Introduction
As a pervasive cancer worldwide, colorectal cancer
(CRC) is the third most commonly diagnosed cancer and the fourth
major cause of cancer-associated mortality, with an estimated 1.2
million new cancer cases and >600,000 mortalities annually
(1). CRC in China ranks among the 10
leading cancers in terms of incidence, with an occurrence rate
ranking sixth in humans. In 2010, there were 132,110 recorded
mortalities due to CRC, with mortality in men and women ranking
fifth and sixth among all cancers, respectively (2). Despite improvements in surgical
techniques, adjuvant chemotherapy and neoadjuvant radiotherapy, the
prognosis of advanced CRC remains unsatisfactory, with a 5-year
relative survival rate of only 10% in stage IV patients (3). Therefore, there is an essential
requirement for a n improved understanding of the biological
features and pathogenesis of CRC, which will be useful in the
investigation of novel treatments and prognostic markers for this
disease.
Forkhead box protein M1 (FoxM1), previously known as
MPP-2, FKHL-16 or Trident, belongs to the abundant family of
forkhead transcription factors, characterized by an evolutionary
conserved winged helix DNA-binding domain termed Forkhead box
(4). It has been acknowledged that
the expression of FoxM1 is associated with proliferating cells,
with levels particularly increasing at the entry to the S-phase of
the cell cycle and reaching a peak during the G2 and M phases, but
that it is absent in quiescent and terminally-differentiated cells
(5). Furthermore, it is ubiquitously
reported that FoxM1 plays a critical role in tumorigenesis,
including angiogenesis, invasion and metastasis, and that the
elevated expression levels of FoxM1 contribute to a poor prognosis
and metastasis in diverse tumors (5).
Emerging data has shown that FoxM1 is overexpressed in solid tumors
of the bladder, prostate, lungs, ovaries, breasts, liver, kidneys,
stomach, pancreas and colon (6).
Furthermore, it is not unexpected that the decreased expression of
FoxM1 when using FoxM1 short hairpin RNA leads to the reduction of
cell proliferation, migration and invasion in human tumor cell
lines (7). Specifically, the
expression of FoxM1 is significantly observed in samples from colon
cancer patients compared with lower expression in corresponding
adjacent normal tissues. Moreover, the enforced expression level of
FoxM1 drastically promotes the proliferation, development and
growth of colon tumors in FoxM1 transgenic mice (8). Nevertheless, the underlying molecular
regulatory mechanisms of FoxM1 in the promotion of CRC remain
unclear, and the roles of FoxM1 in the process of CRC development
and progression remain to be elucidated.
Caveolin-1 (Cav-1), a principal structural component
of caveolae and a 21- to 24-kDa integral membrane protein acting as
a scaffolding protein, is implicated in various cellular events by
interacting with diverse molecular complexes, and it may play an
irreplaceable role in the transformation and tumorigenesis of cells
(9). Notably, Cav-1 is defined with
an ambiguous role in cancer. The wealth of data available indicate
that the role of Cav-1 as a tumor suppressor or promoter is
dependent on the type of tissue; for example, Cav-1 acts as a tumor
suppressor in lung cancer, ovarian carcinoma and sarcomas, whereas
it act as a tumor promoter in prostate cancer and esophageal
squamous cell carcinomas (10). Based
on previous studies, compared with the increased expression of
Cav-1 in normal gastric tissues, there is a lower expression of
Cav-1 in paired gastric adenocarcinoma tissues; the results
obtained in gastric carcinoma cell lines are also consistent with
these observations, supporting the fact that Cav-1 may be a tumor
suppressor in gastric cancer (11).
With respect to the expression of Cav-1 in colon cancer,
controversy remains. One previous study stated that the expression
level of Cav-1 is decreased in human colon cancer, contributing
greatly to cellular migration and invasiveness (12). By contrast, other emerging evidence
supports the overexpression of Cav-1 in colon cancer in comparison
with normal colon tissues (13).
Considering all the aforementioned evidence, it is necessary and
essential for the expression and role of Cav-1 to be determined in
CRC.
E-cadherin, a single-span transmembrane
glycoprotein, is indispensable for the function of epithelial
adherens junctions through its interactions with adjacent
E-cadherin molecules secreted by contiguous cells (14). It has been widely illustrated in
different assays that the decreased expression of E-cadherin is
implicated in various tumors, such as pancreatic cancer, non-small
cell lung cancer and CRC, by acting in the process of
epithelial-mesenchymal transition (EMT) (15–17). Kong
et al verified that the elevated expression of FoxM1 led to
the loss expression of E-cadherin, which was significantly critical
to the acquisition of EMT in non-small cell lung cancer (16). As alluded to previously, emerging
clinical evidence shows that the downregulation of E-cadherin level
is correlated with a poor prognosis of CRC via acquisition of an
EMT phenotype (17). However, the
underlying precise regulating mechanisms of E-cadherin remain
unknown.
Based on the published literature, although a
diverse array of studies can be found on FoxM1, Cav-1 and
E-cadherin, respectively, there are no studies concerning the
associations among these proteins in CRC. The aim of the present
study was to determine the clinicopathological features of FoxM1,
Cav-1 and E-cadherin in CRC, and to investigate the potential
associations among these proteins.
Materials and methods
Human tissue samples
The expression of FoxM1, Cav-1 and E-cadherin in CRC
was analyzed employing a human CRC and normal tissue microarray (US
Biomax Inc., Rockville, MD, USA). The tissue microarray contained
30 samples of CRC, 8 samples of normal tissues, 2 samples of lung
metastases derived from CRC, 5 samples of lymph node metastases
derived from CRC and 3 samples of ovary metastases derived from
CRC. Each sample had 2 cores from the same specimen. The specimens
were obtained from 17 men and 13 women. The tumor stage
(tumor-node-metastasis classification) for the CRC was stage II for
7 samples, stage III for 17 samples and stage IV for 6 samples
(18). The differentiation for the
CRC was poorly-differentiated for 1 specimen,
moderately-differentiated for 18 specimens and well-differentiated
for 8 specimens (the remaining 3 specimens were of mucinous
carcinoma). The use of the tissue samples was approved by the
Institutional Review Board of Shanghai Jiaotong University
Affiliated First People's Hospital (Shanghai, China). The patients'
clinical information for the tissue microarray, including gender,
age, pathological diagnosis, clinical stage, histological grade and
metastasis, was provided by US Biomax.
