1 h, the cells were incubated with the primary antibodies
against RhoA, fibrillarin or B23 at 4°C overnight, followed by
another incubation with the species-specific anti-IgG
FITC-conjugated secondary antibody for 1 h at room temperature.
Cells were washed three times with PBS (10 min each wash)
subsequent to each incubation. The distribution of the target
protein in the cells was analyzed by confocal laser scanning
microscopy (LSM 510; Carl Zeiss Microscopy GmbH, Jena, Germany).
For the counterstaining of cell nuclei with B23 or fibrillarin,
cells were incubated with nuclear fluorochrome Hoechst 33342 (10
mg/l) for 15 min at room temperature, followed by three washes in
PBS.
Western blotting assay
Cells were washed with cold PBS and transferred to a
1.7 ml centrifuge tube (Genetimes Biological Mall) where they were
centrifuged at 200 × g for 5 min at 4°C. Lysis buffer (50 mM Tris,
150 mM NaCl, 1 mM ethylenediaminetetraacetic acid and 1% Triton
X-100; pH, 7.4; Sinopharm Chemical Reagent Co., Ltd., Shanghai,
China) was added according to the proportion of 1 ml lysis buffer
in 107 cells. Following agitation for 15 min at 4°C,
centrifugation at 12,000 × g was performed for 15 min at 4°C. The
supernatant was collected as whole cell protein extract. The sample
proteins of whole HEp-2 cell lysate (20 µg protein) were run on
12.5% sodium dodecyl sulfate-polyacrylamide gels, followed by
transferal onto polyvinyl difluoride (PVDF) membranes (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). GAPDH was used as a loading
control. The PVDF membranes were initially blocked with 5% milk in
TBS-T (NaCl, 80 g/l; KCl, 2 g/l; Tris, 30 g/l; Tween-20, 0.1%; pH
