Introduction
Cervical cancer is the fourth most common cause of
cancer and the fourth most common cause of cancer-associated
mortality in women, worldwide (1). In
2012, there was ~528,000 novel cases of cervical cancer diagnosed,
and 266,000 mortalities (1). In
total, ~70% of cervical cancers are identified in developing
countries (1). Lung cancer is the
most common cause of cancer-associated mortality in men and women
worldwide, accounting for 1.56 million mortalities in 2012
(2). Primary liver cancer is the
sixth most common type of cancer and the second most common cause
of cancer-associated mortality, worldwide (2); in 2012, there were 782,000 novel cases
diagnosed and 746,000 mortalities (2). Hepatocellular carcinoma is the most
common type of liver cancer, accounting for ~80% of all types of
primary liver cancer (3). Although
conventional cancer therapies, such as surgery, chemotherapy and
radiotherapy, are used for the treatment of the majority of solid
tumors, successful therapeutic outcomes are often limited, due to
side effects associated with treatment, drug toxicity and the
development of multidrug resistance (4,5).
Therefore, novel therapeutic strategies with improved treatment
efficacies and decreased toxicity with no adverse effects are
urgently required.
Recently, the use of oncolytic viruses in the
treatment of cancer has progressed significantly (6–8). Oncolytic
viruses selectively kill tumor cells by replicating in tumor cells
exclusively. In 1999, the Sendai virus strain Tianjin was isolated
from the lungs of a marmoset (9).
Tianjin is a novel genotype of the Sendai virus, which has been
demonstrated to possess oncolytic activities in various types of
tumor cell (10–13). In a previous study by the present
authors, ultraviolet-inactivated Sendai virus strain Tianjin
(UV-Tianjin) particles were prepared, and it was demonstrated that
they inhibited the growth of murine colon carcinoma cells via the
induction of the immune response and apoptosis in mice (14). However, the underlying molecular
mechanisms by which Sendai virus or UV-Tianjin induced anticancer
effects remain unclear.
Clinical and experimental studies have demonstrated
that anticancer effects are mediated via various mechanisms, such
as alteration of carcinogen metabolism, induction of DNA repair
systems, activation of immune responses, suppression of cell cycle
progression and promotion of apoptosis (15–17).
Apoptosis may be induced by various chemotherapeutic agents, which
are important for the prevention and treatment of cancer (18–20). The
aim of the present study was to investigate the anticancer and
proapoptotic effects of UV-Tianjin against the human cervical
cancer HeLa, human small cell lung cancer NCI-H446 and human
hepatocellular carcinoma Hep 3B cell lines in vitro, and to
analyze the possible mechanisms underlying apoptosis induced by
UV-Tianjin. The present study also aimed to provide novel insights
concerning the antitumor mechanisms of the Sendai virus.
Materials and methods
Reagents
RPMI-1640 medium, Dulbecco's modified Eagle's medium
(DMEM) and fetal bovine serum (FBS) were obtained from Thermo
Fisher Scientific, Inc. (Waltham, MA, USA).
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT)
was purchased from Sigma-Aldrich (St. Louis, MO, USA). Hoechst
33342 and caspase-3, −8 and −9 Colorimetric Assays were purchased
from KeyGen Biotech Co., Ltd. (Nanjing, China). Annexin
V-fluorescein isothiocyanate (FITC)/propidium iodide (PI) Apoptosis
kit was obtained from Biovision, Inc. (Milpitas, CA, USA). JC-10
Mitochondrial Membrane Potential (MMP) Assay kit was purchased from
Fanbo Science & Technology Co., Ltd. (Beijing, China).
Monoclonal rabbit anti-caspase-3 (dilution, 1:500; catalog no.,
9665), monoclonal mouse anti-caspase-8 (dilution, 1:500; catalog
no., 9746), polyclonal rabbit anti-caspase-9 (dilution, 1:500;
catalog no., 9502) and monoclonal rabbit anti-apoptosis protease
activating factor-1 (Apaf-1; dilution, 1:500; catalog no., 8723)
antibodies were obtained from Cell Signaling Technology, Inc.
(Danvers, MA, USA). Polyclonal rabbit anti-cytochrome c (cyt
c; dilution, 1:500; catalog no., sc-7159), polyclonal rabbit
anti-Fas (dilution, 1:200; catalog no., sc-715), polyclonal rabbit
anti-Fas ligand (FasL; dilution, 1:500; catalog no., sc-6237),
polyclonal rabbit anti-Fas-associated protein with death domain
(FADD; dilution, 1:500; catalog no., sc-5559), monoclonal mouse
anti-β-actin (dilution, 1:1,000; catalog no., sc-47778), goat
anti-mouse immunoglobin (Ig)G-horseradish peroxidase (HRP;
dilution, 1:2,000; catalog no., sc-2005) and goat anti-rabbit
IgG-HRP (dilution, 1:5,000; catalog no., sc-2004) antibodies were
purchased from Santa Cruz Biotechnology, Inc. (Dallas, TX,
USA).
Cell and virus cultivation
HeLa cells were obtained from the Cell Bank of Type
Culture Collection of Chinese Academy of Sciences (Shanghai, China)
and cultured in RPMI 1640 medium supplemented with 10% FBS at 37°C
in a humidified atmosphere of 5% CO2. NCI-H446 and Hep
3B cells were kindly provided by the General Hospital of Tianjin
Medical University (Tianjin, China) and cultured in DMEM containing
10% FBS at 37°C in a humidified atmosphere of 5% CO2.
Sendai virus strain Tianjin particles (GenBank: EF679198) were
cultured in chorioallantoic fluid obtained from 9–11 day-old
chicken eggs. After incubation for 72 h, the allantoic fluid was
carefully collected and then centrifuged (5810R; Eppendorf,
Hamburg, Germany) at 1,000 × g for 10 min at 4°C to pellet debris.
The virus-containing supernatant was purified by discontinuous
sucrose gradient centrifugation and inactivated by UV irradiation
(99 mJ/cm2), as previously described (21). Inactivated viruses cannot replicate,
but retain cell fusion activity.
Cell viability assay
A total of 5×103 cells/well (HeLa, NCI-H446 and Hep
3B) were seeded onto 96-well plates for 24 h. After incubation with
various doses of UV-Tianjin [multiplicity of infection (MOI), 0,
10, 20, 40, 80, 200 and 400] for 24 or 48 h, MTT solution (5 mg/ml)
was added to each well and incubated for 4 h at 37°C. The medium
containing MTT was aspirated and treated with 100 µl dimethyl
sulfoxide. Absorbance at a wavelength of 570 nm was measured using
a microplate reader.
