Introduction
Oral squamous cell carcinoma (OSCC) is one of the
most common malignancies of the head and neck. It is characterized
by local invasiveness and a high rate of cervical lymph node
metastasis (1,2). In particular, lymph node metastasis has
been revealed to directly affect the prognosis of patients
(3). Despite advances in combined
therapies, including radical surgery, radiotherapy and neo-adjuvant
chemotherapy, the poor prognosis observed for patients with OSCC
has not significantly improved during the last decade (4,5).
Therefore, investigation to better understand the mechanism of
metastasis in OSCC is required, in order to provide a more
effective therapeutic strategy.
It has been previously demonstrated that
epithelial-mesenchymal transition (EMT) is linked with OSCC
progression, including cell invasion and metastasis (6,7). Tumor
cells undergoing EMT aid in the dissemination of carcinoma cells,
in which stationary epithelial tumor cells typically lose an
epithelial phenotype, including cell polarity and adherent
junctions, and acquire a narrow, fibroblast-like mesenchymal
morphology and invasive properties. In addition, these cells
exhibit a decrease in epithelial markers, including E-cadherin and
β-catenin, and an increase in mesenchymal markers, including
N-cadherin and vimentin (8,9). These alterations to the cell phenotype
facilitates the cells in losing their adhesiveness, dissolving the
extracellular matrix, which depends on matrix metalloproteinases
(MMPs), and spreading to the surrounding tissue (10). However, little information is
available regarding the type of EMT induced by chemokines.
The chemokine receptor C-X-C chemokine receptor type
4 (CXCR4), a G-protein-coupled receptor that is widely expressed on
the membranes of neutrophils, lymphocytes and monocytes, and less
often on epithelial cells, is the cognate receptor of stromal
cell-derived factor-1α (SDF-1α; also referred to as CXCL12)
(9,11). Several studies illustrate that SDF-1
and CXCR4 are associated with tumor progression and the development
of lymph node and distant metastasis (12–15). In
addition, CXCR4 expression has been demonstrated to be upregulated
in a variety cancer types, including breast, ovarian, kidney and
esophageal cancer (16–19), and elevated levels of CXCR4 in
patients with cancer have been associated with a poor patient
outcome (20,21).
The present authors have previously reported that an
overexpression of CXCR4 is associated with lymph node metastasis in
OSCC, and is significantly correlated with EMT-associated proteins
(22). Further analysis concerning
the regulation of CXCR4 expression accompanying EMT may be useful
for the treatment of metastatic lymph nodes of OSCC. The present
study used a gene silencing method to evaluate the effect of CXCR4
on EMT in vitro and in vivo in OSCC.
Materials and methods
Cell culture
Tongue squamous cell carcinoma (TSCCA) cells were
purchased from the Chinese Academy of Medical Sciences Institute of
Basic Medical Sciences (Beijing, China) and were grown in minimum
essential medium (MEM; Thermo Fisher Scientific, Inc., Waltham, MA,
USA) supplemented with 10% fetal bovine serum (FBS; Thermo Fisher
Scientific, Inc.). Cell cultures were incubated at 37°C in a 5%
CO2 atmosphere.
Small interfering (si) RNA-mediated
downregulation of CXCR4 expression
The siRNAs used in the present study were chemically
synthesized by GenePharma Co., Ltd. (Shanghai, China). The
sequences were as follows: CXCR4 siRNA sense,
5′-CCGACCUCCUCUUUGUCAUTT-3′; negative control siRNA sense,
5′-UUCUCCGAACGUGUCACGUTT-3′. For transfection, 5×105
cells were seeded into each well of a 6-well plate and grown
overnight until they reached 50–80% confluency. The cells were
washed, placed in opti-MEM® reduced serum medium (Thermo
Fisher Scientific, Inc.) and transfected with siRNA using
Lipofectamine® 2000 (Invitrogen™; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol.
Subsequent to a 6 h transfection, the medium was changed to MEM
(Thermo Fisher Scientific, Inc.), and the cells were cultured at
37°C in 5% CO2. Three groups were generated for all
experiments: Blank control group; negative siRNA control group; and
CXCR4 siRNA group.
Semi-quantitative reverse
transcription-polymerase chain reaction (RT-PCR) analysis
Total RNA was extracted from the TSCCA cells using
TRIzol Reagent (Invitrogen™; Thermo Fisher Scientific, Inc.),
according to the manufacturer's protocol. cDNA was synthesized
using M-MLV Reverse Transcriptase (Invitrogen™; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol. A 25
µl reaction containing 1 µl of the reverse transcription product
was used for semi-quantitative PCR. The RT-qPCR cycling conditions
were as follows: 30 sec at 95°C, followed by 40 cycles of 5 sec at
95°C and 34 sec at 60°C. CXCR4 was amplified using the following
primers: Forward, 5′-CACTGGTGTCGGTCTCTGC-3′; and reverse,
5′-TGATTGAGTCAATGAAGTGGC-3′, which were purchased from Sangon
Biotech Co., Ltd. (Shanghai, China). Human β-actin was used as an
internal control, and the primer sequence was as follows: Forward,
5′-CACAGCAAGAGAGGCATCC-3′; and reverse, 5′-CTGGGGTGTTGAAGGTCTC-3′.
