Introduction
Acute myeloid leukemia (AML) is a genetically
heterogeneous clonal disorder caused by the build up of somatic
mutations in hematopoietic progenitor cells, which affects the
regulation of self-renewal, survival, proliferation and
differentiation (1). The induction of
differentiation is a desired consequence of chemopreventive and
therapeutic agents, as it often results in the elimination of
premalignant or malignant cells (2).
Numerous studies focus on selectively killing tumor cells through
differentiation induction (3).
Therefore, the development of novel differentiation-inducing drugs
for blocking AML is of clinical significance.
For the past 30 years, studies have explored the use
of diallyl disulfide (DADS), the main active component of the
cancer-fighting allyl sulfides found in garlic, as it has been
shown to reduce the initiation of carcinogen-induced cancers and
inhibit the proliferation of various types of cancer cells
(4). The actions of DADS include the
regulation of cell cycle arrest, induction of apoptosis and cell
differentiation, and inhibition of cell invasion (5–7). Previous
studies at the Cancer Research Institute, University of South China
(Hengyang, China) confirmed that DADS can inhibit the proliferation
of human leukemia cells in vivo in a dose-dependent manner
(8,9).
DADS exhibits a dual effect: A medium dose (>1.25 mg/l) can
induce apoptosis in human leukemia cells (8,9), and a
small dose (<1.25 mg/l) can induce human leukemia cell
differentiation (2). The mechanisms
of inducing differentiation involve: G2/M-phase cell
cycle arrest; histone acetylation; the regulation of regulatory
gene expression, including signal transducer and activator of
transcription 3, v-myc avian myelocytomatosis viral oncogene
homolog, Fos proto-oncogene and Jun proto-oncogene regulation; and
the upregulation of cyclin-dependent kinase inhibitor 1 expression
(10–12). Proteomic analysis was also used to
explore differentially expressed proteins in DADS-induced
differentiation in human leukemia HL-60 cells (13). The results showed decreased expression
of calreticulin (CRT), an endoplasmic reticulum-resident protein,
in differentiated HL-60 cells induced by DADS (13), suggesting that CRT is involved in
DADS-mediated induction of differentiation in HL-60 cells. Although
the role of DADS as an antitumor agent has been established, its
exact cytotoxic mechanism in differentiation is not entirely
clear.
CRT, a multi-process calcium-buffering chaperone of
the endoplasmic reticulum, is essential for numerous cellular
functions, including lectin-like chaperoning, Ca2+
storage and signaling, the regulation of gene expression, cell
adhesion, wound healing, cancer and autoimmunity (14). Recently, numerous studies have shown
that CRT is important in tumorigenesis and prognosis (15,16). The
increased expression of CRT was indicated in the urine samples of
patients with bladder cancer, indicating CRT as a biomarker in
bladder cancer (17). In gastric
cancer, positive immunohistochemical staining of CRT was correlated
with high microvessel density, serosal and perineural invasion,
lymph node metastasis and poor patient survival (18). However, in neuroblastoma, which is the
most common malignancy in infants, positive immunohistochemical
staining for CRT was associated with a better prognosis and patient
survival (19). CRT was also
differentially expressed in colorectal cancer (20). CRT-overexpressing gastric cancer cells
demonstrate increased proliferation rates, while CRT-knockdown
gastric cancer cell lines demonstrated decreased proliferation
rates (20). In addition,
CRT-overexpressing cells showed greater wound healing and migration
rates compared with CRT-knockdown cells (21). In a recent preliminary study of 33
breast cancer patients, Kabbage et al (22) observed a potential association between
CRT overexpression and axillary lymph node metastasis in breast
cancer patients.
Certain studies have indicated that CRT is
upregulated in all AML subtypes in the French-American-British
classification of leukemia cells (23,24). In
addition, the expression of CRT was significantly reduced during
certain drug-induced leukemia cell differentiation (25).
In the present study, CRT was hypothesized to be an
important factor in DADS-induced proliferation, migration and
differentiation in human leukemia HL-60 cells. The present study
examined the role of CRT on migration and differentiation in human
leukemia HL-60 cells treated with DADS. These results may lead to a
better understanding of the antitumor molecular mechanisms of DADS
and provide essential knowledge for the development of
differentiation inducers to treat leukemia.
Materials and methods
Materials and reagents
Fluka Chemika Company DADS was purchased from
Sigma-Aldrich (Buchs, Switzerland). The Total RNA Kit II extraction
kit was purchased from Omega Bio-Tek, Inc. (Norcross, GA, USA). A
High Capacity cDNA Reverse Transcription kit was purchased from
Promega Corporation (Madison, WI, USA). The Bestar™ qPCR RT Kit
(#DBI-2220) and Bestar™ SybrGreen qPCR Mastermix (#DBI-2043) were
purchased from DBI Bioscience (Ludwigshafen, Germany). The CRT
(mouse monoclonal; #ab22683; dilution, 1:500), GAPDH (mouse
monoclonal; #ab8245; dilution, 1:500), cluster of differentiation
(CD)11b (rabbit monoclonal; #ab52478; dilution, 1:200) and CD33
(mouse monoclonal; #ab119860; dilution, 1:200) primary antibodies,
and the horseradish peroxidase-conjugated secondary antibodies
[goat anti-mouse IgG H&L (#ab6789; dilution, 1:1,000) and goat
anti-rabbit IgG H&L (#ab6721; dilution, 1:1,000)] were
purchased from Abcam (Cambridge, UK). Small interfering RNA (siRNA)
transfection reagent, siRNA transfection medium, CRT siRNA and a
O-GlcNAc Western Blot Detection Kit were purchased from Santa Cruz
Biotechnology (Dallas, TX, USA). Nitroblue tetrazolium (NBT) was
purchased from Sigma-Aldrich (St. Louis, MO, USA). The Invitrogen
plasmids (pcDNA3.1-12GS0643-IG-3, KL121217003) and vector
(pcDNA3.1) were obtained from Thermo Fisher Scientific, Inc.
