Introduction
Macroautophagy, which is also referred to as
autophagy, is a protein degradation process in eukaryotic cells.
The role of autophagy in cancer treatment has been extensively
studied, yet the results remain controversial. Although certain
studies indicate that autophagy participates in chemotherapy
resistance, the mechanism is not yet clear (1). A recent series of studies suggest that
activating molecule in Beclin 1-regulated autophagy protein 1
(Ambra1) is an important factor in regards to the association
between autophagy and apoptosis, and may control the reciprocal
conversion between the two processes to decide the resulting cell
death or survival (2–4). Therefore, Ambra1 may be an important
factor of autophagy involved in cancer treatment and chemotherapy
resistance. The present review focuses on the role of Ambra1 in
autophagy and apoptosis and assesses the implications for cell
survival and chemotherapy resistance.
Autophagy process and its dual role in cell
death and survival
Autophagy is an evolutionarily conserved
lysosome-dependent cellular catabolic degradation process in
eukaryotic cells (5). In general,
basal autophagy exists in cells to maintain cellular homeostasis
through the degradation of long-lived proteins, protein aggregates
and damaged organelles. However, autophagy is rapidly upregulated
under adverse conditions, including nutrient deprivation, hypoxia,
radiation and anticancer drugs, to recycle energy and supply
macromolecules for biosynthesis, leading to the cells adapting to
the stress and survival (5–13). Three major types of autophagy have
been reported, including macroautophagy, microautophagy and
chaperone-mediated autophagy (CMA) (10). Autophagy is highly regulated by a
series of autophagy-related genes (ATGs) that are essential for the
formation, maturation and traffic of autophagosomes (8,14). At
present, >30 ATGs have been identified in yeast, the majority of
which have a mammalian homolog (15).
Several complexes are essential at the initial stage
of autophagy, including the mammalian target of rapamycin (mTOR)
complex, Unc-51 like kinase-1 (ULK1) complex and the mammalian
ortholog of the ATG6/vacuolar protein-sorting protein (Vps)30
(Beclin 1)-class III phosphatidylinositol 3-kinase (CIII
PI3K/Vps34) complex (Vps34 complex) (16–18). The
mTOR complex includes two distinct complexes, mTOR complex 1
(mTORC1) and mTOR complex 2 (mTORC2). The mTORC1 is a sensor of
amino acids, adenosine triphosphate (ATP), growth factors and
reactive oxygen species (ROS). Usually, mTORC1 combines with the
ULK1 complex, leading to the phosphorylation of ULK1 and mammalian
ATG13 (mATG13), thus deactivating the ULK1 complex and blocking
autophagy (Fig. 1A) (16).
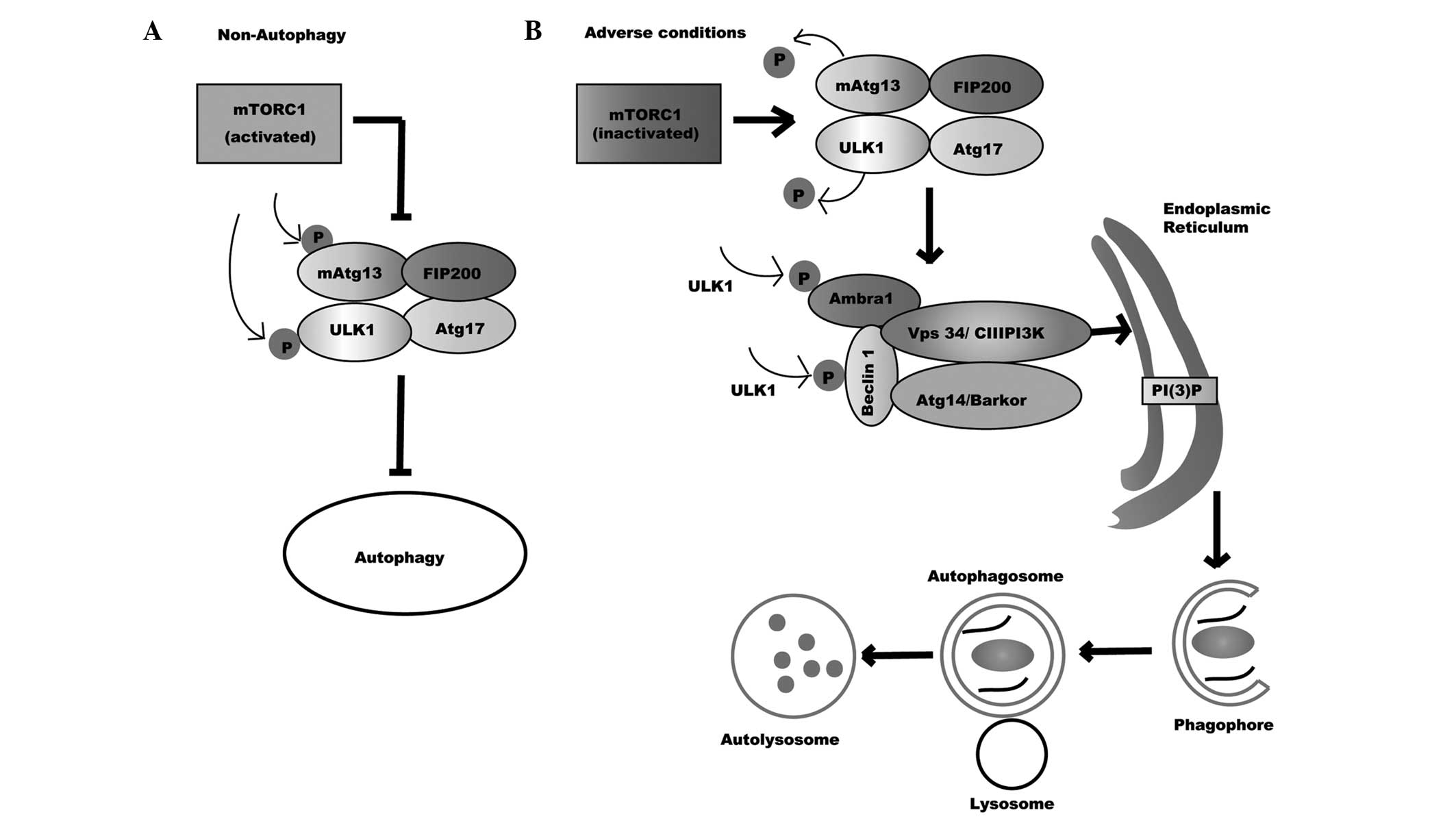 | Figure 1.Autophagy process. (A) Non-autophagy,
mTORC1 binds and deactivates ULK1 complex through phosphorylating
ULK1 and mATG13, thus blocking autophagy. (B) Under adverse
conditions, the inactivation of mTORC1 makes ULK1 and mATG13
dephosphorylate, prompting the activation of the ULK1 complex.