Tissue immunohistochemistry
Standard immunohistochemical procedures were applied
using anti-FoxM1 rabbit polyclonal antibody (1:100 dilution;
catalog no. sc-500; Santa Cruz Biotechnology Inc., Dallas, TX,
USA), anti-Cav-1 mouse monoclonal antibody (1:200 dilution; catalog
no. 610406; BD Biosciences, Franklin Lakes, NJ, USA) and
anti-E-cadherin mouse monoclonal antibody (1:200 dilution; catalog
no. 610182; BD Biosciences). First, the 4-µm slices were dried in
an incubator at 60°C, then dewaxed in xylene, washed twice for 10
min each and rehydrated using a gradient from 100, 95, 85, 75 and
50% ethanol to pure distilled water with 5-min washes,
respectively. Antigen retrieval was performed by heating the slides
dipped in 10 mM sodium citrate (pH 6.0) at 95°C for 30 min.
Endogenous peroxidase activity was blocked by use of 3% hydrogen
peroxide for 10 min at room temperature. The sections were
incubated with 10% normal goat serum diluted with
phosphate-buffered saline for ~30 min at room temperature. The
slides were then incubated with the anti-FoxM1, anti-Cav-1 and
anti-E-cadherin antibodies, respectively, at 4°C overnight. The
specimens were later incubated with peroxidase-conjugated
antibodies at normal room temperature for 1 h. The secondary
antibody used for FoxM1 was peroxidase-conjugated Affinipure goat
anti-rabbit immunoglobulin (Ig)G(H+L) (1:500 dilution; catalog no.
SA00001-2). The secondary antibody used for Cav-1 and E-cadherin
was peroxidase-conjugated Affinipure goat anti-mouse IgG(H+L)
(1:500 dilution; catalog no. SA00001-1); All secondary antibodies
were purified from antisera by immunoaffinity chromatography using
antigens coupled to agarose beads. The slides were stained with
diaminobenzidine and then counterstained with Mayer's hematoxylin.
The tissue sections were dehydrated using 5-min washes with 50, 75,
85, 95 and 100% ethanol, respectively, and then washed twice with
xylene for 10 min each. The slides were finally mounted with
coverslips and assessed by microscopic examination (Axio Scope.A1;
Zeiss AG, Oberkochen, Germany).
Immunohistochemistry score
Immunohistochemical scoring was implemented by two
independent observers who were blinded to the clinical data.
Depending completely on the proportion of positive cells and the
intensity of cell staining in the field of view, the cell nuclei
and cytoplasmic staining of FoxM1, the cytoplasmic and membranous
staining of Cav-1, and the membranous staining of E-cadherin were
relatively divided into three groups: Negative, weak and strongly
positive. Collectively, the staining intensity of cells was
classified into four intensity scores: No staining, 0; light
staining, 1; moderate staining, 2; and dark staining, 3. The
percentage of positively-stained cells was classified into five
grades with percentage scores as follows: 10% staining, 0; 10–25%
staining, 1; 25–50% staining, 2; 50–75 staining, 3; and >75%
staining, 4. The staining positivity of FoxM1, Cav-1 and E-cadherin
was calculated using the following formula: Total score = intensity
score × percentage score. Based on the total score of each tissue,
results of ≤3, >3 to ≤6 and >6 were defined as negative,
weakly-positive and strongly-positive, respectively.
Statistical analysis
The Wilcoxon rank sum test was performed to
determine the significance of the association between the
expression of FoxM1/Cav-1/E-cadherin and clinical parameters. The
association between FoxM1 and Cav-1, FoxM1 and E-cadherin, and
Cav-1 and E-cadherin protein expression was analyzed utilizing
Spearman's test (r; P-value). In all tests, P<0.05 was
considered to indicate a statistically significant difference. SPSS
16.0 software (SPSS Inc., Chicago, IL, USA) was used for the
statistical analyses.
Results
Expression of FoxM1, Cav-1 and
E-cadherin in CRC tissues and surrounding normal tissues
The expression of FoxM1, Cav-1 and E-cadherin in CRC
tissues and paired normal tissues was evaluated by
immunohistochemistry. The expression of FoxM1 was observed mainly
in the nuclear compartment and cytoplasm, and the staining of FoxM1
was weakly- or strongly-positive in CRC in comparison with negative
in the adjacent normal tissue (Fig.
1A). A significant difference between CRC tissues and paired
normal tissues was observed (P=8.5416×10−6). Cav-1
expression was detected predominantly in the cytoplasm and cell
membrane, and the extent of Cav-1 staining in the CRC tissues was
weakly- or strongly-positive as compared with negative in the
paired normal tissues (P=9.6306×10−6), suggesting that
the expression of Cav-1 is upregulated in CRC (Fig. 1B). E-cadherin expression was primarily
found in the membrane of epithelial cells, and staining for
E-cadherin in the CRC and surrounding normal colorectal tissues was
found to be negative (or weakly-positive) and strongly-positive,
respectively (Fig. 1C). There was a
significant difference in the expression of E-cadherin between the
CRC tissues and paired normal tissues (P=0.0044).