7.4; Sinopharm Chemical Reagent Co., Ltd.) for 1 h at room
temperature and subsequently incubated with the primary antibodies
against RhoA and GAPDH at 4°C overnight. Following the incubation
of the membranes with the anti-IgG HRP-conjugated secondary
antibody for 1 h at room temperature, ECL reagents were used to
indicate the positive bands on the membrane, according to the
manufacturer's protocol. The bands were detected using the Typhoon
9400 (GE Healthcare Life Sciences).
Statistical analysis
A comparison of the amount of total RhoA protein
following treatment with varying concentrations of actinomycin D
was evaluated by one-way analysis of variance using SPSS 16.0
software (SPSS, Inc., Chicago, IL, USA). P<0.05 was considered
to indicate a statistically significant difference.
Results
The subcellular redistribution of
nucleolar RhoA following actinomycin D treatment in HEp-2
cells
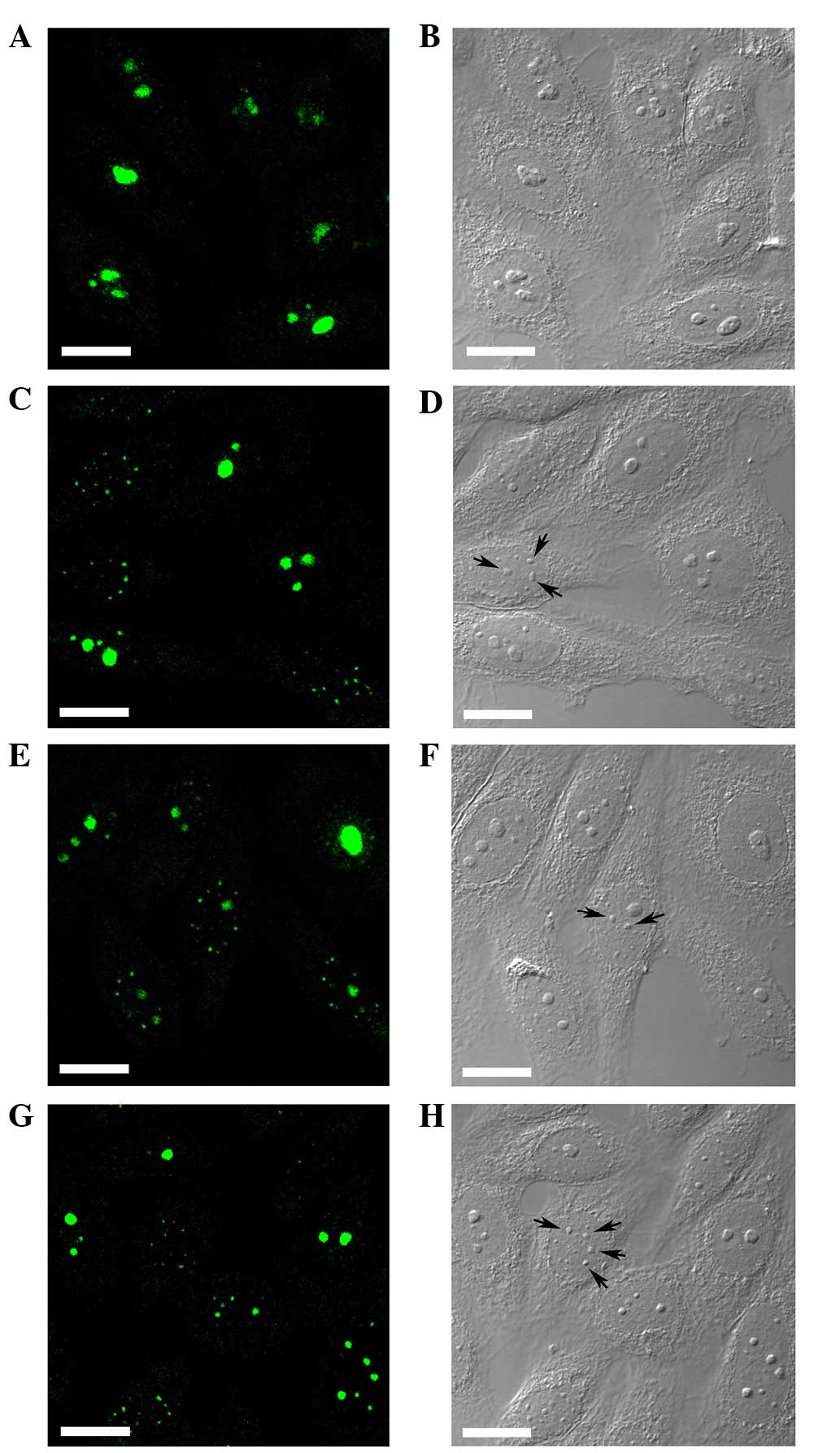
Confocal laser scanning microscopy revealed the
nuclear localization of RhoA with predominant accumulation in the
cell nucleolus in HEp-2 cells with no drug treatment (Fig. 1A), whereas treatment with actinomycin
D altered the subcellular distribution of RhoA within the cell
nucleus. Corresponding differential interference contrast pictures
revealed large nucleoli in untreated HEp-2 cells (Fig. 1B). Accompanied by a decrease in
nucleolar staining, a bright nucleoplasmic speckle staining of RhoA
was visualized in the cells (Fig. 1C, E
and G). However, the segregated nucleoli or nucleolus-like
bodies were observed in the cells treated with actinomycin D
(Fig. 1D, F and 1H; arrows).