(Waltham, MA, USA).
Cell culture
Human AML HL-60 cells, obtained from Central South
University (Hengyang, China), were incubated in RPMI-1640 culture
medium (Hyclone; GE Healthcare Life Sciences, Logan, UT, USA)
containing 10% calf serum (Gibco; Thermo Fisher Scientific, Inc.)
at 37°C, at saturated humidity and 5% CO2. Cell cultures
were replaced with fresh medium every 2–3 days. Cells in
logarithmic growing phase were used for experiments. DADS was
diluted to 1.25 mg/l in culture medium.
Wright-Giemsa staining
Cells were collected, smeared on a slide and
air-dried. Subsequently, 2–3 drops of Wright-Giemsa dye solution
were added to the cells, and after 1 min, the slide was rinsed with
distilled water. The sample was dried and observed under a
microscope.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Expression levels of the CRT gene in HL-60 cells
were measured by SYBR Green RT-qPCR. β-actin was used as an
internal control. All primer sequences were designed by Premier 5.0
software (Premier Biosoft International, Palo Alto, CA, USA) and
synthesized by Takara Bio Inc. (Otsu, Japan). The primer sequences
were as follows: β-actin sense, 5′-GGACCTGACTGACTACCTC-3′ and
antisense, 5′-TAGTCGTTCGTCCTCATAC-3′; and CRT sense,
5′-GGAAGATGAGGAGGAAGATGTC-3′ and antisense,
5′-CAGGAAGGAGAGCAGATGAAAT-3′. Total cell RNA was extracted
according to the instructions of the Total RNA Kit II (Omega
Bio-Tek, Inc.), and the RNA purity and concentration were measured
using a NanoDrop® ND-1000 (Thermo Fisher Scientitic,
Inc.). Total RNA was subjected to RT by first heating 10 µl of the
annealing mixture, which contained 2 µg total RNA and 1 µl 0.5
µg/µl oligo (dT)18, to 70°C for 3 min. After cooling to
37°C for 10 min, 2 µl 10X RT reaction buffer, 4 µl 2.5 mM dNTP
mixture, 1 µl RNase inhibitor and 200 units of moloney murine
leukemia virus reverse transcriptase were added. In a PCR machine
(Applied Biosystems; Thermo Fisher Scientific, Inc.), the mixture
was first incubated at 37°C for 1 h, then heated to 95°C for 5 min
and cooled on ice. For qPCR, a 25-µl reaction mixture, consisting
of 1 µl cDNA, 1 µl Primer1, 1 µl Primer2, 2×1 unit Taq Master Mix,
2.5 µl 2.5 mM dNTPs, 2.5 µl 10X PCR buffer, 1.5 µl MgCl2
and 10,000-fold diluted SYBR Green, was used. All PCR reactions
were performed at 95°C for 5 min, followed by 40 cycles of 95°C for
10 sec, 59°C for 15 sec, and 72°C for 20 sec. In order to establish
the melting curve of PCR products obtained from the reaction, the
samples were then heated between 72 and 99°C, rising 1°C every 5
sec. The results were quantified by dissociation and amplification
curves (26).
Fast-forward transfection cell
transfection
Experimental plasmid (pcDNA3.1-12GS0643-IG-3),
negative plasmid (pcDNA3.1-neg-IRES2-EGFP) and vectors (pcDNA3.1),
0.4 µg each, were dissolved in Tris-ethylenediaminetetraacetic acid
(TE) buffer (pH 7–8; minimum DNA concentration, 0.1 µg/µl) with
RPMI-1640 without serum, proteins or antibiotics to a total volume
of 60 µl. Attractene transfection reagent (Qiagen, Inc., Valencia,
CA, USA) (1.5 µl) was added to the DNA and mixed by pipetting or
vortexing, and if necessary, the sample was centrifuged for a few
seconds to remove any liquid from the top of the tube. The samples
were incubated for 10–15 min at room temperature to allow
transfection complex formation, and then added to 500 µl of freshly
harvested cell suspension (to a final cell density of
0.4–1.6×105 cells in 500 µl), which was added to a well
of a 24-well plate. Cells were incubated with the transfection
complexes under normal growth conditions and assayed 24 h after the
fast-forward transfection.
RNA interference
Cells were collected through centrifugation and the
concentration was adjusted to 1–2×106 cells/ml. The
cells were then washed once with 2 ml siRNA transfection medium.
Two various preparations were used: Solution A, consisting of 6 µl
CRT siRNA duplex in 100 µl siRNA transfection medium, and solution
B, including 6 µl siRNA transfection reagent in 100 µl siRNA
transfection medium. Solution A was then directly added to solution
B using a pipette, and the solutions were mixed gently and
incubated for 30 min at room temperature. siRNA transfection medium
(0.8 ml) was added to each tube containing the siRNA transfection
reagent mixture, and the mixture was overlaid onto cells. Cells
were incubated for 5–7 h at 37°C in a CO2 incubator, and
then 1 ml of normal growth medium containing two times the normal
serum amount (8% calf serum) was directly added without removing
the transfection mixture. Cells were incubated for an additional
18–24 h, after which the medium was removed and replaced with fresh
normal growth medium. At 24–72 h, cells were harvested for
analysis.
Western blot analysis
Total cell proteins from the different groups were
harvested (untreated HL-60 cells were used as the normal control),
and 25 µg of each sample was mixed with 5X sodium dodecyl sulfate
(SDS) loading buffer at 5:1 ratio, denatured by heating, and
separated by 10% SDS-polyacrylamide gel electrophoresis. Samples
were transferred to a nitrocellulose membrane, and membranes were
blocked with Tris-buffered saline and Tween 20 (TBST) containing 5%
non-fat milk for 2 h. The membrane was washed with TBST three
times, and incubated with the CRT, CD11b, CD33 or GAPDH (internal
control) primary antibodies for 2 h at 37°C, followed by three
washes with TBST for 15 min each. The membranes were incubated with
the corresponding secondary antibody for 1 h, washed with TBST
three times for 10 min each, and finally developed using BeyoECL
Plus (Beyotime Biotechnology, Turku, Finland).