Activated ULK1 then phosphorylates Beclin 1 and Ambra1, leading to
the formation and activation of the Vps34 complex. Activated ULK1
directs the Vps34 complex to the endoplasmic reticulum, where Vps34
catalyzes the transformation of PI into PI-3-P, which recruits the
specific autophagic proteins that are required for the formation of
phagophores. Subsequently, the phagofores elongate and fuse to form
autophagosomes. After the maturation of autophagosomes, the outer
membrane of it eventually fuses with lysosomes to form
autophagolysosomes. Lastly, the contents of the autolysosomes are
digested by lysosomal hydrolases. mTORC1, mammalian target of
rapamycin complex 1; ULK1, Unc-51 like kinase-1; mATG13, mammalian
autophagy-related gene 13; Ambra1, activating molecule in Beclin
1-regulated autophagy protein 1; Vps34/CIII PI3K, class III
phosphatidylinositol 3-kinase; ATG17, autophagy-related gene 17;
FIP200, mammalian ortholog of ATG17; ATG14/Barkor,
autophagy-related gene 14; PI(3)P,
phosphatidylinositol-3-phosphate. |
The ULK1 complex contains the mammalian ortholog of
ATG1 (ULK1/2), mATG13, the mammalian ortholog of ATG17 (FIP200) and
ATG101 (17). Under adverse
conditions, the inactivation of mTORC1 results in the
dephosphorylation of ULK1 and mATG13, prompting the activation of
the ULK1 complex (16,17). The activated ULK1 phosphorylates
Beclin 1 and Ambra1 to promote the formation and activation of the
Vps34 complex. In addition, the activated ULK1 directs the Vps34
complex to the endoplasmic reticulum (ER), where Vps34 catalyzes
the transform of phosphatidylinositol (PI) into PI-3-phosphate,
which recruits the specific autophagic proteins that are required
for the formation of phagophores (Fig.
1B) (16–18).
The Vps34 complex is the core machinery of autophagy
initiation, and is composed by Beclin 1, Vps34/CIII PI3K and Vps15
(19,20). Beclin 1, a B-cell lymphoma
(Bcl)-2-homology (BH)3 domain only protein, is identified as a
binding protein of Bcl-2. It is an indispensible protein in
mammalian autophagy induction (21–23).
Beclin 1 contains three identified structural domains, including a
BH3 domain, a central coiled-coil domain (CCD) and an
evolutionarily conserved domain (ECD) (18,24–26). On
the one hand, Bcl-2 family members, including Bcl-2, Bcl-extra
large and myeloid cell leukemia 1, interact with the BH3 domain to
block the interaction of Beclin 1 with Vps34/CIII PI3K, decreasing
Vps34/CIII PI3K activity and negatively regulating Beclin
1-dependent autophagy (18,21–26) On the
other hand, Beclin 1 directly binds to Vps34 with the ECD and CCD
domains to form the Vps34 complex and arouse autophagic cascades
(18). In addition, Beclin 1
regulates autophagy through various steps by associating with other
specific proteins, including Ambra1, ATG14/Barkor, UV radiation
resistance-associated gene and RUN and cysteine rich domain
containing Beclin 1 interacting protein at the CCD domain (18,22,26–28).
In one previous study, Ambra1, a cofactor of Beclin 1, was also
shown to be an inseparable part of the core Vps34 complex and a
positive regulator of autophagy (2).
Subsequently, the phagofores elongate and fuse to form
double-membrane vesicles called autophagosomes. In the process of
phagofore elongation, two ubiquitin-like conjugation systems are
required, including the Atg12-Atg5 conjugation and ATG8/LC3-PE
(phosphatidylethanolamine) system. Following the maturation of
autophagosomes, the outer membrane eventually fuses with lysosomes
to form autolysosomes (8,14,15). The
contents within the autolysosomes are digested by lysosomal
hydrolases, the resultant macromolecules of which are recycled and
catabolized, thus producing energy to aid the adaptation of cells
to starvation or stress and contributing to cell survival (6,12).
Although autophagy is primary to contribute to cell
survival, in certain conditions, it can directly lead to cell
death, particularly in the cells with apoptotic machinery
deficiency (6,12). This type of cell death is called
autophagic cell death or type II programmed cell death (6,12). At
present, the mechanisms of cell death directly induced by autophagy
have not been fully clarified. The recognized interpretation is
that sustained autophagy leads to excessive degradation of
necessary proteins and organelles, or induces the high threshold
apoptosis (29). In cancer cells,
autophagy has been previously indicated to be upstream to apoptosis
in ER stress-induced death (30).
Therefore, autophagy is a highly regulated process
that has a dual role in cell death or survival. In addition, the
role of autophagy in determining the outcome of cells is dependent
on the context and cell type.
Autophagy in cancer therapeutic
responsiveness and chemotherapy resistance
Autophagy in cancer treatment
Previously, a growing body of evidence has revealed
that a variety of cancer treatments, including chemotherapy,
irradiation, endocrine therapy and molecular-targeted therapy, can
induce autophagy in diverse cancer cell lines, and that
induced-autophagy may be associated with therapeutic effects
(1,7,19).
Unfortunately, the role of autophagy in cancer treatment remains
controversial. Certain studies have shown that autophagy induced by
anticancer treatment is a pro-death mechanism. For example, histone
deacetylase inhibitor, suberoylanilide hydroxamic acid,
5-fluorouracil, sorafenib and imatinib, could respectively induce
cell death by autophagy in the cells of breast cancer, colorectal
cancer (CRC), hepatocellular carcinoma and glioma; while the
inhibition of autophagy with chemical reagents, such as
3-methyladenine, or small interfering RNAs (siRNAs), including
Beclin1 or ATG5, may suppress cell death (31–34). As a
result, autophagy is beneficial to cancer treatment. On the
contrary, increasing studies have shown that autophagy induced by
anticancer treatment plays a protective role (35–55).
Therefore, the blockage of autophagy is beneficial to cancer
treatment. Based on the results, >30 clinical trials have been
opened to investigate the effectiveness of chloroquine (CQ) or
hydroxychloroquine (HCQ) plus or minus chemotherapeutic or
targeted-drugs in human cancers (1).
CQ and its derivative HCQ raise the lysosomal pH and ultimately
inhibit the fusion between autophagosomes and lysosomes, thus
preventing the maturation of autophagosomes into autolysosomes, and
blocking a late step of autophagy (56,57).
Notably, numerous trials have supplied evidence of preliminary
anticancer activity of CQ or HCQ (1).
Autophagy in chemotherapy
resistance
Chemotherapy is one of the major means of cancer
treatment. However, the resistance of cancer cells to drugs
seriously limits their use. The mechanisms underlying drug
resistance are numerous, including reduced drug absorption, the
reduced ability of drugs to eliminate cells due to diverse changes
in cells, increased efflux pump and so on (58,59). To
date, these mechanisms are not entirely understood.