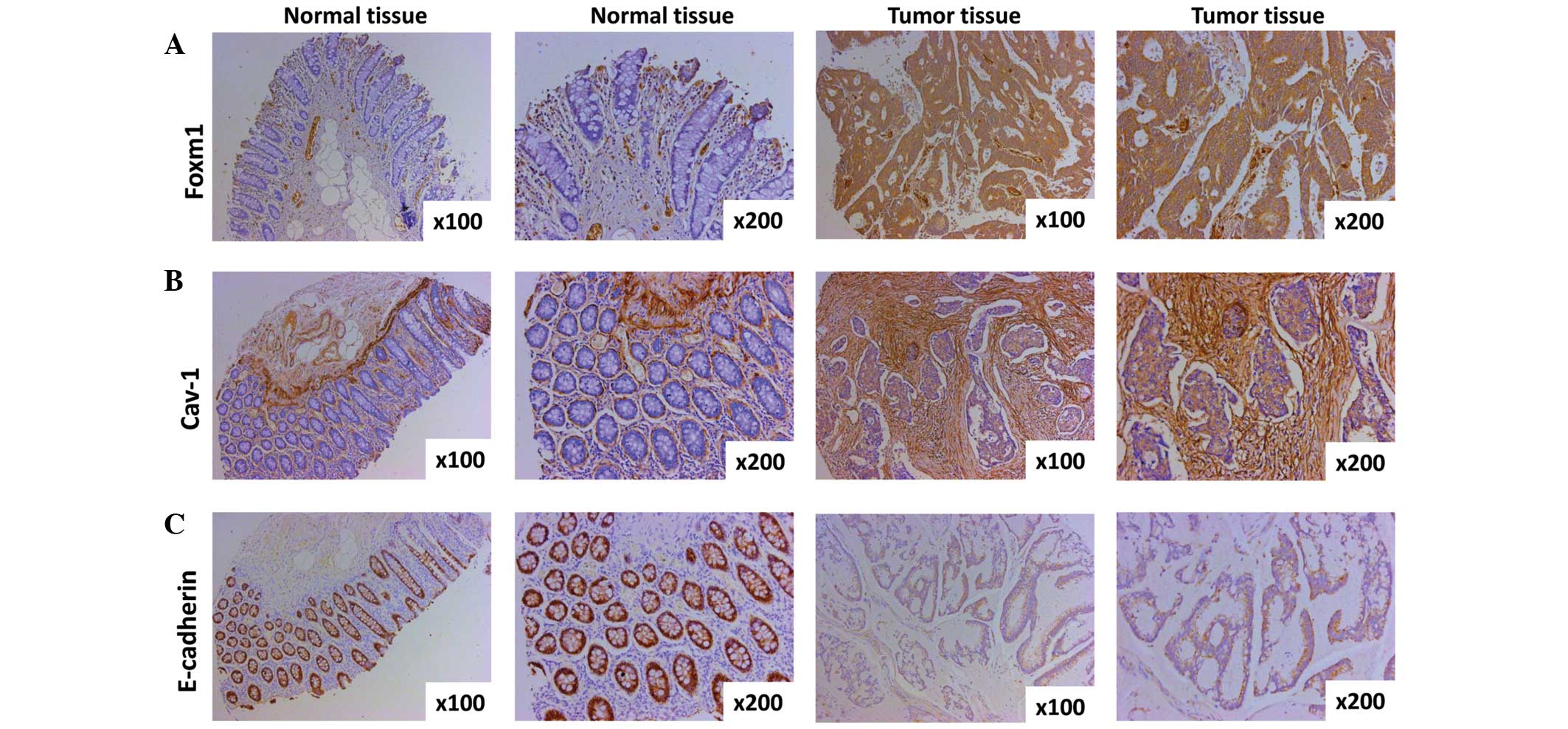 | Figure 1.Expression of FoxM1, Cav-1 and
E-cadherin in CRC specimens and normal tissues as detected by
immunohistochemistry. (A) Representative images of FoxM1 protein
expression in CRC tissues and surrounding normal tissues are shown
(magnification, ×100 and ×200). The negative immunostaining of
FoxM1 is displayed in the majority of the adjacent normal
colorectal tissue cells, whereas the immunostaining of FoxM1 in the
CRC tissues cells is strongly positive. (B) Cav-1 expression shown
by immunohistochemical staining utilizing Cav-1 antibody
(magnification, ×100 and ×200). The expression of Cav-1 in normal
colorectal tissue cells is negative, however, the majority of areas
of the CRC tissue cells are positive. (C) Typical images of
E-cadherin protein expression in CRC tissues and paired normal
tissue (magnification, ×100 and ×200). Almost all normal tissue
cells are strongly positive, yet the majority of CRC cells are
weakly positive or negative. CRC, colorectal cancer; FoxM1,
Forkhead box protein M1; Cav-1, caveolin-1. |
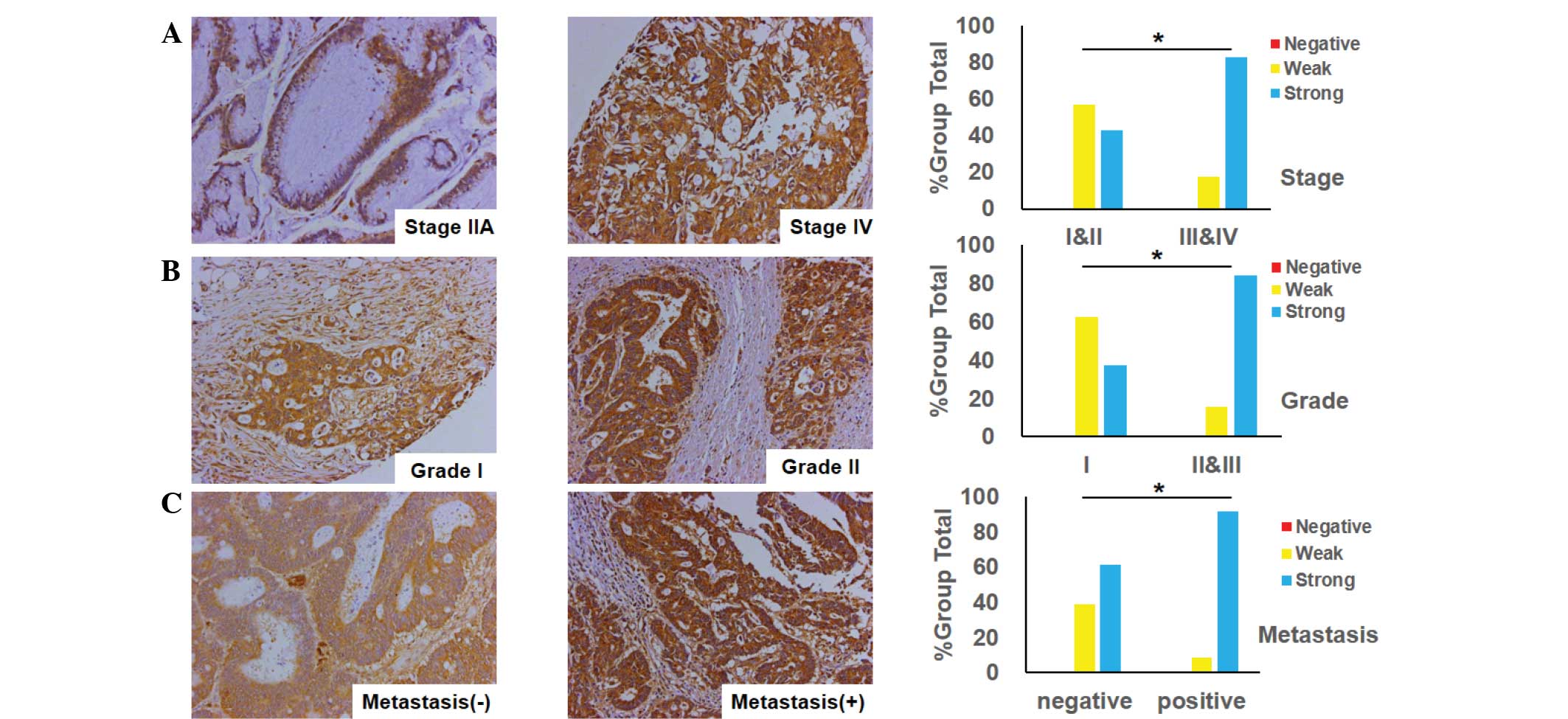
Association between FoxM1 expression
and clinicopathological features in CRC
The association between FoxM1 expression and the
clinicopathological features in CRC was investigated using the
immunohistochemistry tissues. The observations of the study
supplied unequivocal evidence that the expression of FoxM1 was
closely associated with tumor differentiation, tumor stage and its
metastasis. A positive association was found between FoxM1
expression and tumor stage, indicating that the expression of FoxM1
was higher in advanced-stage tissues (stages III and IV) in
contrast with early-stage tissues (stages I and II) (Fig. 2A). Additionally, when comparing
well-differentiated (grade I), and moderately- and
poorly-differentiated tumors (grades II and III), the FoxM1
expression was markedly increased in the moderately-differentiated
tumors (Fig. 2B), and there was a
significant difference between the two groups (grade I vs. grades
II and III; P=0.0355). Furthermore, the expression of FoxM1 was
much lower in non-metastatic samples than that in samples with
distant organ or lymph node metastasis (Fig. 2C).
