Introduction
Gastric cancer is one of the most common types of
cancer worldwide, particularly in East Asian populations. However,
while the precise mechanism that underlies gastric carcinogenesis
is not fully understood, multiple genetic alterations in tumor
suppression genes, oncogenes, cell adhesion molecules, the growth
factor/receptor system and DNA repair genes have been observed to
be involved in this process. Among the molecular markers that
exhibit variable expression levels, the expression of early growth
response-1 (EGR-1), which is considered a tumor suppressor gene in
a number of cell types, was observed to be elevated in gastric
cancer tissues (1). EGR-1 is
functionally implicated in numerous critical biological processes,
including inflammation, cell proliferation, differentiation,
vascular wound response and cancer progression (2). Furthermore, it has been suggested that
EGR-1 may play a significant role in carcinogenesis and cancer
progression in the stomach via the alteration of tumor cell
behaviors, including migration and invasion (3). However, the effects of EGR-1 expression
on human gastric cancer progression, particularly in precancerous
lesions, have yet to be investigated. The aims of this study were
to measure the expression levels of EGR-1 in the target mucosa of
patients with early gastric cancer and precancerous lesions, and to
evaluate whether EGR-1 affects oncogenic phenotypes of human
gastric cancer cells.
Materials and methods
Patients and specimens
The expression levels of EGR-1 protein were
evaluated in 19 gastric tissues harvested from patients with
chronic gastritis (n=6), low-grade dysplasia (LGD; n=6), high-grade
dysplasia (HGD; n=4) or adenocarcinoma (ADC; n=3) by endoscopic
biopsy. All patients had undergone endoscopic mucosal resection or
endoscopic submucosal dissection for known dysplasia or T1 ADC at
Chonnam National University Hospital, Korea, between July and
December 2014. All resected specimens were evaluated by
histological examination on the basis of the Vienna classification
system (4). Patients were considered
to be infected with Helicobacter pylori if the results of at
least one of three diagnostic tests (rapid urease test, histology
results and [13C]-urea breath test) were positive. All
specimens were collected with the informed consent of patients, and
the study was approved by the ethics committee of Chonnam National
University Hospital (IRB no. CNUH-2014-144).
Cell culture and transfection
Human gastric carcinoma AGS cells were purchased
from the American Type Culture Collection (Manassas, VA, USA).
Cells were cultured at 37°C in a 5% CO2 atmosphere with
Dulbecco's modified Eagle's medium (Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) supplemented with 10%
heat-inactivated fetal bovine serum, 1% penicillin-streptomycin and
0.2% gentamycin (50 mg/ml solution; Gibco; Thermo Fisher
Scientific, Inc.). For transfection studies, cells were seeded on
six-well plates and incubated overnight. Adhered cells were then
transiently transfected with the pcDNA3.1/EGR-1 (I293F) expression
vector (provided by Professor Yong Han Lee), which expresses a
dominant active form of EGR-1 [(DA)-EGR-1].
MTT assay
Cell viability was evaluated using the
3-(4,5-dimethylthiazole-2-yl)-2,5-diphenyl tetrazolium bromide
(MTT) assay. Briefly, AGS cells were seeded in 96-well plates and
allowed to adhere overnight at 37°C. Following incubation, 20 µl
MTT solution was added to each well and the plates were incubated
for 4 h at 37°C with 5% CO2. The MTT solution was then
removed, and 200 µl dimethyl sulfoxide was added to each well.
Viability was quantified by measuring the optical density of each
well at a wavelength of 550 nm (OD550).
Cell migration assay
To assess the effects of EGR-1 expression on cell
migration, cell migration assays were performed in six-well plates
(Corning® 3516 Costar® six-well × 16.8 ml
flat bottom cell culture microplate; Corning Life Sciences,
Tewksbury, MA, USA). To create wound gaps, AGS cells were seeded on
the culture plate inserts, and gently removed using sterile
tweezers after 0, 6, 12, 24 and 48 h of incubation. The progress of
wound closure was monitored and photographed using an inverted
microscope. Wound size was measured at six different positions on
the photographs, and the average wound size at each position was
calculated.