The protective role of autophagy in cancer treatment
suggests that autophagy may be involved in anticancer drug
resistance. In fact, the role of autophagy in anticancer drug
resistance has been confirmed by the results from several breast
cancer cell lines (60–65). Epirubicin can induce autophagy in
MCF-7 cells, and the induced-autophagy protects the cells from
death by blocking apoptosis (60).
Similarly, this reagent also induces autophagy in derived MCF-7er
cells (a type of induced epirubicin-resistant cell), and the
inhibition of induced-autophagy by chemical inhibitors, such as
baflomycin A1, or small hairpin RNAs (shRNAs), including sh-Beclin
1 and sh-ATG7, restores the sensitivity of MCF-7er cells to
epirubicin through enhancing apoptosis (60). MCF-7er cells also generate resistance
to paclitaxel (PTX) and vinorelbine (NVB), which suggests that the
cells obtain a multidrug resistance (MDR) phenotype (61). Furthermore, the inhibition of PTX and
NVB induced-autophagy makes the cells more sensitive to these
drugs, indicating the autophagy is also involved in MDR development
(61). In addition, Chittaranjan
et al (62) indicated that the
blockage of epirubicin-induced autophagy augments the anticancer
effects of epirubicin in triple-negative breast cancer MDA-MB-231
cells, derived resistant MDA-MB-231-R8 cells and SUM159PTR75 cells.
These results suggest that autophagy facilitates epirubicin
resistance development by blocking apoptosis. Additionally, other
studies also demonstrate that the induced-autophagy in various
breast cancer cells contributes to the development of resistance to
paclitaxel, tamoxifen or herceptin (63–65).
Notably, these studies indicated that derived resistant breast
cancer cells obtained an increased capability for autophagy
compared with the parental cells, which further suggests that
autophagy is involved in the drugs resistance. Nevertheless, the
molecular mechanisms of autophagy in drug resistance are complex
and not yet established. A number of factors or signaling pathways
may participate in the process, including epidermal growth factor
receptor signaling, the PI3K/protein kinase B/mTOR axis, tumor
protein 53 and mitogen-activated protein kinase 14/p38 signaling
(1). In the future, additional
studies in vitro and in vivo are required to confirm
the role of autophagy in drug resistance and to explore the
underlying mechanisms.
Ambra1 is a positive factor of
autophagy
Ambra1 is a Beclin 1-interacting protein that
contains a WD40 domain. It is primarily expressed in neural tissues
and is essential for normal neural tube development (3). In mouse embryos, the functional
deficiency of Ambra1 results in serious neural tube defects, which
are associated with autophagy impairment, accumulation of
ubiquitinated proteins, unbalanced cell proliferation and excessive
apoptotic cell death (3). The
overexpression of Ambra1 in rapamycin-treated human fibroblast
2FTGH cells has been shown to significantly increase basal and
rapamycin-induced autophagy (3). On
the contrary, downregulation of Ambra1 results in an evident
decrease in autophagy induced by rapamycin and nutrient deficiency
(3). Therefore, Ambra1 is a positive
factor of autophagy induction. The functions of Ambra1 in autophagy
regulation are mainly through interaction with mTORC1, ULK1, Beclin
1, dynein light chain 1/2 (DLC 1/2) and Bcl-2 located at the
mitochondria (mito-Bcl-2) (2,66–69).
Normally, Ambra1 physically interacts with mTORC1,
which results in Ambra1 deactivation by phosphorylation at Ser 52
(Fig. 2A) (67). However, under stress conditions,
mTORC1 inactivates and ULK1 activates, which results in the
activation of Ambra1 and Beclin 1 through phosphorylation and leads
to autophagy induction (Fig. 1B)
(67). Recently, Nazio et al
(67) have found that Ambra1 is a
ULK1-binding partner that is required for ULK1 stability and kinase
activity. On autophagy induction, Ambra1 mediates ULK1
Lys-63-linked ubiquitylation through interaction with the E3-ligase
tumor necrosis receptor associated factor 6 (Fig. 2A) (67).
Ubiquitylation enhances ULK1 stability, kinase activity and
self-association; therefore, there is a positive regulation loop
between Ambra1 and ULK1 in autophagy regulation (67).
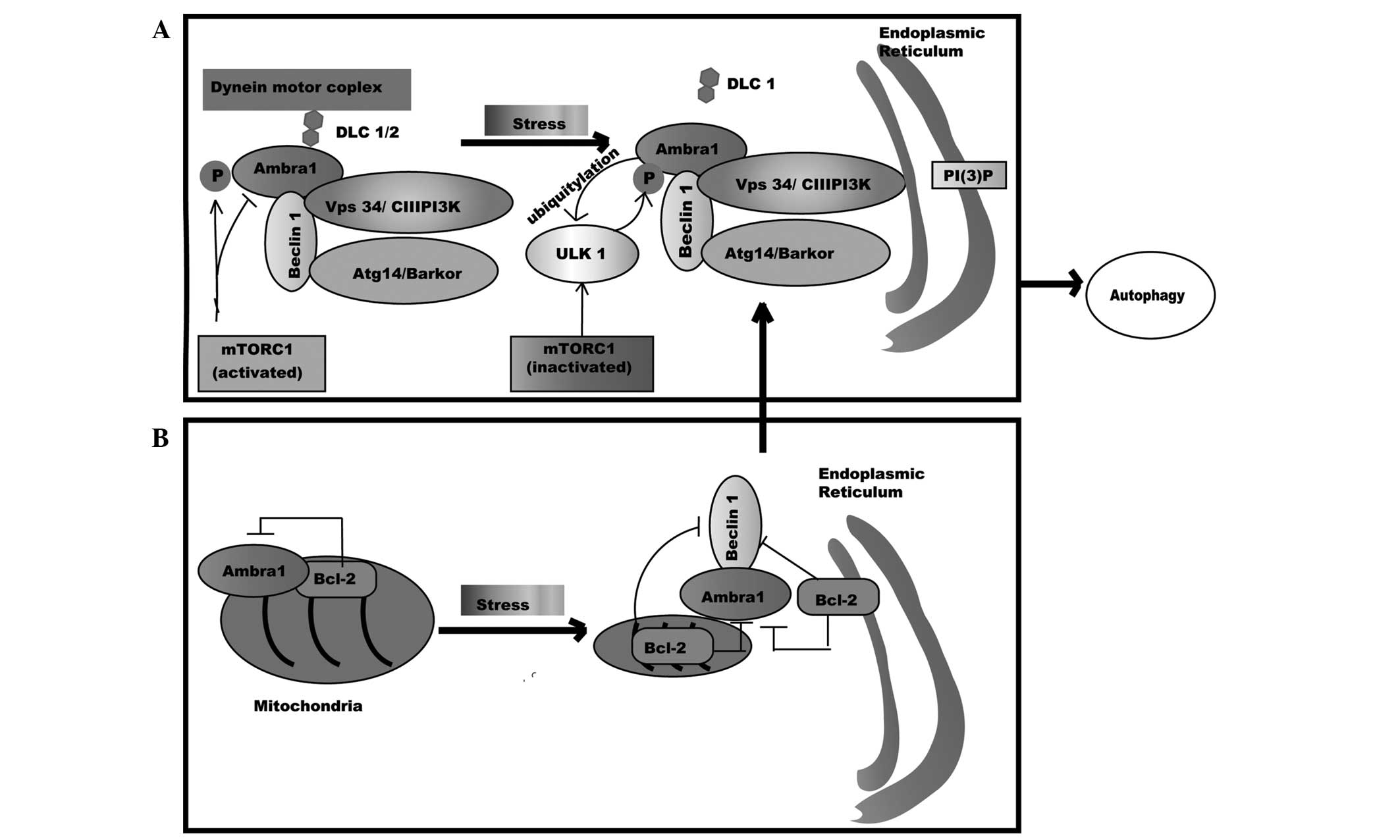 | Figure 2.Ambra1 in autophagy. (A) Ambra1
physically interacts with mTORC1, deactivating mTORC1 by
phosphorylation; however, the association between Ambra1 and the
Beclin1-Vps34 complex is deactivated by binding to the dynein motor
complex through interplay with DLC 1/2. While under stress
settings, Ambra1 is activated through phosphorylation by ULK1, the
association of Ambra1-DLC1 and the Beclin1-Vps34 complex is
released from the dynein motor complex, and the core complex
translocates to the ER and accelerates the formation of PI(3)P, which leads to autophagosome formation.