Altered subcellular localization of
nucleolar protein B23 in HEp-2 cells following actinomycin D
treatment
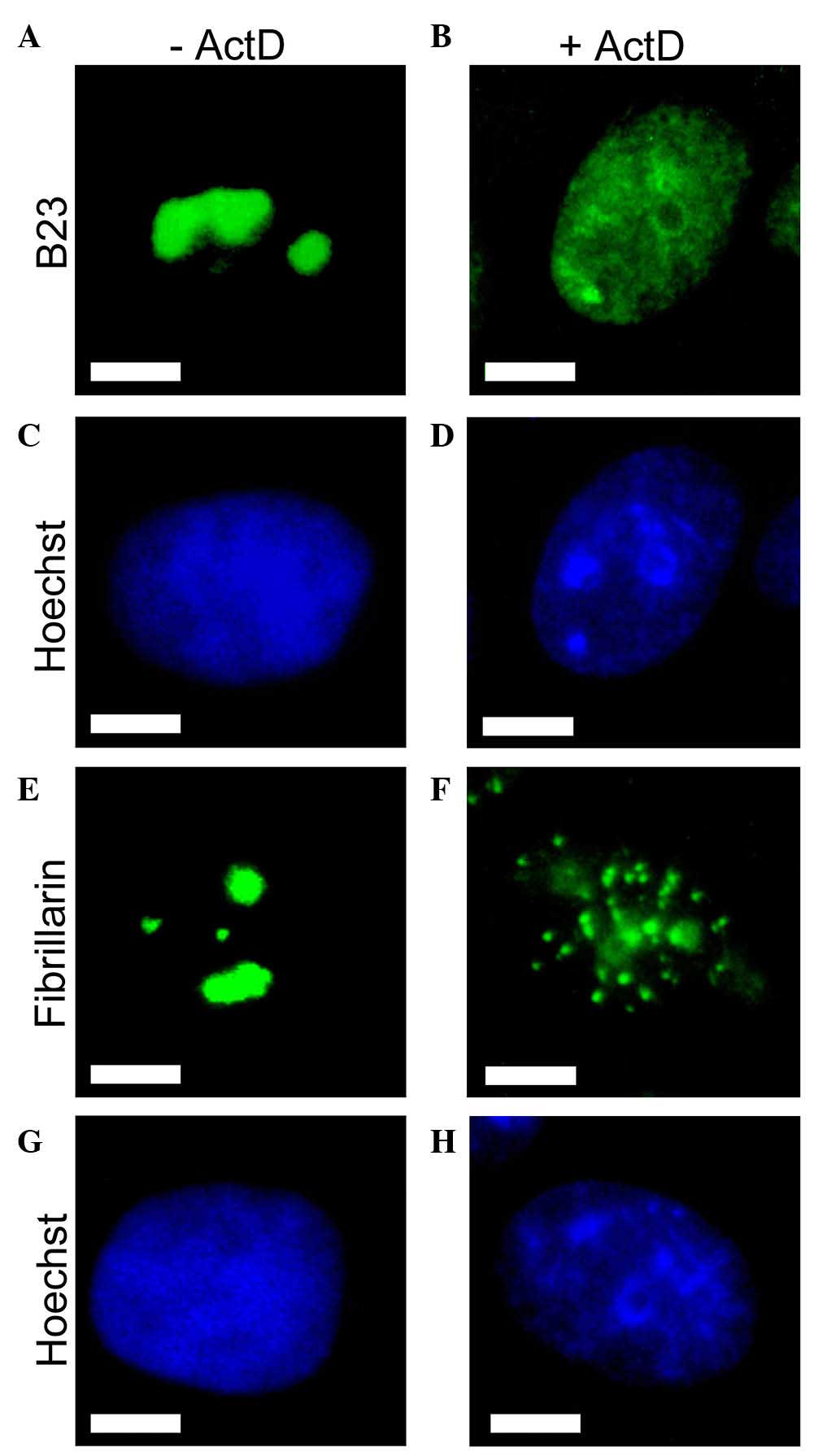
Intracellular localization of two abundant nucleolar
proteins fibrillarin and B23 was additionally examined by indirect
immunofluorescence (Fig. 2).
Nucleolar protein B23 is well-known to relocalize following
treatment with actinomycin D in numerous cell types, including
HEp-2 cells (21–28). The staining for B23 was concentrated
in the nucleoli of untreated HEp-2 cells (Fig. 2A), but was redistributed diffusely
throughout the nucleus following 4 h of treatment with 0.05 mg/l
actinomycin D (Fig. 2B). This
redistribution of B23 may act as a marker for effective treatment
by actinomycin D. Another abundant nucleolar protein, fibrillarin,
demonstrated exclusively nucleolar localization in untreated cells
(Fig. 2E). Consistent with the
previous finding, a redistribution of fibrillarin to the
nucleoplasmic small entities was induced by actinomycin D (Fig. 2F).