Evaluation of EGR-1 expression
The EGR-1 protein expression levels were measured
within tissues from patients with normal mucosa (n=6), LGD (n=6),
HGD (n=4) or ADC (n=4) using a human enzyme-linked immunosorbent
assay (ELISA) kit (Cloud-Clone Corp., Houston, TX, USA), according
to the manufacturer's instructions. The detection limit of the
assay for EGR-1 was 0.1 ng/ml. All measurements were performed in
duplicate.
For immunohistochemical staining, 4-µm-thick
sections were generated from the 19 gastric tissues from subjects
with normal mucosa, LGD, HGD or ADC. The tissue sections were then
deparaffinized, rehydrated and retrieved with retrieval buffer.
Tissues were treated with peroxidase-blocking solution (Dako,
Glostrup, Denmark) to block endogenous peroxidase activity, and
incubated at 4°C overnight with polyclonal rabbit anti-human EGR-1
antibody (diluted 1:100). After washing with Tris-buffered saline
containing Tween-20 (TBST), tissues were stained using the Dako
Real™ Envision HRP/DAB detection system (Dako). Stained tissues
were viewed and photographed using a light microscope. The
immunoreactivity of each sample (intensity, total area and pattern
of immunostaining) was evaluated independently by two observers who
had no knowledge of the clinical outcomes. In cases where there was
a discrepancy, a consensus was reached after further evaluation.
The staining intensity was graded using the following numerical
scale: 0, no staining of cancer cells; 1, weak staining; 2,
moderate staining; and 3, strong staining. Specifically, we
analyzed the staining intensity within the nucleus and cytoplasm,
respectively. The overall scores were calculated as the sum of the
staining intensity in the nuclei and the cytoplasm. As such, these
scores could range from 0 to 6. Specimens with a score >3 were
regarded as EGR-1 expression-positive, while those with a score ≤3
were considered EGR-1 expression-negative.
Western blot analysis
Cells were lysed in RIPA buffer (1 M Tris-HCl, 150
mM NaCl, 1% Triton X-100 and 2 mM EDTA) containing a phosphatase
inhibitor and a protease inhibitor cocktail (Sigma-Aldrich, St.
Louis, MO, USA). The cell lysates (20 µg protein) were then
separated by 10% sodium dodecyl sulphate-polyacrylamide gel
electrophoresis and electrophoretically transferred to
polyvinylidene fluoride membranes. After blocking with 5% skimmed
milk in TBST for 1 h, membranes were blotted with EGR-1- or
β-actin-specific antibodies (Cell Signaling Technology, Danvers,
MA, USA) at room temperature for 3 h. After washing three times
with TBST, membranes were incubated with horse-radish peroxidase
(HRP)-conjugated anti-rabbit immunoglobulin secondary antibodies
(1:2,000 dilution; Cell Signaling Technology) at room temperature
for 1 h. After three washes with TBST, membranes were incubated
with enhanced chemiluminescence reagent, and protein bands were
visualized using a Fuji LAS-3000 image analyzer (Fuji Film, Tokyo,
Japan).
Statistical analyses
Differences in EGR-1 protein levels and in the
intensity values obtained from the EGR-1 immunohistochemical
staining analyses were evaluated using a non-parametric
Kruskal-Wallis test. Chi-square tests (linear by linear
association) were used to compare the nuclear and cytoplasmic
expression of EGR-1 according to the histological grades.
Associations between EGR-1 expression and clinical factors were
evaluated by Chi-square tests and Mann-Whitney U tests. The results
of the cell viability (MTT) and migration assays were assessed
using two-tailed Student's t-tests. P<0.05 was considered to
indicate a statistically significant difference. All analyses were
conducted using Statistical Package for Social Sciences (SPSS/PC)
20.0 software (SPSS Statistics Inc., Chicago, IL, USA).