(B) Ambra1 is docked at the mitochondria by Bcl-2. Under stress
conditions, Ambra1 is separated from mito-Bcl-2 and relocated to
the outer membrane of mitochondria. Ambra1 then competes with mito-
and ER-Bcl-2 to bind Beclin1 and prompts Beclin 1-dependent
autophagy. Ambra1, activating molecule in Beclin 1-regulated
autophagy protein 1; mTORC1, mammalian target of rapamycin complex
1; ULK1, Unc-51 like kinase-1; Vps34/CIII PI3K, class III
phosphatidylinositol 3-kinase; ATG14/Barkor, autophagy-related gene
14; DLC, dynein light chain; PI(3)P,
phosphatidylinositol-3-phosphate; ER, endoplasmic reticulum; Bcl-2,
B cell lymphoma-2. |
As a binding protein of Beclin 1, Ambra1 can
directly interact with it at the CCD domain; equally, Beclin 1 can
bind to Ambra1 at the central region. The two proteins are the
primary elements of the Vps34 complex. In addition, Ambra1 can
modify the function of Beclin 1 through regulation the
ubiquitylation of it at lysine 437 (68). The Ambra1-damage specific DNA binding
protein 1-Cullin-4A complex is an E3 ligase for K63-linked
ubiquitylation of Beclin 1 (70,71). The
ubiquitination of Beclin 1 enhances the association of it with
Vps34 and promotes the activation of Vps34, which is required for
starvation-induced autophagy (70).
Furthermore, studies have shown that the downregulation of Ambra1
leads to a reduced capability of Beclin 1 to interact with Vps34,
and a decrease in Vps34 activation (20,72).
Therefore, Ambra1 triggers autophagy through interaction with
Beclin 1 to activate Vps34 kinase and to promote Vps34 complex
formation at the beginning stages of autophagy.
Ambra1 has been found to dynamically bind DLC 1/2,
which leads the Ambra1-Beclin 1-Vps34 complex to tether to the
dynein motor complex, resulting in core complex deactivation
(2,66,68).
During autophagy, Ambra1 is phosphorylated by ULK1 and the
Ambra1-DLC1 and Beclin 1-Vps34 complexes are released from the
dynein motor complex. Subsequently, the core complex translocates
to the ER and initiates autophagic cascades (Fig. 2A). In addition, Ambra1 can also
regulate autophagy through dynamic combination with mito-Bcl-2
(68). Usually, a pool of Ambra1
proteins is docked at the mitochondria by Bcl-2; however, these
proteins remain separate from Bcl-2 and relocate on the outer
membrane of the mitochondria when autophagy occurs (68). Ambra1 then competes with mito-Bcl2 and
Bcl-2 resided at the endoplasmic reticulum to bind Beclin 1, thus
prompting Beclin 1-dependent autophagy (Fig. 2B). Previously, Ambra1-Bcl2 interaction
has been found to decrease at the mitochondria, whereas
Ambra1-Beclin 1 interaction increases at the mitochondria and ER
following the initiation of autophagy (68). Notably, the Ambra1-Bcl-2 interaction
at the mitochondria is disrupted by autophagy and apoptosis
induction (68). As a result, the
dynamic subcellular localization of Ambra1 is also an important
factor of autophagy regulation.
In summary, Ambra1 is a pro-autophagy factor, and
the function of Ambra1 in autophagy induction is a complex process
that requires additional studies.
Ambra1 is a negative factor of apoptosis
execution
Ambra1 has been found to be important for apoptosis
execution (66). As previously
mentioned, the functional deficiency of Ambra1 in mouse embryos
leads to excessive apoptotic cell death (3). Similarly, the downregulation of Ambra1
in adult neural stem cells results in an increase in basal
apoptosis and an augmented sensitivity to DNA-damage-induced death
(73). In addition, in 2FTG cells and
CRC SW620 cells, the downregulation of Ambra1 with siRNA results in
increased sensitivity of the cells to staurosporine- and
etoposide-induced apoptosis, while the overexpression of Ambra1
makes the cells undergo autophagy and survival more easily
(4). Therefore, the expression of
Ambra1 is negatively associated with apoptosis. Pagliarini et
al (74) have found that Ambra1
is rapidly degraded by caspases and calpains when apoptosis is
induced by staurosporine in human fibroblast 2FTGH cells. The
phenomenon has also been observed in SW620 cells during
etoposide-induced apoptosis (4).
Previously, caspases have been found to be responsible for Ambra1
cleavage at the D482 site, whereas calpains are involved in
complete Ambra1 degradation (74). In
addition, caspase-uncleavable Ambra1D482A mutant 2FTGH
cells confer increased resistance to staurosporine- and
etoposide-induced cell death (74).
Thus, Ambra1 is an important target of apoptotic proteases
resulting in the dismantling of the autophagic machinery.