RhoA protein content is unchanged
following actinomycin D treatment
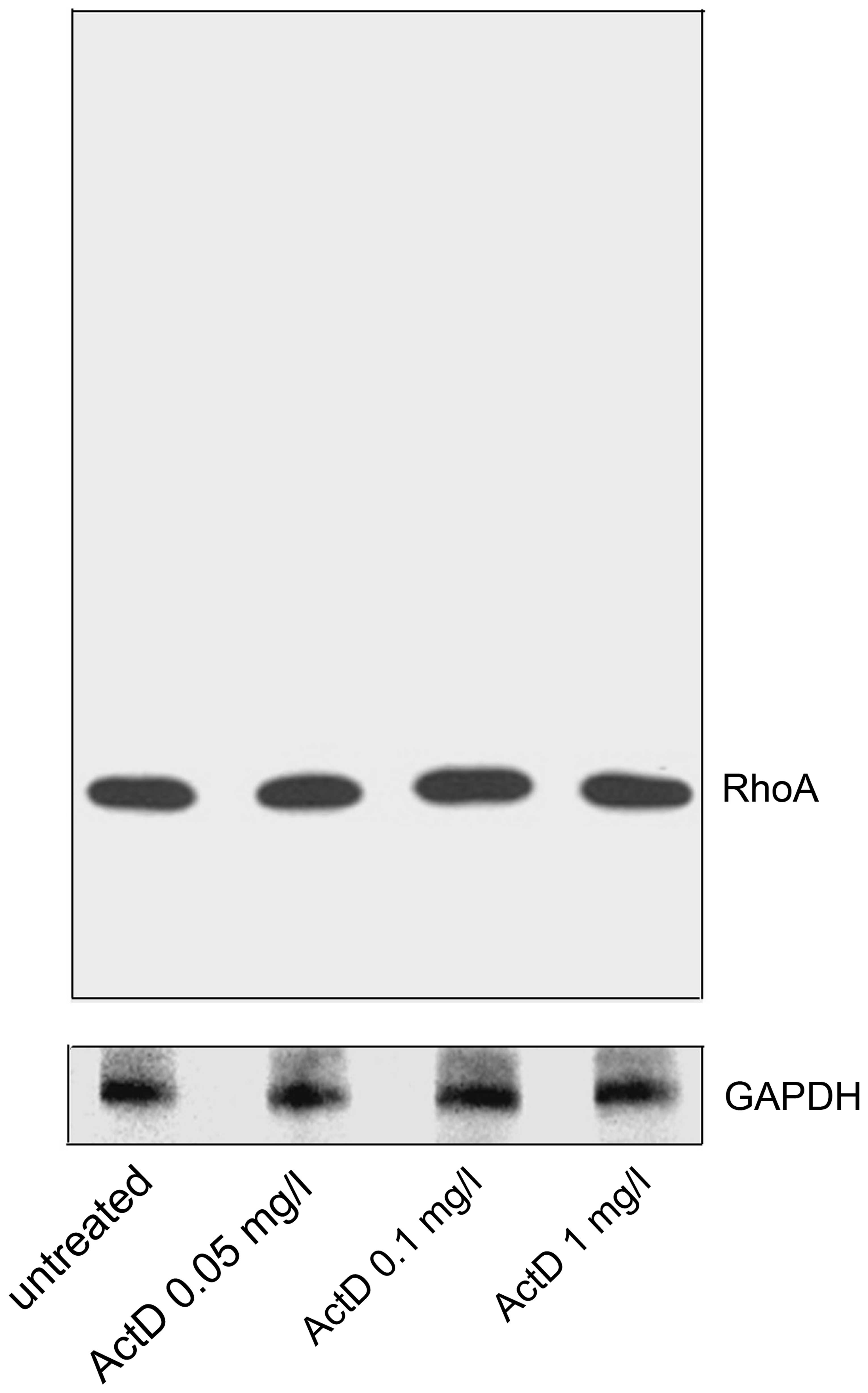
Following treatment with actinomycin D for 4 h,
HEp-2 cells were washed with PBS and lysed under denaturing
conditions. The protein amounts from whole-cell lysate were
compared through immunoblotting analysis. Regardless of the various
concentrations of actinomycin D, immunoblotting revealed a single
band exclusively at 21 kDa for all untreated and actinomycin
D-treated groups (Fig. 3).
Statistical analysis on the amount of total RhoA protein following
various treatments with actinomycin D demonstrated no change in
total cellular RhoA content (P>0.05). Therefore, the actinomycin
D treatment did not affect the net protein amount of RhoA, whilst
it did affect the cellular distribution of the RhoA protein.