A 2% agarose gel electrophoresis was used to analyze the amplified
products.
Western blotting
Following transfection and culturing for 48 h, the
cells were washed three times with ice-cold phosphate-buffered
saline (PBS) and lysed in RIPA-buffer (EMD Millipore, Billerica,
MA, USA). The cell lysates were cleared by centrifugation at 10,000
× g for 5 min and protein concentrations of the supernatants were
determined using a BCA protein assay kit (Thermo Fisher Scientific,
Inc.). Subsequently, ~30 µg protein was separated by 10% sodium
dodecyl sulfate-polyacrylamide gel electrophoresis. The gel was
transferred onto polyvinylidene difluoride membranes and probed
with the following primary antibodies: Anti-CXCR4 (dilution, 1:200;
#ab58176), anti-N-cadherin (dilution, 1:1,500; #ab98952), anti-MMP2
(dilution, 1:5,000; #ab92536), anti-MMP9 (dilution, 1:800;
#ab137867) (Abcam, Cambridge, MA, USA); and anti-E-cadherin
(dilution, 1:800; #ZS-7870; Zhongshan Golden Bridge Biotechnology
Co., Ltd., Beijing, China). The membranes were incubated with
primary antibodies 4°C overnight, followed by incubation with
horseradish peroxidase-conjugated secondary antibodies (dilution,
1:15,000; #ZB-5301 and #ZB-5303; Zhongshan Golden Bridge
Biotechnology Co., Ltd.) 37°C for 2 h. Immunoreactive protein bands
were visualized using an enhanced chemiluminescence detection
system (Bio-Rad Laboratories, Inc., Hercules, CA, USA) and
subsequent exposure of the membrane to Hyperfilm (Thermo Fisher
Scientific, Inc.). The same membrane was probed with a β-actin
antibody (dilution, 1:2,000; #TA-09; Zhongshan Golden Bridge
Biotechnology Co., Ltd.) as a loading control. Denistometric
analysis was performed using Image J 1.47ver software (National
Institutes of Health, Bethesda, MD, USA).
Wound healing assay
Cells were cultured in 6-well plates and incubated
overnight. Scratches were created by scratching a straight line
with a 20 µl tip vertically in the center of the dish. Cells were
transfected with CXCR4 siRNA. The dishes were washed with PBS once
to remove the detached cells, and a primary image of the scratch
was taken under a microscope. A total of 24 and 48 h following
transfection, the width of the scratches was observed and measured
using Image J v1.47 software (National Institutes of Health,
Bethesda, MA, USA). The relative distance was calculated as the
mean width of the cell scratch following transfection / the width
prior to transfection.
Transwell invasion assays
A cell invasion assay was performed using invasion
chambers (BD Biosciences, Franklin Lakes, NJ, USA). In total, 50 µl
Matrigel (BD Biosciences) and 200 µl cell suspension
(1×105 cells/ml) was added to the inserts. MEM
containing 10% FBS was added to the lower chamber. The cells were
cultured in a CO2 incubator at 37°C for 48 h. The
noninvasive cells in the upper chamber were cleared with cotton
swabs and invasive cells were fixed with 95% ethanol for 10 min and
stained with 0.5% crystal violet. Cells that had penetrated through
the polyethylene terephthalate membrane were counted in 10
representative microscopic fields (magnification, ×400).
Annexin V-fluorescein isothiocyanate
(FITC)/propidum iodide (PI) double-staining
An Annexin V-FITC Apoptosis Detection kit (BD
Biosciences) was used to assess the apoptosis of transfected cells.
TSCCA cells, which were transfected for 48 h, were collected and
washed with PBS. Subsequently, the cells were resuspended in 1X
Binding Buffer at a density of 1×106 cells/ml. In total,
5 µl Annexin V-FITC and 5 µl PI were added to the cells, and they
were incubated at room temperature for 15 min. Subsequently, the
cells were analyzed by flow cytometry, using BD FACSDiva™ software
v6.1.3 and BD CellQuest™ Pro (BD Biosciences).
Nude mouse tumor xenograft model
A total of 18 BALB/c Nude 4-week-old female mice
(weight, 15–18 g) were purchased from the Cancer Institute and
Hospital, Chinese Academy of Medical Sciences (Beijing, China). All
animal experimental protocols were approved by the Tianjin Medical
University Animal Care and Use Committee. All mice were maintained
under specific pathogen-free conditions: Food and water were
sterilized by high-pressure steam and feed was sterilized by Co-60
irradiation, and the light/dark cycle was 12 h, with a temperature
of 24±1°C and relative humidity of 55±5%. In total, 3 mice were
injected subcutaneously with 1×107 TSCCA cells, in a
volume of 200 µl PBS. The mice were monitored daily and all 3 mice
formed tumors subcutaneously. When the tumor size reached ~10 mm in
length, the tumors were surgically removed, cut into 1
mm3 pieces and re-seeded into the inguinal region of 15
mice. Two weeks later, the mice were randomized into three groups
(5 mice/group) to receive treatment: Blank control group; negative
control siRNA-treated group; and CXCR4 siRNA-treated group. A
mixture of 5 µl Lipofectamine 2000 and 15 µl siRNA (20 nmol/l)
mixture was injected into the xenograft tumor model in a multi-site
injection manner. Mice in the blank control group received 20 µl
PBS only. The mice were treated every 4 days and tumors were
measured with a caliper. Tumor volume was calculated as follows:
Tumor volume (mm3) = [tumor length (mm) × tumor width
(mm) × 2] / 2. At the end of the 22-day observation period, the
mice bearing xenograft tumors were sacrificed and the tumor tissues
were removed for formalin fixation and preparation of
paraffin-embedded sections.