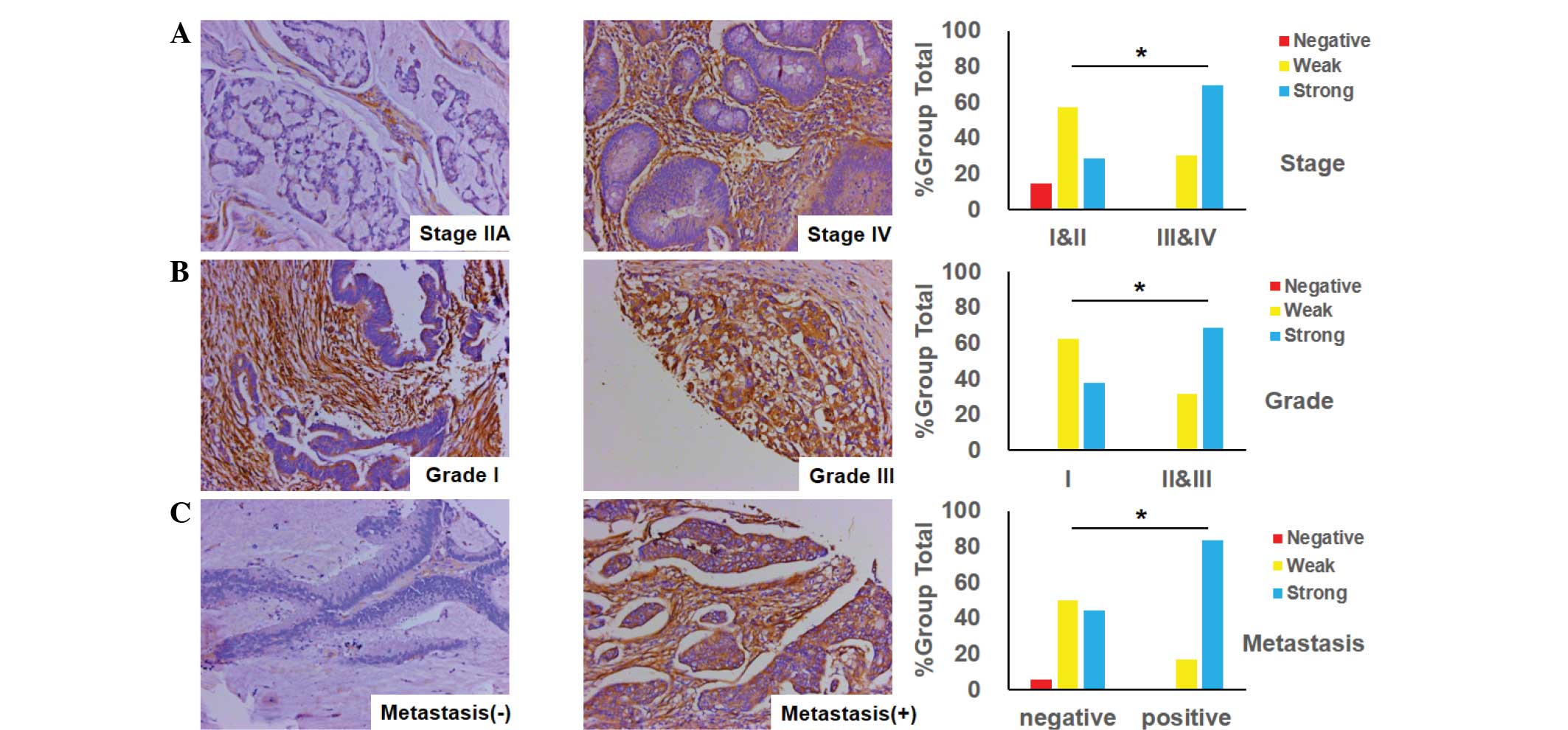
Association between Cav-1 expression
and clinicopathological features in CRC
The status of Cav-1 expression was an important
factor for CRC patients, as it was implicated in tumor
differentiation and stage, and its metastasis. Fig. 3A shows that the level of Cav-1
expression in advanced-stage CRC (stages III and IV) was markedly
positive, while conversely, the level of Cav-1 expression in
early-stage CRC (stages I and II) was weak. There was a significant
difference in Cav-1 expression between tumor stages (P=0.0477). In
addition, it was identified that the expression of Cav-1 in poorly-
and moderately-differentiated CRC tumors (grades II and III) was
stronger compared with the weak expression of Cav-1 in
well-differentiated tumors (grade I) (Fig. 3B). Moreover, it was found that the
expression of Cav-1 in specimens of CRC with lymph node or distant
organ metastasis was more elevated than that in other samples with
no metastasis (Fig. 3C).