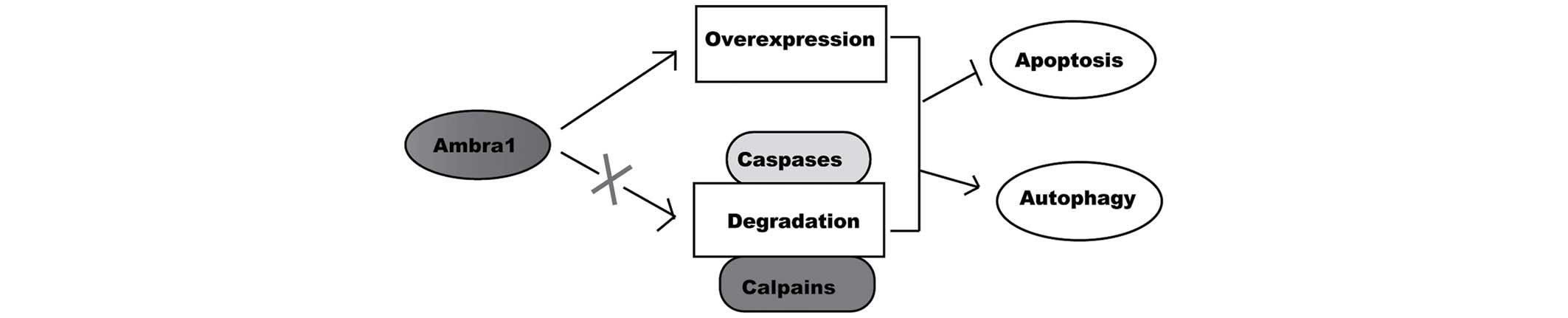
Therefore, the overexpression or damaged degradation of Ambra1
leads to its accumulation, and makes the cells avoid apoptosis and
more easily undergo autophagy and survival (Fig. 3). Furthermore, Ambra1 preferentially
binds the pool of mito-Bcl-2 proteins, and the Ambra1-Bcl-2
interaction is disrupted by the induction of apoptosis and
autophagy, which indicates that Ambra1 has a double function in the
regulation of autophagy and apoptosis (68). During autophagy induction, mito-Bcl-2
separates from Ambra1 leading to the increased release of Bcl-2,
which may enhance the anti-apoptotic function of Bcl-2 (69).
Therefore, Ambra1 is an important factor of
apoptosis execution, and the level at which it is expressed will
determine whether the cell undergoes autophagy or apoptosis.
Ambra1 in cell survival and implications for
chemotherapy resistance
Previously, studies have indicated that there is a
complex association between autophagy and apoptosis (13,75). For
example, the important apoptotic proteins, including Bcl-2 family
members and caspases, participate in the regulation of autophagy,
whereas numerous autophagic proteins, including Beclin 1, Ambra1,
ATG5 and ATG12, are involved in apoptosis execution (75). Therefore, these two processes can
mutually regulate and transform, depending on the context. A study
by Hou et al (76)
demonstrated the autophagic degradation of active caspase-8 during
tumor necrosis factor superfamily member 10-induced autophagy,
indicating that only one of these two processes can prevail at a
time.
As has been previously shown, Ambra1 is an important
factor at the crossroad between autophagy and apoptosis, which may
control the balance and conversion between autophagy and apoptosis.
Ambra1 may primarily play a pro-survival role due to the positive
induction of autophagy, which has been previously demonstrated in
in vivo (mouse embryos) and in vitro (2FTG cells,
SW620 cells and neural stem cells) studies (3,4,73,74).
According to these results, the accumulation of Ambra1 in cells
will promote autophagy occurrence and suppress apoptosis execution,
which may decrease the effectiveness of chemotherapy. Therefore,
Ambra1 may be a negative factor in cancer treatment and prognosis,
and involved in drug resistance. Previously, it has been reported
that Ambra1 is expressed in ~63.9% patients with pancreatic ductal
adenocarcinoma (77). Furthermore,
the increased expression of Ambra1 is marginally associated with
perineural invasion (P=0.063), which is a negative prognostic
factor of cancer therapy, and significantly associated with poor
overall survival (P=0.032) (77).
Nevertheless, no more data currently exists on the expression of
Ambra1 in other human cancers and its role in chemotherapy
resistance. In addition, the intracellular distribution and
interplay of Ambra1 with other proteins, such as Bcl-2, can also
modify the function of Ambra1 and control the conversion between
autophagy and apoptosis to determine cell death or survival
(68). Therefore, Ambra1 is an
important factor in deciding the fate of cells. Increased levels of
Ambra1 due to overexpression or damaged degradation, in addition to
abnormal distribution and interaction with other proteins, may
shift the balance towards autophagy instead of apoptosis in order
to help the cells to survive, thus leading to the resistance to
anticancer drugs.
Conclusion
It is not surprising that autophagy is involved in
the resistance of cancer to anticancer drugs, due to its dual role
in cell death and survival. However, the current knowledge of the
molecular mechanisms of autophagy in drug resistance remains
superficial. Ambra1 is an important factor in regards to the
association between autophagy and apoptosis, and for switching
between these two processes. The accumulation of Ambra1 promotes
autophagy occurrence, protecting cells from apoptosis. In addition,
the abnormal distribution and interplay of Ambra1 with other
proteins can also modify its function. Therefore, Ambra1 may be a
novel target for cancer treatment, and involved in chemotherapy
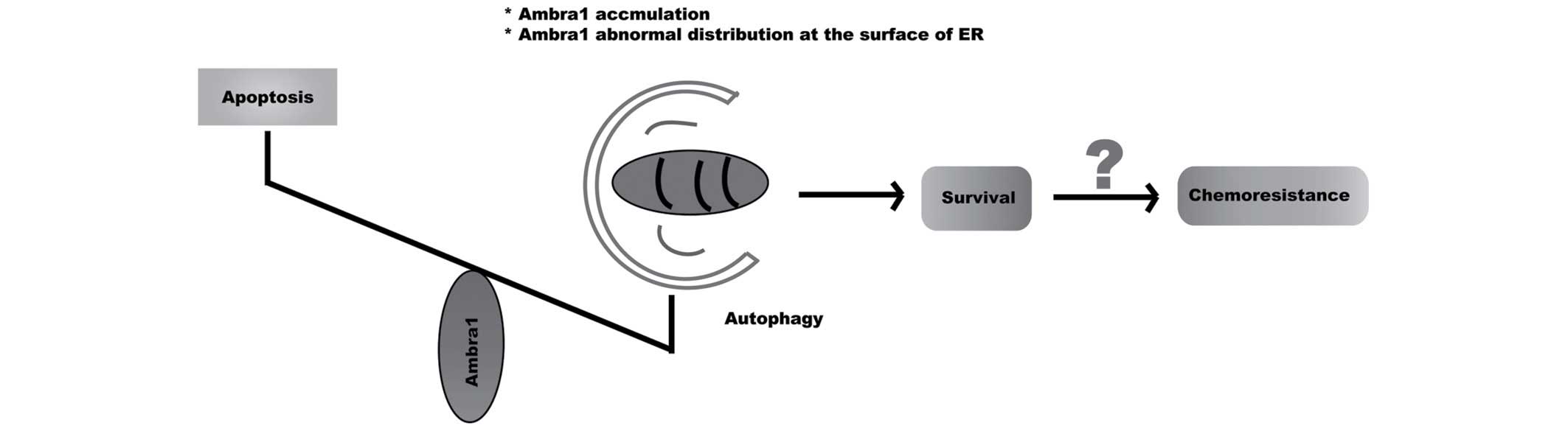
resistance. Based on these findings, it is possible that, in
resistant cells, Ambra1 demonstrates accumulation or abnormal
distribution at the surface of the ER, which tips the balance
towards autophagy and accelerates cell survival, thus decreasing
the effectiveness of chemotherapy and generating resistance to the
drugs (Fig. 4). In the future,
rigorous studies in vivo and in vitro are required to
confirm this theory and to investigate the potential mechanism.