Hematoxylin and eosin (H&E) and
immunohistochemistry staining
H&E staining was performed on the
formalin-fixed, paraffin-embedded mice tumor tissues to observe the
pathological alterations of the tumor under a microscope. For
immunohistochemistry, tumor tissue sections were deparaffinized,
rehydrated and incubated with primary antibodies, used previously
in western blotting, overnight at 4°C. The tissues were then
incubated with biotin-labeled secondary antibody (KIT9710; Maxim
Biomedical, Inc., Rockville, MD, USA) for 1 h at room temperature,
incubated with diaminobenzidine (Zhongshan Golden Bridge
Biotechnology Co., Ltd.) and counterstained with hematoxylin.
Slides were dehydrated with various concentrations of alcohol and
soaked in xylene, and then mounted with neutral balsam and
visualized using a light microscope.
Statistical analysis
The data are presented as the mean ± standard
deviation of at least three independent experiments. One-way
analysis of variance was performed using SPSS version 17.0 software
(SPSS, Inc., Chicago, IL, USA). P<0.05 was considered to
indicate a statistically significant difference.
Results
CXCR4 siRNA inhibited the expression
of CXCR4 in TSCCA cells
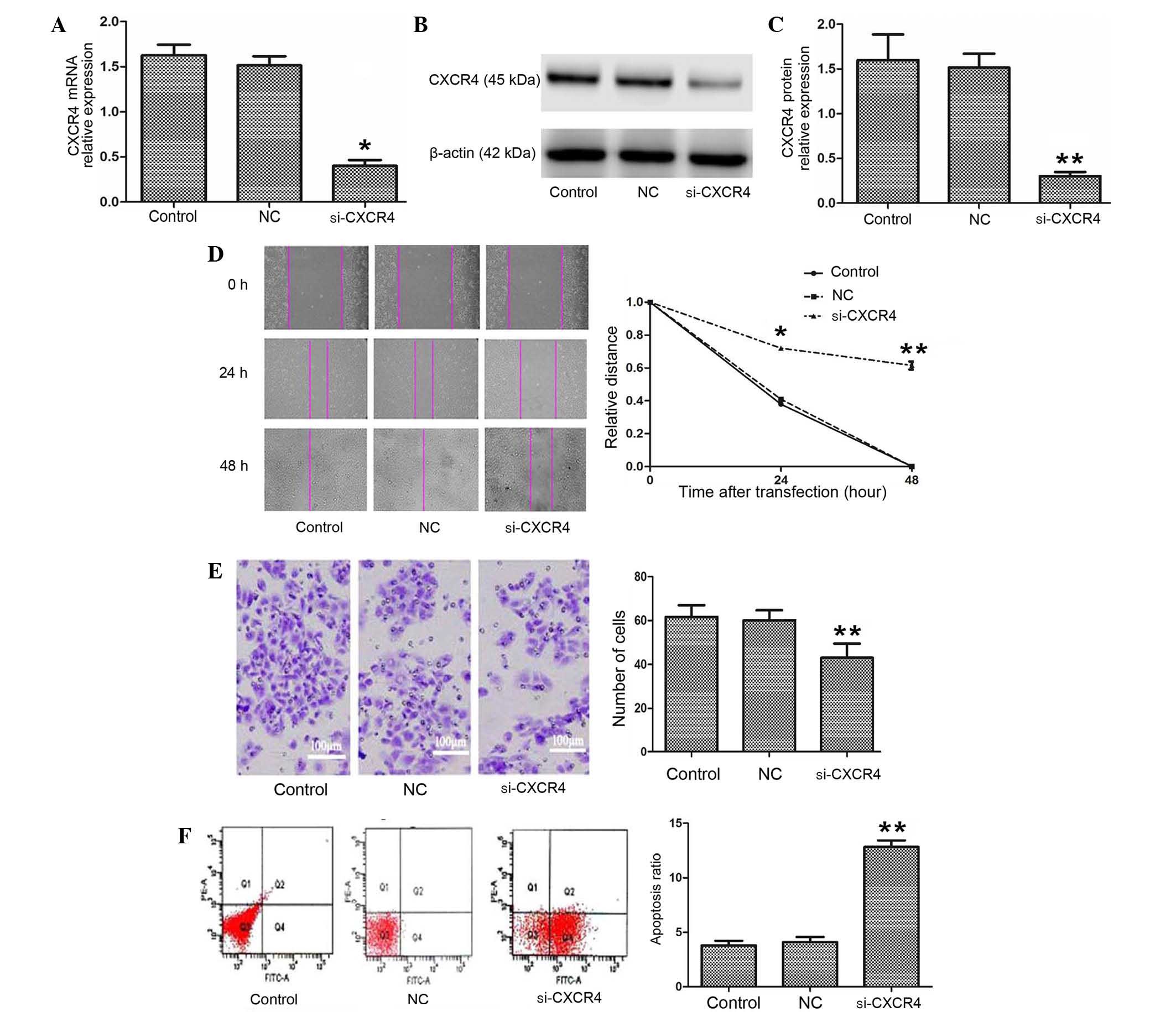
The expression of CXCR4 mRNA and protein were
confirmed using semi-quantitative RT-PCR and western blotting. As
shown in Fig. 1A-C, CXCR4 mRNA and
protein expression in the CXCR4 siRNA group were clearly
downregulated compared with the blank control and negative control
groups (P=0.003). The results indicate that CXCR4 siRNA
significantly silenced the expression of CXCR4 in TSCCA cells.