Nuclear redistribution of RhoA is
reversible
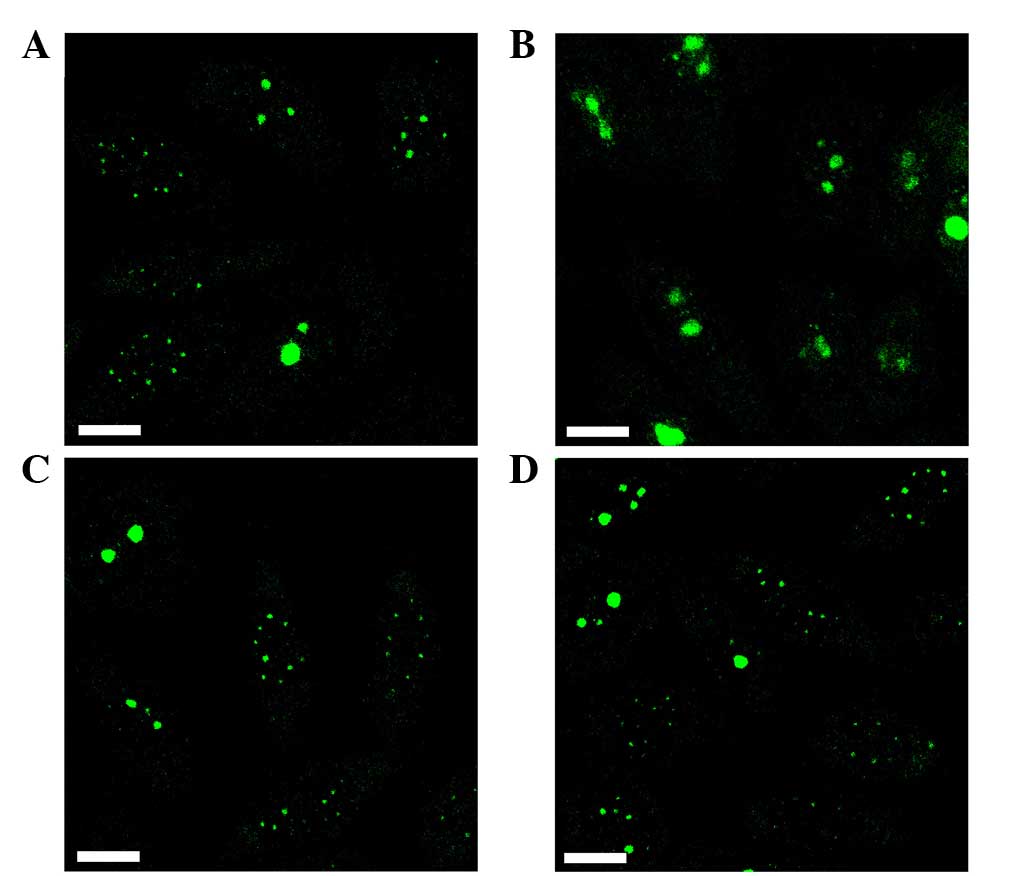
To investigate the persistence and reversibility of
the actinomycin D-induced nuclear redistribution of RhoA, HEp-2
cells were initially treated with actinomycin D, followed by an
additional cultivation in fresh culture medium without actinomycin
D for at least 24 h. When HEp-2 cells were incubated with 0.05 mg/l
actinomycin D for 4 h, a redistribution of RhoA was observed
(Fig. 4A). Removal of the drug from
the culture medium allowed the reaccumulation of RhoA in the cell
nucleoli 24 h later (Fig. 4B).
However, reaccumulation was not observed following initial
treatment with an increased concentration of actinomycin D (1 mg/l
for 1 h) (Fig. 4C), even when the
incubation time without actinomycin D was extended to 27 h
(Fig. 4D).
Discussion
Over the past few decades, the understanding of
nucleolar function has changed markedly with the concept of the
plurifunctional nucleolus (9).
Proteomic studies have suggested that the cell nucleolus may be
involved in a wide range of cellular processes independent of
ribosome biogenesis (29). Extensive
evidence indicates that the nucleolus affects the response to
cellular stress, and regulates the cell cycle and cell growth
(29). Perturbations to the nucleolus
have been reported in a wide range of cellular diseases, from
autoimmunity to cancer (30).