Results
Expression of EGR-1 protein is
associated with gastric carcinogenesis
We first analyzed the EGR-1 protein expression
levels in gastric precancerous lesions and cancer tissues using an
EGR-1-specific ELISA. A total of 19 patients (median age, 65 years;
10 males) were included in this study. The baseline demographic
data of the patients are summarized in Table I. Nine (47.4%) of the 19 patients were
infected with H. pylori. The results of these assays
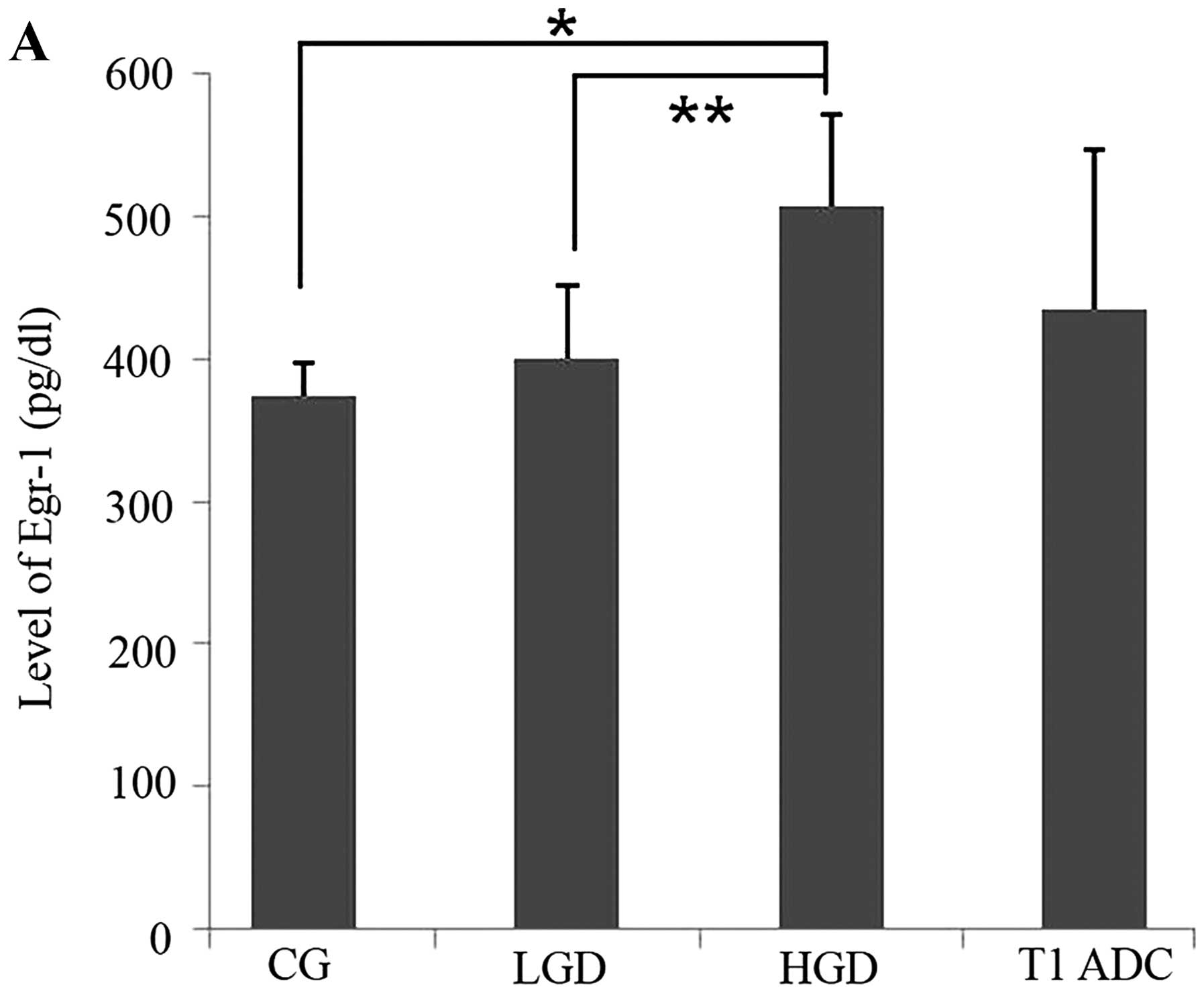
indicated that there was a positive correlation between EGR-1
expression and the histological grade (P=0.016, Fig. 1A). In addition, we assessed EGR-1
expression by immunohistochemical staining of formalin-fixed,
paraffin-embedded tissue blocks obtained from the 19 enrolled
patients (Fig. 1B). Nuclear and
cytoplasmic expression of EGR-1 increased according to the
histological grade (P for trends=0.003 and 0.003, respectively),
and there was a positive association between the sum of the nuclear
and cytoplasmic EGR-1 expression levels and the histological grade
(P=0.003, Fig. 1C). When the
expression of EGR-1 was classified as positive or negative,
according to the immunohistochemistry score, there was a
statistically significant association between the positive
expression of EGR-1 and the histological grade (P for trend=0.001).