Silencing of CXCR4 inhibited migration
and invasion and stimulated apoptosis in TSCCA cells
To better understand the biological role of CXCR4,
the present study investigated the motility and invasion ability of
cells using wound healing and Transwell assays. The relative
distance in the wound healing assay was greater in the CXCR4 siRNA
group compared with the blank control and negative control groups
24 h and 48 h following transfection (P=0.036 and P=0.008,
respectively; Fig. 1D). The Transwell
assay demonstrated that the number of invading cells was 61.20±5.71
and 60.50±5.58 in the blank control and negative control groups,
respectively, which was significantly increased compared with the
CXCR4 siRNA group (44.3±8.59; F=19.936; P<0.001; Fig. 1E). Subsequently, the effect of CXCR4
siRNA on cell apoptosis was analyzed using Annexin V and PI double
staining. The Annexin V-positive early-phase apoptotic cells were
significantly increased in the CXCR4 siRNA group (13.23±0.35%)
compared with the blank control and negative control groups
(3.30±0.72 and 3.40±0.44%, respectively; F=351.676; P=0.001 and
P=0.004, respectively; Fig. 1F). The
data suggest that CXCR4 is involved in the progression of OSCC.
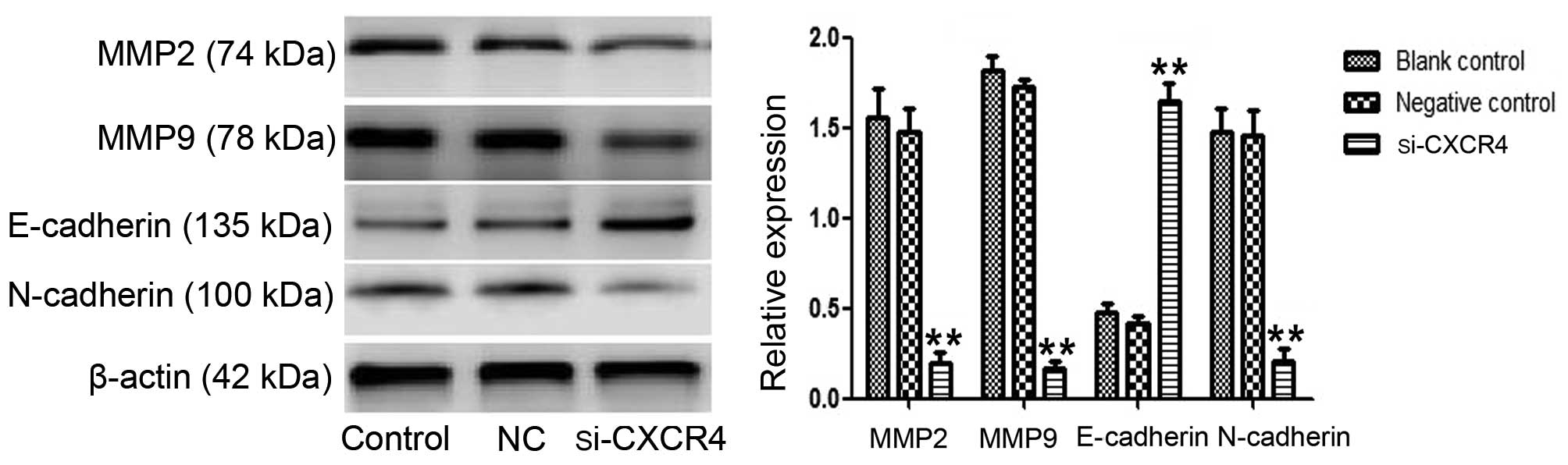
Expression of EMT-associated proteins
were regulated by silencing of CXCR4
To further verify the role of CXCR4 in EMT in TSCCA
cells, the protein expression levels of EMT-associated proteins,
N-cadherin, E-cadherin and MMP2/9 were evaluated by western
blotting. The result revealed that the expression level of the
epithelial marker E-cadherin was elevated (P<0.001), while the
mesenchymal marker N-cadherin was clearly attenuated (P=0.002). In
addition, the expression of MMP2/9, which are assisting factors in
EMT, was inhibited (P<0.001) (Fig.