Hoechst staining
HeLa, NCI-H446 or Hep 3B cells (2×105 cells per
well) were seeded onto 6-well plates. Following treatment with
UV-Tianjin (MOI, 40 or 80) for 24 h, the cells were washed twice
with phosphate-buffered saline (PBS) and incubated with 250 µl
Hoechst 33342 for 20 min in the dark. After rinsing with PBS, the
cells were examined using a fluorescence microscope (E600; Nikon
Corporation, Tokyo, Japan). Cells with nuclei that exhibited
characteristic bright blue fluorescence, due to condensed or
fragmented chromatin, were considered to be apoptotic.
Annexin V-FITC/PI staining
The externalization of phosphatidylserine was
determined by flow cytometric analysis of cell staining using an
Annexin V-FITC conjugate and PI according to the manufacturer's
protocol. The cells (2×105 cells/well) were seeded in 6-well
plates, cultured for 24 h and subsequently treated with UV-Tianjin
(MOI, 20, 40 or 80) for an additional 24 h. The cells were then
trypsinized (Thermo Fisher Scientific, Inc.), washed twice with
PBS, centrifuged at 400 × g for 5 min to discard the supernatant,
and resuspended in 500 µl binding buffer (FITC/PI Apoptosis kit;
Biovision, Inc.). Subsequently, Annexin V-FITC (5 µl) and PI (5 µl)
were added to the suspension and incubated for 10 min at room
temperature in the dark. The cells were evaluated by flow cytometry
(FACSVerse; BD Biosciences, San Jose, CA, USA). Data were analyzed
using CellQuest™software version 5.1 (BD Biosciences) and the
results were expressed as percentages of the cell populations.
Caspase activity assay
Caspase activity was measured using caspases-3, −8
and −9 colorimetric activity assay kits in HeLa cells. Following
incubation with UV-Tianjin (MOI, 20, 40 or 80) for 24 h, HeLa cells
were harvested, lysed with cell extraction buffer (KeyGen Biotech
Co., Ltd.) and phenylmethylsulfonyl fluoride (KeyGen Biotech Co.,
Ltd.), incubated on ice for 30 min and vortexed for 30 sec. The
cell lysates were centrifuged at 12,000 × g for 10 min at 4°C and
the caspase-3, −8 or −9 levels in the supernatant were measured
using the caspase-specific colorimetric kits, according to the
manufacturer's protocol. The optical density was measured using a
microplate reader (BioTek Instruments, Inc.) at a wavelength of 405
nm.
Measurement of MMP
The fluorescent cationic dye, JC-10, was used to
determine the MMP in HeLa cells. JC-10 is capable of selectively
accessing the mitochondrial membrane, and exhibits a reversible
color change from green to orange as the membrane potential
increases. When MMP is relatively low, JC-10 dye forms monomers and
emits green fluorescence (wavelength, 525–530 nm). By contrast,
when MMP is high, JC-10 aggregates and emits orange fluorescence
(wavelength, 590 nm) (22). The ratio
between green and orange fluorescence provides an estimate of MMP
that is independent of the mitochondrial mass. Briefly, after 24 h
of treatment with UV-Tianjin (MOI, 20, 40 or 80), HeLa cells were
harvested, washed and incubated with JC-10 (5 µM) for 30 min at
37°C in the dark. Subsequently, the cells were analyzed by flow
cytometry, which was performed using the same method as described
for Annexin V-FITC/PI staining.
Western blot analysis
HeLa cells (2×105 cells/well) were seeded onto
6-well plates. After 24 h, the cells were treated with UV-Tianjin
(MOI, 20, 40 or 80) and incubated for an additional 24 h.
Subsequently, the cells were washed in PBS, scraped from the well
and centrifuged at 400 × g for 5 min to discard the supernatant.
Cell lysates were obtained by homogenizing the cell pellets in
lysis buffer (Beijing Solarbio Science & Technology Co., Ltd.,
Beijing, China). Protein concentrations were determined using a
bicinchoninic acid protein assay kit (Beijing Solarbio Science
& Technology Co., Ltd.), with bovine serum albumin (Beijing
Solarbio Science & Technology Co., Ltd.) as the standard.
Protein (40 µg) was separated using 10% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis gel and transferred onto
polyvinylidene difluoride membranes. The membranes were blocked
with 5% milk for 1 h at room temperature and were then washed 3
times (5 min each wash) in TBST [Tris-buffered saline with
Tween-20: 20 mM Tris-HCl (pH 7.6), 150 mM NaCl and 0.05% Tween-20].
The membranes were then incubated with primary antibodies against
cyt c, Apaf-1, caspase-3, −8 and −9, pro-caspase-3, −8 and
−9, Fas, FasL, FADD, β-actin at 4°C overnight and washed 3 times
for 5 min each in TBST. The membranes were then incubated with
anti-mouse IgG HRP-linked antibody or anti-rabbit IgG HRP-linked
antibody for 1 h at room temperature and washed 3 times for 5 min
each in TBST, and the protein levels were measured using a
chemiluminescence kit (Beijing Solarbio Science & Technology
Co., Ltd.). Protein signals were analyzed by ImageJ version 1.43
software (National Institutes of Health, Bethesda, MA, USA).
β-actin served as the internal control.
Statistical analysis
Each experiment was replicated at least three times.
Data are expressed as the mean ± standard deviation. Analysis of
variance was used to analyze differences between the groups. SPSS
software version 18.0 (SPSS, Chicago, IL, USA) was used to perform
all statistical analyses, and P<0.05 was considered to indicate
a statistically significant difference.
Results
UV-Tianjin inhibits the proliferation
of HeLa, NCI-H446 and Hep 3B cells
The cell lines HeLa, NCI-H446 and Hep 3B were
cultured with UV-Tianjin (MOI, 10, 20, 40, 80, 200 and 400) for 24
h or 48 h, and cell proliferation was assessed by MTT assay. As
shown in Fig. 1A, UV-Tianjin
inhibited the proliferation of all three cell lines in a dose- and
time-dependent manner. The results also demonstrated that the HeLa
and NCI-H446 cell lines were more sensitive to UV-Tianjin treatment
compared with the Hep 3B cell line. Notably, the HeLa and NCI-H446
cell lined exhibited almost identical proliferation
characteristics. Microscopic examination confirmed the results
obtained from the MTT assay (Fig.
1B).