Furthermore, additional studies may be helpful for understanding
the mechanism of drugs resistance, and supply novel strategies for
cancer treatment, particularly for overcoming chemotherapy
resistance in a clinical setting.
Acknowledgements
The present study is supported by grants from the
National Natural Science Foundation of China (Beijing, China; grant
no. 81360340) and the Wu Jieping Medical Foundation clinical
research special fund (Beijing, China; grant no.
320.6750.12689).
Glossary
Abbreviations
Abbreviations:
|
Ambra1
|
activating molecule in Beclin
1-regulated autophagy protein 1
|
|
ATG
|
autophagy-related gene
|
|
mTOR
|
mammalian target of rapamycin
|
|
ULK1
|
Unc-51 like kinase-1
|
|
ER
|
endoplasmic reticulum
|
|
DLC1/2
|
dynein light chain 1/2
|
|
Vps34
|
vacuolar protein-sorting protein
34
|
|
Mito-Bcl-2
|
Bcl-2 located at the mitochondria
|
|
ER-Bcl-2
|
Bcl-2 resided at the endoplasmic
reticulum
|
References
|
1
|
Sui X, Chen R, Wang Z, Huang Z, Kong N,
Zhang M, Han W, Lou F, Yang J, Zhang Q, et al: Autophagy and
chemotherapy resistance: A promising therapeutic target for cancer
treatment. Cell Death Dis. 4:e8382013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Fimia GM, Corazzari M, Antonioli M and
Piacentini M: Ambra1 at the crossroad between autophagy and cell
death. Oncogene. 32:3311–3318. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Fimia GM, Stoykova A, Romagnoli A, Giunta
L, Di Bartolomeo S, Nardacci R, Corazzari M, Fuoco C, Ucar A,
Schwartz P, et al: Ambra1 regulates autophagy and development of
the nervous system. Nature. 447:1121–1125. 2007.PubMed/NCBI
|
|
4
|
Gu W, Wan D, Qian Q, Yi B, He Z, Gu Y,
Wang L and He S: Ambra1 is an essential regulator of autophagy and
apoptosis in SW620 cells: Pro-survival role of Ambra1. PLoS One.
9:e901512014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Klionsky DJ: Autophagy: From phenomenology
to molecular understanding in less than a decade. Nat Rev Mol Cell
Biol. 8:931–937. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kroemer G and Jäättelä M: Lysosomes and
autophagy in cell death control. Nat Rev Cancer. 5:886–897. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kondo Y, Kanzawa T, Sawaya R and Kondo S:
The role of autophagy in cancer development and response to
therapy. Nat Rev Cancer. 5:726–734. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Xie ZP and Klionsky DJ: Autophagosome
formation: Core machinery and adaptation. Nat Cell Biol.
9:1102–1109. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Levine B: Cell biology: Autophagy and
cancer. Nature. 446:745–747. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ravikumar B, Futter M, Jahreiss L,
Korolchuk VI, Lichtenberg M, Luo S, Massey DC, Menzies FM,
Narayanan U, Renna M, et al: Mammalian macroautophagy at a glance.
J Cell Sci. 122:1707–1711. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Baehrecke EH: Autophagy: Dual roles in
life and death? Nat Rev Mol Cell Biol. 6:505–510. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Levine B and Yuan J: Autophagy in cell
death: An innocent convict? J Clin Invest. 115:2679–2688. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Maiuri MC, Zalckvar E, Kimchi A and
Kroemer G: Self-eating and self-killing: Crosstalk between
autophagy and apoptosis. Nat Rev Mol Cell Biol. 8:741–752. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Mizushima N, Yoshimori T and Ohsumi Y: The
role of Atg proteins in autophagosome formation. Annu Rev Cell Dev
Biol. 27:107–132. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Rubinsztein DC, Shpilka T and Elazar Z:
Mechanisms of autophagosome biogenesis. Curr Biol. 22:R29–R34.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Dunlop EA and Tee AR: mTOR and autophagy:
A dynamic relationship governed by nutrients and energy. Semin Cell
Dev Biol. 36:121–129. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wong PM, Puente C, Ganley IG and Jiang XJ:
The ULK1 complex: Sensing nutrient signals for autophagy
activation. Autophagy. 9:124–137. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kang R, Zeh HJ, Lotze MT and Tang D: The
Beclin 1 network regulates autophagy and apoptosis. Cell Death
Differ. 18:571–580. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lorin S, Hamaїb A, Mehrpour M and Codogno
P: Autophagy regulation and its role in cancer. Semin Cancer Biol.
23:361–379. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Funderburk SF, Wang QJ and Yue Z: The
Beclin 1-VPS34 complex-at the crossroads of autophagy and beyond.
Trends Cell Biol. 20:355–362. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liang XH, Jackson S, Seaman M, Brown K,
Kempkes B, Hibshoosh H and Levine B: Induction of autophagy and
inhibition of tumorigenesis by beclin 1. Nature. 402:672–676. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Maiuri MC, Criollo A, Tasdemir E, Vicencio
JM, Tajeddine N, Hickman JA, Geneste O and Kroemer G: BH3-only
proteins and BH3 mimetics induce autophagy by competitively
disrupting the interaction between Beclin 1 and Bcl-2/Bcl-X (L).
Autophagy. 3:374–376. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sinha S and Levine B: The autophagy
effector Beclin 1: A novel BH3-only protein. Oncogene. 27(Suppl 1):
S137–S148. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fu LL, Cheng Y and Liu B: Beclin-1:
Autophagic regulator and therapeutic target in cancer. Int J
Biochem Cell Biol. 45:921–924. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Toton E, Lisiak N, Sawicka P and
Rybczynska M: Beclin-1 and its role as a target for anticancer
therapy. J Physiol Pharmacol. 65:459–467. 2014.PubMed/NCBI
|
|
26
|
He C and Levine B: The Beclin 1
interactome. Curr Opin Cell Biol. 22:140–149. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Matsunaga K, Saitoh T, Tabata K, Omori H,
Satoh T, Kurotori N, Maejima I, Shirahama-Noda K, Ichimura T, Isobe
T, et al: Two Beclin 1-binding proteins, Atg14L and Rubicon,
reciprocally regulate autophagy at different stages. Nat Cell Biol.