2). This is consistent with the phenomenon that CXCR4 silencing
suppresses cell invasion. Taken together, these results suggested
that downregulation of CXCR4 facilitates cells to reverse their
mesenchymal characteristic.
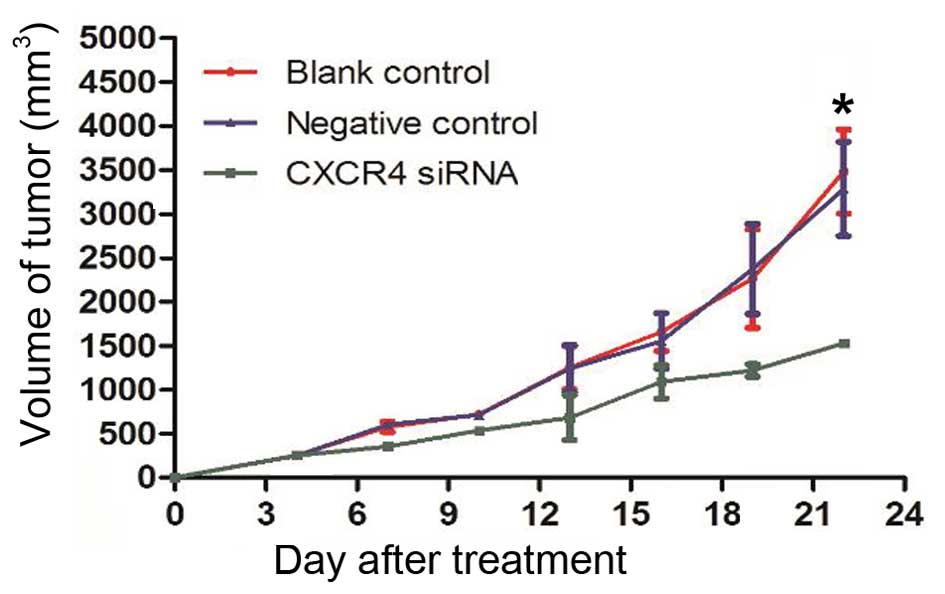
Growth inhibitory effects of CXCR4
siRNA were examined in vivo in a nude mouse model
The results of the present in vitro
experiments suggested that CXCR4 siRNA had an effect on tumor
suppression. To further confirm this conclusion, tumor growth in a
xenograft model was performed using a Lipofectamine-mediated siRNA
therapy approach. At the beginning of treatment, the mean tumor
volumes of the mice in the blank control, negative control and
CXCR4 siRNA groups were 81.55±20.88, 89.63±46.30 and 84.74±18.44
mm3, respectively, with no statistically significant
differences among these three groups. On day 13, a rapid increase
in the tumor volumes in the blank control (1163.90±295.62
mm3) and negative control (1171.30±327.60
mm3) groups was observed compared to the CXCR4 siRNA
group (682.33±429.69 mm3) (P=0.027). This inhibitory
effect was aggravated with increasing time. At the end of the
experiment, a significant decrease in tumor volume was observed in
the CXCR4 siRNA group (1574.90±27.20 mm3) compared with
the blank control and negative control groups (3512.12±870.05
mm3 and 3349.70±920.88 mm3, respectively)
(P=0.012; Fig. 3).
Silencing of CXCR4 suppressed TSCCA
EMT in the xenograft model
The histopathological alterations in the tumor
tissues from the three treatment groups were observed using H&E
staining. Cells in the control and blank groups were sheet
arranged, with large nuclei. These cells exhibited multi
morphology, pathological cell division and giant tumor cells, with
marked atypia. By contrast, the cells in the CXCR4 siRNA group
exhibited smaller nuclei, lighter staining and a reduction in the
amount of neovasculature, and apoptosis and necrosis was observed
(Fig. 4A). Subsequently,
EMT-associated proteins were evaluated in these tissues using
immunohistochemistry. Compared with the blank control and negative
control groups, the expression of CXCR4, N-cadherin and MMP2/9 in
the CXCR4 siRNA group were significantly reduced, and the
expression of E-cadherin was increased (Fig. 4B). These results are consistent with
the conclusion in vitro.
