Introduction
Ovarian cancer is one of three most common
gynecological malignant tumors, along with cervical cancer and
endometrial carcinoma, with the highest morbidity and mortality
among these types of tumors. In 2008, it was estimated that 225,500
women were diagnosed with ovarian cancer and 140,200 women
succumbed to this disease worldwide; however, these figures vary
geographically (1). The fourth most
common type of cancer among gynecological malignant tumors
worldwide is epithelial ovarian cancer (EOC) (2). Due to the unfavorable anatomical
location of ovarian cancer, ~70% of ovarian cancer patients are
diagnosed at advanced stages, and the 5-year survival rate of these
patients is 20–30% (2). At present,
surgery and chemotherapy are the major treatment modalities of
ovarian cancer, but the effectiveness of treatment is poor. The
prognosis of patients with ovarian cancer is poor, due to late
diagnosis, since there are non-specific symptoms and a lack of
effective screening methods (2).
Furthermore, the etiology and early events in the progression of
ovarian cancer are poorly understood, and the genetic mechanism of
ovarian cancer development is extremely complex. Deletion, mutation
and/or altered expression of oncogenes and tumor suppressor genes
are important in ovarian cancer development (3). Speckle-type POZ (pox virus and zinc
finger protein) protein (SPOP) gene deletion or loss of
heterozygosity (LOH) in its loci has been reported in 57.8% of
breast cancer patients (4). In
addition, in 24 types of cancer, including ovarian cancer, copy
number analysis of SPOP amplification, LOH, deletion and mutation
revealed that LOH frequently appears in the SPOP locus (4).
SPOP is an E3 ubiquitin ligase adaptor protein,
which promotes substrate ubiquitination, leading to proteolysis of
the substrate via the 26S proteasome. In addition, the SPOP gene is
located on chromosome 17q21.33, which has frequent deletions and
mutations (5). Recent studies on
prostate and endometrial cancer demonstrated that the SPOP gene was
frequently mutated and its locus was observed to undergo LOH, which
indicated that SPOP may act as a tumor suppressor gene (5–8). In breast
cancer, the SPOP gene frequently undergoes copy number loss, which
also indicates that SPOP functions as a tumor suppressor gene
(4). Similarly in ovarian cancer, the
SPOP gene has been revealed to frequently undergo copy number loss
(4–6).
However, the role of SPOP gene in ovarian cancer has not been
confirmed.
A number of studies have revealed that SPOP is
frequently altered in human cancer. However, there are few studies
that focus on the genetic aberrations of the SPOP gene in ovarian
cancer. The aim of the present study was to investigate the role of
the SPOP gene in ovarian tumorigenesis, and to evaluate numerical
aberrations of chromosome 17 and SPOP gene alterations in an
ovarian cancer tissue microarray (TMA) with fluorescence in
situ hybridization (FISH).
Materials and methods
TMA
A TMA of ovarian cancer was purchased from Xi'an
Ailina Biotechnology Co., Ltd. (catalog no. BC11115a; Xi'an,
China), which consisted of 100 cases and 100 spots, including 5
cases of clear cell carcinoma, 63 cases of serous EOC, 10 cases of
mucous EOC, 2 cases of endometrial carcinoma, 10 cases of
metastatic adenocarcinoma and 10 cases of normal ovarian tissue.
Evaluation of the TMA by the present study was approved by Xi'an
Ailina Biotechnology Ethics Committee (The Second Affiliated
Hospital of Chongqing Medical University, Chongqing, China).
FISH
FISH technique was used on the TMA, with a
SPOP-specific DNA fluorescence probe used as the experimental group
and a centromere-specific DNA fluorescence probe for chromosome 17
[chromosome enumeration probe 17 (CEP17)] for the control group.
Fluorescence in situ hybridization Detection kit (catalog
no., G 100984R-8; included SPOP probe, CEP17 probe, SSC buffer,
RNase A, pepsin and hybridization buffer) was purchased from
Agilent Technologies, Inc. (Cedar Creek, TX, USA). The SPOP probe,
labeled red (rhodamine), covers a 190 Kb region of 17q21.33, which
includes the whole SPOP genome. The CEP17 control probe was labeled
green [fluorescein isothiocyanate (FITC)]. The FISH technique was
performed according to the manufacturer's protocol. Briefly, the
TMA samples were dewaxed, and underwent proteolysis for 10 min. The
probes were then added, followed by protein denaturation for 10 min
at 85°C and hybridization for 15–17 h at 37°C. The hybridization
was observed using a fluorescence microscope (BX51; Olympus
Corporation, Tokyo, Japan) equipped with selective filters for the
fluorochromes used.
