Introduction
Esophageal cancer is currently the sixth leading
cause of cancer-associated mortality and the eighth most diagnosed
type of cancer in China (1), where
the incidence of esophageal cancer is markedly increasing, most
likely due to dietary habits (2).
Although great efforts have been made towards the improvement of
diagnosis and therapy, the prognosis of patients with esophageal
cancer remains poor (3). Common
features of esophageal cancer are metastasis and relapse, which are
responsible for the low 5-year survival rates of ~14% (4). Thus, to investigate the mechanism of
cancer cell invasion may provide new insight into cancer
progression and aid in developing improved therapies for esophageal
cancer.
Long non-coding RNAs (lncRNAs) are >200
nucleotides in length and possess no protein-coding capacity
(5). lncRNAs have been reported to
function in multiple biological processes associated with cancer
progression, such as proliferation (6), apoptosis (7) and invasion (8) of cancer cells. The gene lncRNA
plasmacytoma variant translocation 1 (PVT1), which is located at
8q24.21, plays an oncogenic role in various types of human cancer,
including pancreatic cancer (9), lung
cancer (10) and gastric cancer
(11). However, lncRNA PVT1
expression in esophageal cancer and the underlying mechanisms of
its effects remain unclear.
The epithelial-to-mesenchymal transition (EMT) has
been confirmed to be important in cell invasion in different cancer
types (12,13). EMT is closely associated with the
transformation and infiltration of tumor cells. Much research on
the association between lncRNAs and tumor development has focused
on the expression of EMT-associated proteins, such as the
epithelial marker E-cadherin, and the mesenchymal markers
N-cadherin and vimentin (14). The
breakdown of tight junctions is associated with the loss of
epithelial markers and acquisition of mesenchymal makers (15–17).
The current study was performed with the aim of
assessing the expression of lncRNA PVT1 in esophageal cancer, and
exploring the potential mechanism of lncRNA PVT1 in cell invasion.
In addition, the effect of abnormal expression of lncRNA PVT1 in
the regulation of EMT marker expression was investigated.
Materials and methods
Clinical samples
A total of 77 patients with esophageal cancer who
had undergone routine surgery at Huai'an First People's Hospital
(Huai'an, China) between May 2008 and November 2013 were enrolled.
Esophageal cancer tissues and the adjacent tissues were obtained
from the 77 patients; all the samples were immediately snap-frozen
in liquid nitrogen and stored at −80°C until RNA extraction. Each
patient provided written informed consent, and the study was
approved by the Ethics Committee of Huai'an First People's
Hospital.
Cell culture
Esophageal cancer cell lines (Kyse140, TE-1, EC18,
Eca-109 and HKESC1) and a normal esophageal cell line (NEEC) were
obtained from the Cell Bank of Type Culture Collection of the
Chinese Academy of Sciences (Shanghai, China), and cultured in
Invitrogen RPMI-1640 medium with 10% fetal bovine serum (FBS) and
100 U/ml penicillin (all from Thermo Fisher Scientific, Inc.,
Carlsbad, CA, USA) at 37°C in an atmosphere of 5%
CO2.
Plasmid construction and cell
transfection
The sequence of lncRNA PVT1 was synthesized by
Genewiz, Inc. (Suzhou, China). The plasmid was subcloned into a
lentiviral vector (pLV-GFP; Addgene, Inc., Cambridge, MA, USA) and
then co-transfected into HEK-293T cells (SiXin Bio Company,
Shanghai, China) with Lentiviral Packaging Mix (Genewiz, Inc.,
Suzhou, China). TE-1 cells were transfected with lentivirus to
produce cells with high expression of lncRNA PVT1 (LV-lncRNA), and
underwent selection with G418. Cells transfected with LV-vector
constituted the control group.
RNA interference
Small interfering RNA (siRNA; 50 nM) specifically
targeting lncRNA PVT1 and a corresponding scrambled siRNA negative
control (Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA) were
transfected into Eca-109 cells in 6-well plates using Invitrogen
Lipofectamine 2000 reagent (Thermo Fisher Scientific, Inc.),
according to the manufacturer's instructions.