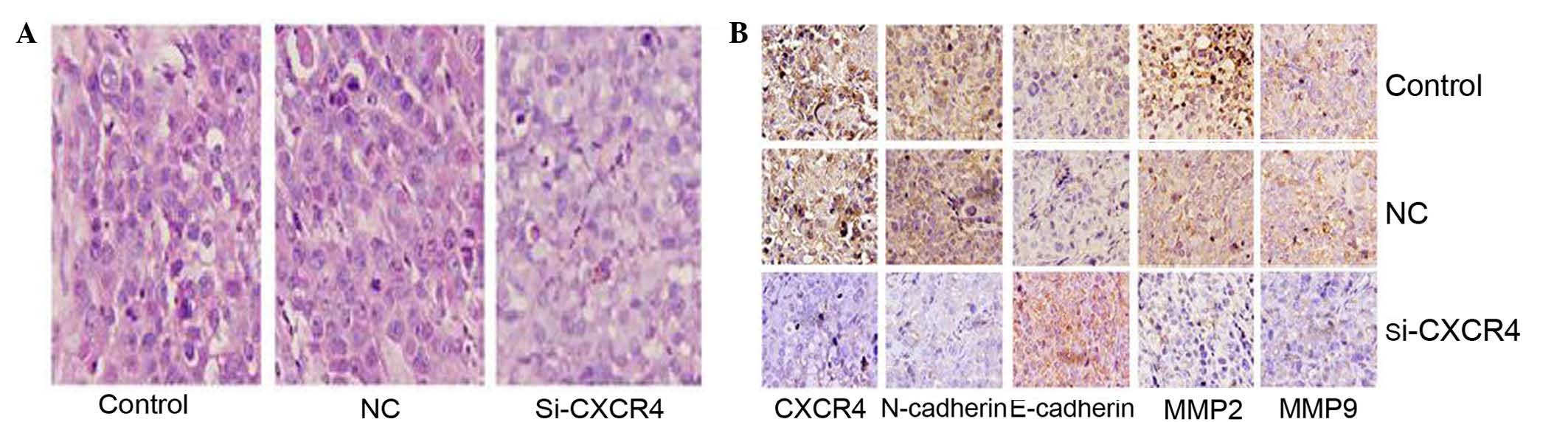 | Figure 4.H&E and immunohistochemistry
analysis of TSCCA xenograft tumors. (A) Histopathological
alterations of tumor tissues from nude mice models were observed
using H&E staining. Compared with the negative control and
blank control groups, cells in the si-CXCR4 group exhibited smaller
nuclei, lighter staining, less neovasculature and more apoptosis
and necrosis. (B) Immunohistochemistry analysis of CXCR4,
N-cadherin, E-cadherin, MMP2/9 in TSCCA xenograft tumors. CXCR4 and
N-cadherin positive staining were observed in the cytoplasm and
membrane, and were suppressed in the si-CXCR4 group. E-cadherin was
located in the membrane and expressed more clearly in the si-CXCR4
group. MMP2/9 was located in the cytoplasm and was inhibited in the
si-CXCR4 (magnification, ×400). TSCCA, tongue squamous cell
carcinoma; CXCR4, C-X-C chemokine receptor type 4; si-CXCR4, small
interfering RNA to CXCR4; control, blank control group; NC,
negative control small interfering RNA group; MMP, matrix
metalloproteinases. |
Discussion
Chemokines are a superfamily of small secreted
proteins that have been identified as attractants of leukocytes to
sites of infection and inflammation, and are pivotal in host
defense mechanisms. They are locally produced in tissues and act on
various cells through interactions with a subset of
seven-transmembrane G-protein-coupled receptors (23). Among the numerous chemokines and their
receptors, SDF-1, also referred to as the CXCL12/CXCR4 system, has
been demonstrated to be involved in lymph node or distant
metastasis in several types of cancer (17,24).
There is accumulating evidence that the CXCR4 system
may facilitate lymph node metastasis in OSCC. Almofti et al
(25) reported that invasion and
recurrence of tumors was strongly associated with CXCR4 expression,
and a CXCR4-positive group of patients had a poorer prognosis
compared with a CXCR4-negative group. In addition, Uchida et
al (26) examined the expression
of 13 types of chemokine receptors and chemokines in OSCC cells,
and revealed that upregulation of CXCR4 mRNA was detectable only in
lymph node metastatic cells, including HNt and B88 cells, in
comparison with nonmetastatic OSCC cells and normal gingival
epithelial cells. A subsequent study confirmed that lymph node
metastasis, loss in body weight and increase in tumour volumes were
significantly inhibited in mice inoculated with siRNA against CXCR4
cells compared with mice inoculated with control cells (27). The present study demonstrated that
CXCR4 was highly expressed in TSCCA cells. Subsequently, the
present study verified that siRNA-mediated CXCR4 silencing
significantly inhibited TSCCA migration and invasion using wound
healing and Transwell assays. These results indicate that CXCR4
expression is a possible marker of highly-invasive OSCC.
Previous studies have revealed interactions between
CXCR4 and EMT, which involves a series of events leading to tumor
invasion and metastasis (28). In a
study concerning hepatocellular cancer, intervention with exogenous
SDF-1 induced invasion and downregulated E-cadherin expression, and
upregulated vimentin expression in HepG2 cells; however, this
effect was not observed in CXCR4-depleted carcinoma (29). Zhu et al (30) silenced CXCR4 in glioma U87 cells and
revealed that EMT was inhibited. In addition, the authors
demonstrated that the production of transforming growth factor
(TGF)-β and β-catenin were decreased by CXCR4 silencing, indicating
that the effect of CXCR4 on EMT may be associated with its function
of affecting transcription factors. Another study revealed that
SDF-1/CXCR4 signaling induced pancreatic cancer EMT through the
activation of the Hedgehog pathway (31). The present study demonstrated that in
the CXCR4 siRNA group of cells, the expression of N-cadherin and
MMP2/9 were attenuated, while E-cadherin was upregulated, and the
same results were observed in xenograft models. In addition,
treatment of xenografted tumors with CXCR4 siRNA resulted in
histological alterations to tumor cells in the present study.
However, further studies are required to clarify the mechanism of
CXCR4 regulating OSCC tumor metastasis via EMT.
