Introduction
Osteosarcoma (OS), known as the most common primary
malignant tumor arising from bone, is derived from mesenchymal
cells, which generate osteoid and immature bone (1–3). It has
been demonstrated that OS has a significant age distribution; the
first peak appears during adolescence (10–14 years old), and the
second peak occurs in elderly adults (>65 years) (1). Although advanced treatments, such as
surgery, chemotherapy and radiotherapy, are used, the 5-year
survival rate of OS patients is 60–70% (4). It has been reported that the morbidity
of males diagnosed with OS is 1.4 times higher than that of females
(5). Furthermore, OS mainly occurs in
the long bones, particularly in the metaphyses, proximal tibia,
proximal humerus and distal femur, while ~10% of OS occurs in the
axial skeleton, most frequently in the pelvis (5,6).
Meanwhile, ~30% of OS patients have the potential for pulmonary
metastatic disease, and the 5-year survival rates are considered to
be <20% (7). It has been shown
that the pathogenesis of OS contributes to the interactions between
genetic and environmental risk factors, including smoking, air
population and ionizing radiation (4,8–10). Numerous studies have focused on the
association of microRNAs (miRNAs/miRs) and the development of OS
and its underlying mechanisms (1,10).
miRNAs are a class of novel small non-coding RNA
molecules that can regulate the gene expression of target mRNA in
animals and plants (11–13). Previous studies have suggested that
miRNAs exhibit significant functions in certain physiological
processes, including cell proliferation, differentiation,
apoptosis, angiogenesis, stress resistance and the self-renewal of
stem cells (14,15). In addition, miRNAs may function as
tumor suppressors and oncogenes in specific cases, and certain
miRNAs are also viewed as candidate targets for intervention
therapy or molecular markers for the early diagnosis and prognosis
of tumors (16–18). Recently, studies have demonstrated
that various miRNAs, including miR-125b, miR-143, miR-20a,
miR-199a-3p, miR-34a and miR-150, may be closely associated with
tumor metastasis and invasion in OS (19,20). Among
them, miR-150, which is abundant in lymphocytes, acts as a
sensitive marker that indicates bone marrow damage and lymphocyte
depletion (21,22). It is well documented that miR-150
plays an essential role as an oncogene in different tumors,
including gastric cancer, OS, lung cancer and hepatoblastoma
(23,24). Ezrin, known as a member of the
Ezrin-Radixin-Moesin protein family, is a cytoskeleton linkage
protein that actively participates in the regulation of growth and
metastasis of cancer cells (25).
Ezrin is also defined as a predominant component of the surface
structure of cells, being involved in adhesion to the extracellular
matrix, cell-to-cell communication and the signaling pathway of
receptor tyrosine kinase (26). Due
to the function of membrane-cytoskeleton linking, Ezrin has been
found to be essential for the metastasis of OS in a rat model,
while the high expression of Ezrin in dogs with OS has also been
correlated with the early progress of metastasis (27). The present study aimed to investigate
the effect of miRNA-150 on the proliferation and metastasis of the
human OS F5M2 cell line, in order to provide a basis for the
treatment of OS.
Materials and methods
Cells culture and grouping design
The human OS F5M2 cell line was acquired from the
American Type Culture Collection (Manassas, VA, USA) and cultured
in RPMI-1640 medium that contained 10% fetal bovine serum (FBS;
Gibco, Life Technologies, Carlsbad, CA, USA), 100 U/ml penicillin G
and 100 U/ml streptomycin (Invitrogen, Life Technologies, Carlsbad,
CA, USA) at 37°C with 5% CO2. The optimal F5M2 subline
during logarithmic growth phase was harvested with 0.05% trypsin
(containing 0.02% EDTA). The harvested F5M2 subline samples were
assigned to five groups: i) Control group (untreated F5M2 cell
subline); ii) miR-150 low-dose group (F5M2 cells transfected with
20 nM miR-150 mimics); iii) miR-150 high-dose group (F5M2 cells
transfected with 50 nM miR-150 mimics); iv) negative control (NC)
group (F5M2 cells transfected with 20 nM miR-150 NC, acting as the
control of the miR-150 mimics group and the miR-150 inhibitors
group); v) inhibitor group (F5M2 cells transfected with 20 nM
miR-150 inhibitor). miR-150 mimic, miR-150-negative control and
miR-150 inhibitor were provided by Guangzhou RiboBio Co., Ltd.
(Guangzhou, China).