An important aspect regarding nucleolus structure
and function are the morphological variations that exist between
normal somatic and neoplastic or malignant cells (31). For over a century, an increase in the
size and number of cell nucleoli has been utilized as a marker of
aggressive tumors (31,32). Contemporary studies have inferred that
nucleoli may have a broad role in malignant transformation.
Specifically, the extra-ribosomal functions of the nucleolus
position the organelle as a central integrator of cellular
proliferation and stress signaling and are emerging as important
mechanisms for modulating how oncogenes and tumor suppressors
operate in normal and malignant cells (33).
Research has demonstrated that RhoA serves a key
role in governing extra- and intracellular signaling transduction,
in addition to being involved in numerous biological processes,
including tumorigenesis (34). RhoA,
as the most extensively investigated member of the Rho protein
family, acts as a molecular switch in cells, regulating signal
transduction from cell surface receptors to intracellular target
molecules, and is involved in a number of biological processes,
including cell morphology, motility and tumor progression (34). Without destroying the cells, the
present study demonstrated a redistribution of nucleolar RhoA
between the nucleoli and the nucleoplasm with a speckled staining
pattern following the treatment of HEp-2 cells with actinomycin D.
Furthermore, immunoblotting revealed a single band exclusively at
21 kDa, indicating that an actinomycin D-induced decrease in
nucleolar RhoA was not accompanied by any alteration in RhoA
protein content, biochemical modification or cleavage of RhoA.
These results are concordant with previous findings regarding the
nucelolar protein fibrillarin following treatment with actinomycin
D. The present results demonstrating that the nucleolar protein B23
translocated to the nucleoplasm, along with redistribution of
nucleolar fibrillarin to the nucleoplasm following treatment with
actinomycin D, are also consistent with previous observations made
in various types of cells (24,28). As
actinomycin D may displace RNA polymerases by binding DNA and
subsequently inhibiting RNA synthesis (35), the present results may reflect the
redistribution of RhoA between the nucleoli and nucleoplasm when
RNA synthesis is inhibited.
At low concentrations (0.04–0.05 mg/l), actinomycin
D selectively inhibits transcription that is mediated by RNA
polymerase I in the nucleolus, but not transcription mediated by
RNA polymerase II and III (36). The
reaccumulation of RhoA occurred between the nucleoplasm and the
nucleoli following removal of the lower concentration (0.05 mg/l)
of actinomycin D, which extended the aforementioned results due to
the observation that the effect on the subcellular redistribution
of RhoA induced by low concentrations of actinomycin D was
reversible. However, due to the action of actinomycin D on RNA
polymerases II and III at increased concentrations, even subsequent
to an extended period of drug removal, the displacement of RhoA was
irreversible upon treatment of cells with increased concentrations
of actinomycin D, which led to severe inhibition of RNA synthesis.
The DFC and the edge of the FC are the nucleolar regions where
transcription takes place (7).
Although the exact location of RhoA in respect to the nucleolar
component remains to be elucidated, considering the similar
nucleolar localization of RhoA and transcription sites, the
observation of actinomycin D-induced redistribution of nucleolar
RhoA to the nucleoplasm revealed direct evidence that the residency
of RhoA in the nucleolus was dependent on active transcription.
In line with the present results, the βII-subunit of
tubulin, a major subunit of microtubules that was previously
considered to exist solely in the cytoplasm of cells, has also been
identified to localize to the nuclei, with a strong aggregation in
the nucleoli of numerous types of carcinoma cells (37,38).
Following treatment with the drug taxol, which inhibits mitosis
targeted at the microtubules of dividing cells, βII-tubulin
underwent a rearrangement in the nuclei and demonstrated the
formation of micronuclei (37).
Increasing evidence emerged that these cellular proteins may be
dynamic, crossing through the cytoplasm, nucleoplasm and nucleolus
to perform their roles, and a number of these proteins may remain
to be elucidated (16,18,38).
In the current era of translational medicine, the
newly recognized roles of the nucleolus raise the possibility that
cancer drug discovery and emerging chemotherapy may identify a
foundation in the nucleolus or nucleolar proteins to a greater
degree than had been anticipated (35). Knowledge of the location, function and
duration of the residency of proteins in the nucleolus is vital in
identifying additional drug targets in the nucleolus.