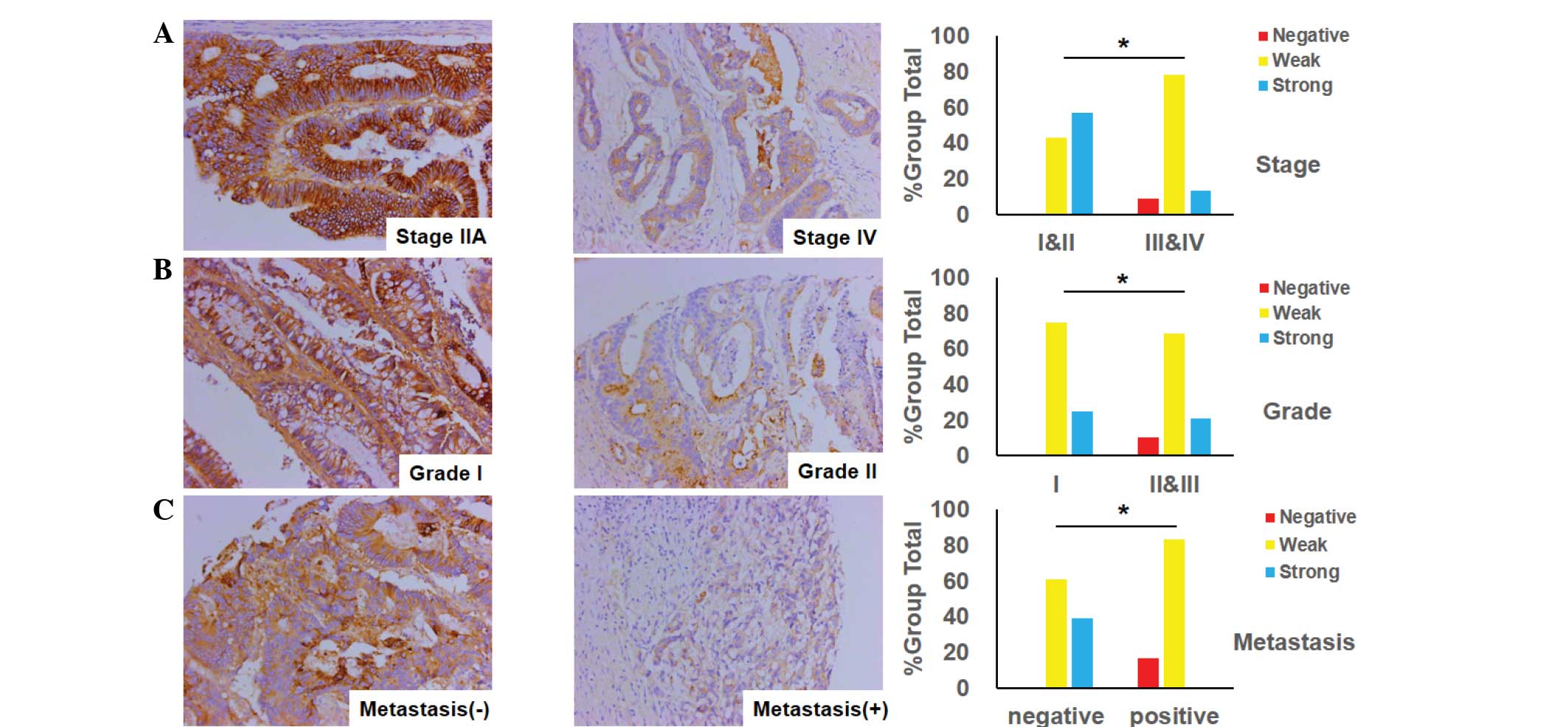
Association between the expression of
the epithelial marker E-cadherin and clinicopathological features
in CRC
The association between E-cadherin expression and
clinicopathological variables is presented in Fig. 4. As illustrated in Fig. 4A, the expression of E-cadherin in
advanced-stage CRC (stages III and IV) was observed to be more
significantly decreased than in early-stage CRC (stages I and II).
Similarly, the loss of expression of E-cadherin was more marked in
the poorly- or moderately-differentiated tumors of CRC than in the
well-differentiated tumors, as demonstrated in Fig. 4B. Consistent with the aforementioned
observation, the expression of E-cadherin in CRC with lymph node or
distant organ metastasis was attenuated more significantly than
that in non-metastatic tissues (Fig.
4C).
Correlation among FoxM1, Cav-1 and
E-cadherin expression
The strong or moderate staining of FoxM1 was
observed in 30 out of 30 CRC tissues. Notably, among the 30 tissues
strongly or moderately stained for FoxM1, there was 29 that were
strongly or moderately stained for Cav-1. Similarly, the results
showed that the expression of FoxM1 was strong or moderate in all
the tissues stained strongly- or moderately for Cav-1 (29 out of
29). These observations suggested that FoxM1 expression in the CRC
tissues had a positive correlation with Cav-1 expression (r=0.569,
P=0.0103) (Fig. 5A). With respect to
the correlation between FoxM1 and E-cadherin expression, it was
demonstrated that the increased expression of FoxM1 was always
accompanied by the decreased expression of E-cadherin in the same
CRC tissues Fig. 5A. On the other
side, the elevated expression of E-cadherin was concomitant with
the attenuated expression of FoxM1 in the same CRC tissues
(Fig. 5B). It was clearly shown that
a negative correlation existed between FoxM1 and E-cadherin
expression (r=−0.397, P=0.0299) (Fig.
5B). Consistent with these analogous observations, the
increased expression of Cav-1 was observed together with the
decreased expression of E-cadherin in corresponding CRC tissues.
Additionally, Cav-1 expression was attenuated in contrast with the
high E-cadherin expression in corresponding CRC specimens (Fig. 5C), indicating a negative association
between Cav-1 and E-cadherin (r=−0.362, P=0.0492).
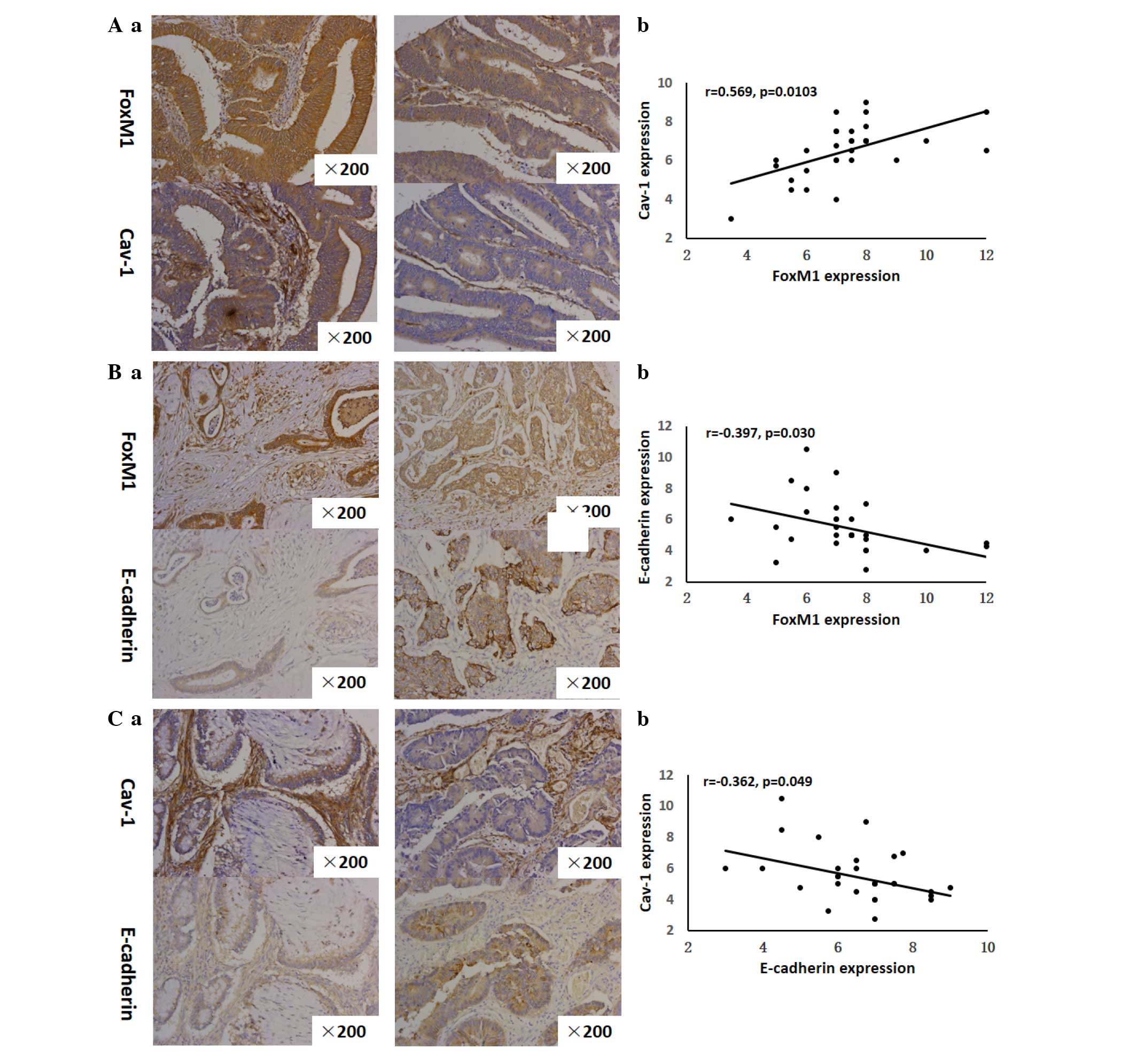 | Figure 5.Correlation between FoxM1 and Cav-1
expression, FoxM1 and E-cadherin expression, and Cav-1 and
E-cadherin expression. (Aa), FoxM1 and Cav-1 expression was
detected by immunohistochemistry using antibodies for FoxM1 and
Cav-1, respectively. Representative images of strongly-positive
FoxM1 and Cav-1 staining (left 2 panels) and moderate FoxM1 and
Cav-1 staining (right 2 panels) in CRC are shown (magnification,
×200). (Ab) Direct correlation between FoxM1 and Cav-1 expression
in CRC samples (n=30; Pearson's correlation test, r=0.533). Certain
dots on the graphs represent more than one sample (overlapped
scores). (Ba) FoxM1 and E-cadherin expression were detected by
immunohistochemistry using antibodies for FoxM1 and E-cadherin,
respectively. Representative images of strongly-positive FoxM1 and
weak E-cadherin staining (left 2 panels) and weakly-positive for
FoxM1 and strongly-positive E-cadherin staining (right 2 panels) in
CRC are shown (magnification, ×200). (Bb), Direct corelation
between FoxM1 expression and E-cadherin expression in CRC samples
(n=30; Pearson's correlation test, r=−0.356). Certain dots on the
graphs represent more than one sample (overlapped scores). (Ca)
Cav-1 and E-cadherin expression was detected by
immunohistochemistry using antibodies for Cav-1 and E-cadherin,
respectively. Representative images of strongly-positive Cav-1 and
weakly-positive E-cadherin staining (left 2 panels) and
weakly-positive Cav-1 and strongly-positive E-cadherin staining
(right 2 panels) in CRC are shown (magnification, ×200). (Cb)