11:385–396. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Itakura E, Kishi C, Inoue K and Mizushima
N: Beclin 1 forms two distinct phosphatidylinositol 3-kinase
complexes with mammalian Atg14 and UVRAG. Mol Biol Cell.
19:5360–5372. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Choi KS: Autophagy and cancer. Exp Mol
Med. 44:109–120. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Corazzari M, Fimia GM and Piacentini M:
Dismantling the autophagic arsenal when it is time to die:
Concerted AMBRA1 degradation by caspases and calpains. Autophagy.
8:1255–1257. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Shingu T, Fujiwara K, Bögler O, Akiyama Y,
Moritake K, Shinojima N, Yokoyama T and Kondo S: Inhibition of
autophagy at a late stage enhances imatinib-induced cytotoxicity in
human malignant glioma cells. Int J Cancer. 124:1060–1071. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Xiong HY, Guo XL, Bu XX, Zhang SS, Ma NN,
Song JR, Hu F, Tao SF, Sun K, Li R, et al: Autophagic cell death
induced by 5-FU in Bax or PUMA deficient human colon cancer cell.
Cancer Lett. 288:68–74. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lee YJ, Won AJ, Lee J, Jung JH, Yoon S,
Lee BM and Kim HS: Molecular mechanism of SAHA on regulation of
autophagic cell death in tamoxifen-resistant MCF-7 breast cancer
cells. Int J Med Sci. 9:881–893. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tai WT, Shiau CW, Chen HL, Liu CY, Lin CS,
Cheng AL, Chen PJ and Chen KF: Mcl-1-dependent activation of Beclin
1 mediates autophagic cell death induced by sorafenib and SC-59 in
hepatocellular carcinoma cells. Cell Death Dis. 4:e4852013.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Li J, Hou N, Faried A, Tsutsumi S and
Kuwano H: Inhibition of autophagy augments 5-fluorouracil
chemotherapy in human colon cancer in vitro and in vivo model. Eur
J Cancer. 46:1900–1909. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
de la Cruz-Morcillo MA, Valero ML,
Callejas-Valera JL, Arias-González L, Melgar-Rojas P, Galán-Moya
EM, García-Gil E, García-Cano J and Sánchez-Prieto R: P38MAPK is a
major determinant of the balance between apoptosis and autophagy
triggered by 5-fluorouracil: Implication in resistance. Oncogene.
31:1073–1085. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Sasaki K, Tsuno NH, Sunami E, Tsurita G,
Kawai K, Okaji Y, Nishikawa T, Shuno Y, Hongo K, Hiyoshi M, et al:
Chloroquine potentiates the anti-cancer effect of 5-fluorouracil on
colon cancer cells. BMC Cancer. 10:3702010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Sasaki K, Tsuno NH, Sunami E, Kawai K,
Hongo K, Hiyoshi M, Kaneko M, Murono K, Tada N, Nirei T, et al:
Resistance of colon cancer to 5-fluorouracil may be overcome by
combination with chloroquine, an in vivo study. Anticancer Drugs.
23:675–682. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yang PM, Liu YL, Lin YC, Shun CT, Wu MS
and Chen CC: Inhibition of autophagy enhances anticancer effects of
atorvastatin in digestive malignancies. Cancer Res. 70:7699–7709.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Paillas S, Causse A, Marzi L, de Medina P,
Poirot M, Denis V, Vezzio-Vie N, Espert L, Arzouk H, Coquelle A, et
al: MAPK14/p38α confers irinotecan resistance to TP53-defective
cells by inducing survival autophagy. Autophagy. 8:1098–1112. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Liu D, Yang Y, Liu Q and Wang J:
Inhibition of autophagy by 3-MA potentiates cisplatin-induced
apoptosis in esophageal squamous cell carcinoma cells. Med Oncol.
28:105–111. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Ding ZB, Hui B, Shi YH, Zhou J, Peng YF,
Gu CY, Yang H, Shi GM, Ke AW, Wang XY, et al: Autophagy activation
in hepatocellular carcinoma contributes to the tolerance of
oxaliplatin via reactive oxygen species modulation. Clin Cancer
Res. 17:6229–6238. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Guo XL, Li D, Sun K, Wang J, Liu Y, Song
JR, Zhao QD, Zhang SS, Deng WJ, Zhao X, et al: Inhibition of
autophagy enhances anticancer effects of bevacizumab in
hepatocarcinoma. J Mol Med (Berl). 91:473–483. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Shi YH, Ding ZB, Zhou J, Hui B, Shi GM, Ke
AW, Wang XY, Dai Z, Peng YF, Gu CY, et al: Targeting autophagy
enhances sorafenib lethality for hepatocellular carcinoma via ER
stress-related apoptosis. Autophagy. 7:1159–1172. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zhao M, Yang M, Yang L, Yu Y, Xie M, Zhu
S, Kang R, Tang D, Jiang Z, Yuan W, et al: HMGB1 regulates
autophagy through increasing transcriptional activities of JNK and
ERK in human myeloid leukemia cells. BMB Rep. 44:601–606. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Han W, Pan H, Chen Y, Sun J, Wang Y, Li J,
Ge W, Feng L, Lin X, Wang X, et al: EGFR tyrosine kinase inhibitors
activate autophagy as a cytoprotective response in human lung
cancer cells. PLoS One. 6:e186912011. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Wang Y, Peng RQ, Li DD, Ding Y, Wu XQ,
Zeng YX, Zhu XF and Zhang XS: Chloroquine enhances the cytotoxicity
of topotecan by inhibiting autophagy in lung cancer cells. Chin J
Cancer. 30:690–700. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Kang R, Tang D, Schapiro NE, Livesey KM,
Farkas A, Loughran P, Bierhaus A, Lotze MT and Zeh HJ: The receptor
for advanced glycation end products (RAGE) sustains autophagy and
limits apoptosis, promoting pancreatic tumor cell survival. Cell
Death Differ. 17:666–676. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Shin SW, Kim SY and Park JW: Autophagy
inhibition enhances ursolic acid-induced apoptosis in PC3 cells.