In conclusion, the results of the present study
provided in situ evidence that actinomycin D induced a
redistribution of nucleolar RhoA to the nucleoplasm in human
carcinoma HEp-2 cells. In addition, the results indicated that the
residency of RhoA in the cell nucleolus is associated with active
RNA synthesis. To the best of our knowledge, these results provide
the first information regarding nucleolar RhoA in association with
nucleolar function. Additional investigations on the nuclear or
nucleolar functioning of RhoA are underway and may provide novel
considerations for cancer therapy.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (Beijing, China; grant no.
81372404), the Specialized Research Fund for Senior Personnel
Program of Jiangsu University (Zhenjiang, China; grant no.
11JDG129) and the Postdoctoral Foundations of China (Beijing,
China; grant no. 2012M521018) and Jiangsu province (Nanjing, China;
grant no. 1201025B) granted to Dr Yueying Li.
References
|
1
|
Misteli T: Protein dynamics: Implications
for nuclear architecture and gene expression. Science. 291:843–847.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Scheer U, Thiry M and Goessens G:
Structure, function and assembly of the nucleolus. Trends Cell
Biol. 3:236–241. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bártová E, Horáková AH, Uhlírová R, Raska
I, Galiová G, Orlova D and Kozubek S: Structure and epigenetics of
nucleoli in comparison with non-nucleolar compartments. J Histochem
Cytochem. 58:391–403. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hernandez-Verdun D: The nucleolus today. J
Cell Sci. 99:465–471. 1991.PubMed/NCBI
|
|
5
|
Spector DL: Macromolecular domains within
the cell nucleus. Annu Rev Biochem. 9:265–315. 1993. View Article : Google Scholar
|
|
6
|
Shaw PJ and Jordan EG: The nucleolus. Annu
Rev Cell Dev Biol. 11:93–121. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Scheer U and Hock R: Structure and
function of the nucleolus. Curr Opin Cell Biol. 11:385–390. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Muro E, Gébrane-Younís J, Jobart-Malfait
A, Louvet E, Roussel P and Hernandez-Verdun D: The traffic of
proteins between nucleolar organizer regions and prenucleolar
bodies governs the assembly of the nucleolus at exit of mitosis.
Nucleus. 1:201–211. 2010. View Article : Google Scholar
|
|
9
|
Pederson T: The plurifunctional nucleolus.
Nuclei Acid Res. 26:3871–3876. 1998. View Article : Google Scholar
|
|
10
|
Pederson T: The nucleolus. Cold Spring
Harb Perspect Biol. 3:a0006382011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mackay DJ and Hall A: Rho GTPases. J Biol
Chem. 273:20685–20688. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sotiropoulos A, Gineitis D, Copeland J and
Treisman R: Signal-regulated activation of serum response factor is
mediated by changes in actin dynamics. Cell. 98:159–169. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kamai T, Yamanishi T, Shirataki H, Takagi
K, Asami H, Ito Y and Yoshida K: Overexpression of RhoA, Rac1, and
Cdc42 GTPases is associated with progression in testicular cancer.
Clin Cancer Res. 10:4799–4805. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dubash AD, Guilluy C, Srougi MC, Boulter
E, Burridge K and García-Mata R: The small GTPase RhoA localizes to
the nucleus and is activated by Net1 and DNA damage signals. PloS
One. 6:e173802011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Guilluy C, Dubash AD and García-Mata R:
Analysis of RhoA and Rho GEF activity in whole-cells and the cell
nucleus. Nat Protoc. 6:2050–2060. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li YY, Chen YC and Xu J: Factors
influencing RhoA protein distribution in the nucleus. Mol Med
Report. 4:1115–1119. 2011.
|
|
17
|
Li YY, Chen YC, Tao Y, Xu J and Chen M:
RhoA protein is generally distributed in the nuclei of cancer
cells. Oncol Rep. 24:1005–1009. 2010.PubMed/NCBI
|
|
18
|
Xu J, Li Y, Yang X, Chen Y and Chen M:
Nuclear translocation of Small G protein RhoA via active
transportation in gastric cancer cells. Oncol Rep. 30:1878–1882.