The cells were cultured in 50 ml complete culture
medium, and grown to 30–50% density. Lipofectamine® 2000
(Invitrogen, Life Technologies) and miR-150 mimic/miR-150
inhibitor/miR-150 negative control were prepared in a sterile
Eppendorf tube (Eppendorf microfuge, Brinkmann Instruments, Inc.,
Westbury, NY, USA). The diluted miR-150 mimic/miR-150
inhibitor/miR-150 negative control and Lipofectamine 2000 were
mixed gently to a total volume of 100 µl, and incubated at room
temperature for 20 min. A total of 100 µl of transfection solution
was added to each cell with gentle agitation. Cells were
subsequently incubated at 37°C in an atmosphere of 5%
CO2, followed by removal of transfection solution and
replacement with complete medium 6–8 h later. Following 48 to 72 h
of culture, the cells were harvested and the total protein was
extracted. The experiment was repeated 3 times for each group.
The study was approved by the ethics committee of
the Second Military Medical University (Shanghai, China).
Proliferation assay
Transfected F5M2 cells of each group were seeded in
seven 96-well plates (1×104 cells per well) using
methylthiazolyldiphenyl-tetrazolium bromide (MTT) assay
(Sigma-Aldrich, St. Louis, MO, USA). The absorbance (OD value) was
determined at 490 nm (one plate per day) on a microplate
spectrophotometer (SpectraMax, Molecular Devices, Sunnyvale, CA,
USA). After seven days, the average absorbance was calculated. Cell
growth curves were obtained for the comparison of growth conditions
among groups.
Cell migration in vitro
Transwell inserts (24-well insert; pore size, 8 µm)
were applied to investigate the effect of miR-150 on the migration
and invasion of the F5M2 cells. Transwell inserts were coated with
Matrigel (50 µl per filter) (BD Biosciences, Franklin Lakes, NJ,
USA) based on the manufacturer's instructions. Following
transfection, the F5M2 cells were transferred onto the
Matrigel-coated upper inserts (at a density of 1×105 per
Transwell), which contained serum-free Dulbecco's modified Eagle's
medium (DMEM) and then incubated at 37°C for 48 h. DMEM containing
10% FBS was added to the lower inserts and further incubation was
performed for 24 h. The cells which migrated to the lower inserts
were fixed with 90% ethanol and stained with crystal violet, and
the cells that remained on the top of the filters were scrubbed
off. Six random visual fields of each insert were counted
subsequent to being observed under a microscope. Migration assay
was performed in the same manner as the aforementioned procedures
but without the Matrigel coating. The cells that penetrated the
upper side of the membrane were fixed with ethanol and stained.
Finally, the cells were counted under a microscope (Eclipse E600;
Nikon Inc., Melville, NY, USA).
Apoptosis test
Apoptotic cells were analyzed by double staining
with Annexin V-FITC/PI Apoptosis Assay (BD Biosciences Pharmingen,
San Diego, CA, USA). Following transfection, adherent cells were
trypsinized, centrifuged (400 × g) and washed three times in
phosphate-buffered saline. The cells were resuspended with 300 µl
staining solution containing fluorescein isothiocyanate
(FITC)-conjugated Annexin V antibody and propidium iodide (PI).
Incubation was performed on ice for 15 min and the cells were
analyzed using flow cytometry (BD Biosciences). The number of
apoptotic cells (Annexin+/PI−) was recorded
by applying CellQuest Software (BD Biosciences Pharmingen).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was isolated using TRIzol reagent
(Invitrogen, Life Technologies) and then reverse transcription was
performed. Designed PCR primers for miR-150 were as follows:
Forward, 5′-TGTCCTGCGGGAGTGGTA-3′ and reverse,
5′-TGATGCTGGTGGGTCTGC-3′. The length of the PCR product was 387 bp.
Primers were synthesized by Invitrogen (Shanghai, China). PCR
conditions were predenaturation at 95°C for 5 min, denaturation at
94°C for 50 sec, annealing at 58°C for 50 sec and elongation at
72°C for 1 min, for a total of 30 cycles. PCR products were
separated by electrophoresis on 2% agarose gels and then the
results were analyzed.