Fluorescence signals were scored using a
well-established criteria (9). The
interpretation of the signals followed the same interpretation
criteria as previous studies using human epidermal growth factor
(HER-2) gene (3) and p16 gene
(10). A normal cell has two copies
of chromosome 17 (shown by two green spots) and two copies of SPOP
gene (shown by two red spots). For the SPOP FISH test the FISH
ratio was calculated as follows: FISH ratio (SPOP/CEP17) = number
of red rhodamine fluorescent SPOP signals / number of green FITC
fluorescent CEP17 signals. FISH was reported using the following
ratios: Deletion of SPOP gene, SPOP/CEP17 <0.7 or SPOP
signals/nucleus <1.5; normal SPOP gene, 0.7≤SPOP/CEP17≤2.0;
amplification of SPOP gene, SPOP/CEP17 >2.0; monosomy, >10%
cells with CEP17 signals/nucleus <1.5; polysomy, >10% cells
with CEP17 signals/nucleus >2.5; disome, CEP17 signals/nucleus =
1.5–2.5. The results were reviewed by two pathologists (The Center
of Pathological Diagnosis, Chongqing Medical University, Chongqing,
China).
Statistical analysis
SPSS version 19.0 software for Windows (IBM SPSS,
Armonk, NY, USA) was used. Bivariate analysis was performed using
χ2 test or likelihood-ratio test, and Fisher's exact
probability method when required. P<0.05 was considered to
indicate a statistically significant difference.
Results
Status of SPOP gene and CEP17
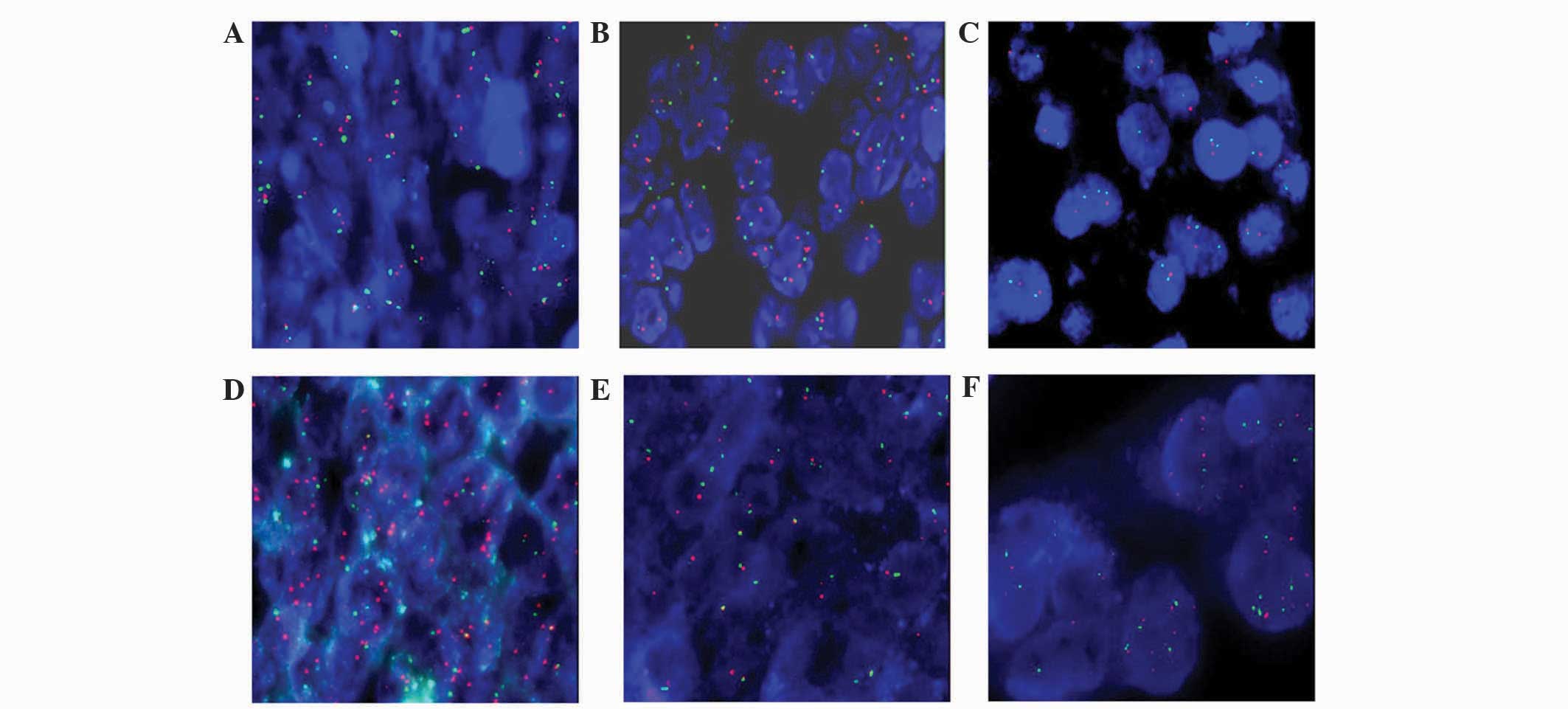
In total, 98 out of 100 cases from the TMA were
evaluated; 2 cases of serous EOC could not be analyzed, due to
unclear fluorescence with FISH. No alterations in the SPOP gene or
CEP17 were observed in normal ovarian tissue using FISH (Fig. 1A). In ovarian cancer, 37 cases (37/88;
42.05%) were classified as disome and the SPOP genes were normal
(Fig. 1B). Comparatively, the SPOP
genes in 51 cases (51/88; 57.95%) were abnormal as follows:
deletion, 1 case (Fig. 1C);
amplification, 1 case (Fig. 1D);
monosomy, 45 cases (Fig. 1E); and
polysomy, 4 cases (Fig. 1F).
Differences in SPOP genes between
histological types and grades of ovarian cancer
The status of SPOP genes in ovarian cancer are shown
in Table I. The status of the SPOP