Invasion assay
Transwell inserts (pore diameter, 8 µm; diameter,
6.3 mm) were inserted in the chamber of a 24-well transwell plate.
Serum-free RPMI-1640 with BD Matrigel (5 mg/ml; dilution, 8:1; BD
Biosciences, Franklin Lakes, NJ, USA) was added into the bottom of
the Transwell chamber and plates were incubated at 37°C overnight.
Cells (1×106 cells/ml) of the different groups (with
untreated HL-60 cells used as the normal control) were added into
the upper chamber, and RPMI-1640 medium containing serum was added
into the lower chamber. The plates were incubated for 48 h, the
supernatant was removed, and then the plates were washed with
phosphate-buffered saline (PBS). Samples were fixed with 10%
formalin for 20–30 min, stained with hematoxylin for 10–20 min and
air-dried. The invading cells were counted using an inverted
microscope.
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-tetrazolium bromide (MTT)
assay
Cell viability was determined using a conventional
MTT reduction assay (27). HL-60
cells (1×104 cells/well) in 96-well plates were treated
with 1.25 mg/l DADS for 48 h. Untreated HL-60 cells were used as
the normal control. The medium was removed and 100 µl of dimethyl
sulfoxide was added to each well to dissolve the formazan dye
crystals for 15 min. The optical density was measured at an
absorbance of 490 nm with a microplate reader.
Differentiation assay
NBT reduction was performed as previously described
(28). Cells were incubated in
96-well culture plates and collected through centrifugation. Cells
were the resuspended with 200 µl NBT solution. Following 60 min of
culture, centrifugation was performed, 200 µl DMSO was added and
the cells were oscillated for 20 min. Absorbance was measured at
570 nm.
Flow cytometry (FCM) analysis
To evaluate the effect of CRT siRNA transfection and
CRT overexpression on HL-60 cells, CD11b and CD33 levels were
evaluated. Cells from the different groups (with untreated HL-60
cells included as the normal control) were collected, centrifuged
at 500 × g for 5 min, washed twice with PBS, and the cell
concentration was adjusted to 1×106 cells/ml. Cells were
then fixed with 70% ethanol. Prior to FCM analysis, the cells were
washed twice again with PBS and 500 µl 0.2% Triton-X 100 was added.
The mixture was incubated on ice for 10 min. After washing the
samples twice again with PBS, 50 µl primary antibody was added and
cells were incubated on ice for 30–45 min. After washing with PBS,
50 µl specific fluorescent secondary antibody was added, and cells
were incubated on ice for 30 min. Subsequent to washing with PBS,
the cells were immediately measured by FCM (FlowJo version 9;
FlowJo, LLC, Ashland, OR, USA).
Statistics
SPSS 12.0 statistical software (SPSS, Inc., Chicago,
IL, USA) was used to analyze the results. Experimental data were
presented as mean ± standard deviation, and the differences between
experimental and control groups were detected by the Student's
t-test. P<0.05 was considered to indicate a statistically
significant difference.
Results
Calreticulin is downregulated during
DADS-induced differentiation in HL-60 cells
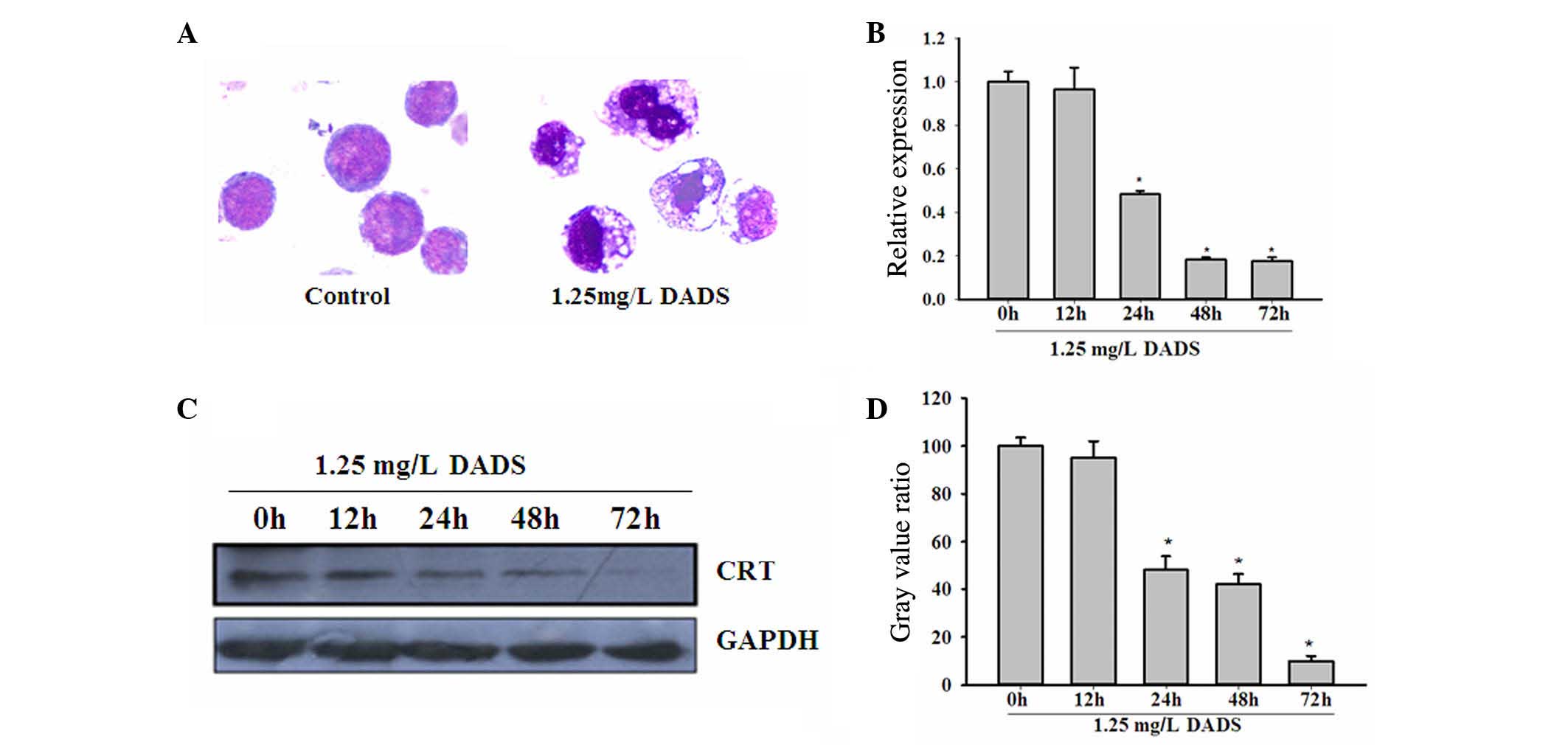
As shown in Fig. 1A,
the treatment of HL-60 cells with 1.25 mg/l DADS for 48 h induced
cells to differentiate into granulocyte-like cells. The
cytomorphoses of the DADS-treated cells were clearly varied from
the controls. RT-PCR confirmed the significant downregulation of
CRT messenger RNA (mRNA) levels in HL-60 cells following treatment
with 1.25 mg/l DADS in a time-dependent manner, compared with
untreated HL-60 cells (P=0.042; Fig.