Western blotting
Total proteins were extracted and detected by
western blot analysis. For further study, the protein was
electrophoresed by 12% SDS-PAGE, then transferred onto
polyvinylidene difluoride (PVDF) membranes (EMD Millipore,
Billerica, MA, USA). Subsequent to being blocked with 10% skimmed
milk for 1 h, the membranes were washed with TBS. The goat
polyclonal antibody against ezrin (C-15; di1ution, 1:200; Santa
Cruz Biotechnology Inc., Dallas, TX, USA), was added at a 1:1,000
dilution, using U6 as the internal control; horseradish peroxidase
(HRP)-labeled immunoglobulin G (Jackson ImmunoResearch, West Grove,
PA, USA) was supplemented as the secondary antibody at a 1:1,000
dilution. Subsequently, chemiluminescence immunoblotting detection
reagents were applied to the PVDF membranes. X-ray was utilized for
exposure and visualization, and then the expression level of Ezrin
was analyzed by the intensity of electrophoresis.
Statistical analysis
Statistical analyses were conducted using SPSS 18.0
software (SPSS, Inc., Chicago, IL, USA). Data were expressed as the
mean ± standard deviation. Measurement data were by compared by
applying a t-test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Expression of miR-150 and Ezrin
protein
miR-150 expression in each group was detected by
RT-qPCR and the 2−ΔΔCT method. Higher miR-150 expression
levels were detected in the miR-150 low-dose and high-dose groups
compared with the control group at 48 h post-transfection (both
P<0.05). When compared with the control group, the expression of