gene in ovarian cancer and normal ovarian tissues differed
significantly (P<0.01; Table II).
In addition for ovarian cancer, the status of the SPOP gene in
serous EOC and clear cell carcinoma differed significantly
(P<0.01), while the status of the SPOP gene of serous EOC and
other histological subtypes did not differ significantly
(P>0.05; Tables III and IV). The present data revealed that the SPOP
gene differed significantly between grade 3 and grade 1 serous EOC
(P<0.01). Similarly, the status of the SPOP gene differed
significantly between grade 3 and grade 2 serous EOC (P<0.01).
However, the status of the SPOP gene of grade 1 and grade 2 did not
differ significantly in serous EOC (P=0.104) (Tables V and VI).
 | Table I.SPOP/CEP17 in ovarian cancer and
normal ovarian tissues. |
Table I.
SPOP/CEP17 in ovarian cancer and
normal ovarian tissues.
| Ratio
(SPOP/CEP17) | Status of SPOP
gene | Status of CEP17 | Ovarian cancer
tissue, n | Normal ovarian
tissue, n |
|---|
| Total |
|
| 88 | 10 |
| Ratio >2 | Amplification | – | 1 | 0 |
| 0.7≤ratio≤2;
CEP17<1.5 | – | Monosomy | 45 | 0 |
| 0.7≤ratio≤2;
1.5≤CEP17≤2.5 | – | Disome | 37 | 10 |
| 0.7≤ratio≤2; CEP17
>2.5 | – | Polysomy | 4 | 0 |
| Ratio <0.7 | Deletion | – | 1 | 0 |
| Table II.SPOP gene status between ovarian
cancer and normal ovarian tissues. |
Table II.
SPOP gene status between ovarian
cancer and normal ovarian tissues.
|
|
| SPOP gene status |
|---|
|
|
|
|
|---|
| Tissue type | Total, n | Abnormal, n | Normal, n |
|---|
| Ovarian
cancera | 88 | 51 | 37 |
| Normal ovarian | 10 | 0 | 10 |
| Table III.SPOP gene status between different
types of EOC. |
Table III.
SPOP gene status between different
types of EOC.
|
|
| SPOP gene status |
|---|
|
|
|
|
|---|
| Histological
type | Total, n | Abnormal, n | Normal, n |
|---|
| Total | 88 | 51 | 37 |
| Serous EOC | 41 | 20 | 61 |
| Mucinous EOC | 4 | 6 | 10 |
| Clear cell
carcinoma | 0 | 5 | 5 |
| Endometrioid
carcinoma | 0 | 2 | 2 |
| Metastatic
adenocarcinoma | 6 | 4 | 10 |
| Table IV.Bivarate analysis of speckle-type POZ
(pox virus and zinc finger protein) gene status between different
types of EOC. |
Table IV.