1B). Western blot analysis was consistent with the RT-PCR
results, showing a decrease in CRT protein levels in HL-60 cells
treated with 1.25 mg/l DADS in a time-dependent manner (P=0.021;
Fig. 1C and D).
CRT affects proliferation of
DADS-treated HL-60 cells
The present study then analyzed the functional role
of CRT in DADS-induced effects on HL-60 cells using CRT
overexpression and CRT downregulation mediated by siRNA. The RT-PCR
and western blot analyses confirmed that CRT knockdown by RNA
interference was ~70%. In the DADS+siRNA-treated group, CRT
expression was further inhibited (P=0.027). Notably, upon treatment
of untransfected and CRT-overexpressing cells with 1.25
mg·l−1 DADS for 48 h, CRT levels were significantly
reduced compared with the corresponding untreated cells (P=0.019),
indicating that DADS could downregulate CRT expression in HL-60
cells (Fig. 2A and B).
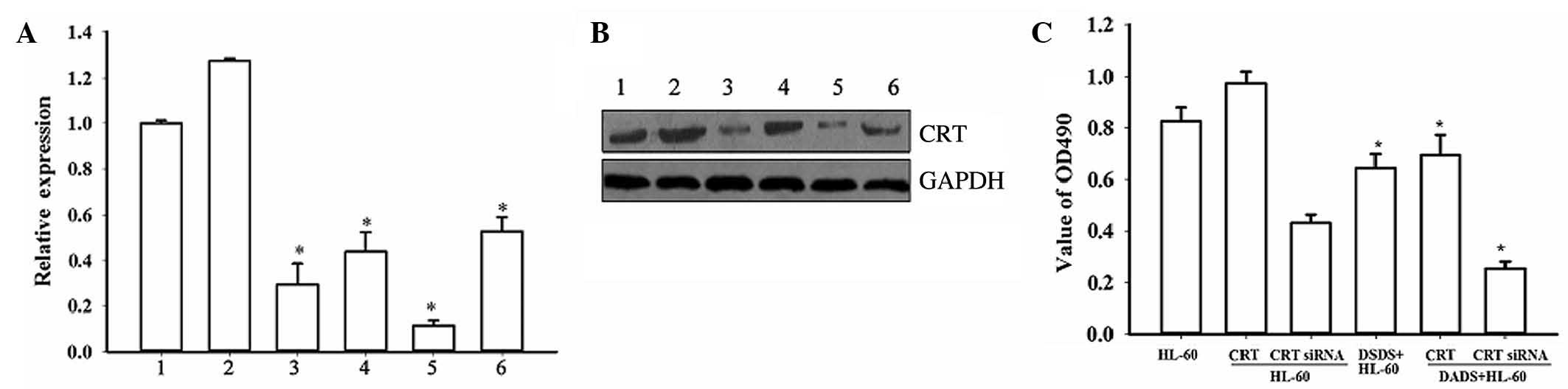 | Figure 2.Effect of CRT overexpression and CRT
downregulation on proliferation of HL-60 cells induced by DADS.
Inhibition of CRT expression by siRNA and CRT overexpression by
plasmid transfection in HL-60 cells were evaluated by (A) reverse
transcription-polymerase chain reaction: 1, HL-60; 2, HL-60/CRT; 3,
HL-60/CRT siRNA; 4, DADS+HL-60; 5, DADS+HL-60/CRT siRNA; 6,
DADS+HL-60/CRT (*P<0.05 vs. control cells) and (B) western blot
analysis: 1, HL-60; 2, HL-60/CRT; 3, HL-60/CRT siRNA; 4,
DADS+HL-60; 5, DADS+HL-60/CRT siRNA; 6, DADS+HL-60/CRT. (C)
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-tetrazolium bromide
analysis in CRT-overexpressing and CRT-downregulated HL-60 cells,
with and without DADS treatment. *P<0.05 vs. control cells. CRT,
calreticulin; DADS, diallyl disulfide; siRNA, small interfering
RNA; GAPDH, glyceraldehyde 3-phosphate dehydrogenase. |
MTT assays demonstrated that downregulation of CRT
by siRNA resulted in decreased proliferation of HL-60 cells
compared with the controls, while CRT overexpression increased cell
proliferation (Fig. 2C). These
results indicate that CRT has a function in HL-60 cell
proliferation. Notably, following treatment with DADS for 48 h, the
proliferation rates of all cells were decreased compared with the
corresponding untreated groups (P=0.038). Together, these data
indicate that CRT is involved in cell proliferation in DADS-induced
HL-60 cells.