Isolation of total RNA and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA was extracted from the collected tissues
using Invitrogen TRIzol reagent (Thermo Fisher Scientific, Inc.),
and mRNA was reverse transcribed into cDNA. lncRNA PVT1 expression
was quantified using a LightCycler Brilliant SYBR Green RT-qPCR kit
(Roche Applied Science, Indianapolis, IN, USA) according to the
manufacturer's protocol, with the following primers: Forward,
TTGGCACATACAGCCATCAT; and reverse, GCAGTAAAAGGGGAACACCA. β-actin
was used for normalization, with the following primers: Forward,
AGCGAGCATCCCCCAAAGTT; and reverse, GGGCACGAAGGCTCATCATT. PCR was
then performed on ABI PRISM 7500 System (Applied Biosystems; Thermo
Fisher Scientific, Inc.). The PCR conditions were as follows: 94°C
for 2 min, followed by 94°C for 30 sec, 60°C for 30 sec and 72°C
for 1 min for 30 cycles, and 72°C for 10 min. Every independent
experiment was performed three times. The 2−∆∆Cq method
(18) was used to evaluate the
results of RT-qPCR in all the experiments.
Wound healing assay
The transfected cells were plated into 6-well plates
and cultured with RPMI-1640 medium. After 24 h, the transfected
cells were wounded with a pipette tip. Serum-free RPMI-1640 medium
was added, and wound closure was observed for 48 h.
Transwell assay
A transwell invasion assay was performed with
BioCoat Matrigel (BD Biosciences, San Jose, CA, USA) and invasion
chambers (Merck Millipore, Eschborn, Germany) with an 8-µm pore
size. The transfected cells were treated with
trypsin/ethylenediaminetetraacetic acid solution, washed once with
serum-containing RPMI-1640 medium. Cells (1×105) in 0.2 ml
serum-free RPMI-1640 medium were seeded on a Transwell apparatus.
RPMI-1640 containing 10% FBS (600 µl) was added to the lower
chamber. According to the manufacturer's instructions, an invasion
assay was conducted following the same procedure, with the
exception that the filters of the transwell chambers were coated
with 45 µg Matrigel. Following incubation of the cells for 24 h at
37°C in a 5% CO2 incubator, cells on the top surface of
the insert were removed by wiping with a cotton swab. Cells that
invaded to the bottom surface of the insert were fixed in the 100%
precooled methanol for 10 min, stained in 0.5% crystal violet for
30 min, then rinsed in phosphate-buffered saline (PBS) and
subjected to microscopic inspection. The values for invasion were
obtained by counting three fields per membrane and represented the
average of three independent experiments.
Western blot analysis
Total proteins were collected from the established
cells and quantified using a bicinchoninic acid Protein Assay Kit
(Beyotime, Haimen, China). Proteins were fractionated by sodium
dodecyl sulfate polyacrylamide gel electrophoresis, transferred to
polyvinylidene difluoride membranes, blocked in 5% dry milk at 37°C
temperature for 1 h, and immunostained with antibodies at 4°C
overnight. Antibodies against vimentin (rabbit monoclonal
anti-human vimentin; 1:100; ab76601) and glyceraldehyde 3-phosphate
dehydrogenase (GAPDH; rabbit monoclonal anti-human GAPDH; 1:5,000;
ab70699), N-cadherin (rabbit polyclonal anti-human N-cadherin;
1:5,000; ab18203) and E-cadherin (rabbit polyclonal anti-human
E-cadherin; 1:1,000; ab15148) were purchased from Abcam (Cambridge,
MA, USA). Next, the membranes were washed four times with PBS
containing 0.1% Tween 20 (PBS-T; Sigma-Aldrich, St. Louis, MO,
USA), and the secondary antibody (goat polyclonal anti-rabbit
antibody; 1:1,000; ab6721; Abcam) was then added in PBS-T for 1 h
at 37°C. The membranes were next washed three times for 15 min with
PBS-T. The staining was revealed using a Pierce enhanced
chemiluminescence kit (Thermo Fisher Scientific, Inc.), and exposed
in a Molecular Imager ChemiDoc XRS System (Bio-Rad Laboratories,
Inc., Hercules, CA). The integrated density of the band was
quantified by ImageJ version 1.48u software (https://imagej.nih.gov/ij/).
Statistical methods
Statistical analysis was performed using STATA 11
software (StataCorp LP, College Station, TX, USA), and presented
with GraphPad Prism version 4.0 (GraphPad Software, San Diego, CA,
USA). The results obtained from experiment in vitro assays
are presented as the mean ± standard error of the mean from five
separate experiments in triplicates per experiment, and the data
was analyzed by Wilcoxon rank-sum (Mann-Whitney) test. P<0.05
was considered to indicate statistically significant
differences.