Bivarate analysis of speckle-type POZ
(pox virus and zinc finger protein) gene status between different
types of EOC.
| Histological
type | χ2 | P-value |
|---|
| Serous EOC vs.
mucinous EOC | 2.635 | 0.105a |
| Serous EOC vs. clear
cell |
| 0.006b |
| Serous EOC vs.
endometrioid |
| 0.118b |
| Serous EOC vs.
metastatic | 0.195 | 0.658a |
| Table V.SPOP gene status between pathological
grades of serous epithelial ovarian cancer. |
Table V.
SPOP gene status between pathological
grades of serous epithelial ovarian cancer.
|
|
| SPOP gene status |
|---|
|
|
|
|
|---|
| Grade | Total | Abnormal, n | Normal, n |
|---|
| Total | 61 | 41 | 20 |
| 1 | 2 | 8 | 10 |
| 2 | 8 | 6 | 14 |
| 3 | 31 | 6 | 37 |
| Table VI.Bivarate analysis of speckle-type POZ
(pox virus and zinc finger protein) protein gene status between
pathological grades of serous epithelial ovarian cancer. |
Table VI.
Bivarate analysis of speckle-type POZ
(pox virus and zinc finger protein) protein gene status between
pathological grades of serous epithelial ovarian cancer.
| Grade | χ2 | P-value |
|---|
| 1 vs. 2 |
|
0.104a |
| 2 vs. 3 |
4.006 |
0.045b |
| 3 vs. 1 | 14.443 |
<0.001c |
Discussion
The present study demonstrated that there is a
significant difference in the status of SPOP genes between ovarian
cancer and normal ovarian tissues using FISH assay on a TMA. The
present results reveal that in ovarian cancer, SPOP gene copies
undergo deletion and amplification, and chromosome 17 undergoes
monosomy, disome and polysomy. In addition, out of 88 cases of
ovarian cancer, there were 45 cases of monosomy 17 (45/88; 51.14%);
monosomy 17 results in the deletion of SPOP gene. By contrast, the
present study observed that the SPOP gene and chromosome 17 were
unaltered in normal ovarian tissues (Tables I–III).
The data gathered in the present study indicated
that the deletion of the SPOP gene, in association with monosomy
17, may contribute to ovarian carcinogenesis. In addition, the
present results demonstrate that there is a high proportion (45/88;
51.14%) of monosomy 17 in ovarian cancer (Table I), and indicates that chromosome 17
has undergone loss of a certain number of tumor suppressor genes.
Furthermore, there may be a positive association between
histological subtypes and tumor grade and the deletion of the SPOP
gene in ovarian cancer (Table IV and
VI, respectively). The deletion of
the SPOP gene appeared in a large proportion of serous EOC tissues
(41/61; 67.21%), particularly in grade 3 EOC (31/37; 83.78%)
(Tables III–VI).
Previously, several studies concerning the genetic
alterations in ovarian carcinogenesis have identified specific
molecular markers that may provide a novel method of diagnosing
ovarian cancer; therefore, these markers may potentially increase
the prognosis of ovarian cancer patients and provide novel
therapeutic strategies (11–14). The present study demonstrated that the
SPOP gene loses its function when it is inactive, since the
inactivation of the SPOP gene is primarily caused by deletion, LOH
or mutation of the gene (4,6). However, investigation concerning the
SPOP protein indicated that SPOP protein function may also alter
when it translocates between the cell nucleus and the cytoplasm in
clear cell renal cell carcinoma (ccRCC), which indicates that the
SPOP protein serves as a regulatory hub to promote ccRCC
tumorigenesis (15).
In addition to the inactivation of the SPOP gene,
certain studies have identified that inactivation of certain genes
located in chromosomal region 17q21, including breast cancer 1 and
putative oncoprotein nm23, may contribute to ovarian carcinogenesis
(16). Additionally, the present
study data may indicate that a number of oncogenes that are
associated with polysomy 17, including HER-2 and growth factor
receptor bound protein 7 gene, may contribute to ovarian
carcinogenesis (4). Therefore, the
present study hypothesizes that alterations in chromosome 17 may be
important in ovarian carcinogenesis.