Conversely, gender, age, presence of H. pylori and tumor
location were not associated with EGR-1 expression (Table II).
 | Table I.Baseline demographic data for the
patients enrolled in study. |
Table I.
Baseline demographic data for the
patients enrolled in study.
|
| Median age
(range) | Gender (M:F) | Positive for H.
pylori (%) |
|---|
| Chronic gastritis
(n=6) | 39 (28–71) | 3:3 | 3
(50) |
| LGD (n=6) | 67 (54–76) | 4:2 | 3
(50) |
| HGD (n=4) | 72 (48–80) | 1:3 | 0 (0) |
| T1 ADC (n=3) | 59 (44–68) | 2:1 |
3 (100) |
| Table II.Correlation between EGR-1 expression
and clinicopathological features. |
Table II.
Correlation between EGR-1 expression
and clinicopathological features.
|
| EGR-1−
n=6 | EGR-1+
n=13 | P-value |
|---|
| Age (mean ± SD) | 45.3±19.6 | 64.6±10.8 | 0.062 |
| Gender (M:F) |
3:3 |
7:6 | 0.999 |
| Positive H.
pylori | 3 (50.0%) | 6 (46.2%) | 0.999 |
| Location of
specimen |
|
| 0.459 |
| Lower
1/3 | 5 (83.3%) | 7 (53.8%) |
|
| Mid
1/3 | 0 (0%) | 4 (30.8%) |
|
| Upper
1/3 | 1 (16.7%) | 2 (15.4%) |
|
| Pathology |
|
| 0.001 |
| Chronic
gastritis | 6 (100%) | 0 (0%) |
|
| LGD | 0 (0%) | 6 (46.2%) |
|
| HGD | 0 (0%) | 4 (30.8%) |
|
| T1
ADC | 0 (0%) | 3 (23.1%) |
|
EGR-1 overexpression promotes cell
proliferation in gastric cancer cells
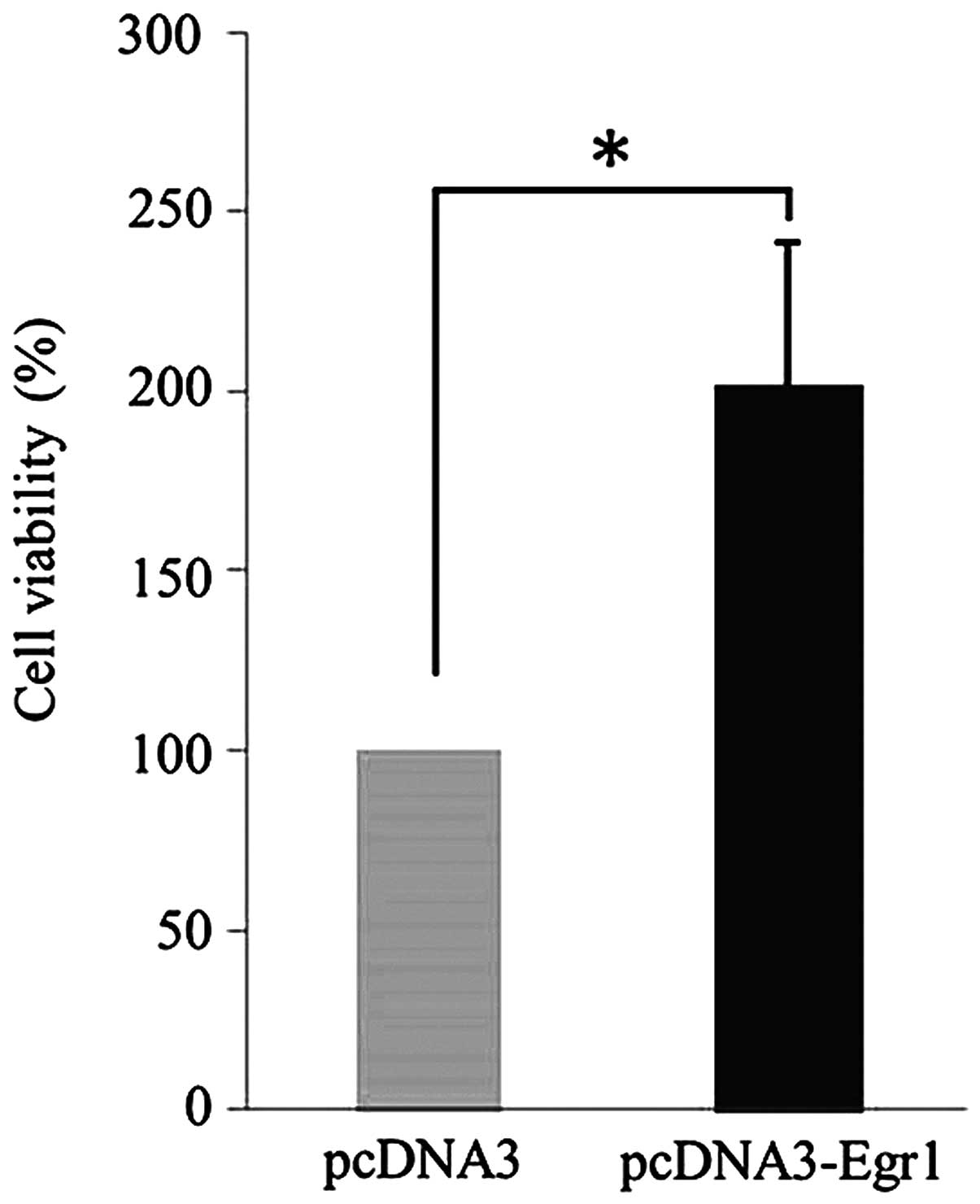
MTT assays were used to assess the potential effects
of EGR-1 expression on the proliferation of human gastric cancer
cells. Compared with cells transfected with the empty vector, AGS