Results
lncRNA PVT1 is increased in human
esophageal cancer tissues
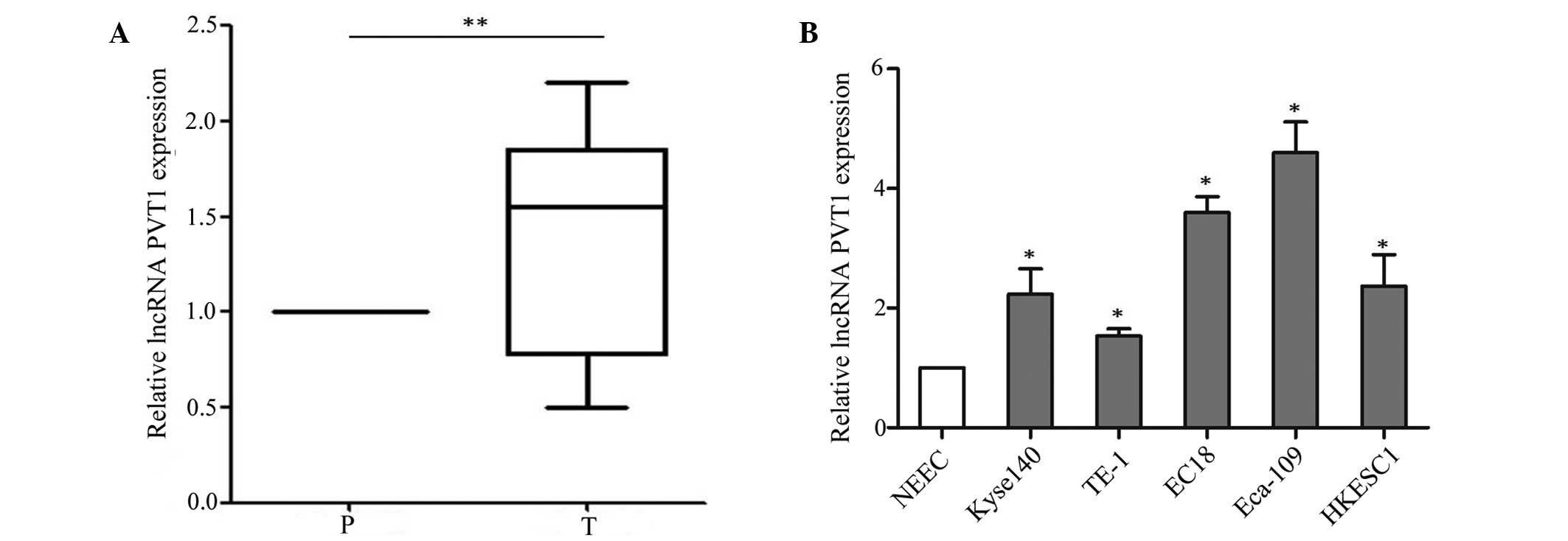
First, the expression of lncRNA PVT1 was analyzed in
esophageal cancer samples (n=77) and adjacent tissues by RT-qPCR.
lncRNA PVT1 expression was significantly higher in esophageal
cancer tissues than adjacent normal tissues (P=0.002) (Fig. 1A). Its expression showed no
association with gender, age, histological type or tumor size;
however, there were significant associations with tumor stage
(P=0.009) and metastasis (P<0.001) (Table I). The aberrant expression level of
lncRNA PVT1 in the cancer tissues suggested that lncRNA PVT1 may
play an important role in the development of esophageal cancer.
Furthermore, the expression of lncRNA PVT1 was assessed in
esophageal cancer cell lines (Kyse140, TE-1, EC18, Eca-109 and
HKESC1) and a normal esophageal cell line (NEEC). The RT-qPCR assay
indicated that lncRNA PVT1 was higher in esophageal cancer cell
lines (Kyse140, TE-1, EC18, Eca-109 and HKESC1) than the normal
esophageal cell line (NEEC) (P=0.016) (Fig. 1B). Based on this expression pattern,
the TE-1 and Eca-109 cell lines were selected to verify the effect
of lncRNA PVT1.
 | Table I.Expression level of lncRNA PVT1 in
esophageal cancer tissues. |
Table I.