SPOP is an adaptor for E3 ligase Cullin3, a nuclear
protein, which promotes substrate protein ubiquitination and
degradation by the 26S proteasome (4,6). Sequence
analysis indicates that the SPOP protein contains a nuclear
localization signal (NLS), which locates the SPOP protein to the
cell nucleus leading to a promotion of cell apoptosis (15). The function of the SPOP protein is
altered when it translocates from the nucleus to the cytoplasm, and
in this situation it is referred to as SPOP-cyto, since it lacks
the NLS (15).
Recent studies concerning the structures of
SPOP-substrate complexes have revealed that SPOP-substrates require
SPOP-binding consensus motifs φ-π-S-S/T-S/T (φ, nonpolar; π, polar)
as a prerequisite for binding to the SPOP protein (17). Certain protein substrates, including
death-domain-associated-protein (Daxx), are involved in
transcription, cell-cycle regulation and apoptosis, and may bind to
the MATH domain of the SPOP protein, leading to ubiquitination and
degradation in the proteasome of the substrate. Therefore, the SPOP
protein serves as a regulator of cellular function (6). At present, known SPOP substrates include
macro H2A, Puckered, Daxx, glioma-associated-oncogene and
phosphatase and tensin homolog (17).
Several studies concerning SPOP gene function
indicate that the mutation or LOH of the SPOP gene is critical in
prostate and breast cancer (4,5). In
addition, the SPOP protein is key in ubiquitin-mediated degradation
of steroid receptor coactivator-3 by 26S proteasome in breast
cancer (4). These results suggest
that SPOP may act as a tumor suppressor in breast and prostate
cancers. The SPOP protein is primarily located in the cell nucleus,
but SPOP-cyto and hypoxia drive the SPOP protein to become
accumulated in the cytoplasm in kidney cancer, where the function
of the SPOP protein may alter from proapoptotic to antiapoptotic
and induce proliferation (15). This
indicates that the SPOP protein may act as a tumor promoter in
ccRCC.
The contradiction of tumor-promoting and
tumor-suppressing roles of SPOP may be partially attributed to the
following factors: Difference in substrate function due to
different subcellular localization of the SPOP protein, and a
difference in the expression of SPOP substrates owing to diverse
cell and cancer varieties. In breast cancer, the SPOP protein binds
to substrates that are tumor-promoters and therefore plays a
tumor-suppressing role (4). In
prostate cancer, the mutated SPOP gene does not bind to substrates
that are tumor-suppressors, and so SPOP may play a
tumor-suppressing role (5,6). In conclusion, the role of SPOP is
associated with the status of the SPOP protein and the function of
substrates (6).
TMA is a cost- and time-efficient method for
high-throughput analysis. The TMA used in the present study
contained 90 spots of ovarian cancer and 10 spots of normal ovarian
tissue. Combined with the FISH technique, TMA limits the damage to
specimens and creates consistent experimental conditions, thus
ensuring the stability of experimental results.
In conclusion, the FISH technique is an experimental
method that uses a SPOP DNA probe (red) and a CEP17 probe (green).
It is a valuable method that concurrently detects numerical
abnormalities in chromosome 17 and numerical alterations in SPOP
gene copies. The results of the present study indicate that the
deletion of the SPOP gene was observed in a large proportion of
serous EOC cases, particularly in grade 3 serous EOC. The use of
molecular diagnosis technologies allows for a high-throughput
analysis of genes, therefore providing novel insights into gene
expression profiles in ovarian cancer. Consequently, it may promote
the development of novel biomarkers or aid in the identification of
a novel therapy for human cancer. The present study has revealed
that investigating the molecular mechanisms of the deletion of the
SPOP gene in ovarian cancer is an important direction for future
study.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (Beijing, China; grant no.
81100443), Chongqing Yuzhong District Science and Technology Plan
projects (Chongqing, China; grant no. 20110303) and Chongqing
Municipal Health Bureau Scientific Research Project (Chongqing,
China; grant no. 20121039).
References
|
1
|
Lowe KA, Chia VM, Taylor A, O'Malley C,
Kelsh M, Mohamed M, Mowat FS and Goff B: An international
assessment of ovarian cancer incidence and mortality. Gynecol
Oncol. 130:107–114. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mayr D, Kanitz V, Amann G, Engel J, Burges
A, Löhrs U and Diebold J: HER-2/neu amplification in ovarian
tumours: A comprehensive immunohistochemical and FISH analysis on
tissue microarrays. Histopathology. 48:149–156. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chao WR, Lee MY, Lin WL, Chen CK, Lin JC,
Koo CL, Sheu GT and Han CP: HER2 amplification and overexpression
are significantly correlated in mucinous epithelial ovarian cancer.