cells transfected with the pcDNA3.1/EGR-1 (I293F) expression
vector, which encodes (DA)-EGR-1, exhibited enhanced rates of
proliferation (P=0.021, Fig. 2).
Overexpression of EGR-1 promotes the
migration of gastric cancer cells
We examined whether EGR-1 expression promotes the
migration of gastric cancer cells using a wound-healing assay.
Compared with the cells transfected with the empty vector, AGS
cells transfected with the EGR-1 expression vector exhibited
significant reductions in wound sizes (Fig. 3A). Furthermore, quantitative analyses
revealed that overexpression of EGR-1 stimulated gastric cancer
cell migration (Fig. 3B).
EGR-1 expression promotes the
phosphorylation of p38 MAPK and AKT
While transfection of AGS cells with the EGR-1
expression vector resulted in increased levels of AKT and p38 MAPK
phosphorylation, this EGR-1 overexpression had no effect on the
total expression levels of AKT and MAPK (Fig. 4). Therefore, these findings indicate
that EGR-1 promotes the activation of p38 MAPK and AKT.
Discussion
The principal findings of this study were as
follows: i) expression of EGR-1 protein, as detected by ELISA, was
associated with gastric carcinogenesis; ii) there was an increase
in the nuclear and cytoplasmic levels of EGR-1 expression in
accordance with the histological grades; iii) EGR-1 overexpression
promoted cell proliferation and migration in gastric cancer cells
in vitro; and iv) overexpression of EGR-1 resulted in
increased phosphorylation of p38 MAPK and AKT in vitro.