In addition to supporting metastasis, CXCR4
regulates tumor growth and apoptosis (25,32). The
present study demonstrated that inhibition of CXCR4 expression
promoted cell apoptosis of OSCC cells in vitro and,
following xenograft tumor model construction in nude mice, the
tumor volume in the CXCR4 siRNA treatment group was significantly
decreased compared with the blank control and negative control
groups. A previous study demonstrated that CXCR4 promoted cancer
cell proliferation by altering the expression of >1,500 genes
involved in the cell cycle, apoptosis and multiple signaling
pathways using microarray analysis technology (33). Some of these genes, including
mitogen-activated protein kinases, TGF-β and MMP, are central in
EMT (33). In addition, the
determination of cellular fates of apoptosis or survival during
tumor progression is highly affected by EMT; EMT confers cancer
cells resistant to apoptosis (34,35).
Therefore, growth inhibition and apoptosis promotion may be
attributable to the EMT conferred by CXCR4 siRNA.
The present results demonstrate that overexpression
of CXCR4 is significantly associated with EMT in OSCC cells. In
view of the importance of CXCR4, the expression of CXCR4 in OSCC
may be used as a molecular target of diagnosis, not only for lymph
node metastatic potential, but also for EMT. In addition, blockade
of CXCR4 may be investigated as a potential therapeutic target for
OSCC treatment.
Acknowledgements
The present study was supported by the Tianjin
Science and Technology Committee (Tianjin, China; grant no.
12JCYBJC33800).
References
|
1
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2012. CA Cancer J Clin. 62:10–29. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Noguti J, De Moura CF, De Jesus GP, Da
Silva VH, Hossaka TA, Oshima CT and Ribeiro DA: Metastasis from
oral cancer: An overview. Cancer Genomics Proteomics. 9:329–335.
2012.PubMed/NCBI
|
|
3
|
Patel SG, Amit M, Yen TC, Liao CT,
Chaturvedi P, Agarwal JP, Kowalski LP, Ebrahimi A, Clark JR, Cernea
CR, et al: Lymph node density in oral cavity cancer: Results of the
International consortium for outcomes research. Br J Cancer.
109:2087–2095. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Warnakulasuriya S: Global epidemiology of
oral and oropharyngeal cancer. Oral Oncol. 45:309–316. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sano D and Myers JN: Metastasis of
squamous cell carcinoma of the oral tongue. Cancer Metastasis Rev.
26:645–662. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Micalizzi DS, Farabaugh SM and Ford HL:
Epithelial-mesenchymal transition in cancer: Parallels between
normal development and tumor progression. J Mammary Gland Biol
Neoplasia. 15:117–134. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Smith A, Teknos TN and Pan Q: Epithelial
to mesenchymal transition in head and neck squamous cell carcinoma.
Oral Oncol. 49:287–292. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zheng H and Kang Y: Multilayer control of
the EMT master regulators. Oncogene. 33:1755–1763. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Oberlin E, Amara A, Bachelerie F, Bessia
C, Virelizier JL, Arenzana-Seisdedos F, Schwartz O, Heard JM,
Clark-Lewis I, Legler DF, et al: The CXC chemokine SDF-1 is the
ligand for LESTR/fusin and prevents infection by
T-cell-line-adapted HIV-1. Nature. 382:833–835. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelial-mesenchymal transitions in development and disease.
Cell. 139:871–890. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Murdoch C: CXCR4: Chemokine receptor
extraordinaire. Immunol Rev. 177:175–184. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Teng F, Tian WY, Wang YM, Zhang YF, Guo F,
Zhao J, Gao C and Xue FX: Cancer-associated fibroblasts promote the
progression of endometrial cancer via the SDF-1/CXCR4 axis. J
Hematol Oncol. 9:82016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yang DL, Xin MM, Wang JS, Xu HY, Huo Q,
Tang ZR and Wang HF: Chemokine receptor CXCR4 and its ligand CXCL12
expressions and clinical significance in bladder cancer. Genet Mol
Res. 14:17699–17707. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Guo Q, Gao BL, Zhang XJ, Liu GC, Xu F, Fan
QY, Zhang SJ, Yang B and Wu XH: CXCL12-CXCR4 axis promotes
proliferation, migration, invasion, and metastasis of ovarian
cancer. Oncol Res. 22:247–258. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wu W, Qian L, Chen X and Ding B:
Prognostic significance of CXCL12, CXCR4, and CXCR7 in patients
with breast cancer. Int J Clin Exp Pathol. 8:13217–13224.
2015.PubMed/NCBI
|
|
16
|
Gros SJ, Kurschat N, Drenckhan A, Dohrmann
T, Forberich E, Effenberger K, Reichelt U, Hoffman RM, Pantel K,
Kaifi JT and Izbicki JR: Involvement of CXCR4 chemokine receptor in
metastastic HER2-positive esophageal cancer. PLoS One.
7:e472872012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Müller A, Homey B, Soto H, Ge N, Catron D,
Buchanan ME, McClanahan T, Murphy E, Yuan W, Wagner SN, et al:
Involvement of chemokine receptors in breast cancer metastasis.