miR-150 was evidently decreased (both P<0.05), while there was
no statistically significant difference between the control and the
NC groups (P>0.05) (Table I). The
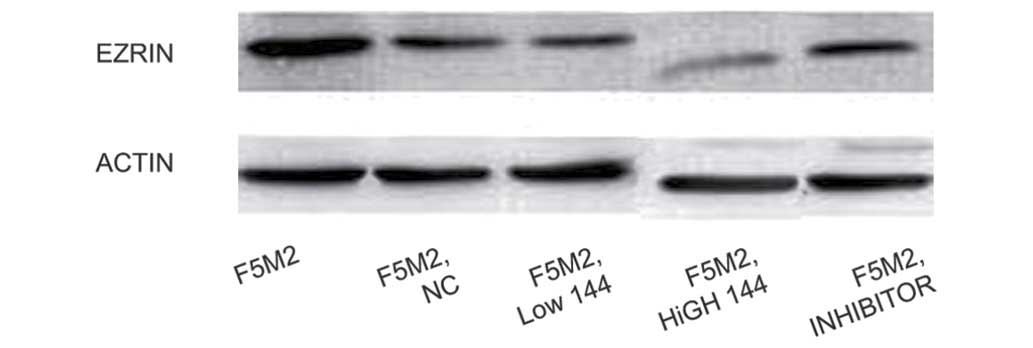
level of Ezrin expression in each group was detected by western
blot analysis. It was observed that Ezrin expression levels in the
low-dose and high-dose groups were notably lower than that in the
control group, and the expression of Ezrin in the high-dose group
was markedly reduced compared with the low-dose group (Fig. 1).
 | Table I.Expression of miR-150 in each
group. |
Table I.
Expression of miR-150 in each
group.
| Group | mean ± SD | MD | 95% CI | P-value |
|---|
| Control | 19.67±2.16 |
|
|
|
| Negative control | 17.33±4.50 | −2.34 | −5.66 to
10.34 | 0.462 |
| miR-150 low-dose | 66.00±10.08 | 46.33 | −62.85 to −29.81 | 0.002 |
| miR-150
high-dose | 75.17±8.23 | 55.50 | −69.14 to −41.86 | <0.001 |
| Inhibitor | 7.83±2.71 | −11.84 | 6.29 to
17.39 | 0.004 |
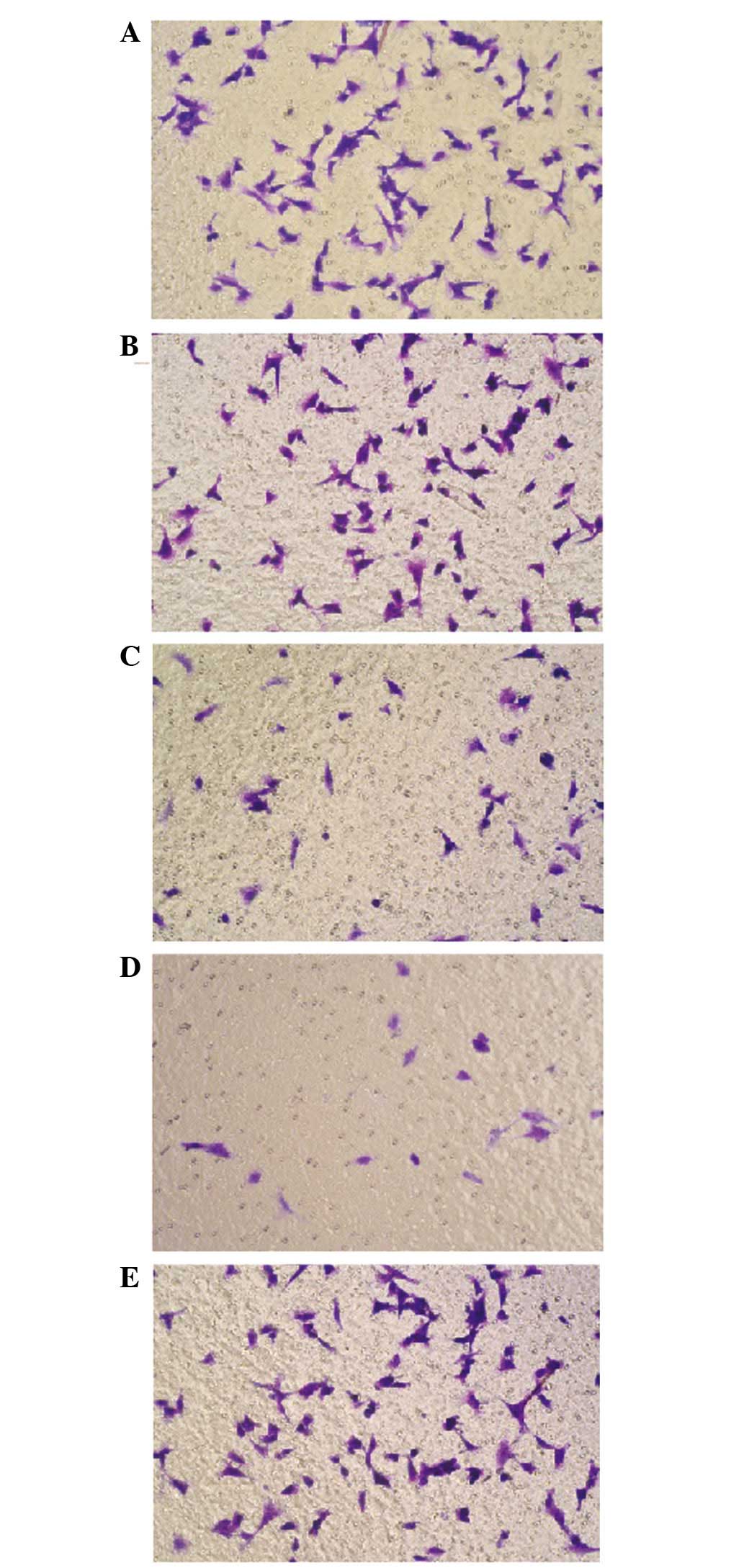
Migration ability of F5M2 cells and
transfected F5M2 cells
A paired samples t-test was applied to compare the
migration ability through the poly-carbonate membrane in the
Transwell experiment. Subsequent to being transfected with miR-150,
the migration ability of the miR-150 low-dose and high-dose groups
decreased in contrast to the F5M2 control cells without
transfection. Compared with the control group, no significant
effect on migration ability was found in the NC and inhibitor
groups (both P>0.05) (Fig. 2).