Biochim Biophys Acta. 1823:451–457. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Schoenlein PV, Periyasamy-Thandavan S,
Samaddar JS, Jackson WH and Barrett JT: Autophagy facilitates the
progression of ERalpha-positive breast cancer cells to antiestrogen
resistance. Autophagy. 5:400–403. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
O'Donovan TR, O'Sullivan GC and McKenna
SL: Induction of autophagy by drug-resistant esophageal cancer
cells promotes their survival and recovery following treatment with
chemotherapeutics. Autophagy. 7:509–524. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Chen YS, Song HX, Lu Y, Li X, Chen T,
Zhang Y, Xue JX, Liu H, Kan B, Yang G and Fu T: Autophagy
inhibition contributes to radiation sensitization of esophageal
squamous carcinoma cells. Dis Esophagus. 24:437–443. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Xu CX, Zhao L, Yue P, Fang G, Tao H,
Owonikoko TK, Ramalingam SS, Khuri FR and Sun SY: Augmentation of
NVP-BEZ235′s anticancer activity against human lung cancer cells by
blockage of autophagy. Cancer Biol Ther. 12:549–555. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Mirzoeva OK, Hann B, Hom YK, Debnath J,
Aftab D, Shokat K and Korn WM: Autophagy suppression promotes
apoptotic cell death in response to inhibition of the PI3K-mTOR
pathway in pancreatic adenocarcinoma. J Mol Med (Berl). 89:877–889.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Xu WH, Liu ZB, Hou YF, Hong Q, Hu DL and
Shao ZM: Inhibition of autophagy enhances the cytotoxic effect of
PA-MSHA in breast cancer. BMC Cancer. 14:2732014. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Fedorko M: Effect of chloroquine on
morphology of cytoplasmic granules in maturing human leukocytes-an
ultrastructural study. J Clin Invest. 46:1932–1942. 1967.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Klionsky DJ, Baehrecke EH, Brume JH, Chu
CT, Codogno P, Cuervo AM, Debnath J, Deretic V, Elazar Z, Eskelinen
EL, et al: A comprehensive glossary of autophagy-related molecules
and processes (II edition). Autophagy. 7:1273–1294. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Gottesman MM: Mechanisms of cancer drug
resistance. Annu Rev Med. 53:615–627. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Szakács G, Paterson JK, Ludwig JA,
Booth-Genthe C and Gottesman MM: Targeting multidrug resistance in
cancer. Nat Rev Drug Discov. 5:219–234. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Sun WL, Chen J, Wang YP and Zheng H:
Autophagy protects breast cancer cells from epirubicin-induced
apoptosis and facilitates epirubicin-resistance development.
Autophagy. 7:1035–1044. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Sun WL, Lan D, Gan TQ and Cai ZW:
Autophagy facilitates multidrug resistance development through
inhibition of apoptosis in breast cancer cells. Neoplasma.
62:199–208. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Chittaranjan S, Bortnik S, Dragowska WH,
Xu J, Abeysundara N, Leung A, Go NE, DeVorkin L, Weppler SA, Gelmon
K, et al: Autophagy inhibition augments the anticancer effects of
epirubicin treatment in anthracycline-sensitive and -resistant
triple-negative breast cancer. Clin Cancer Res. 20:3159–3173. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Samaddar JS, Gaddy VT, Duplantier J,
Thandavan SP, Shah M, Smith MJ, Browning D, Rawson J, Smith SB,
Barrett JT and Schoenlein PV: A role for macroautophagy in
protection against 4-hydroxytamoxifen-induced cell death and the
development of antiestrogen resistance. Mol Cancer Ther.
7:2977–2987. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Vazquez-Martin A, Oliveras-Ferraros C and
Menendez JA: Autophagy facilitates the development of breast cancer
resistance to the anti-HER2 monoclonal antibody trastuzumab. PLoS
ONE. 4:e62512009. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Ajabnoor GM, Crook T and Coley HM:
Paclitaxel resistance is associated with switch from apoptotic to
autophagic cell death in MCF-7 breast cancer cells. Cell Death Dis.
3:e2602012. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Di Bartolomeo S, Corazzari M, Nazio F,
Oliverio S, Lisi G, Antonioli M, Pagliarini V, Matteoni S, Fuoco C,
Giunta L, et al: The dynamic interaction of AMBRA1 with the dynein
motor complex regulates mammalian autophagy. J Cell Biol.
191:155–168. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Nazio F, Strappazzon F, Antonioli M,
Bielli P, Cianfanelli V, Bordi M, Gretzmeier C, Dengjel J,
Piacentini M, Fimia GM and Cecconi F: mTOR inhibits autophagy by
controlling ULK1 ubiquitylation, self-association and function
through AMBRA1 and TRAF6. Nat Cell Bio. 15:406–416. 2013.
View Article : Google Scholar
|
|
68
|
Strappazzon F, Vietri-Rudan M, Campello S,
Nazio F, Florenzano F, Fimia GM, Piacentini M, Levine B and Cecconi
F: Mitochondrial BCL-2 inhibits AMBRA1-induced autophagy. EMBO J.
30:1195–1208. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Tooze1 SA and Codogno P: Compartmentalized
regulation of autophagy regulators: Fine-tuning AMBRA1 by Bcl-2.
EMBO J. 30:1185–1186. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Xia P, Wang S, Du Y, Zhao Z, Shi L, Sun L,
Huang G, Ye B, Li C, Dai Z, et al: WASH inhibits autophagy through
suppression of Beclin 1 ubiquitination. EMBO J. 32:2685–2696. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Xia P, Wang S, Huang G, Du Y, Zhu P, Li M
and Fan Z: RNF2 is recruited by WASH to ubiquitinate AMBRA1 leading
to downregulation of autophagy. Cell Res. 24:943–958. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Abrahamsen H, Stenmark H and Platta HW:
Ubiquitination and phosphorylation of Beclin 1 and its binding
partners: Tuning class III phosphatidylinositol 3-kinase activity
and tumor suppression. FEBS Lett. 586:1584–1591. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Yazdankhah M, Farioli-Vecchioli S, Tonchev
AB, Stoykova A and Cecconi F: The autophagy regulators Ambra1 and
Beclin 1 are required for adult neurogenesis in the brain
subventricular zone. Cell Death Dis. 5:e14032014. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Pagliarini V, Wirawan E, Romagnoli A,
Ciccosanti F, Lisi G, Lippens S, Cecconi F, Fimia GM, Vandenabeele
P, Corazzari M and Piacentini M: Proteolysis of Ambra1 during
apoptosis has a role in the inhibition of the autophagic
pro-survival response. Cell Death Differ. 19:1495–1504. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Booth LA, Tavallai S, Hamed HA,
Cruickshanks N and Dent P: The role of cell signalling in the
crosstalk between autophagy and apoptosis. Cell Signal. 26:549–555.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Hou W, Han J, Lu C, Goldstein LA and
Rabinowich H: Autophagic degradation of active caspase-8: A
crosstalk mechanism between autophagy and apoptosis. Autophagy.
6:891–900. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Ko YH, Cho YS, Won HS, Jeon EK, An HJ,
Hong SU, Park JH and Lee MA: Prognostic significance of
autophagy-related protein expression in resected pancreatic ductal
adenocarcinoma. Pancreas. 42:829–835. 2013. View Article : Google Scholar : PubMed/NCBI
|