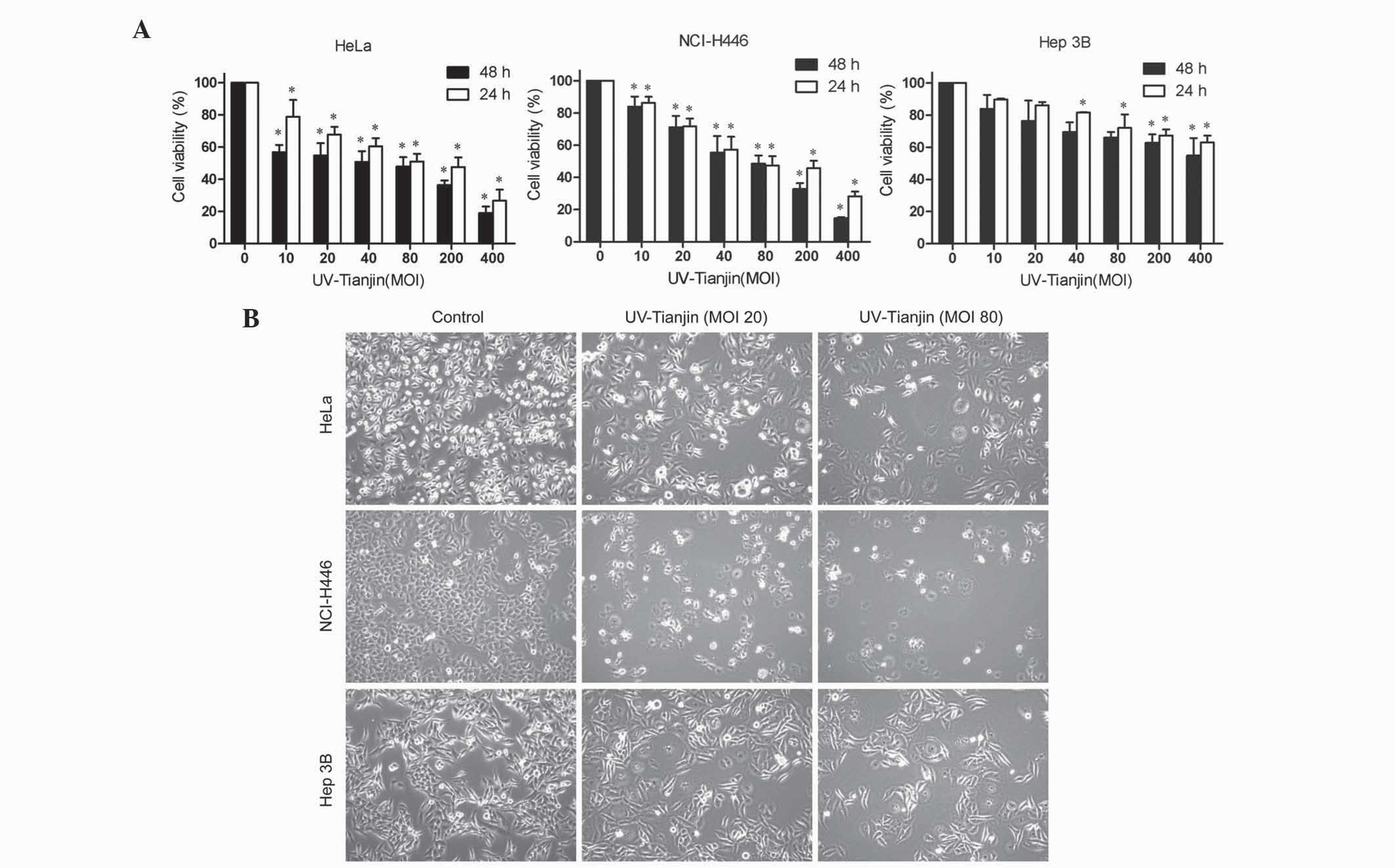 | Figure 1.UV-Tianjin treatment exhibits
antiproliferative effects in human cervical carcinoma HeLa, human
small cell lung cancer NCI-H446 and human hepatocellular carcinoma
Hep 3B cells. (A) HeLa, NCI-H446 and Hep 3B cells were treated with
various doses of UV-Tianjin (MOI, 0, 10, 20, 40, 80, 200 and 400)
for 24 h or 48 h and cell viability was determined by
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay.
Data are presented as the mean ± standard deviation of three
experiments performed in quadruplicate. *P<0.05 vs. untreated
control cells. (B) Representative photographs of HeLa, NCI-H446,
and Hep 3B cells under a fluorescence microscope after UV-Tianjin
(MOI, 40 and 80) treatment for 24 h (magnification, ×100).
UV-Tianjin, ultraviolet-inactivated Sendai virus strain Tianjin;
MOI, multiplicity of infection. |
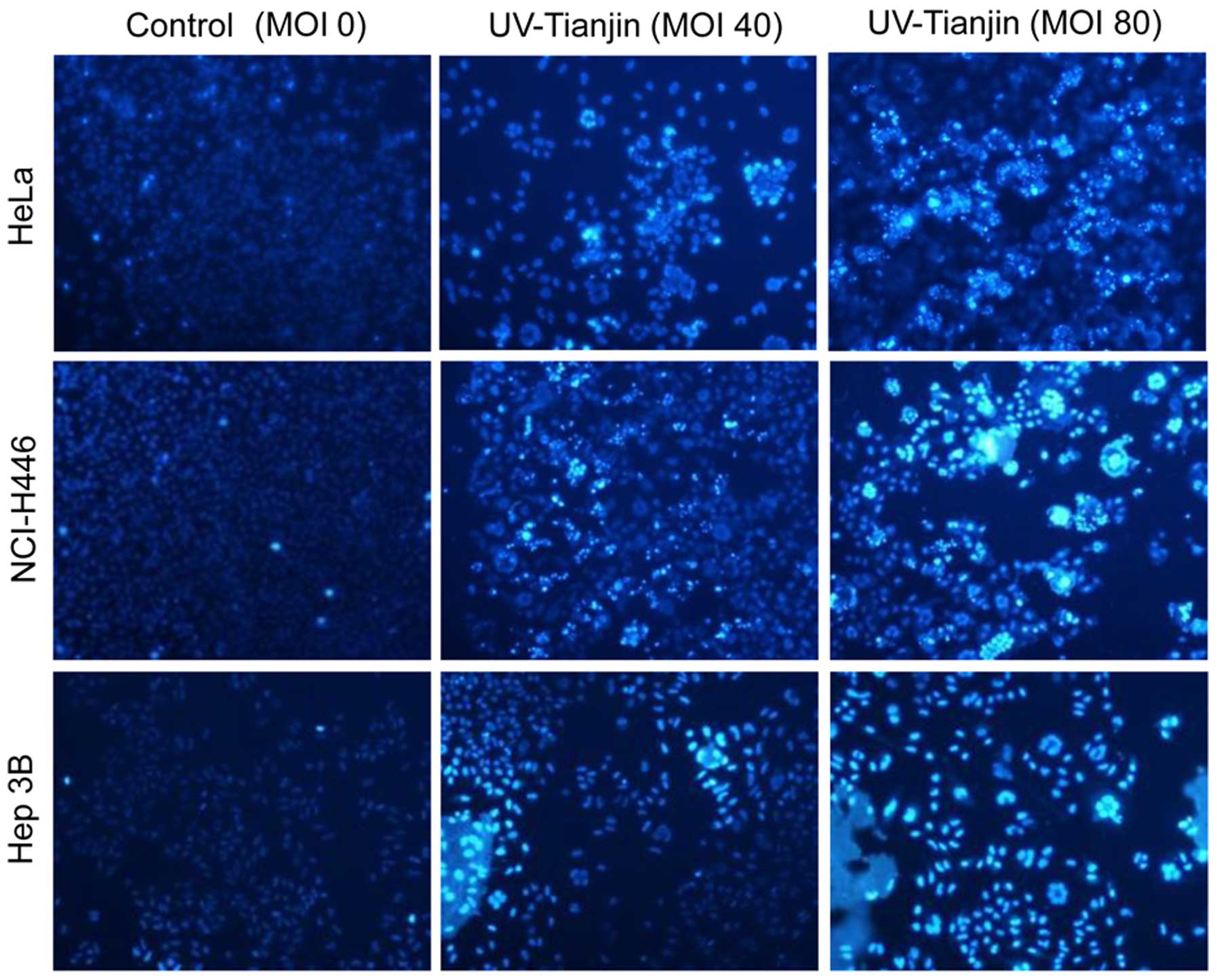
UV-Tianjin induces apoptosis of HeLa,
NCI-H446 and Hep 3B cells
To examine whether the anti-proliferative effects of
UV-Tianjin are due to the induction of apoptosis, the cell lines
HeLa, NCI-H446 and Hep 3B were stained with Hoechst 33342 dye and
observed under a fluorescence microscope. Compared with the
untreated groups, a higher percentage of apoptotic cells were
observed in the UV-Tianjin-treated groups (Fig. 2). Furthermore, a dose-dependent
increase in apoptosis was observed in all three cell lines.