Expression level of lncRNA PVT1 in
esophageal cancer tissues.
|
|
| lncRNA PVT1
expression, n |
|
|---|
|
|
|
|
|
|---|
| Factor | Samples, n | High (≥median) | Low (<median) | P-value |
|---|
| Total | 77 | 39 | 38 |
|
| Gender |
|
|
| 0.554 |
| Male | 33 | 18 | 15 |
|
|
Female | 44 | 21 | 23 |
|
| Age (years) |
|
|
| 0.420 |
|
<60 | 36 | 20 | 16 |
|
| ≥60 | 41 | 19 | 22 |
|
| Histological
types |
|
|
| 0.424 |
| Squamous
cell carcinoma | 38 | 21 | 17 |
|
|
Adenocarcinoma | 39 | 18 | 21 |
|
| Tumor stage |
|
|
| 0.009a |
| I–II | 35 | 12 | 23 |
|
|
III–IV | 42 | 27 | 15 |
|
| Metastasis |
|
|
|
<0.001a |
| No | 47 | 16 | 31 |
|
| Yes | 30 | 23 | 7 |
|
lncRNA PVT1 regulates the invasion of
cell lines in vitro
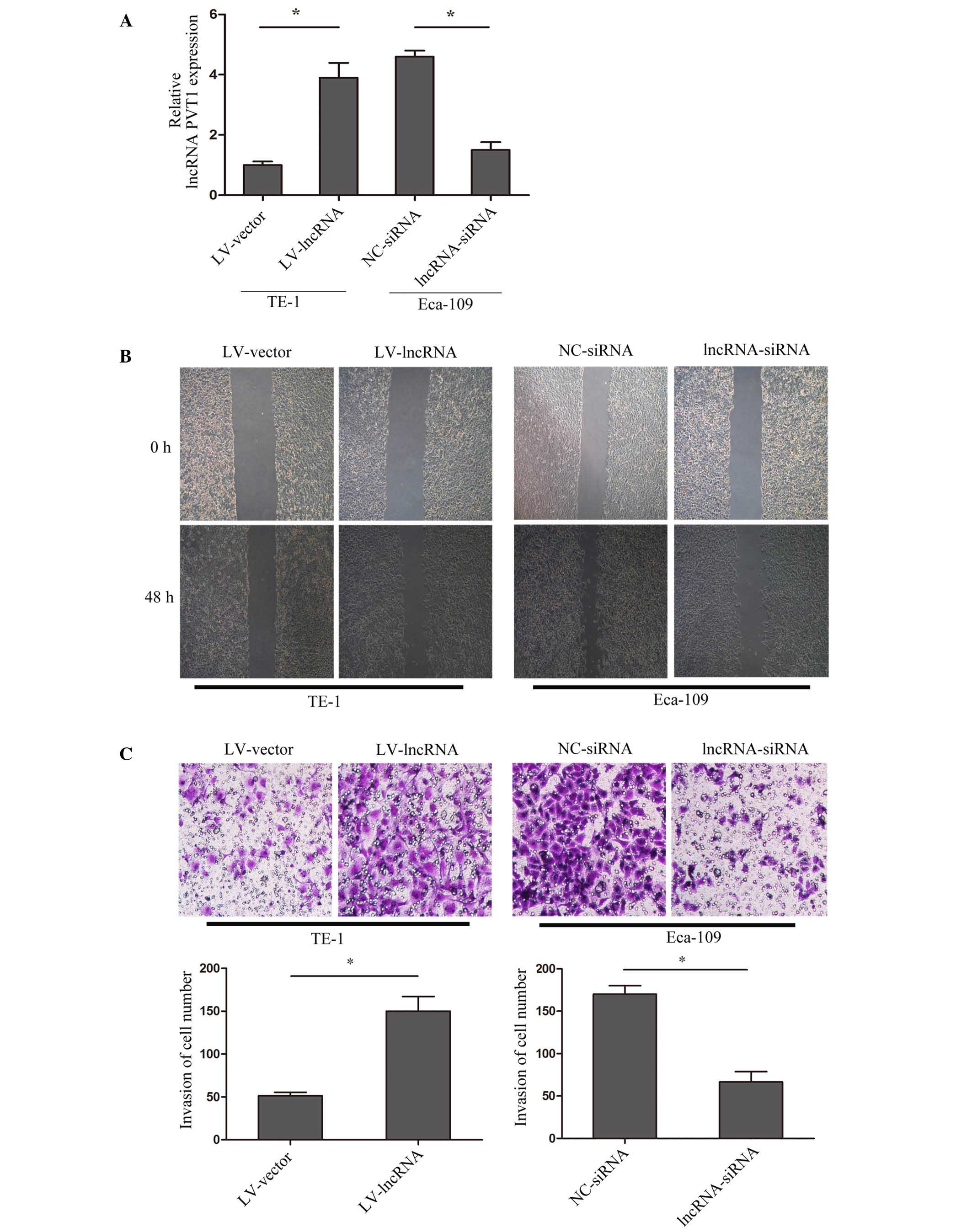
To explore the mechanism of lncRNA PVT1 and cell
invasion in esophageal cancer, lncRNA PVT1 expression was
upregulated in TE-1 cells by transfection with lentivirus (named
LV-lncRNA), and downregulated in Eca-10 cells by siRNA interference
(named lncRNA-siRNA). The transfection efficiency was validated by
RT-qPCR (Fig. 2A). A wound healing
assay indicated that upregulation of lncRNA PVT1 promoted the
healing of cells, while suppressed lncRNA PVT1 inhibited healing
(P=0.009) (Fig. 2B). Furthermore, a
matrigel invasion assay showed that upregulated lncRNA PVT1
promoted cell invasion, while suppressed lncRNA PVT1 inhibited cell
invasion (P=0.0031) (Fig. 2C). The
results suggested that lncRNA PVT1 acts to regulate esophageal
cancer cell invasion.