Direct correlation between Cav-1 and E-cadherin expression in CRC
samples (n=30; Pearson's correlation test, r=−0.342). Certain dots
on the graphs represent more than one sample (overlapped scores).
CRC, colorectal cancer; FoxM1, Forkhead box protein M1; Cav-1,
caveolin-1. |
Discussion
The current study aimed to determine the critical
associations among FoxM1, Cav-1 and E-cadherin in the development
and progression of CRC. Based on the results, it was found that
there was a positive correlation between FoxM1 and Cav-1
expression. Conversely, there was a negative correlation between
FoxM1 and E-cadherin expression, and a negative correlation between
Cav-1 and E-cadherin. This novel clinical evidence suggested that
high FoxM1 expression, elevated Cav-1 expression and low E-cadherin
expression may be critical contributors to CRC pathogenesis and
aggressiveness. Collectively, considering the evidence presented, a
novel FoxM1/Cav-1/E-cadherin signaling pathway may exist in CRC,
directly impacting EMT.
A growing body of evidence has convincingly
demonstrated that FoxM1 plays an irreplaceable role in cancer
initiation by modulating the G1-S and G2-M
phases of the cell cycle, thus inducing cell proliferation and
progression (19). Transcription
molecules Skp2 and Cks1, components of the Skp-Cullin1-F-box
complex, are controlled by FoxM1, which contributes greatly to the
G1-S transition. Moreover, it is widely reported that
FoxM1 is essential for the G2-M phase transition through
regulators such as Cyclin A2, Cyclin B, survivin, centromere
protein A and Aurora A kinase, which are under control of FoxM1 at
the transcriptional level (20). It
is accepted worldwide that FoxM1 is not only involved in cell
proliferation, senescence and angiogenesis, which are indispensable
for tumorigenesis, but that it is also implicated in EMT and
metastasis, which are critical to tumor development and progression
(20). Numerous clinical studies and
experimental results have demonstrated that FoxM1, acknowledged as
an oncogene, contributes to diseases such as Ewing's sarcoma,
ovarian cancer, esophageal squamous cell carcinoma, gastric cancer
and CRC by regulating the protein expression essential for
angiogenesis, EMT and metastasis (7,21–24). One previous study provided convincing
evidence that the increased expression of FoxM1 was clearly
correlated with the poor prognosis of gastric cancer by
downregulating E-cadherin expression in the cells of gastric cancer
(21). In our previous study several
lines of investigation were followed to determine the crucial role
of FoxM1 in CRC, and the elevated expression of FoxM1 in in
vivo and in intro experiments promoted the migration,
invasion and metastasis of CRC. Conversely, the decreased
expression of FoxM1 via the transfection of cells with small
interfering RNA did the opposite (23). In the present study, it was
demonstrated that the expression of FoxM1 was markedly associated
with tumor stage (P=0.0209), grade (P=0.0355) and lymph node
metastasis (P=0.0038). It was previously illustrated by Yang et
al (25) that the overexpression
of FoxM1 was accompanied by the decreased expression of E-cadherin
in breast cancer, which was a pivotal step for the acquisition of
an EMT phenotype in tumor invasion and metastasis. An unexpected
finding in a study by Wierstra demonstrated that transcription
factor FoxM1c can directly bind to the murine and human E-cadherin
promoter as a novel target gene in tumors in vitro (26). In the present study, there was a
negative correlation between FoxM1 and E-cadherin in CRC (Fig. 5B), indicating that FoxM1 may
negatively regulate E-cadherin in the metastasis of CRC. Although
the underlying mechanism of FoxM1 regulating the tumorigenicity and
metastasis of cancer is unclear, all these observations support the
fact that FoxM1 may be a valuable prognostic hallmark or serve as a
novel therapeutic target in CRC.