CRT affects cell invasion in
DADS-treated HL-60 cells
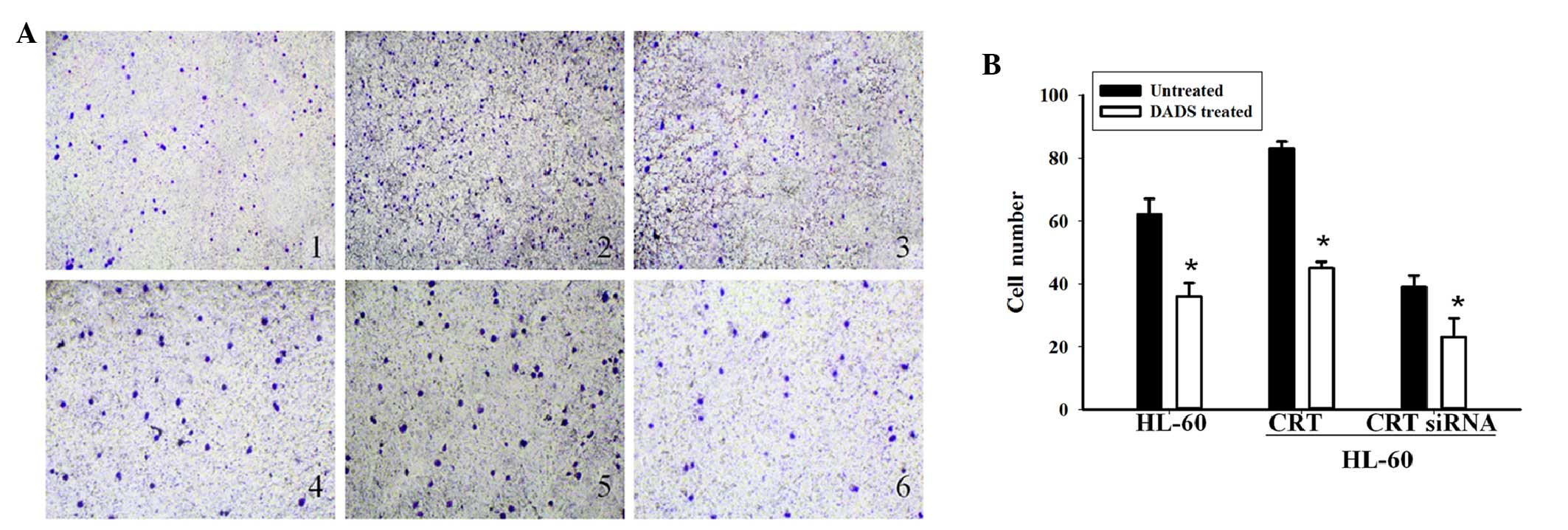
The present study next investigated the role of CRT
in cell invasion induced by DADS treatment. Transwell Matrigel
invasion assays showed that the number of HL-60 cells that
penetrated through the Matrigel in the non-transfected group was
62±6.37, compared with 83±11.39 in the CRT overexpression group and
39±2.43 in the CRT siRNA group (Fig. 3A
and B). These results indicate that CRT overexpression can
increase the ability of invasion in HL-60 cells and CRT siRNA
transfection can decrease the ability of invasion. Following
treatment with DADS for 48 h, the number of HL-60 cells that
penetrated through the Matrigel was 36.2±7.59 in the
non-transfected group, compared with 45.6±4.10 and 23.45±3.86 in
the CRT overexpressing and downregulated groups, respectively.
Cellular invasion in the DADS treated groups was decreased compared
with the untreated groups (P=0.034). These results indicate that
CRT is involved in cell invasion in DADS-induced HL-60 cells.
CRT affects cell differentiation in
DADS-treated HL-60 cells
The present study finally investigated the role of
CRT in cell differentiation induced by DADS treatment. As shown in
Fig. 4A, FCM analysis showed that,
compared with control cells, the expression of CD33 in the CRT
siRNA group was markedly reduced (93.53 vs. 51.22%, respectively),
while the CRT-overexpressed groups showed slightly increased CD33
expression (95.76%). Following treatment with DADS for 48 h, the
expression of CD33 was significantly decreased in non-transfected
HL-60 cells (55.46%), CRT-overexpressing HL-60 cells (63.09%), and
CRT siRNA HL-60 cells (31.46%) (P=0.037). As shown in Fig. 4B, compared with the control cells
(1.00%), the CRT-overexpressed group showed decreased CD11b
expression (0.81%) and the CRT siRNA group showed significantly
increased CD11b expression (7.37%). After DADS treatment for 48 h,
the control group, CRT-overexpressed group and the CRT siRNA group
all showed increased CD11b expression (8.24, 6.55 and 12.42%,
respectively; P=0.033). NBT reduction assays showed decreased NBT
reduction activity in the CRT overexpression group and increased
NBT reduction in the CRT siRNA group (Fig. 4C). Following treatment with DADS, the
NBT reduction abilities in all groups were increased (P=0.024).