Subsequently, the number of apoptotic cells was
estimated using flow cytometry. As shown in Fig. 3A and B, for HeLa cells, the early
apoptotic cells accounted for 36.01±4.44, 45.98±3.26 and
64.73±4.82% subsequent to a 24 h incubation with UV-Tianjin at MOI
20, 40 and 80, respectively, compared with 21.88±4.36, 26.57±2.84
and 33.04±2.24% in NCI-H446 cells and 10.22±1.77, 15.31±2.97 and
27.07±2.46% in Hep 3B cells. A dose-dependent increase in early
apoptotic rate was observed in all three cell lines (P<0.01;
Fig. 3B). In addition, the results
demonstrated that the HeLa cell line was most susceptible to
UV-Tianjin treatment. Consequently, the HeLa cell line was selected
for further experiments.
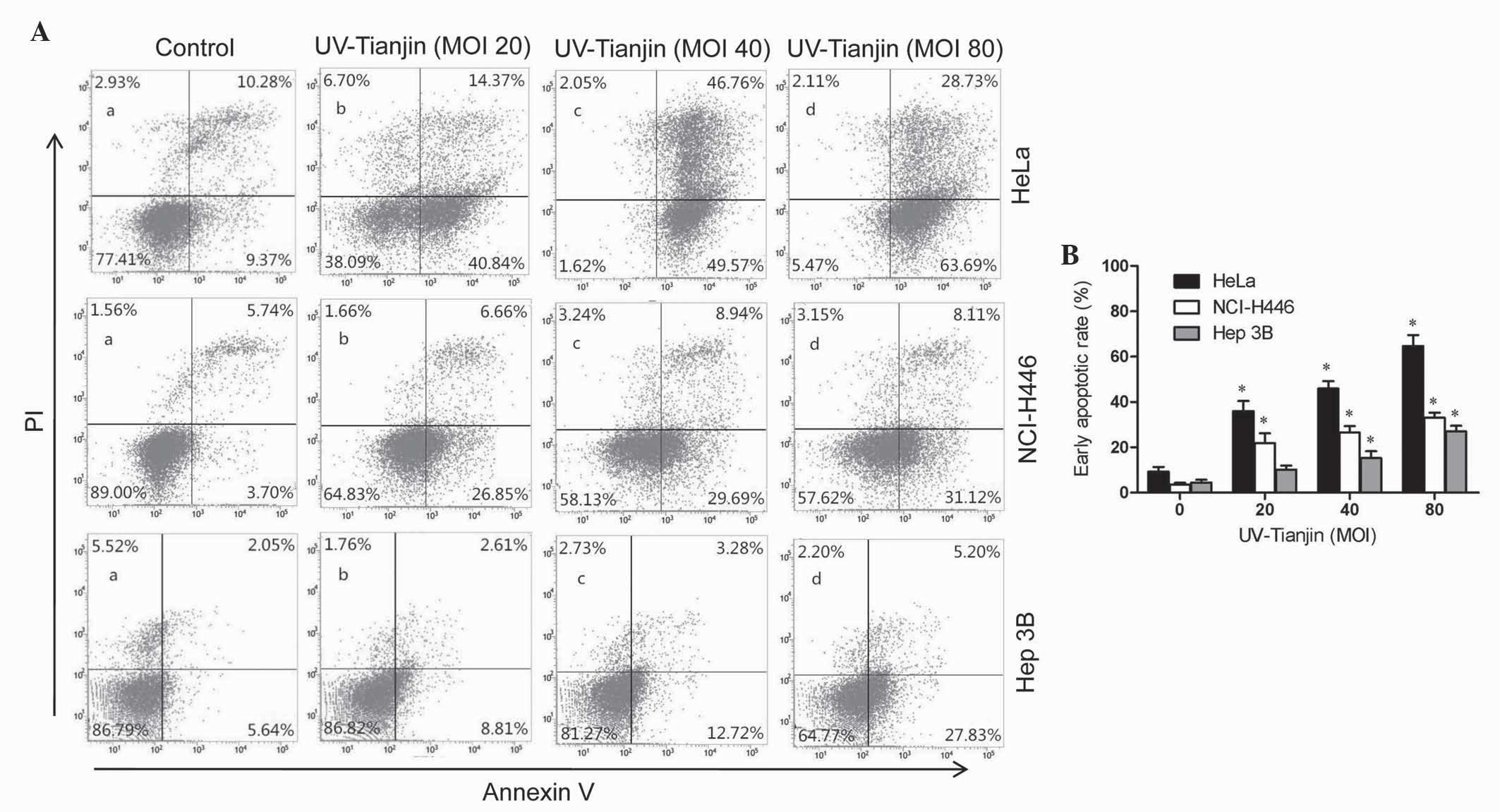 | Figure 3.Annexin V-FITC/PI staining of human
cervical carcinoma HeLa, human small cell lung cancer NCI-H446 and
human hepatocellular carcinoma Hep 3B cells following treatment
with UV-Tianjin (MOI, 0, 20,40 and 80). (A) Following treatment
with UV-Tianjin (MOI, 0, 20, 40 and 80) for 24 h, HeLa, NCI-H446
and Hep 3B cells were stained with Annexin V-FITC/PI and analyzed
by flow cytometry. The lower right quadrant (Annexin
V+/PI−) represents early apoptosis, whereas
the upper right quadrant (Annexin V+/PI+)
represents late apoptosis and necrosis. (B) Early apoptotic rate
(%) of HeLa, NCI-H446 and Hep 3B cells as determined by flow
cytometry. Data are expressed as the mean ± standard deviation of
three independent experiments. *P<0.01 vs. untreated control
cells. FITC, fluorescein isothiocyanate; PI, propidium iodide;
UV-Tianjin, ultraviolet-inactivated Sendai virus strain Tianjin;
MOI, multiplicity of infection. |
UV-Tianjin disrupts MMP in HeLa
cells
Early apoptosis is usually accompanied by collapse
of the mitochondrial membrane, which results in MMP disruption. In
the present study, the effect of UV-Tianjin on MMP was analyzed
using a mitochondria-specific dye, JC-10. As shown in Fig. 4A and B, flow cytometry demonstrated
that the intensity of orange fluorescence was significantly
decreased following UV-Tianjin treatment for 24 h, indicating MMP
disruption. A total of 48.77±4.12, 54.61±2.53 and 63.79±2.20% of
cells exhibited MMP disruption following treatment with UV-Tianjin
at MOI, 20, 40 and 80, respectively, which suggests that UV-Tianjin
treatment decreased the MMP of HeLa cells in a dose-dependent
manner.