It has been reported that Cav-1, also known as
caveolin or VIP21, which was first identified in the caveolin
family, plays an indispensable role in the physiological functions
of the cell by functioning in surface signaling, intracellular
cholesterol transport and endocytosis (27). Nevertheless, there is intense
controversy concerning the role of Cav-1, either as a tumor
suppressor or as a tumor promoter, in the process of cancer
development and progression (10).
One possible explanation accounting for the different roles of
Cav-1 is the tumor type. A growing body of studies has shown that
the decreased expression of Cav-1 can be recognized as a tumor
suppressor in ovarian carcinoma (28), gastric cancer (11) and breast carcinoma (29), meanwhile, Cav-1 overexpression as a
tumor promoter has been found in hepatocellular carcinoma (30), bladder cancer (31) and pancreatic cancer (15). Based on the available literature, the
role of Cav-1 in CRC appears to be highly inconsistent. One study
demonstrated that the overexpression of Cav-1 in colon cancer cells
markedly weakened proliferation, development, invasion, viability
and metastasis (32). Another study
published by Patlolla et al (33) demonstrated that Cav-1 was
overexpressed in the tissues and cell lines of human colon cancer.
Besides that, emerging novel evidence has revealed that the
expression of Cav-1 is a biphasic process in CRC, meaning that the
expression of Cav-1 is commonly downregulated in early-stage CRC
and then upregulated in advanced-stage CRC, manifesting the change
in the role of Cav-1 in CRC from a oncogene to an anti-oncogene
during tumor progression (34). In
the present study, it was illustrated that the expression of Cav-1
was elevated in CRC tissues in comparison with paired normal
tissues, and that the expression of Cav-1 was closely associated
with tumor stage (P=0.0477), grade (P=0.0189) and lymph node
metastasis (P=0.0112), which meant that it may act as a tumor
promoter during the development of the tumor. Consistent with these
observations, it was previously demonstrated that the elevated
expression of Cav-1 in bladder cancer was tightly correlated with
cell migration and invasiveness in contrast with the control group,
and the expression of Cav-1 was elevated in the process of
acquiring an EMT phenotype (31). Our
previous study verified that the staining of Cav-1 in pancreatic
cancer tissues was strongly positive compared with the paired
normal tissues, and when Cav-1 expression was elevated in the
pancreatic cancer cells, the abilities of cell migration and
metastasis were strengthened, E-cadherin expression was
significantly decreased and typical EMT morphological changes were
caused (15). In the present study,
it was found that the overexpression of Cav-1 in CRC was
concomitant with the decreased expression of E-cadherin, which
meant that Cav-1 may negatively regulate E-cadherin expression and
play significant roles in the progression of CRC.
Depletion of E-cadherin, one of the predominant
adhesive molecules of epithelial cells, plays a prominent role in
the invasion and metastasis of diverse neoplasms, including bladder
cancer, hepatocellular carcinoma and breast cancer, via acquisition
of the EMT phenotype (25,30,31). In
the current study, it was shown that E-cadherin expression was
decreased in CRC tissues, which is a key step in the process of
acquiring an EMT phenotype, and the expression of E-cadherin was
found to be closely associated with the clinicopathological
features of the disease (Fig. 4).
Diverse studies have revealed that there may be a range of
regulatory mechanisms of E-cadherin in different cancers. In
ovarian cancer, it was shown that transforming growth factor-β
played critical roles in tumorigenesis via decreasing E-cadherin
expression (35). Moreover, it was
demonstrated in non-small cell lung cancer that the expression of
E-cadherin was regulated by microRNA-10b (36). However, the specific regulatory
mechanism of E-cadherin in CRC remained unclear. Supporting data
published by Yang et al (25)
demonstrated that there was a correlation between FoxM1 and
E-cadherin in breast cancer. Additionally, another study showed
that Cav-1 can regulate the expression of E-cadherin in bladder
cancer (31). In our previous study,
it was verified that there was a novel FoxM1-Cav-1-E-cadherin
signaling pathway in pancreatic cancer (15). Therefore, it may be concluded that a
FoxM1-Cav-1-E-cadherin signaling pathway may exist in CRC.
Taken together, the results of the present study not
only showed the expression of FoxM1, Cav-1 and E-cadherin in CRC
tissues and normal tissues, but also revealed associations between
FoxM1, Cav-1 and E-cadherin expression and clinicopathological
features, respectively. More importantly, there may be a
FoxM1-Cav-1-E-cadherin signaling pathway in CRC, which may be a
promising therapeutic target for CRC patients.
Acknowledgements
The study was supported in part by grants from the
Shanghai Municipal Human Resources and Social Security Bureau (nos.
2012040 and 13PJD024), the Shanghai Health and Family Planning
Commission (no. XYQ2013092) and the Shanghai Municipal Science and
Technology Commission (no. 14411966800).
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen W, Zheng R, Zhang S, Zhao P, Zeng H,
Zou X and He J: Annual report on status of cancer in China, 2010.
Chin J Cancer Res. 26:48–58. 2014.PubMed/NCBI
|
|
3
|
Brenner H, Kloor M and Pox CP: Colorectal
cancer. Lancet. 383:1490–1502. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Halasi M and Gartel AL: Targeting FOXM1 in
cancer. Biochem Pharmacol. 85:644–652. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Alvarez-Fernandez M and Medema RH: Novel
functions of FoxM1: From molecular mechanisms to cancer therapy.
Front Oncol. 3:302013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Pilarsky C, Wenzig M, Specht T, Saeger HD
and Grützmann R: Identification and validation of commonly
overexpressed genes in solid tumors by comparison of microarray
data. Neoplasia. 6:744–750. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Christensen L, Joo J, Lee S, Wai D, Triche
TJ and May WA: FOXM1 is an oncogenic mediator in Ewing Sarcoma.
PloS One. 8:e545562013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yoshida Y, Wang IC, Yoder HM, Davidson NO
and Costa RH: The forkhead box M1 transcription factor contributes
to the development and growth of mouse colorectal cancer.
Gastroenterology. 132:1420–1431. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Schwencke C, Braun-Dullaeus RC, Wunderlich
C and Strasser RH: Caveolae and caveolin in transmembrane
signaling: Implications for human disease. Cardiovasc Res.
70:42–49. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Quest AF, Gutierrez-Pajares JL and Torres
VA: Caveolin-1: An ambiguous partner in cell signalling and cancer.