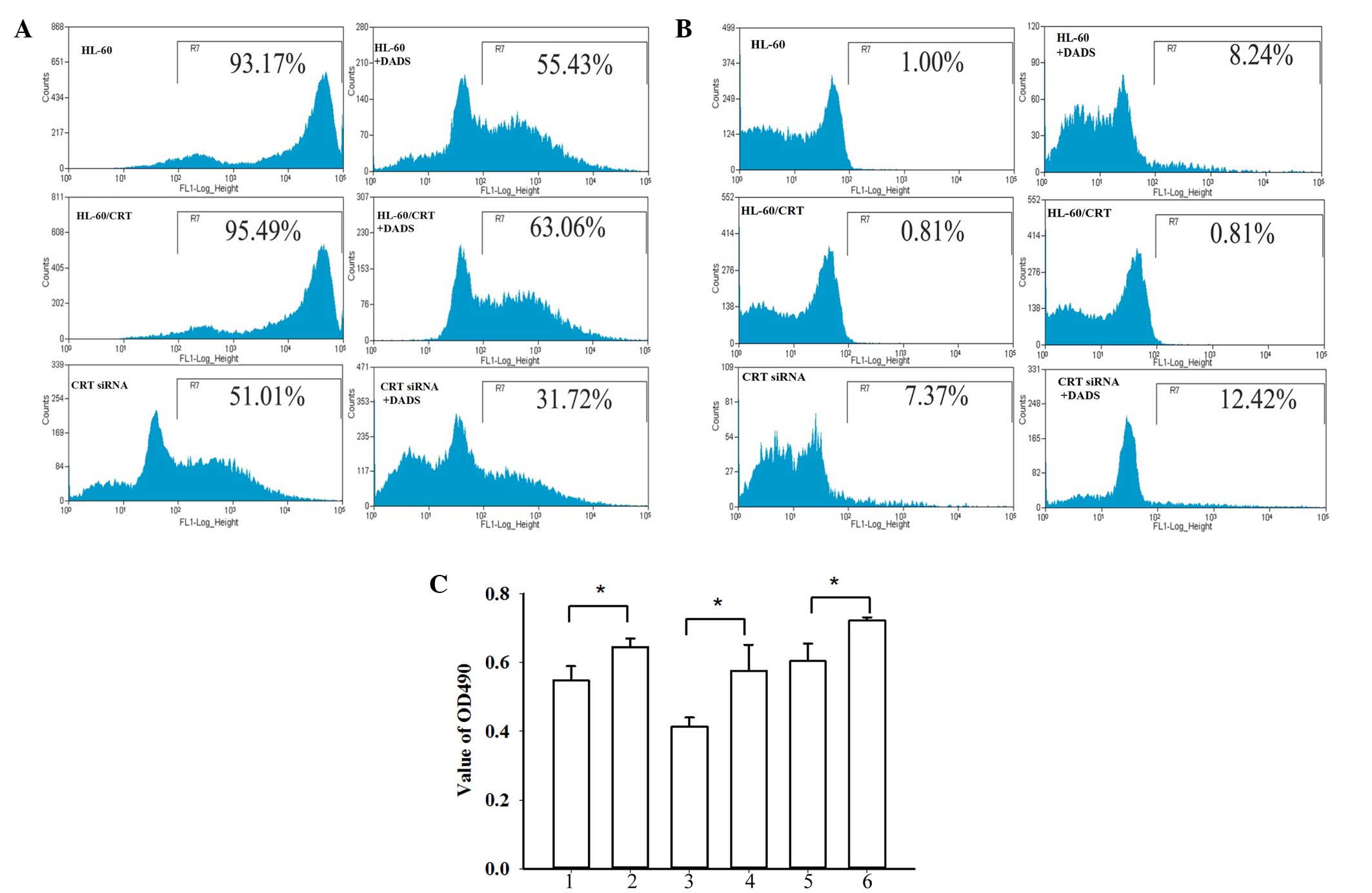 | Figure 4.Role of CRT in cell differentiation of
HL-60 cells induced by DADS. (A) Representative flow cytometry
results showing CD33 expression. (B) Representative flow cytometry
results showing CD11b expression. (C) Nitroblue tetrazolium
reduction assays: 1, HL-60; 2, DADS+ HL-60; 3, HL-60/CRT; 4,
DADS+HL-60/CRT; 5, HL-60/CRT siRNA; 6, DADS+HL-60/CRT siRNA. Data
are the mean ± standard deviation of three experiments. *P<0.05
vs. control; Student's t-test. CRT, calreticulin; DADS, diallyl
disulfide; CD33, cluster of differentiation 33; CD11b, cluster of
differentiation 11b; OD, optical density. |
Discussion
Large numbers of cytokines and drugs have been shown
to induce tumor cell differentiation. Among these, the role of
allyl sulfides, compounds that are found in garlic, in the
prevention and treatment of tumors has attracted attention
(29). DADS, the most active
anticancer ingredient in garlic, induces differentiation in several
tumor types, including HL-60 cells (5). However, the exact mechanism underlying
this process remains unclear.
Previous studies have demonstrated that DADS may
induce differentiation in leukemia cells (12,30). These
studies analyzed and identified differentially expressed proteins
during DADS-induced differentiation in HL-60 cells, including CRT.
CRT is a multi-functional protein that regulates various cellular
functions, such as calcium homeostasis and intracellular adhesion
(31). In addition, increased CRT
expression levels have been observed in several cancer tissues,
including hepatoma, neuroblastoma, mammary gland cancer, bladder
cancer and colon cancer (15). These
observations imply that high CRT expression levels could be
associated with carcinogenesis in these cancers.
Subsequent to treatment with DADS, the morphology of
the HL-60 cells used in the present study demonstrated
differentiation into granulocyte-like cells. Consistent with a
previous study (5), the results of
the RT-PCR and western blot analysis in the current study confirmed
the significant downregulation of CRT mRNA and proteins in
DADS-treated HL-60 cells compared with untreated cells. These data
suggest that CRT is closely associated with DADS-induced
differentiation in human leukemia HL-60 cells.
To further investigate the role of CRT in
DADS-induced cellular effects, CRT expression was inhibited using
siRNA and overexpressed CRT by plasmid transfection in HL-60 cells.
MTT assays showed that the downregulation of CRT resulted in the
inhibited proliferation of HL-60 cells, while CRT overexpression
increased proliferation. After treatment with DADS, cellular
proliferation in all cell groups was decreased compared with the
control. This indicates that CRT is involved in cell proliferation
in DADS-induced HL-60 cells. Transwell invasion assays showed that
CRT overexpression can increase cellular invasion ability and that
CRT downregulation can decrease invasion ability in HL-60 cells.