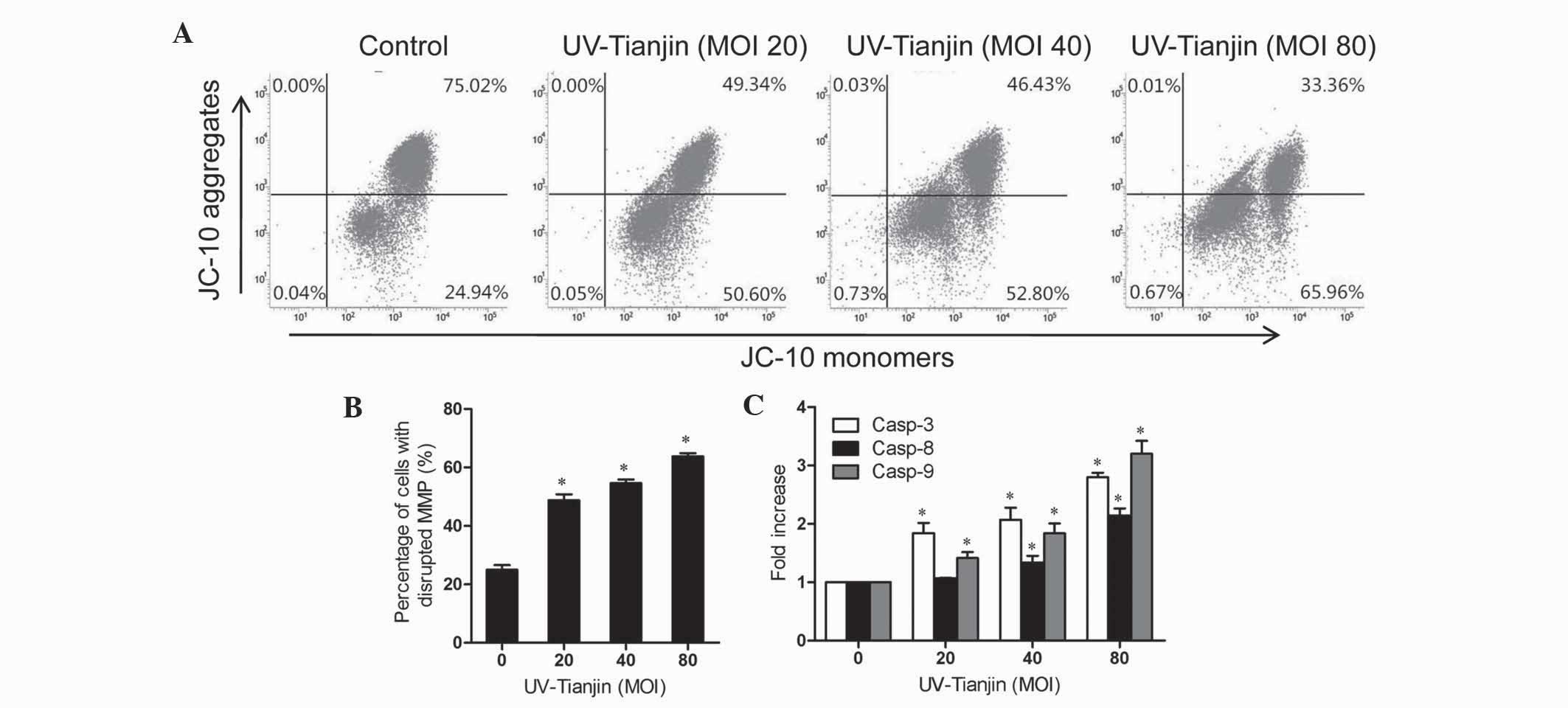 | Figure 4.Effects of UV-Tianjin on MMP and casp
activity in human cervical carcinoma HeLa cells. (A) MMP was
decreased in UV-Tianjin-treated HeLa cells. Cells were treated with
various doses of UV-Tianjin (MOI, 0, 20, 40 and 80) for 24 h and
stained with JC-10. Fluorescence was measured by flow cytometry.
(B) Histograms show the percentage of cells with a depolarized
mitochondrial membrane. (C) HeLa cells were treated with UV-Tianjin
(MOI, 20, 40, 80) for 24 h and analyzed by casp-3, −8, and −9
activity assays. Results are expressed as the fold increase in casp
activity of apoptotic cells compared to that of non-treated cells.
Data are expressed as the mean ± standard deviation of three
experiments. *P<0.05 vs. untreated control cells. UV-Tianjin,
ultraviolet-inactivated Sendai virus strain Tianjin; MOI,
multiplicity of infection; MMP, mitochondrial membrane potential;
casp, caspase. |
UV-Tianjin-induced apoptosis is
caspase-dependent in HeLa cells
To further investigate the proapoptotic mechanism of
UV-Tianjin, caspase-3, −8, and −9 activity in HeLa cells was
investigated following incubation with UV-Tianjin (MOI, 20, 40 and
80) for 24 h. As shown in Fig. 4C, at
a MOI of 80, caspase-3, −8 and −9 activity was increased 2.80±0.15,
2.14±0.25 and 3.20±0.44 fold, respectively, compared with the
corresponding control groups. These results indicated that
UV-Tianjin-induced apoptosis may occur via a caspase-dependent
pathway.