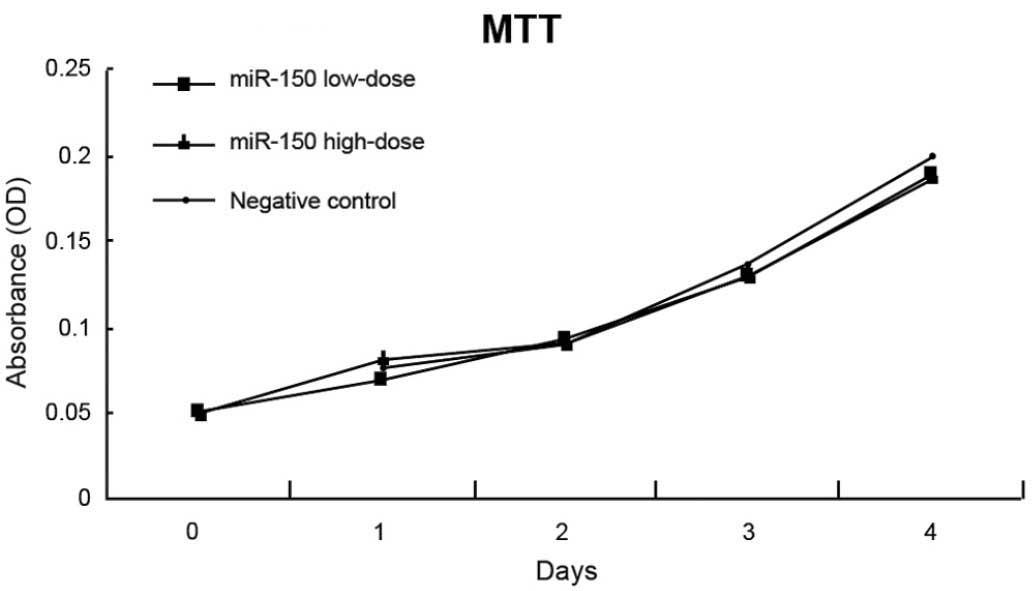
Proliferation and apoptosis of F5M2
cells
The absorbance of untreated F5M2 cells, F5M2 cells
transfected with high-dose miR-150 mimics, F5M2 cells transfected
with low-dose miR-150 mimics, F5M2 cells transfected with
inhibitors and F5M2 cells in the NC group were continuously
detected for seven days, and then growth curves were drawn
(Fig. 3). According to the MTT assay,
the cell growth curves of the F5M2 cells in the different groups
were close to each other, indicating no marked differences between
the proliferation of the groups (P>0.05). The cell growth curves
of the F5M2 cells in the high-dose and control groups also showed
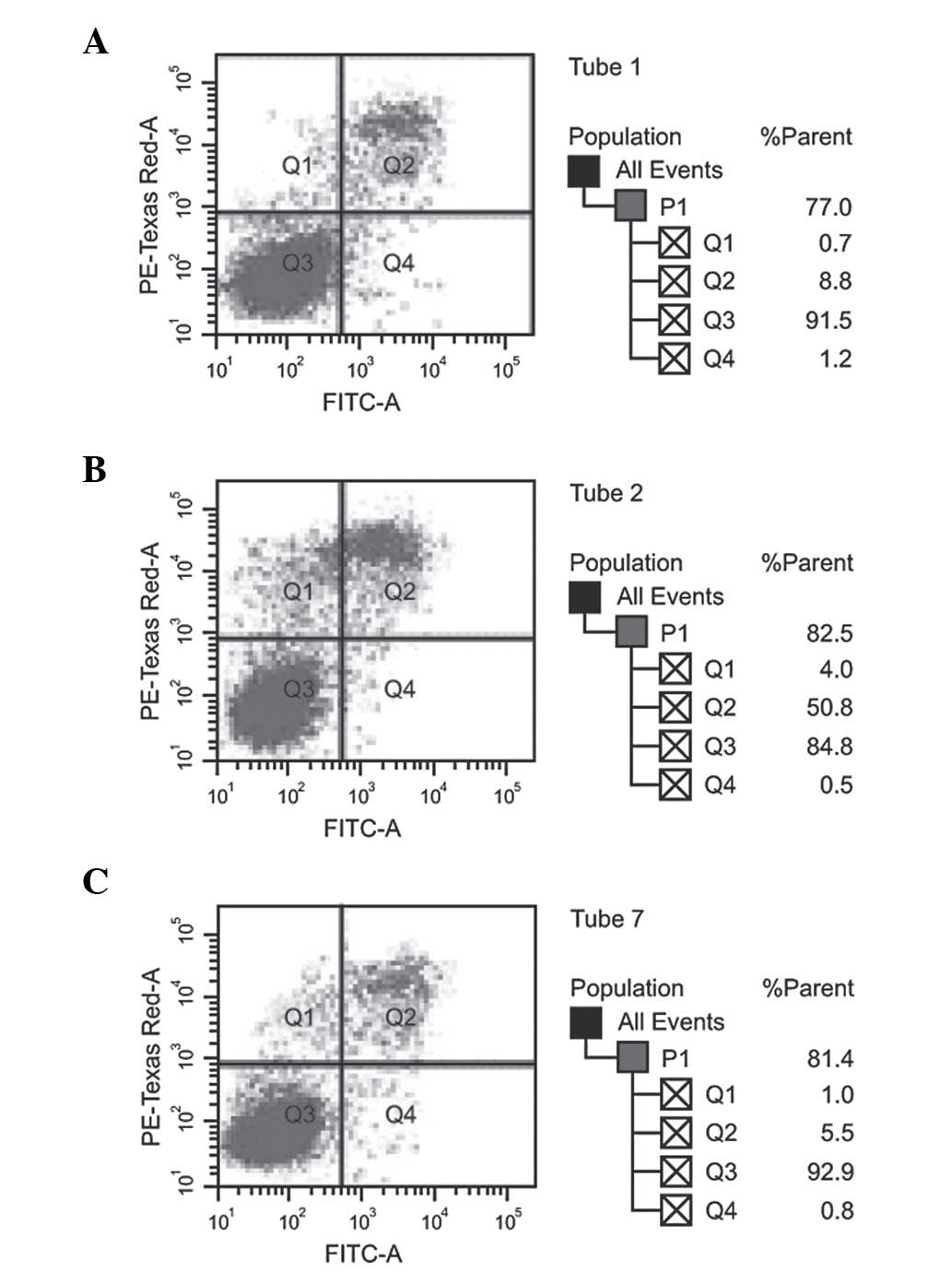
no significant differences (P>0.05). Fig. 4 shows that the apoptotic capacity of
the high-dose and low-dose miR-150-transfected F5M2 cells did not
differ from that of the control F5M2 cells upon flow cytometry
(P>0.05). Additionally, no marked statistical difference in the
total apoptosis rate was observed between the NC and control groups
(P>0.05).
Discussion
The results of the present study showed that the
expression of miR-150 in the OS cells significantly improved after
the exogenous transfection of miR-150 mimics. Watanabe et al
reported that miR-150 can suppress malignant lymphoma by activating
the phosphoinositide 3-kinase-Akt pathway (28). Yokobori et al observed that
regulation of miR-150 expression in esophageal squamous cell
carcinoma could inhibit malignant potential, such as lymph node
metastasis and invasion, by controlling the activity of zinc finger
E-box binding homeobox 1 (29). Thus,
analysis of previous studies concluded that the expression of
miR-150 in highly metastatic OS is lower than that in less
metastatic OS. It was also indicated that the expression of miR-150
in F5M2 cells may be improved through the transfection of miR-150
mimics (30). Similarly, the present
study suggested that the expression of miR-150 shows a positive
association with the transfection concentration of miR-150
mimics.
Another pivotal result from the present study was
that the exogenous overexpression of miR-150 may lead to a
significant decrease in the invasion and metastasis ability of F5M2
cells, indicating a negative dose-dependent association. In
addition, miR-150 had no significant effect on the ability for
proliferation and apoptosis in the F5M2 cells. One potential