Following treatment with DADS, the number of invading HL-60 cells
in all groups was reduced compared with untreated groups. These
results indicate that CRT is involved in cell invasion in
DADS-induced HL-60 cells.
CD33 is a transmembrane receptor expressed on cells
of myeloid lineage and is usually considered to be myeloid specific
(32). CD33 is expressed in >90%
of AML patients, and is not expressed in the hematopoietic stem
cell surface, mature granule cells and other tissues (33). Thus, CD33 is a good target for myeloid
leukemia treatment (34). CD11b, also
known as cluster of differentiation 11b molecule, is expressed on
the surface of numerous leukocytes involved in the innate immune
system, including monocytes, granulocytes, macrophages and natural
killer cells (35). CD11b mediates
inflammation by regulating leukocyte adhesion and migration and has
been implicated in several immune processes, including
phagocytosis, cell-mediated cytotoxicity, chemotaxis and cellular
activation (36). Therefore, CD33 and
CD11b are important indexes to evaluate the differentiation of
myeloid leukemia cells. FCM analysis showed that the expression of
CD33 in the CRT siRNA group was markedly reduced, while expression
was increased in CRT-overexpressed groups. The expression of CD33
was significantly decreased in all cell groups following treatment
with DADS. The expression of CD11b was significantly increased in
the CRT siRNA group, and increased in the CRT-overexpressed group.
The expression of CD11b was significantly increased in all groups
following treatment with DADS. These data demonstrate that
DADS-induced HL-60 cells are differentiated to mature neutrophil
cells. NBT reduction assays showed that NBT reduction was increased
in the CRT siRNA group and reduced in the CRT overexpression group.
After treatment with DADS, NBT reduction abilities were increased.
These results indicate that CRT is involved in cell differentiation
in DADS-induced HL-60 cells.
In conclusion, the present study clearly
demonstrates the downregulation of CRT during DADS-induced
differentiation in HL-60 cells and indicates that CRT is involved
in cell proliferation, invasion and differentiation in DADS-induced
HL-60 cells. The data first demonstrated that CRT could play an
essential role in AML cell proliferation and invasion, and
therefore may be an important target for future research of human
AML. The exact mechanisms underlying DADS-induced tumor cell
differentiation require further investigation.
Acknowledgements
The present study was sponsored by the National
Natural Scientific Foundation of China (grant no. 81400117;
Beijing, China), the Hunan Provincial Natural Science Foundation of
China (grant no. 2015JJ4043; Changsha, China), China Postdoctoral
Science Foundation (grant no. 2014M562115; Beijing, China) and the
returned personnel Initial Funding of University of South China
(grant no. 2014XQD46; Hengyang, China).
References
|
1
|
Rubnitz JE and Inaba H: Childhood acute
myeloid leukaemia. Br J Haematol. 159:259–276. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Iland HJ, Bradstock K, Supple SG, Catalano
A, Collins M, Hertzberg M, Browett P, Grigg A, Firkin F, Hugman A,
et al: Australasian Leukaemia and Lymphoma Group:
All-trans-retinoic acid, idarubicin, and IV arsenic trioxide as
initial therapy in acute promyelocytic leukemia (APML4). Blood.
120:1570–1580. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
DiNardo CD and Cortes JE: New treatment
for acute myelogenous leukemia. Expert Opin Pharmacother.
16:95–106. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Arunkumar R, Sharmila G, Elumalai P,
Senthilkumar K, Banudevi S, Gunadharini DN, Benson CS, Daisy P and
Arunakaran J: Effect of diallyl disulfide on insulin-like growth
factor signaling molecules involved in cell survival and
proliferation of human prostate cancer cells in vitro and in silico
approach through docking analysis. Phytomedicine. 19:912–923. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yi L and Su Qi: Molecular mechanisms for
the anti-cancer effects of diallyl disulfide. Food Chem Toxicol.
57:362–370. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Altonsy MO, Habib TN and Andrews SC:
Diallyl disulfide-induced apoptosis in a breast-cancer cell line
(MCF-7) may be caused by inhibition of histone deacetylation. Nutr
Cancer. 64:1251–1260. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhou Y, Su J, Shi L, Liao Q and Su Q: DADS
downregulates the Rac1-ROCK1/PAK1-LIMK1-ADF/cofilin signaling
pathway, inhibiting cell migration and invasion. Oncol Rep.
29:605–612. 2013.PubMed/NCBI
|
|
8
|
Tan H, Ling H, He J, Yi L, Zhou J, Lin M
and Su Q: Inhibition of ERK and activation of p38 are involved in
diallyl disulfide induced apoptosis of leukemia HL-60 cells. Arch
Pharm Res. 31:786–793. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yi L, Ji XX, Tan H, Lin M, Tang Y, Wen L,
Ma YH and Su Q: Role of Ras-related C3 botulinum toxin substrate 2
(Rac2), NADPH oxidase and reactive oxygen species in diallyl
disulphide-induced apoptosis of human leukaemia HL-60 cells. Clin
Exp Pharmacol Physiol. 37:1147–1153. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yi L, Ji XX, Tan H, Feng MY, Tang Y, Wen L
and Su Q: Involvement of Mcl1 in diallyl disulfide-induced G2/M
cell cycle arrest in HL-60 cells. Oncol Rep. 27:1911–1917.