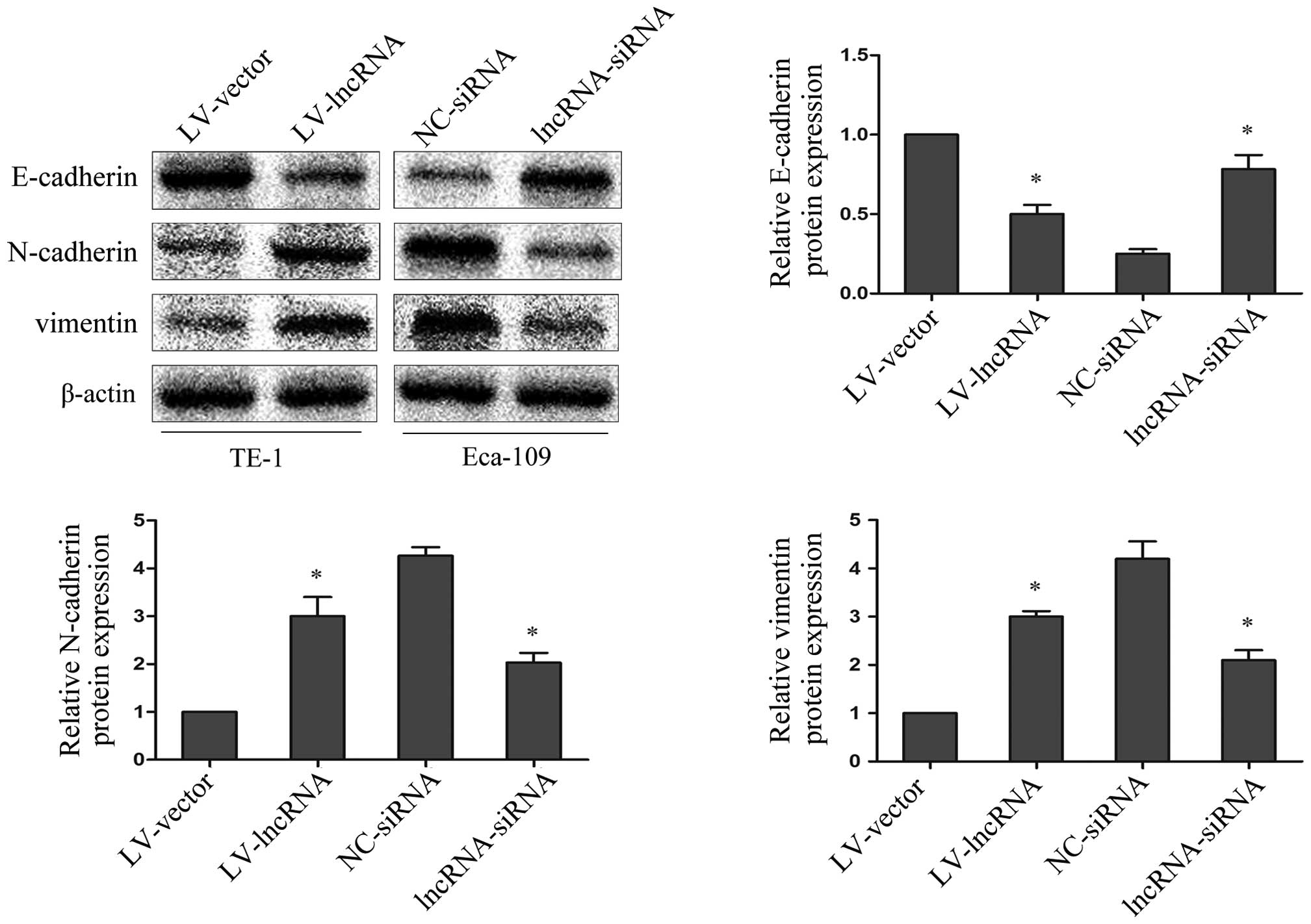
lncRNA PVT1 regulates EMT marker
expression
To explore whether abnormal lncRNA PVT1 expression
levels may be involved in the EMT process, the epithelial marker
E-cadherin, and the mesenchymal markers N-cadherin and vimentin
were investigated by western blotting. Upregulated lncRNA PVT1 in
TE-1 cells resulted in decreased E-cadherin protein expression and
increased N-cadherin and vimentin protein expression; while
suppressed lncRNA PVT1 expression in Eca-109 cells resulted in
increased E-cadherin protein expression and decreased N-cadherin
and vimentin protein expression (P=0.0019) (Fig. 3). This indicates that lncRNA PVT1
contributes to the regulation of EMT marker expression in
esophageal cancer cell lines.
Discussion
lncRNAs have been reported to have critical
regulatory roles in cancer biology (19–21).
Specifically for esophageal cancer, it has already been reported
that lncRNAs contribute to proliferation and invasion (22–24). The
present study sought to provide evidence that upregulation of
lncRNA PVT1 may induce EMT to promote cell invasion in esophageal
cancer. The results indicated that the expression of lncRNA PVT1
was increased in esophageal cancer tissues (n=77) compared to
corresponding adjacent tissues, and was associated with tumor stage
and metastasis. Furthermore, in vitro, upregulation of
lncRNA PVT1 promoted the invasion of esophageal cancer cell lines,
and downregulation of lncRNA PVT1 inhibited the cell invasion.
The major causes of mortality from cancer are
complications arising from cancer cell invasion. Decreased
E-cadherin and elevated N-cadherin and vimentin expression are
hallmarks of EMT, which is a key element in cancer invasion
(25). EMT has been identified to be
associated with tumor invasiveness, metastasis and prognosis
(26,27). Numerous studies have established
functional associations between lncRNAs and key effectors of EMT
occurring in the context of carcinogenesis (28–30). In
addition to cancer progression, EMT contributes to chronic
epithelial injury (31), leading to