explanation for this may be that the overexpression of miR-150
could contribute significantly to the reduction in the migration
and invasion ability of OS F5M2 cells in vitro; the
relatively higher concentration of exogenous miR-150 appears to be
correlated with the inhibitory effect on the migration and invasion
of tumor cells (25,31). Studies have demonstrated that the
metastatic ability of OS is inversely associated with the
expression levels of miR-150. Additionally, the expression level of
miR-150 is inversely associated with the expression of Ezrin, as
observed in the present study, suggesting that miR-150 may inhibit
the expression of Ezrin to a certain extent (32,33).
Ezrin has been suggested to play a role in the
physical and functional connections between the actin cytoskeleton
and the cell membrane (34).
Moreover, Ezrin also has effects on maintaining cell shape and cell
polarity, participating in membrane-interacting pathways, and the
migration, signaling, growth regulation and differentiation of
cells; due to the unique functions of Ezrin, the protein has been
considered to be involved in the metastatic capacity of cancer
(25). According to the result of the
present study, it was concluded that the level of Ezrin in cells
may not only be the marker that distinguishes tumors from
non-tumors, but also that it may predict the metastatic potential
of tumor cells, which is of significance for clinical guidance in
the estimation of OS metastasis and its prognosis. Moreover, the
level of Ezrin expression is positively correlated with the
metastatic ability of OS, suggesting that a decrease in Ezrin
expression is a positive predictive factor of the overall survival
rate of patients with OS (27).
Therefore, the expression of Ezrin can be considered as a novel
area research for the treatment of OS metastasis (35). We also postulate that the expression
of Ezrin may be adversely associated with miR-150 expression.
Accordingly, the exogenous overexpression of miR-150 may decrease
the metastasis of OS through downregulating the expression of Ezrin
(30). Furthermore, with an increase
in exogenous miR-150, the inhibition of Ezrin expression secreted
by F5M2 cells becomes more apparent (36). Thus, there may be a dependent
association between the inhibition of Ezrin expression and the
dosage of exogenous miR-150 in F5M2 cells.