Apoptosis in HeLa cells is induced via
extrinsic and intrinsic pathways by UV-Tianjin
Caspase activation is triggered by two pathways:
Extrinsic and intrinsic apoptotic pathways (23). To further investigate the molecular
mechanism of UV-Tianjin-induced apoptosis in HeLa cells, the
expression levels of various apoptosis-associated proteins,
including cyt c, Apaf-1, Fas, FasL, FADD, caspase-3, −8 and
−9, were examined by western blot analysis. The results revealed
that following UV-Tianjin treatment, the expression of the
intrinsic pathway related proteins, cyt c, Apaf-1, active
caspase-9 and −3, as well as extrinsic pathway-associated proteins,
Fas, FasL, FADD and active caspase-8, were increased in a
dose-dependent manner (Fig. 5). These
results indicated that UV-Tianjin-induced apoptosis in HeLa cells
may involve intrinsic and extrinsic apoptotic pathways.
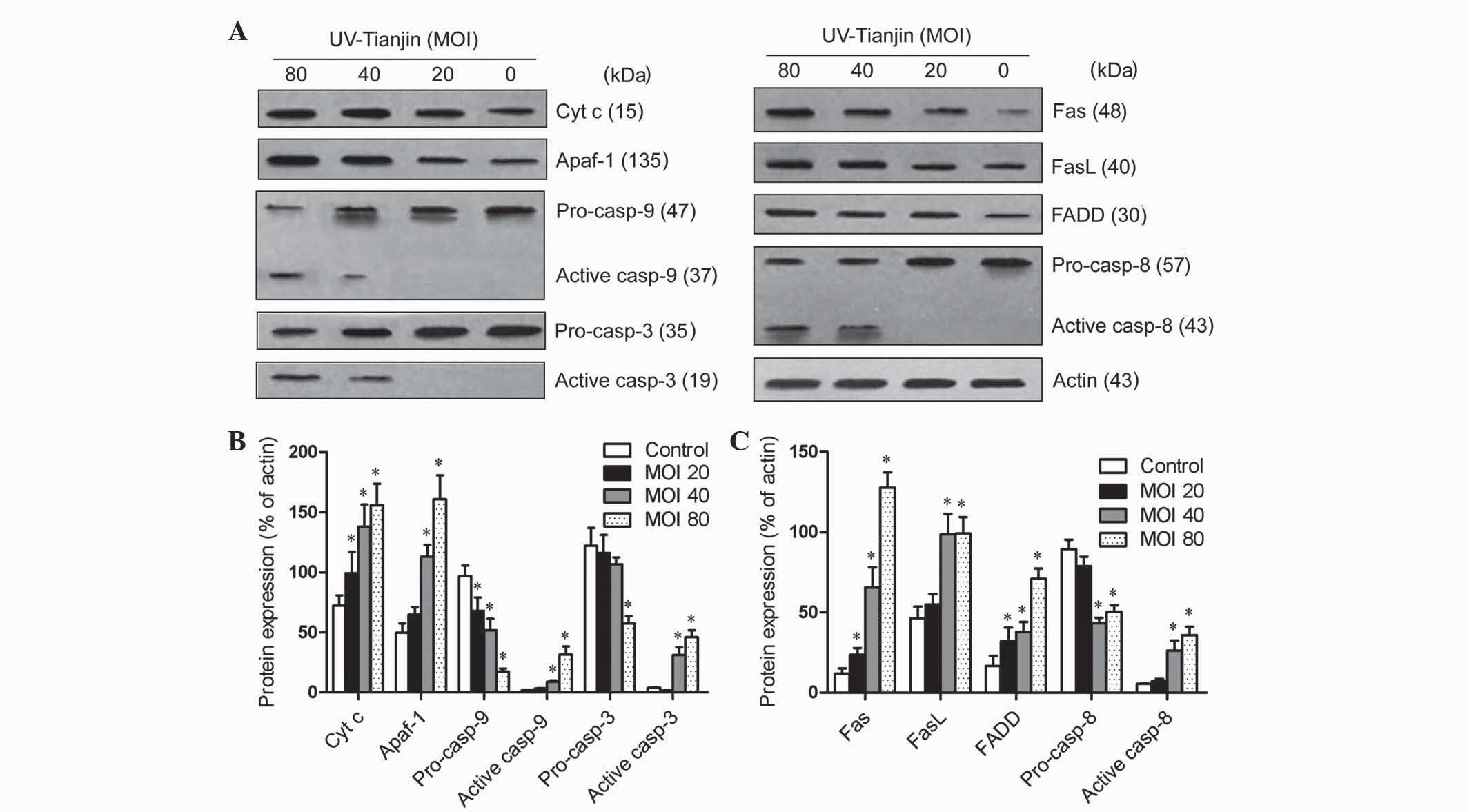 | Figure 5.Effects of UV-Tianjin on the
expression of apoptosis-associated proteins in human cervical
carcinoma HeLa cells. (A) HeLa cells were treated with UV-Tianjin
(MOI, 20, 40, 80) for 24 h and expression levels of
apoptosis-associated proteins were determined by western blot
analysis. (B) Relative expression levels of intrinsic pathway
apoptosis-associated proteins cyt c, Apaf-1, pro-casp-3, active
casp-3, pro-casp-9 and active casp-9 following UV-Tianjin
treatment. (C) Relative expression levels of extrinsic pathway
apoptosis-associated proteins Fas, FasL, FADD, pro-casp-8 and
active casp-8 following UV-Tianjin treatment. β-actin was used as
an internal control. Data are expressed as the mean ± standard
deviation of three independent experiments. *P<0.05 vs.
untreated control cells. UV-Tianjin, ultraviolet-inactivated Sendai
virus strain Tianjin; MOI, multiplicity of infection; cyt c,
cytochrome c; Apaf-1, apoptotic protease activating
factor-1; casp, caspase; FADD, Fas-associated protein with death
domain; FasL, Fas ligand. |
Discussion
The Sendai virus exhibits anticancer activity
against a variety of tumor types (11–13,24).
However, few studies have evaluated the antitumor activities of
Sendai virus in human cervical cancer, human small cell lung cancer
or human hepatocellular carcinoma. In the present study, the novel
genotype of Sendai virus Tianjin strain inhibited the proliferation
of human cervical cancer HeLa, human small cell lung cancer
NCI-H446 and human hepatoma Hep 3B cells in a dose- and
time-dependent manner.