Hum Pathol. 45:810–816. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Li C, Ao J, Fu J, Lee DF, Xu J, Lonard D
and O'Malley BW: Tumor-suppressor role for the SPOP ubiquitin
ligase in signal-dependent proteolysis of the oncogenic
co-activator SRC-3/AIB1. Oncogene. 30:4350–4364. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Geng C, He B, Xu L, Barbieri CE, Eedunuri
VK, Chew SA, Zimmermann M, Bond R, Shou J, Li C, et al: Prostate
cancer-associated mutations in speckle-type POZ protein (SPOP)
regulate steroid receptor coactivator 3 protein turnover. Proc Natl
Acad Sci USA. 110:6997–7002. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mani RS: The emerging role of speckle-type
POZ protein (SPOP) in cancer development. Drug Discovery Today.
19:1498–1502. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kim MS, Je EM, Oh JE, Yoo NJ and Lee SH:
Mutational and expressional analyses of SPOP, a candidate tumor
suppressor gene, in prostate, gastric and colorectal cancers.
APMIS. 121:626–633. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Theurillat JP, Udeshi ND, Errington WJ,
Svinkina T, Baca SC, Pop M, Wild PJ, Blattner M, Groner AC, Rubin
MA, et al: Ubiquitylome analysis identifiesdysregulation of
effector substrates inSPOP-mutant prostate cancer. Science.
346:85–89. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hopman AH, Ramaekers FC, Raap AK, Beck JL,
Devilee P, van der Ploeg M and Vooijs GP: In situ hybridization as
a tool to study numerical chromosome aberrations in solid bladder
tumors. Histochemistry. 89:307–316. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Aravidis C, Panani AD, Kosmaidou Z,
Thomakos N, Rodolakis A and Antsaklis A: Detection of numerical
abnormalities of chromosome 9 and p16/CDKN2A gene alterations in
ovarian cancer with fish analysis. Anticancer Res. 32:5309–5313.
2012.PubMed/NCBI
|
|
11
|
Puig S, Ruiz A, Lázaro C, Castel T, Lynch
M, Palou J, Vilalta A, Weissenbach J, Mascaro JM and Estivill X:
Chromosome 9p deletions in cutaneous malignant melanoma tumors: The
minimal deleted region involves markers outside the p16 (CDKN2)
gene. Am J Hum Genet. 57:395–402. 1995.PubMed/NCBI
|
|
12
|
Caldas C, Hahn SA, da Costa LT, Redston
MS, Schutte M, Seymour AB, Weinstein CL, Hruban RH, Yeo CJ and Kern
SE: Frequent somatic mutations and homozygous deletions of the p16
(MTS1) gene in pancreatic adenocarcinoma. Nat Genet. 8:27–32. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Williamson MP, Elder PA, Shaw ME, Devlin J
and Knowles MA: p16 (CDKN2) is a major deletion target at 9p21 in
bladder cancer. Hum Mol Genet. 4:1569–1577. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Panani AD, Maliaga K, Babanaraki A and
Bellenis I: Numerical abnormalities of chromosome 9 and p16CDKN2A
gene deletion detected by FISH in non-small cell lung cancer.
Anticancer Res. 29:4483–4487. 2009.PubMed/NCBI
|
|
15
|
Li G, Ci W, Karmakar S, Chen K, Dhar R,
Fan Z, Guo Z, Zhang J, Ke Y, Wang L, et al: SPOP promotes
tumorigenesis by acting as a key regulatory hub in kidney cancer.
Cancer Cell. 25:455–468. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kato H, Arakawa A, Suzumori K, Kataoka N
and Young SR: FISH analysis of BRCA1 copy number in
paraffin-embedded ovarian cancer tissue samples. Exp Mol Pathol.
76:138–142. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhuang M, Calabrese MF, Liu J, Waddell MB,
Nourse A, Hammel M, Miller DJ, Walden H, Duda DM, Seyedin SN, et
al: Structures of SPOP-substrate complexes: Insights into molecular
architectures of BTB-Cul3 ubiquitin ligases. Mol Cell. 36:39–50.
2009. View Article : Google Scholar : PubMed/NCBI
|