EGR-1 functions as either a growth-promoting factor
or as a tumor suppressor (5–8), and previous studies demonstrated a
positive correlation between the expression of EGR-1 and gastric
cancer progression during the advanced stages (3,9). The
expression of EGR-1 is rapidly induced by a number of extracellular
stimuli, including growth factors, cytokines, hypoxia,
injury-related stimuli and bacterial toxins (2,10). Keates
et al demonstrated that exposure to cag+ H. pylori
promoted EGR-1 gene expression in gastric epithelial cells,
and that this upregulation may contribute to cancer pathogenesis
(11). Notably, gastric
carcinogenesis is a multifactorial process for which H.
pylori infection is the most significant risk factor (12). Since constant exposure of the gastric
epithelium to H. pylori may cause frequent cellular
activation and upregulation of EGR-1, we speculated that EGR-1
expression may play a role in the progression of gastric cancer,
not only during advanced stages (including invasion or distant
metastasis), but also during early stages (including the
dysplasia-carcinoma carcinogenic pathway). Gastric cancers are
classified histologically into two main types: intestinal and
diffuse. Intestinal-type gastric carcinomas are considered to be
derived from gastric mucosal cells. These carcinomas are also
histologically differentiated based on well-defined glandular
structures that exhibit an expanding growth pattern and which
develop through well-characterized sequential stages, including
chronic gastritis, atrophy, intestinal metaplasia and dysplasia,
which is an atypical change in the epithelium that is considered to
be precancerous (13). Moreover,
there is abundant evidence suggesting that most gastric cancers,
particularly the intestinal subtype, develop via an intestinal
metaplasia-dysplasia-carcinoma carcinogenic pathway (14).
Currently, there are no published studies that have
examined the correlation between EGR-1 expression and precancerous
gastric lesions. A previous study, however, detected high levels of
EGR-1 mRNA and protein expression in precancerous lesions of the
esophagus (10). Likewise, in this
study, we observed that EGR-1 protein expression levels were
elevated in precancerous stomach lesions (LGD and HGD).
Furthermore, compared with tissues obtained from healthy patients,
we observed an increase in the nuclear and cytoplasmic expression
of EGR-1 in gastric tissues harvested from patients with
precancerous lesions by immunohistochemistry. These findings
therefore indicate that EGR-1 may be involved in the progression of
intestinal-type gastric carcinomas at stages as early as the
formation of precancerous lesions, including dysplasia.
To examine the functional significance of EGR-1 in
human gastric cancer cells, we transfected a gastric cancer cell
line with either an EGR-1 overexpression vector or a control
vector. A previous study demonstrated that EGR-1 expression
resulted in increased tumor cell migration and invasion by
promoting β-catenin expression in human gastric cancer cells
(9). In addition, Myung et al
demonstrated that a knockdown of EGR-1 expression in human gastric
cancer cell lines resulted in decreased rates of tumor cell
invasion and migration (3).
Consistent with these findings, our results indicate that EGR-1
overexpression promotes cell proliferation and migration in gastric
cancer cells. Additionally, they demonstrate that EGR-1 expression
promotes the phosphorylation of p38 MAPK and AKT. Thus, EGR-1
activation due to exogenous stimuli, including H. pylori
infection or a high salt diet, may contribute to gastric epithelial
cell hyperproliferation leading to gastric dysplasia. Based on the
American Joint Committee on Cancer and the International Union
Against Cancer (AJCC/UICC) staging, T1 gastric cancers include
tumors that invade the lamina propria, muscularis mucosae or
submucosa (15), and are defined as
unequivocal neoplastic epithelia that are confined to the basement
membrane. Dysplasia is categorized as either low or high grade,
depending on the degree of cytological and architectural atypia of
the pits and surface epithelium (16,17).
Therefore, EGR-1 activation may contribute to the migration of
neoplastic cells, leading to the progression of dysplasia to T1
gastric cancer.
In conclusion, our findings demonstrate that EGR-1
may be involved in the early stages, as well as the latter stages,
of gastric carcinogenesis through the alteration of tumor cell
behaviors.
Acknowledgements
This study was supported by a grant from Chonnam
National University Hospital Biomedical Research Institute
(CRI14014-1).