tissue fibrosis and organ failure (32,33). In
the current study, it was also discovered that abnormal expression
of lncRNA PVT1 could regulate markers of EMT. Upregulated lncRNA
PVT1 in TE-1 cells resulted in decreased E-cadherin expression and
increased N-cadherin and vimentin expression relative to the
control cells, while suppressed lncRNA PVT1 expression in Eca-109
cells resulted in increased E-cadherin expression and decreased
N-cadherin and vimentin expression. These data suggest that lncRNA
PVT1 promotes esophageal cancer invasion by inducing EMT.
Taken together, the findings of the present study
indicate that overexpression of lncRNA PVT1 is associated with
tumor stage and metastasis in esophageal cancer. In addition,
upregulation of lncRNA PVT1 promotes cell invasion by inducing EMT
in vitro. In the future, the pathway linking lncRNA PVT1 and
EMT may be exploited in a therapeutic approach for the treatment of
esophageal cancer.
Acknowledgements
This research was partly supported by Dr Wei Li for
language revision.
References
|
1
|
Shigaki H, Baba Y, Watanabe M, Murata A,
Ishimoto T, Iwatsuki M, Iwagami S, Nosho K and Baba H: PIK3CA
mutation is associated with a favorable prognosis among patients
with curatively resected esophageal squamous cell carcinoma. Clin
Cancer Res. 19:2451–2459. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen Y, Tong Y, Yang C, Gan Y, Sun H, Bi
H, Cao S, Yin X and Lu Z: Consumption of hot beverages and foods
and the risk of esophageal cancer: A meta-analysis of observational
studies. BMC Cancer. 15:4492015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Liao Y, Xue Y, Zhang L, Feng X, Liu W and
Zhang G: Higher heat shock factor 1 expression in tumor stroma
predicts poor prognosis in esophageal squamous cell carcinoma
patients. J Transl Med. 13:3382015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Koshy M, Esiashvilli N, Landry JC, Thomas
CR Jr and Matthews RH: Multiple management modalities in esophageal
cancer: Combined modality management approaches. Oncologist.