However, certain limitations existed in the present
study that may merit a further investigation. Firstly and
importantly, the sample size may have been too small to adequately
address the role of exogenous miR-126 in OS via targeting Ezrin
expression. Moreover, the retrospective study design will have
affected the grouping of the cases and controls. With respect to
those limitations, larger multi-center randomized controlled trials
are necessary for the verification of these findings.
Collectively, miR-150 was negatively correlated with
the invasion and metastatic ability of the OS cells, while a
positive correlation was found with Ezrin expression. The elevated
exogenous expression of miR-150 could inhibit the invasion and
metastatic ability of F5M2 cells via negatively regulating Ezrin
and have no significant effect on proliferation and apoptosis, in
turn providing a novel molecular target for the therapeutic
intervention in OS.
Acknowledgements
The authors would like to acknowledge the reviewers
of this study for providing useful comments on the original
manuscript.
References
|
1
|
Ottaviani G and Jaffe N: The epidemiology
of osteosarcoma. Cancer Treat Res. 152:3–13. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tang N, Song WX, Luo J, Haydon RC and He
TC: Osteosarcoma development and stem cell differentiation. Clin
Orthop Relat Res. 466:2114–2130. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ta HT, Dass CR, Choong PF and Dunstan DE:
Osteosarcoma treatment: State of the art. Cancer Metastasis Rev.
28:247–263. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Broadhead ML, Clark JC, Myers DE, Dass CR
and Choong PF: The molecular pathogenesis of osteosarcoma: A
review. Sarcoma. 2011:9592482011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ritter J and Bielack SS: Osteosarcoma. Ann
Oncol. 21(Suppl 7): vii320–vii325. 2010.PubMed/NCBI
|
|
6
|
Corradi D, Wenger DE, Bertoni F, Bacchini
P, Bosio S, Goldoni M, Unni KK, Sim FH and Inwards CY: Multicentric
osteosarcoma: Clinicopathologic and radiographic study of 56 cases.
Am J Clin Pathol. 136:799–807. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lussier DM, O'Neill L, Nieves LM, McAfee
MS, Holechek SA, Collins AW, Dickman P, Jacobsen J, Hingorani P and
Blattman JN: Enhanced T-cell immunity to osteosarcoma through
antibody blockade of PD-1/PD-L1 interactions. J Immunother.
38:96–106. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gorlick R: Current concepts on the
molecular biology of osteosarcoma. Cancer Treat Res. 152:467–478.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mirabello L, Pfeiffer R, Murphy G, Daw NC,
Patiño-Garcia A, Troisi RJ, Hoover RN, Douglass C, Schüz J, Craft
AW and Savage SA: Height at diagnosis and birth-weight as risk
factors for osteosarcoma. Cancer Causes Control. 22:899–908. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Jones KB, Salah Z, Del Mare S, Galasso M,
Gaudio E, Nuovo GJ, Lovat F, LeBlanc K, Palatini J and Randall RL:
miRNA signatures associate with pathogenesis and progression of
osteosarcoma. Cancer Res. 72:1865–1877. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
O'Connell RM, Rao DS and Baltimore D:
microRNA regulation of inflammatory responses. Annu Rev Immunol.
30:295–312. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Reid G, Kirschner MB and van Zandwijk N:
Circulating microRNAs: Association with disease and potential use
as biomarkers. Crit Rev Oncol Hematol. 80:193–208. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wu Q, Jin H, Yang Z, Luo G, Lu Y, Li K,
Ren G, Su T, Pan Y, Feng B, et al: MiR-150 promotes gastric cancer
proliferation by negatively regulating the pro-apoptotic gene EGR2.
Biochem Biophys Res Commun. 392:340–345. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Baraniskin A, Kuhnhenn J, Schlegel U, Chan
A, Deckert M, Gold R, Maghnouj A, Zöllner H, Reinacher-Schick A,
Schmiegel W, et al: Identification of microRNAs in the
cerebrospinal fluid as marker for primary diffuse large B-cell
lymphoma of the central nervous system. Blood. 117:3140–3146. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Turchinovich A, Weiz L, Langheinz A and
Burwinkel B: Characterization of extracellular circulating
microRNA. Nucleic Acids Res. 39:7223–7233. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
McDermott AM, Heneghan HM, Miller N and
Kerin MJ: The therapeutic potential of microRNAs: Disease
modulators and drug targets. Pharm Res. 28:3016–3029. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mendell JT and Olson EN: MicroRNAs in
stress signaling and human disease. Cell. 148:1172–1187. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Olson EN: MicroRNAs as therapeutic targets
and biomarkers of cardiovascular disease. Sci Transl Med.