References
|
1
|
Kobayashi D, Yamada M, Kamagata C, Kaneko
R, Tsuji N, Nakamura M, Yagihashi A and Watanabe N: Overexpression
of early growth response-1 as a metastasis-regulatory factor in
gastric cancer. Anticancer Res. 22:3963–3970. 2002.PubMed/NCBI
|
|
2
|
Thiel G and Cibelli G: Regulation of life
and death by the zinc finger transcription factor EGR-1. J Cell
Physiol. 193:287–292. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Myung E, Park YL, Kim N, Chung CY, Park
HB, Park HC, Myung DS, Kim JS, Cho SB, Lee WS and Joo YE:
Expression of early growth response-1 in human gastric cancer and
its relationship with tumor cell behaviors and prognosis. Pathol
Res Pract. 209:692–699. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Schlemper RJ, Riddell RH, Kato Y, Borchard
F, Cooper HS, Dawsey SM, Dixon MF, Fenoglio-Preiser CM, Fléjou JF,
Geboes K, et al: The vienna classification of gastrointestinal
epithelial neoplasia. Gut. 47:251–255. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
deBelle I, Huang RP, Fan Y, Liu C, Mercola
D and Adamson ED: p53 and Egr-1 additively suppress transformed
growth in ht1080 cells but Egr-1 counteracts p53-dependent
apoptosis. Oncogene. 18:3633–3642. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yu J, Zhang SS, Saito K, Williams S,
Arimura Y, Ma Y, Ke Y, Baron V, Mercola D, Feng GS, et al: Pten
regulation by AKT-EGR1-ARF-PTEN axis. EMBO J. 28:21–33. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kim J, Lee YH, Kwon TK, Chang JS, Chung KC
and Min DS: Phospholipase D prevents etoposide-induced apoptosis by
inhibiting the expression of early growth response-1 and
phosphatase and tensin homologue deleted on chromosome 10. Cancer
Res. 66:784–793. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mahalingam D, Natoni A, Keane M, Samali A
and Szegezdi E: Early growth response-1 is a regulator of
DR5-induced apoptosis in colon cancer cells. Br J Cancer.
102:754–764. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sun T, Tian H, Feng YG, Zhu YQ and Zhang
WQ: Egr-1 promotes cell proliferation and invasion by increasing
β-catenin expression in gastric cancer. Dig Dis Sci. 58:423–430.
2013.PubMed/NCBI
|
|
10
|
Wu MY, Liang YR, Wu XY and Zhuang CX:
Relationship between Egr-1 gene expression and apoptosis in
esophageal carcinoma and precancerous lesions. World J
Gastroenterol. 8:971–975. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Keates S, Keates AC, Nath S, Peek RM Jr
and Kelly CP: Transactivation of the epidermal growth factor
receptor by cag+ helicobacter pylori induces upregulation of the
early growth response gene Egr-1 in gastric epithelial cells. Gut.
54:1363–1369. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Conteduca V, Sansonno D, Lauletta G, Russi
S, Ingravallo G and Dammacco F: H. pylori infection and gastric
cancer: state of the art (review). Int J Oncol. 42:5–18.
2013.PubMed/NCBI
|
|
13
|
Yuasa Y: Control of gut differentiation
and intestinal-type gastric carcinogenesis. Nat Rev Cancer.
3:592–600. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Correa P: Human gastric carcinogenesis: a
multistep and multifactorial process-first American Cancer Society
award lecture on cancer epidemiology and prevention. Cancer Res.
52:6735–6740. 1992.PubMed/NCBI
|
|
15
|
Edge SB and Compton CC: The American joint
committee on cancer: The 7th edition of the AJCC cancer staging
manual and the future of TNM. Ann Surg Oncol. 17:1471–1474. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Rugge M, Correa P, Dixon MF, Hattori T,
Leandro G, Lewin K, Riddell RH, Sipponen P and Watanabe H: Gastric
dysplasia: the Padova international classification. Am J Surg
Pathol. 24:167–176. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Stolte M: The new vienna classification of
epithelial neoplasia of the gastrointestinal tract: advantages and
disadvantages. Virchows Arch. 442:99–106. 2003.PubMed/NCBI
|