9:147–159. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mercer TR, Dinger ME and Mattick JS: Long
non-coding RNAs: Insights into functions. Nat Rev Genet.
10:155–159. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shi Y, Li J, Liu Y, Ding J, Fan Y, Tian Y,
Wang L, Lian Y, Wang K and Shu Y: The long noncoding RNA SPRY4-IT1
increases the proliferation of human breast cancer cells by
upregulating ZNF703 expression. Mol Cancer. 14:512015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Han L, Zhang EB, Yin DD, Kong R, Xu TP,
Chen WM, Xia R, Shu YQ and De W: Low expression of long noncoding
RNA PANDAR predicts a poor prognosis of non-small cell lung cancer
and affects cell apoptosis by regulating Bcl-2. Cell Death Dis.
6:e16652015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hu L, Wu Y, Tan D, Meng H, Wang K, Bai Y
and Yang K: Up-regulation of long noncoding RNA MALAT1 contributes
to proliferation and metastasis in esophageal squamous cell
carcinoma. J Exp Clin Cancer Res. 34:72015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Huang C, Yu W, Wang Q, Cui H, Wang Y,
Zhang L, Han F and Huang T: Increased expression of the lncRNA PVT1
is associated with poor prognosis in pancreatic cancer patients.
Minerva Med. 106:143–149. 2015.PubMed/NCBI
|
|
10
|
Yang YR, Zang SZ, Zhong CL, Li YX, Zhao SS
and Feng XJ: Increased expression of the lncRNA PVT1 promotes
tumorigenesis in non-small cell lung cancer. Int J Clin Exp Pathol.
7:6929–6935. 2014.PubMed/NCBI
|
|
11
|
Ding J, Li D, Gong M, Wang J, Huang X, Wu
T and Wang C: Expression and clinical significance of the long
non-coding RNA PVT1 in human gastric cancer. Onco Targets Ther.
7:1625–1630. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu SG, Qin XG, Zhao BS, Qi B, Yao WJ,
Wang TY, Li HC and Wu XN: Differential expression of miRNAs in
esophageal cancer tissue. Oncol Lett. 5:1639–1642. 2013.PubMed/NCBI
|
|
13
|
Liu J, Ruan B, You N, Huang Q, Liu W, Dang
Z, Xu W, Zhou T, Ji R, Cao Y, et al: Downregulation of miR-200a
induces EMT phenotypes and CSC-like signatures through targeting
the β-catenin pathway in hepatic oval cells. PLoS One.
8:e794092013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Han Y, Ye J, Wu D, Wu P, Chen Z, Chen J,
Gao S and Huang J: LEIGC long non-coding RNA acts as a tumor
suppressor in gastric carcinoma by inhibiting the
epithelial-to-mesenchymal transition. BMC Cancer. 14:9322014.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Dong H, Xie L, Tang C, Chen S, Liu Q,
Zhang Q, Zheng W, Zheng Z and Zhang H: Snail1 correlates with
patient outcomes in E-cadherin-preserved gastroesophageal junction
adenocarcinoma. Clin Transl Oncol. 16:783–791. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu Y, Li H, Feng J, Cui X, Huang W, Li Y,
Su F, Liu Q, Zhu J, Lv X, et al: Lin28 induces
epithelial-to-mesenchymal transition and stemness via
downregulation of let-7a in breast cancer cells. PLoS One.
8:e830832013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bao YX, Cao Q, Yang Y, Mao R, Xiao L,
Zhang H, Zhao HR and Wen H: Expression and prognostic significance
of Golgiglycoprotein73 (GP73) with epithelial-mesenchymal
transition (EMT) related molecules in hepatocellular carcinoma
(HCC). Diagn Pathol. 8:1972013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Iguchi T, Uchi R, Nambara S, Saito T,
Komatsu H, Hirata H, Ueda M, Sakimura S, Takano Y, Kurashige J, et
al: A long noncoding RNA, lncRNA-ATB, is involved in the
progression and prognosis of colorectal cancer. Anticancer Res.