2012.PubMed/NCBI
|
|
11
|
Wu MH, Huang WG, Tan H, He J and Su Q:
Induction of differentiation by diallyl disulfide through
inhibition of JAK1/STAT3 in human leukemia HL-60 cells. Zhong Guo
Yao Li Xue Tong Bao. 21:580–583. 2005.(In Chinese).
|
|
12
|
Zhao J, Huang WG, He J, Tan H, Liao QJ and
Su Q: Diallyl disulfide suppresses growth of HL-60 cell through
increasing histone acetylation and p21WAF1 expression in
vivo and in vitro. Acta Pharmacol Sin. 27:1459–1466.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
He J, Su Q, Huang WG, Xie HL, Liang SP,
Song Y, Xie JY, Zhou XT, Tan H, Zhao J and Wu MH: Proteomic initial
analysis of differentiation of human myeloid leukemia cells induced
by diallyl disulfide. FEBS J. 272(Suppl 1): S4402005.
|
|
14
|
Wang WA, Groenendyk J and Michalak M:
Calreticulin signaling in health and disease. Int J Biochem Cell
Biol. 44:842–846. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zamanian M, Veerakumarasivam A, Abdullah S
and Rosli R: Calreticulin and Cancer. Pathol Oncol Res. 19:149–154.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu R, Gong J, Chen J, Li Q, Song C, Zhang
J, Li Y, Liu Z, Dong Y, Chen L and Jin B: Calreticulin as a
potential diagnostic biomarker for lung cancer. Cancer Immunol
Immunother. 61:855–864. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kageyama S, Isono T, Matsuda S, Ushio Y,
Satomura S, Terai A, Arai Y, Kawakita M, Okada Y and Yoshiki T:
Urinary calreticulin in the diagnosis of bladder urothelial
carcinoma. Int J Urol. 16:481–486. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen CN, Chang CC, Su TE, Hsu WM, Jeng YM,
Ho MC, Hsieh FJ, Lee PH, Kuo ML, Lee H and Chang KJ: Identification
of calreticulin as a prognosis marker and angiogenic regulator in
human gastric cancer. Ann Surg Oncol. 16:524–533. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hsu WM, Hsieh FJ, Jeng YM, Kuo ML, Chen
CN, Lai DM, Hsieh LJ, Wang BT, Tsao PN, Lee H, et al: Calreticulin
expression in neuroblastoma-a novel independent prognostic factor.
Ann Oncol. 16:314–321. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Alfonso P, Núñez A, Madoz-Gurpide J,
Lombardia L, Sánchez L and Casal JI: Proteomic expression analysis
of colorectal cancer by two-dimensional differential gel
electrophoresis. Proteomics. 5:2602–2611. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen CN, Su TE, Lu YC and Lee H:
Calreticulin regulates cell proliferation and migration in gastric
cancer cell line AGS. FASEB J. 21:A13182007.
|
|
22
|
Kabbage M, Trimeche M, Bergaoui S, Hammann
P, Kuhn L, Hamrita B, ben Nasr H, Chaieb A, Chouchane L and Chahed
K: Calreticulin expression in infiltrating ductal breast
carcinomas: Relationships with disease progression and humoral
immune responses. Tumour Biol. 34:1177–1188. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Park S, Huh HJ, Mun YC, Seong CM, Chung
WS, Chung HS and Huh J: Calreticulin mRNA expression and
clinicopathological characteristics in acute myeloid leukemia.
Cancer Genet. 208:630–635. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kourie HR, Ameye L, Paesmans M and Bron D:
Improved Survival of Calreticulin-Mutated Patients Compared With
Janus Kinase 2 in Primary Myelofibrosis: A Meta-Analysis. Clin
Lymphoma Myeloma Leuk. 16:264–268. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Mans S, Banz Y, Mueller BU and Pabst T:
The angiogenesis inhibitor vasostatin is regulated by neutrophil
elastase-dependent cleavage of calreticulin in AML patients. Blood.
120:2690–2699. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Cao H and Shockey JM: Comparison of TaqMan
and SYBR Green qPCR methods for quantitative gene expression in
tung tree tissues. J Agric Food Chem. 60:12296–12303. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
van Meerloo J, Kaspers GJ and Cloos J:
Cell sensitivity assays: the MTT assay. Methods Mol Biol.
731:237–245. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wu MH, Su Q, Cheng AL, Tan H and Song Y:
Experimental study of HL-60 cell differentiation induced by diallyl
disulfide. Zhong Guo Yao Li Xue Tong Bao. 19:319–322. 2003.(In
Chinese).
|
|
29
|
Ling H, Zhang LY, Su Q, Song Y, Luo ZY,
Zhou XT, Zeng X, He J, Tan H and Yuan JP: Erk is involved in the
differentiation induced by diallyl disulfide in the human gastric
cancer cell line MGC803. Cell Mol Biol Lett. 11:408–423. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
He J, Su Q, Huang WG, Xie HL, Liang SP,
Song Y, Xie JY, Zhou XT, Tan H, Zhao J and Wu MH: Proteomic initial
analysis of differentiation of human myeloid leukemia cells induced
by diallyl disulfide. FEBS J. 272:4402005.
|
|
31
|
Michalak M, Groenendyk J, Szabo E, Gold LI
and Opas M: Calreticulin, a multi-process calcium-buffering
chaperone of the endoplasmic reticulum. Biochem J. 417:651–666.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Walter RB, Appelbaum FR, Estey EH and
Bernstein ID: Acute myeloid leukemia stem cells and CD33-targeted
immunotherapy. Blood. 119:6198–6208. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ayadi L, Abid N, Makni S, Bahri I, Frikha
I and Sellami-Boudawara T: An unusual tumour of the lung.
Pathologica. 107:14–18. 2015.PubMed/NCBI
|
|
34
|
Jurcic JG: What happened to anti-CD33
therapy for acute myeloid leukemia? Curr Hematol Malig Rep.
7:65–73. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Arora M, Poe SL, Ray A and Ray P:
LPS-induced CD11b+Gr1(int)F4/80+ regulatory myeloid cells suppress
allergen-induced airway inflammation. Int Immunopharmacol.
11:827–832. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Heo SK, Noh EK, Yoon DJ, Jo JC, Koh S,
Baek JH, Park JH, Min YJ and Kim H: Rosmarinic acid potentiates
ATRA-induced macrophage differentiation in acute promyelocytic
leukemia NB4 cells. Eur J Pharmacol. 747:36–44. 2015. View Article : Google Scholar : PubMed/NCBI
|