Nature. 410:50–56. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
An H, Xu L, Zhu Y, Lv T, Liu W, Liu Y, Liu
H, Chen L, Xu J and Lin Z: High CXC chemokine receptor 4 expression
is an adverse prognostic factor in patients with clear-cell renal
cell carcinoma. Br J Cancer. 110:2261–2268. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Scotton CJ, Wilson JL, Scott K, Stamp G,
Wilbanks GD, Fricker S, Bridger G and Balkwill FR: Multiple actions
of the chemokine CXCL12 on epithelial tumor cells in human ovarian
cancer. Cancer Res. 62:5930–5938. 2002.PubMed/NCBI
|
|
20
|
Burger JA and Kipps TJ: CXCR4: A key
receptor in the crosstalk between tumor cells and their
microenvironment. Blood. 107:1761–1767. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Duda DG, Kozin SV, Kirkpatrick ND, Xu L,
Fukumura D and Jain RK: CXCL12 (SDF1alpha)-CXCR4/CXCR7 pathway
inhibition: An emerging sensitizer for anticancer therapies? Clin
Cancer Res. 17:2074–2080. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang B, Zhang CP, Zhang S and Wang XD:
Epithelial-mesenchymal transformation-mediated lymph node
metastasis of oral squamous cell carcinoma and its mechanism.
Chinese Journal of Clinical Oncology. 39:1877–1880. 2012.
|
|
23
|
Rossi D and Zlotnik A: The biology of
chemokines and their receptors. Annu Rev Immunol. 18:217–242. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Perissinotto E, Cavalloni G, Leone F,
Fonsato V, Mitola S, Grignani G, Surrenti N, Sangiolo D, Bussolino
F, Piacibello W and Aglietta M: Involvement of chemokine receptor
4/stromal cell-derived factor 1 system during osteosarcoma tumor
progression. Clin Cancer Res. 11:490–497. 2005.PubMed/NCBI
|
|
25
|
Almofti A, Uchida D, Begum NM, Tomizuka Y,
Iga H, Yoshida H and Sato M: The clinicopathological significance
of the expression of CXCR4 protein in oral squamous cell carcinoma.
Int J Oncol. 25:65–71. 2004.PubMed/NCBI
|
|
26
|
Uchida D, Begum NM, Almofti A, Nakashiro
K, Kawamata H, Tateishi Y, Hamakawa H, Yoshida H and Sato M:
Possible role of stromal-cell-derived factor-1/CXCR4 signaling on
lymph node metastasis of oral squamous cell carcinoma. Exp Cell
Res. 290:289–302. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Uchida D, Onoue T, Kuribayashi N, Tomizuka
Y, Tamatani T, Nagai H and Miyamoto Y: Blockade of CXCR4 in oral
squamous cell carcinoma inhibits lymph node metastases. Eur J
Cancer. 47:452–459. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang SS, Han ZP, Jing YY, Tao SF, Li TJ,
Wang H, Wang Y, Li R, Yang Y, Zhao X, et al: CD133(+) CXCR4(+)
colon cancer cells exhibit metastatic potential and predict poor
prognosis of patients. BMC Med. 10:852012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li X, Li P, Chang Y, Xu Q, Wu Z, Ma Q and
Wang Z: The SDF-1/CXCR4 axis induces epithelial-mesenchymal
transition in hepatocellular carcinoma. Mol Cell Biochem.
392:77–84. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhu Y, Yang P, Wang Q, Hu J, Xue J, Li G,
Zhang G, Li X, Li W, Zhou C, et al: The effect of CXCR4 silencing
on epithelial-mesenchymal transition related genes in glioma U87
cells. Anat Rec (Hoboken). 296:1850–1856. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li X, Ma Q, Xu Q, Liu H, Lei J, Duan W,
Bhat K, Wang F, Wu E and Wang Z: SDF-1/CXCR4 signaling induces
pancreatic cancer cell invasion and epithelial-mesenchymal
transition in vitro through non-canonical activation of Hedgehog
pathway. Cancer Lett. 322:169–176. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Thomas RM, Kim J, Revelo-Penafiel MP,
Angel R, Dawson DW and Lowy AM: The chemokine receptor CXCR4 is
expressed in pancreatic intraepithelial neoplasia. Gut.
57:1555–1560. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yu T, Wu Y, Huang Y, Yan C, Liu Y, Wang Z,
Wang X, Wen Y, Wang C and Li L: RNAi targeting CXCR4 inhibits tumor
growth through inducing cell cycle arrest and apoptosis. Mol Ther.
20:398–407. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Valdés F, Alvarez AM, Locascio A, Vega S,
Herrera B, Fernández M, Benito M, Nieto MA and Fabregat I: The
epithelial mesenchymal transition confers resistance to the
apoptotic effects of transforming growth factor Beta in fetal rat
hepatocytes. Mol Cancer Res. 1:68–78. 2002.PubMed/NCBI
|
|
35
|
Robson EJ, Khaled WT, Abell K and Watson
CJ: Epithelial-to-mesenchymal transition confers resistance to
apoptosis in three murine mammary epithelial cell lines.
Differentiation. 74:254–264. 2006. View Article : Google Scholar : PubMed/NCBI
|