Apoptosis is an important physiological process,
which leads to the elimination of redundant or harmful cells during
organism development (25). Apoptosis
is characterized by a number of biological and morphological
alterations, including changes in the MMP, activation of caspases,
DNA fragmentation, membrane blebbing and formation of apoptotic
bodies (26,27). In the present study, Hochest 33342
staining demonstrated that UV-Tianjin-treated HeLa, NCI-H446 and
Hep 3B cells exhibited typical apoptotic morphological alterations,
such as chromatin condensation and nuclear shrinkage. Furthermore,
Annexin V-FITC/PI double staining revealed that UV-Tianjin induced
apoptosis in a dose-dependent manner in all three cell lines and
that the HeLa cell line was most susceptible to UV-Tianjin
treatment. Subsequently, UV-Tianjin-induced apoptosis was further
confirmed in the HeLa cell line, as shown by MMP disruption and
increased caspase activity.
Caspases are expressed in the majority of cell types
as inactive pro-enzymes. Increasing evidence suggests that
activation of caspases initiates apoptosis in various types of cell
(28,29). Within the caspase family, caspase-8
and −9 are the two main initiator caspases in extrinsic and
intrinsic pathways that lead to the activation of effector
caspase-3 (30,31). By contrast to effector caspases, which
are activated by single intrachain cleavage that is mediated by an
initiator caspase, an initiator caspase requires recruitment to a
multimeric adaptor protein complex. Such multimeric protein
complexes are commonly known as apoptosome and death-inducing
signaling complexes; caspase-9 is activated by the Apaf-1
apoptosome and caspase-8 is activated by the adapter FADD (32–34). In
apoptotic cells, cyt c is released from mitochondria to the
cytoplasm, where it binds Apaf-1 (35,36).
Subsequent replacement of adenosine diphosphate by deoxyadenosine
triphosphate (dATP)/ATP in Apaf-1 leads to the formation of a
heptameric apoptosome (37–39). The autocatalytic activation of the
caspase-9 zymogen is catalyzed by the Apaf-1. The present study
demonstrated that UV-Tianjin triggered the intrinsic apoptotic
pathway by activating caspase-9 via Apaf-1 apoptosome activation
and the release of cyt c from the mitochondria in HeLa
cells. The extrinsic pathway is activated by the engagement of
death receptors on the cell surface. Trimerization of the
membrane-bound receptor, Fas, by its natural ligand, FasL, results
in recruitment of the receptor-specific adapter protein, FADD,
which subsequently recruits caspase-8 and promotes the cascade of
pro-caspase activation (40–42). The results of the present study also
demonstrated that UV-Tianjin activates the extrinsic apoptotic
pathway by activating caspase-8 via FADD and Fas activation in HeLa
cells. However, additional studies are required to investigate the
underlying molecular mechanisms of UV-Tianjin-induced apoptosis in
HeLa cells. Further studies are also required to determine whether
the intrinsic and extrinsic apoptotic pathways are involved in the
proapoptotic activity of UV-Tianjin in NCI-H446 and Hep 3B
cells.
In conclusion, the present study revealed that
UV-Tianjin induces apoptosis in HeLa, NCI-H446 and Hep 3B cells,
which indicates that UV-Tianjin may exhibit extensive anticancer
activity in human cancer cells. Furthermore, UV-Tianjin-induced
apoptosis in HeLa cells involves extrinsic and intrinsic apoptotic
pathways. Therefore, UV-Tianjin may present a potential biological
therapeutic agent for the treatment of human cancers.
Acknowledgements
This study was supported by the National Natural
Science Foundation of China, Beijing, China (grant no. 81172168)
and the National Training Program of Innovation and
Entrepreneurship for Undergraduates, Beijing, China (grant no.
201410062004).
References
|
1
|
Prat J, Franceschi S, Denny L and
Lazcano-Ponce E: Cancers of the female reproductive organs. World
Cancer Report 2014, World Health Organization. Stewart BW and Wild
CP: IARC Press. (Lyon). 678–680. 2014.
|
|
2
|
Forman D, Ferlay J, Stewart BW and Wild
CP: The global and regional burden of cancer. World Cancer Report
2014, World Health Organization. Stewart BW and Wild CP: IARC
Press. (Lyon). 28–30. 2014.
|
|
3
|
Theise ND, Chen CJ and Kew MC: Liver
cancer. World Cancer Report 2014, World Health Organization.
Stewart BW and Wild CP: IARC Press. (Lyon). 577–579. 2014.
|
|
4
|
Housman G, Byler S, Heerboth S, Lapinska
K, Longacre M, Snyder N and Sarkar S: Drug resistance in cancer: An
overview. Cancers (Basel). 6:1769–1792. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yu H, Park J, Lee J, Choi K and Choi C:
Constitutive expression of MAP kinase phosphatase-1 confers
multi-drug resistance in human glioblastoma cells. Cancer Res
Treat. 44:195–201. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Guan M, Romano G, Coroniti R and Henderson
EE: Progress in oncolytic virotherapy for treatment of thyroid
malignant neoplasm. J Exp Clin Cancer Res. 33:912014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Woller N, Gürlevik E, Ureche CI,
Schumacher A and Kühnel F: Oncolytic viruses as anticancer
vaccines. Front Oncol. 4:1882014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chiocca EA and Rabkin SD: Oncolytic
viruses and their application to cancer immunotherapy. Cancer
Immunol Res. 2:295–300. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shi JD, Li XM, Liu FY, Jin MJ, Zhang GJ,
Liu M and Li Mei: Isolation and identification of a lethal viral
pathogen from common cotton-eared marmosets. Virologica Sinica.
18:357–361. 2003.
|
|
10
|
Shi LY, Li M, Yuan LJ, Wang Q and Li XM: A
new paramyxovirus, Tianjin strain, isolated from common
cotton-eared marmoset: Genome characterization and structural
protein sequences analysis. Arch Virol. 153:1715–1723. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang Q and Xu X, Yuan Y, Gong X, Chen Z
and Xu X: IPS-1 plays a dual function to directly induce apoptosis
in murine melanoma cells by inactivated Sendai virus. Int J Cancer.
134:224–234. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Nomura M, Shimbo T, Miyamoto Y, Fukuzawa M
and Kaneda Y: 13-Cis retinoic acid can enhance the antitumor
activity of non-replicating Sendai virus particle against
neuroblastoma. Cancer Sci. 104:238–244. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Morodomi Y, Yano T, Kinoh H, Harada Y,
Saito S, Kyuragi R, Yoshida K, Onimaru M, Shoji F, Yoshida T, et
al: BioKnife, a uPA activity-dependent oncolytic Sendai virus,
eliminates pleural spread of malignant mesothelioma via
simultaneous stimulation of uPA expression. Mol Ther. 20:769–777.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Shi L, Chen J, Zhong Q, Li M, Geng P, He
J, Han Z, Sheng M and Tang H: Inactivated Sendai virus strain
Tianjin, a novel genotype of Sendai virus, inhibits growth of
murine colon carcinoma through inducing immune responses and
apoptosis. J Transl Med. 11:2052013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yuan CX, Zhou ZW, Yang YX, He ZX, Zhang X,
Wang D, Yang T, Wang NJ, Zhao RJ and Zhou SF: Inhibition of mitotic
Aurora kinase A by alisertib induces apoptosis and autophagy of
human gastric cancer AGS and NCI-N78 cells. Drug Des Devel Ther.