J Cell Mol Med. 12:1130–1150. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kannan A, Krishnan A, Ali M, Subramaniam
S, Halagowder D and Sivasithamparam ND: Caveolin-1 promotes gastric
cancer progression by up-regulating epithelial to mesenchymal
transition by crosstalk of signalling mechanisms under hypoxic
condition. Eur J Cancer. 50:204–215. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bender FC, Reymond MA, Bron C and Quest
AF: Caveolin-1 levels are down-regulated in human colon tumors, and
ectopic expression of caveolin-1 in colon carcinoma cell lines
reduces cell tumorigenicity. Cancer Res. 60:5870–5878.
2000.PubMed/NCBI
|
|
13
|
Fine SW, Lisanti MP, Galbiati F and Li M:
Elevated expression of caveolin-1 in adenocarcinoma of the colon.
Am J Clin Pathol. 115:719–724. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gumbiner BM: Regulation of
cadherin-mediated adhesion in morphogenesis. Nat Rev Mol Cell Biol.
6:622–634. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Huang C, Qiu Z, Wang L, Peng Z, Jia Z,
Logsdon CD, Le X, Wei D, Huang S and Xie K: A novel FoxM1-caveolin
signaling pathway promotes pancreatic cancer invasion and
metastasis. Cancer Res. 72:655–665. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kong FF, Qu ZQ, Yuan HH, Wang JY, Zhao M,
Guo YH, Shi J, Gong XD, Zhu YL, Liu F, et al: Overexpression of
FOXM1 is associated with EMT and is a predictor of poor prognosis
in non-small cell lung cancer. Oncol Rep. 31:2660–2668.
2014.PubMed/NCBI
|
|
17
|
He X, Chen Z, Jia M and Zhao X:
Downregulated E-cadherin expression indicates worse prognosis in
Asian patients with colorectal cancer: Evidence from meta-analysis.
PloS One. 8:e708582013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Edge S, Byrd DR, Compton CC, Fritz AG,
Greene FL and Trotti A: AJCC Cancer Staging Manual. 7th. Springer;
NY: 2010
|
|
19
|
Huang C, Du J and Xie K: FOXM1 and its
oncogenic signaling in pancreatic cancer pathogenesis. Biochim
Biophys Acta. 1845:104–116. 2014.PubMed/NCBI
|
|
20
|
Lam EWF, Brosens JJ, Gomes AR and Koo CY:
Forkhead box proteins: Tuning forks for transcriptional harmony.
Nat Rev Cancer. 13:482–495. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Miao L, Xiong X, Lin Y, Cheng Y, Lu J,
Zhang J and Cheng N: Down-regulation of FoxM1 leads to the
inhibition of the epithelial-mesenchymal transition in gastric
cancer cells. Cancer Genet. 207:75–82. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Takata A, Takiguchi S, Okada K, Takahashi
T, Kurokawa Y, Yamasaki M, Miyata H, Nakajima K, Mori M and Doki Y:
Clinicopathological and prognostic significance of FOXM1 expression
in esophageal squamous cell carcinoma. Anticancer Res.
34:2427–2432. 2014.PubMed/NCBI
|
|
23
|
Li D, Wei P, Peng Z, Huang C, Tang H, Jia
Z, Cui J, Le X, Huang S and Xie K: The critical role of
dysregulated FOXM1-PLAUR signaling in human colon cancer
progression and metastasis. Clin Cancer Res. 19:62–72. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Huang C, Xie D, Cui J, Li Q, Gao Y and Xie
K: FOXM1c promotes pancreatic cancer epithelial-to-mesenchymal
transition and metastasis via upregulation of expression of the
urokinase plasminogen activator system. Clin Cancer Res.
20:1477–1488. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yang C, Chen H, Tan G, Gao W, Cheng L,
Jiang X, Yu L and Tan Y: FOXM1 promotes the epithelial to
mesenchymal transition by stimulating the transcription of Slug in
human breast cancer. Cancer Lett. 340:104–112. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wierstra I: The transcription factor
FOXM1c binds to and transactivates the promoter of the tumor
suppressor gene E-cadherin. Cell Cycle. 10:760–766. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Williams TM and Lisanti MP: The caveolin
proteins. Genome Biol. 5:2142004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sanna E, Miotti S, Mazzi M, De Santis G,
Canevari S and Tomassetti A: Binding of nuclear caveolin-1 to
promoter elements of growth-associated genes in ovarian carcinoma
cells. Exp Cell Res. 313:1307–1317. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Martins D, Beça FF, Sousa B, Baltazar F,
Paredes J and Schmitt F: Loss of caveolin-1 and gain of MCT4
expression in the tumor stroma: Key events in the progression from
an in situ to an invasive breast carcinoma. Cell Cycle.
12:2684–2690. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Müller R, Gai X, Lu Z, Tu K, Liang Z and
Zheng X: Caveolin-1 Is up-regulated by GLI1 and contributes to
GLI1-driven EMT in hepatocellular carcinoma. PloS one.
9:e845512014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Liang W, Hao Z, Han JL, Zhu DJ, Jin ZF and
Xie WL: CAV-1 contributes to bladder cancer progression by inducing
epithelial-to-mesenchymal transition. Urologic Oncol. 32:855–863.
2014. View Article : Google Scholar
|
|
32
|
Nimri L, Barak H, Graeve L and Schwartz B:
Restoration of caveolin-1 expression suppresses growth,
membrane-type-4 metalloproteinase expression and
metastasis-associated activities in colon cancer cells. Mol
Carcinog. 52:859–870. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
33
|
Patlolla JM, Swamy MV, Raju J and Rao CV:
Overexpression of caveolin-1 in experimental colon adenocarcinomas
and human colon cancer cell lines. Oncol Rep. 11:957–963.
2004.PubMed/NCBI
|
|
34
|
Ha TK, Her NG, Lee MG, Ryu BK, Lee JH, Han
J, Jeong SI, Kang MJ, Kim NH, Kim HJ, et al: Caveolin-1 increases
aerobic glycolysis in colorectal cancers by stimulating
HMGA1-mediated GLUT3 transcription. Cancer Res. 72:4097–4109. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Gao J, Zhu Y, Nilsson M and Sundfeldt K:
TGF-b isoforms induce EMT independent migration of ovarian cancer
cells. Cancer Cell Int. 14:722014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhang J, Xu L, Yang Z, Lu H, Hu D, Li W,
Zhang Z, Liu B and Ma S: MicroRNA-10b indicates a poor prognosis of
non-small cell lung cancer and targets E-cadherin. Clin Transl
Oncol. 17:209–214. 2015. View Article : Google Scholar : PubMed/NCBI
|