2013.PubMed/NCBI
|
|
19
|
Perry RP and Kelly DE: Inhibition of RNA
sythesis by actinomycin D: Characteristic dose-response of
different RNA species. J Cell Physiol. 76:127–139. 1970. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen FM: Binding specificities of
actinomycin D to self-complementary tetranucleotide sequences
-XGCY-. Biochemistry. 27:6393–6397. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Biggiogera M, Fakan S, Kaufmann SH, Black
A, Shaper JH and Busch H: Simultaneous immunoelectron microscopic
visualization of protein B23 and C23 distribution in the HeLa cell
nucleolus. J Histochem Cytochem. 37:1371–1374. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sweet P, Chan PK and Slater LM:
Cyclosporin A and verapamil enhancement of daunorubicin-produced
nucleolar protein B23 translocation in daunorubicin-resistant and
-sensitive human and murine tumor cells. Cancer Res. 49:677–680.
1989.PubMed/NCBI
|
|
23
|
Yung BY, Bor AM and Chan PK: Short
exposure to actinomycin D induced ‘reversible’ translocation of
protein B23 as well as ‘reversible’ inhibition of cell growth and
RNA synthesis in HeLa cells. Cancer Res. 50:5987–5991.
1990.PubMed/NCBI
|
|
24
|
Yao Z, Duan S, Hou D, Wang W, Wang G, Liu
Y, Wen L and Wu M: B23 acts as a nucleolar stress sensor and
promotes cell survival through its dynamic interaction with hnRNPU
and hnRNPA1. Oncogene. 29:1821–1834. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Benavente R, Reimer G, Rose KM, Hügle-Dörr
B and Scheer U: Nucleolar changes after microinjection of
antibodies to RNA polymerase I into the nucleus of mammalian cells.
Chromosoma. 97:115–123. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Puvion-Dutilleul F, Mazan S, Nicoloso M,
Pichard E, Bachellerie JP and Puvion E: Alteration of nucleolar
ultrastructure and ribosome biogenesis by actinomycin D.
Implications for U3 snRNP function. Eur J Cell Biol. 58:149–162.
1992.PubMed/NCBI
|
|
27
|
Rivera-León R and Gerbi SA: Delocalization
of some small nucleolar RNPs after actinomycin D treatment to
deplete early pre-rRNAs. Chromosoma. 105:506–514. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chen M and Jiang P: Altered subcellular
distribution of nucleolar protein fibrillarin by actinomycin D in
HEp-2 cells. Acta Pharmacol Sin. 25:902–906. 2004.PubMed/NCBI
|
|
29
|
Boisvert FM, van Koningsbruggen S,
Navascués J and Lamond AI: The multifunctional nucleolus. Nat Rev
Mol Cell Biol. 8:574–585. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Montanaro L, Treré D and Derenzini M:
Nucleolus, ribosomes, and cancer. Am J Pathol. 173:301–310. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hein N, Hannan KM, George AJ, Sanij E and
Hannan RD: The nucleolus: An emerging target for cancer therapy.
Trends Mol Med. 19:643–654. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Pianese G: Beitrag zur Histologie und
Aetiologie der Carcinoma. Histologische und experimentelle
Untersuchungen. Beitr Pathol Anat Allg Pathol. 142:1–193. 1896.
|
|
33
|
Quin JE, Devlin JR, Cameron D, Hannan KM,
Pearson RB and Hannan RD: Targeting the nucleolus for cancer
intervention. Biochim Biophys Acta. 1842:802–816. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Orgaz JL, Herraiz C and Sanz-Moreno V: Rho
GTPases modulate malignant transformation of tumor cells. Small
GTPases. 5:e290192014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Pickard AJ and Bierbach U: The cell's
nucleolus: An emerging target for chemotherapeutic intervention.
Chem Med Chem. 8:1441–1449. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ochs RL: Methods used to study structure
and function of the nucleolus. Methods Cell Biol. 53:303–321. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Xu K and Ludueña RF: Characterization of
nuclear betaII-tubulin in tumor cells: A possible novel target for
taxol. Cell Motil Cytoskeleton. 53:39–52. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Howard J and Hyman AA: Dynamics and
mechanics of the microtubule plus end. Nature. 422:753–758. 2003.
View Article : Google Scholar : PubMed/NCBI
|