35:1385–1388. 2015.PubMed/NCBI
|
|
20
|
Wang CM, Wu QQ, Li SQ, Chen FJ, Tuo L, Xie
HW, Tong YS, Ji L, Zhou GZ, Cao G, et al: Upregulation of the long
non-coding RNA PlncRNA-1 promotes esophageal squamous carcinoma
cell proliferation and correlates with advanced clinical stage. Dig
Dis Sci. 59:591–597. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ge XS, Ma HJ, Zheng XH, Ruan HL, Liao XY,
Xue WQ, Chen YB, Zhang Y and Jia WH: HOTAIR, a prognostic factor in
esophageal squamous cell carcinoma, inhibits WIF-1 expression and
activates Wnt pathway. Cancer Sci. 104:1675–1682. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li W, Zheng J, Deng J, You Y, Wu H, Li N,
Lu J and Zhou Y: Increased levels of the long intergenic
non-protein coding RNA POU3F3 promote DNA methylation in esophageal
squamous cell carcinoma cells. Gastroenterology. 146:1714–1726.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xie HW, Wu QQ, Zhu B, Chen FJ, Ji L, Li
SQ, Wang CM, Tong YS, Tuo L, Wu M, et al: Long noncoding RNA
SPRY4-IT1 is upregulated in esophageal squamous cell carcinoma and
associated with poor prognosis. Tumour Biol. 35:7743–7754. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Pan F, Yao J, Chen Y, Zhou C, Geng P, Mao
H and Fang X: A novel long non-coding RNA FOXCUT and mRNA FOXC1
pair promote progression and predict poor prognosis in esophageal
squamous cell carcinoma. Int J Clin Exp Pathol. 7:2838–2849.
2014.PubMed/NCBI
|
|
25
|
Kitamura K, Seike M, Okano T, Matsuda K,
Miyanaga A, Mizutani H, Noro R, Minegishi Y, Kubota K and Gemma A:
MiR-134/487b/655 cluster regulates TGF-β-induced
epithelial-mesenchymal transition and drug resistance to gefitinib
by targeting MAGI2 in lung adenocarcinoma cells. Mol Cancer Ther.
13:444–453. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Guo S, Xu X, Tang Y, Zhang C, Li J, Ouyang
Y, Ju J, Bie P and Wang H: miR-15a inhibits cell proliferation and
epithelial to mesenchymal transition in pancreatic ductal
adenocarcinoma by down-regulating Bmi-1 expression. Cancer Lett.
344:40–46. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yamada S, Fuchs BC, Fujii T, Shimoyama Y,
Sugimoto H, Nomoto S, Takeda S, Tanabe KK, Kodera Y and Nakao A:
Epithelial-to-mesenchymal transition predicts prognosis of
pancreatic cancer. Surgery. 154:946–954. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li SP, Xu HX, Yu Y, He JD, Wang Z, Xu YJ,
Wang CY, Zhang HM, Zhang RX, Zhang JJ, et al: LncRNA HULC enhances
epithelial-mesenchymal transition to promote tumorigenesis and
metastasis of hepatocellular carcinoma via the miR-200a-3p/ZEB1
signaling pathway. Oncotarget. 7:2016.(Epub ahead of print) doi:
10.18632/oncotarget.9883.
|
|
29
|
Wang Y, Liu Z, Yao B, Dou C, Xu M, Xue Y,
Ding L, Jia Y, Zhang H, Li Q, et al: Long non-coding RNA TUSC7 acts
a molecular sponge for miR-10a and suppresses EMT in hepatocellular
carcinoma. Tumour Biol. 2016.(Epub ahead of print).
|
|
30
|
Sun R, Qin C, Jiang B, Fang S, Pan X, Peng
L, Liu Z, Li W, Li Y and Li G.: Down-regulation of MALAT1 inhibits
cervical cancer cell invasion and metastasis by inhibition of
epithelial-mesenchymal transition. Mol Biosyst. 12:952–962. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Vitalone MJ, Naesens M, Sigdel T, Li L,
Hseih S and Sarwal MM: The dual role of epithelial-to-mesenchymal
transition in chronic allograft injury in pediatric renal
transplantation. Transplantation. 92:787–795. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
López-Novoa JM and Nieto MA: Inflammation
and EMT: An alliance towards organ fibrosis and cancer progression.
EMBO Mol Med. 1:303–314. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Mucsi I and Rosivall L:
Epithelial-mesenchymal transition in renal tubular cells in the
pathogenesis of progressive tubulo-interstitial fibrosis. Acta
Physiol Hung. 94:117–131. 2007. View Article : Google Scholar : PubMed/NCBI
|