6:239ps32014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhou X, Wei M and Wang W: MicroRNA-340
suppresses osteosarcoma tumor growth and metastasis by directly
targeting ROCK1. Biochem Biophys Res Commun. 437:653–658. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bao YP, Yi Y, Peng LL, Fang J, Liu KB, Li
WZ and Luo HS: Roles of microRNA-206 in osteosarcoma pathogenesis
and progression. Asian Pac J Cancer Prev. 14:3751–3755. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jacob NK, Cooley JV, Yee TN, Jacob J,
Alder H, Wickramasinghe P, Maclean KH and Chakravarti A:
Identification of sensitive serum microRNA biomarkers for radiation
biodosimetry. PLoS One. 8:e576032013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang Y, Liu D, Chen X, Li J, Li L, Bian
Z, Sun F, Lu J, Yin Y, Cai X, et al: Secreted monocytic miR-150
enhances targeted endothelial cell migration. Mol Cell. 39:133–144.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bezman NA, Chakraborty T, Bender T and
Lanier LL: miR-150 regulates the development of NK and iNKT cells.
J Exp Med. 208:2717–2731. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang N, Wei X and Xu L: miR-150 promotes
the proliferation of lung cancer cells by targeting P53. FEBS Lett.
587:2346–2351. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Park HR, Jung WW, Bacchini P, Bertoni F,
Kim YW and Park YK: Ezrin in osteosarcoma: comparison between
conventional high-grade and central low-grade osteosarcoma. Pathol
Res Pract. 202:509–515. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hunter KW: Ezrin, a key component in tumor
metastasis. Trends Mol Med. 10:201–204. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Khanna C, Wan X, Bose S, Cassaday R, Olomu
O, Mendoza A, Yeung C, Gorlick R, Hewitt SM and Helman LJ: The
membrane-cytoskeleton linker ezrin is necessary for osteosarcoma
metastasis. Nat Med. 10:182–186. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Watanabe A, Tagawa H, Yamashita J, Teshima
K, Nara M, Iwamoto K, Kume M, Kameoka Y, Takahashi N, Nakagawa T,
et al: The role of microRNA-150 as a tumor suppressor in malignant
lymphoma. Leukemia. 25:1324–1334. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yokobori T, Suzuki S, Tanaka N, Inose T,
Sohda M, Sano A, Sakai M, Nakajima M, Miyazaki T, Kato H and Kuwano
H: MiR-150 is associated with poor prognosis in esophageal squamous
cell carcinoma via targeting the EMT inducer ZEB1. Cancer Sci.
104:48–54. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Miao J, Wu S, Peng Z, Tania M and Zhang C:
MicroRNAs in osteosarcoma: Diagnostic and therapeutic aspects.
Tumour Biol. 34:2093–2098. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Cai H, Lin L, Cai H, Tang M and Wang Z:
Prognostic evaluation of microRNA-210 expression in pediatric
osteosarcoma. Med Oncol. 30:4992013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhao H, Guo M, Zhao G, Ma Q, Ma B, Qiu X
and Fan Q: miR-183 inhibits the metastasis of osteosarcoma via
downregulation of the expression of Ezrin in F5M2 cells. Int J Mol
Med. 30:1013–1020. 2012.PubMed/NCBI
|
|
33
|
Zhu J, Feng Y, Ke Z, Yang Z, Zhou J, Huang
X and Wang L: Down-regulation of miR-183 promotes migration and
invasion of osteosarcoma by targeting Ezrin. Am J Pathol.
180:2440–2451. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Neisch AL and Fehon RG: Ezrin, radixin and
moesin: Key regulators of membrane-cortex interactions and
signaling. Curr Opin Cell Biol. 23:377–382. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Clark JC, Dass CR and Choong PF: A review
of clinical and molecular prognostic factors in osteosarcoma. J
Cancer Res Clin Oncol. 134:281–297. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kafchinski LA and Jones KB: MicroRNAs in
osteosarcomagenesis. Adv Exp Med Biol. 804:119–127. 2014.
View Article : Google Scholar : PubMed/NCBI
|