9:487–508. 2015.PubMed/NCBI
|
|
16
|
Fionda C, Abruzzese M, Zingoni A, Soriani
A, Ricci B, Molfetta R, Paolini R, Santoni A and Cippitelli M:
Nitric oxide donors increase PVR/CD155 DNAM-1 ligand expression in
multiple myeloma cells: Role of DNA damage response activation. BMC
Cancer. 15:172015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Freeman-Keller M and Weber JS:
Anti-programmed death receptor 1 immunotherapy in melanoma:
Rationale, evidence and clinical potential. Ther Adv Med Oncol.
7:12–21. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Park KC, Heo JH, Jeon JY, Choi HJ, Jo AR,
Kim SW, Kwon HJ, Hong SJ and Han KS: The novel histone deacetylase
inhibitor, N-hydroxy-7-(2-naphthylthio) hepatonomide, exhibits
potent antitumor activity due to cytochrome-c-release-mediated
apoptosis in renal cell carcinoma cells. BMC Cancer. 15:192015.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen J, Lin C, Yong W, Ye Y and Huang Z:
Calycosin and genistein induce apoptosis by inactivation of
HOTAIR/p-Akt signaling pathway in human breast cancer MCF-7 Cells.
Cell Physiol Biochem. 35:722–728. 2015.PubMed/NCBI
|
|
20
|
Nishizaki T, Kanno T, Tsuchiya A, Kaku Y,
Shimizu T and Tanaka A:
1-[2-(2-Methoxyphenylamino)ethylamino]-3-(naphthalene-1-yloxy)propan-2-ol
may be a promising anticancer drug. Molecules. 19:21462–21472.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kaneda Y, Nakajima T, Nishikawa T,
Yamamoto S, Ikegami H, Suzuki N, Nakamura H, Morishita R and Kotani
H: Hemagglutinating virus of Japan (HVJ) envelope vector as a
versatile gene delivery system. Mol Ther. 6:219–226. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Smiley ST, Reers M, Mottola-Hartshorn C,
Lin M, Chen A, Smith TW, Steele GD and Chen LB: Intracellular
heterogeneity in mitochondrial membrane potentials revealed by a
J-aggregate-forming lipophilic cation JC-1. Proc Natl Acad Sci USA.
88:3671–3675. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bredesen DE: Apoptosis: Overview and
signal transduction pathways. J Neurotrauma. 17:801–810. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gao H, Gong XC, Chen ZD, Xu XS, Zhang Q
and Xu XM: Induction of apoptosis in hormone-resistant human
prostate cancer PC3 cells by inactivated Sendai virus. Biomed
Environ Sci. 27:506–514. 2014.PubMed/NCBI
|
|
25
|
Kerr JF, Wyllie AH and Currie AR:
Apoptosis: A basic biological phenomenon with wide-ranging
implications in tissue kinetics. Br J Cancer. 26:239–257. 1972.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Schwartzman RA and Cidlowski JA:
Apoptosis: The biochemistry and molecular biology of programmed
cell death. Endocr Rev. 14:133–151. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Bergamaschi G, Rosti V, Danova M, Lucotti
C and Cazzola M: Apoptosis: Biological and clinical aspects.
Haematologica. 79:86–93. 1994.PubMed/NCBI
|
|
28
|
Frejlich E, Rudno-Rudzińska J, Janiszewski
K, Salomon L, Kotulski K, Pelzer O, Grzebieniak Z, Tarnawa R and
Kielan W: Caspases and their role in gastric cancer. Adv Clin Exp
Med. 22:593–602. 2013.PubMed/NCBI
|
|
29
|
Ola MS, Nawaz M and Ahsan H: Role of Bcl-2
family proteins and caspases in the regulation of apoptosis. Mol
Cell Biochem. 351:41–58. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cohen GM: Caspases: The executioners of
apoptosis. Biochem J. 326:1–16. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Pop C and Salvesen GS: Human caspases:
Activation, specificity, and regulation. J Biol Chem.
284:21777–21781. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Muzio M, Chinnaiyan AM, Kischkel FC,
O'Rourke K, Shevchenko A, Ni J, Scaffidi C, Bretz JD, Zhang M,
Gentz R, et al: FLICE, a novel FADD-homologous ICE/CED-3-like
protease, is recruited to the CD95 (Fas/APO-1) death-inducing
signaling complex. Cell. 85:817–827. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Srinivasula SM, Ahmad M, Fernandes-Alnemri
T and Alnemri ES: Autoactivation of procaspase-9 by Apaf-1-mediated
oligomerization. Mol Cell. 1:949–957. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hua Q, Wu D, Chen W, Yan Z, Yan C, He T,
Liang Q and Shi Y: Molecular determinants of caspase-9 activation
by the Apaf-1 apoptosome. Proc Natl Acad Sci USA. 46:16254–16261.
2014. View Article : Google Scholar
|
|
35
|
Li P, Nijhawan D, Budihardjo I,
Srinivasula SM, Ahmad M, Alnemri ES and Wang X: Cytochrome c and
dATP-dependent formation of Apaf-1/caspase-9 complex initiates an
apoptotic protease cascade. Cell. 91:479–489. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Liu X, Kim CN, Yang J, Jemmerson R and
Wang X: Induction of apoptotic programin cell-free extracts:
Requirement for dATP and cytochrome c. Cell. 86:147–157. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kim HE, Du F, Fang M and Wang X: Formation
of apoptosome is initiated by cytochrome c induced dATP hydrolysis
and subsequent nucleotide exchange on Apaf-1. Proc Natl Acad Sci
USA. 102:17545–17550. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Bao Q, Lu W, Rabinowitz JD and Shi Y:
Calcium blocks formation of apoptosome by preventing nucleotide
exchange in Apaf-1. Mol Cell. 25:181–192. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Acehan D, Jiang X, Morgan DG, Heuser JE,
Wang X and Akey CW: Three-dimensional structure of the apoptosome:
Implications for assembly, procaspase-9 binding, and activation.
Mol Cell. 9:423–432. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Nagata S and Golstein P: The Fas death
factor. Science. 267:1449–1456. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Waring P and Müllbacher A: Cell death
induced by the Fas/Fas ligand pathway and its role in pathology.
Immunol Cell Biol. 77:312–317. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Kruidering M and Evan GI: Caspase-8 in
apoptosis: The beginning of ‘the end’? IUBMB Life. 50:85–90. 2000.
View Article : Google Scholar : PubMed/NCBI
|