Introduction
Breast cancer is one of the most common cancers in
women, and is the leading cause of cancer-induced mortality
(1). With the development of
therapeutic methods, including surgical resection, radiotherapy and
particularly chemotherapy, the outcomes in breast cancer patients
have improved in the last decades (2,3). Multiple
anti-cancer drugs have been developed for chemotherapy in cancer
therapy. As a natural compound isolated from the bark of the
Taxus baccata plant, paclitaxel could bind to the β-tubulin
subunit of microtubules, thereby stabilizing it and forcing
dividing cancer cells to arrest in the M phase of the cell cycle,
thus promoting cell death (4).
Paclitaxel has been successfully used in clinical treatment for
>2 decades, which significantly improved the outcomes of breast
cancer patients (4).
However, acquired drug resistance during treatment
occurs in numerous patients and contributes to 90% of the
chemotherapy failures (5). Thus, to
explore how cancer cells can eliminate the damage effect of
chemotherapeutic drugs such as paclitaxel and improve drug
sensitivity could be important for the treatment of cancers. A
crucial cause of drug resistance may be the increased expression of
efflux pumps (6). Certain
transmembrane proteins could transport drugs out of cancer cells.
It has been reported that human transporter associated with antigen
processing-like, also known as ATP binding cassette (ABC) B9, and
other members of the ABC transporter family could mainly contribute
to paclitaxel resistance (7,8). In addition, increased metabolism of
chemotherapeutic agents by cytochrome p450, various β-tubulin
mutations and other mechanisms could together contribute to the
resistance to paclitaxel and other chemotherapeutic agents
(9,10). Therefore, reducing drug resistance may
improve the treatment of breast and other cancers.
MicroRNAs (miRs) are a class of small non-coding
RNAs that can control the expression of multiple genes via binding
to their 3′-untranslated region (UTR) and consequently inducing
post-transcriptional repression or degradation of the messenger
RNAs (mRNAs) (11). Recent studies
have reported that microRNAs are important in drug resistance
(12,13). For example, miR-29 could modulate
collagen type I alpha 1 expression to improve cisplatin-induced
cytotoxicity in ovarian cancer cells (14), and miR-301a increased doxorubicin
resistance in osteosarcoma cells by regulating AMP-activated
protein kinase α1 (15). In addition,
paclitaxel resistance could be also regulated by miRs (16).
miR-24 has been widely reported to be involved in
multiple cancers and to mediate tumor growth and apoptosis
(17,18). However, the role of miR-24 in cancer
is complex and remains unclear. In the present study, it was
observed that miR-24 was significantly decreased in
paclitaxel-resistant (PR) MCF-7 human breast cancer cells. To
further investigate the role of miR-24 in drug resistance of breast
cancer, MCF-7/PR cells were established, and the effect of miR-24
on the sensitivity to paclitaxel of MCF-7/PR cells was studied both
in vitro and in vivo.
Materials and methods
Cell culture and establishment of PR
cell lines
MCF-7 and SKBR3 human breast cancer cells were
obtained from the American Type Culture Collection (Manassas, VA,
USA). Cells were cultured in Dulbecco's modified Eagle medium
(containing 25 mM glucose and 10% fetal bovine serum) (GE
Healthcare Life Sciences HyClone Laboratories, Logan, UT, USA) at
37°C and 5% CO2. To establish PR cell lines, the cells
were incubated with 1 µM paclitaxel (Sigma-Aldrich; Merck
Millipore, Darmstadt, Germany) for different days. The culture
medium was changed every 2 days in the presence of 1 µM
paclitaxel.
miR-24 mimic and miR-24 inhibitor
transfection
MCF-7 and MCF-7/PR cells were transfected with
miR-24 mimic or miR-24 inhibitor using Lipofectamine 2000
(Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
according to the manufacturer's protocol. A nonspecific control
(Shanghai GenePharma Co., Ltd., Shanghai, China) was used for the
control cells. The sequences were as follows: miR-24 mimic sense,
5′-uggcucaguucagcaggaacag-3′ and antisense,
5′-guuccugcugaacugagccauu-3′; and miR-24 inhibitor,
5′-cuguuccugcugaacugagcca-3′. Upon transfection, the mRNA
expression of miR-24 was measured by quantitative polymerase chain
reaction (qPCR) assay.
3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2-H-tetrazoliumbromide
(MTT) and reverse transcription (RT)-PCR assay
The half maximal inhibitory concentration
(IC50) of paclitaxel was calculated by cell viability
using an MTT assay. After the cells were treated with 1 µM
paclitaxel for 60 days to induce drug resistance, miR-24 mimic was
pre-transfected into passage-60 cells for 24 h. Then, different
concentrations of paclitaxel were added to the culture medium for
another 24 h, and the cell viability was determined as described
previously (19). RT-qPCR was
conducted to determine the expression level of miR-24 as previously
described (20). The primers for
miR-24 were those present in TaqMan™ MicroRNA Assays (Applied
Biosystems; Thermo Fisher Scientific, Inc.), and the microRNA
expression was normalized to U6 small nuclear RNA (RNU6) expression
according to the ΔΔCq method, as previously described
(21). Other primers used were: ABCB9
forward, 5′-GCTCTGGGAGAGACCTTCCT-3′ and reverse
5′-GAGCGGAAGAGACAGTTTCG-3′; glyceraldehyde 3-phosphate
dehydrogenase forward, 5′-CCCACTCCTCCACCTTTGAC-3′ and reverse,
5′-CATACCAGGAAATGAGCTTGACAA-3′; and RNU6 forward,
5′-CTCGGTTCGGCAGCACA-3′ and reverse,
5′-AACGCTTCACGAATTTGCGT-3′.
Hoechst staining and western blot
analysis
After transfection with miR-24 mimic for 24 h, the
MCF-7 and MCF-7/PR cells were treated with 10 µM paclitaxel for
another 24 h. Then, Hoechst 33342 staining was performed as
previously described (19). For
western blot analysis, following different treatments, MCF-7 and
MCF-7/PR cells were lysed, and western blotting was performed as
previously described (19). All
western blot analyses were performed ≥3 times. The blots were
analyzed by Quantity One v4.62 software (National Institutes of
Health, Bethesda, MD, USA). Primary antibodies were all purchased
from Santa Cruz Biotechnology, Inc. (Dallas, TX, USA), including
anti-B-cell lymphoma (Bcl)-2 (cat. no. sc-492; dilution 1:500),
anti-Bcl-2-associated × protein (Bax) (cat. no. sc-526; dilution
1:1,000), anti-cleaved caspase-3 (cat. no. sc-7148; dilution
1:1,000), anti-ABCB9 (cat. no.sc-46744; dilution 1:1,000) and
anti-β-actin (cat. no. sc-1616; dilution 1:5,000).
In vitro scratch assay
MCF-7 and MCF-7/PR cells were seeded in 12-well
plates and grown in complete medium. The cells were transfected
with miR-24 mimic and incubated with 10 µM paclitaxel. After 48 h
of incubation, the confluent monolayer cells were scraped by
pipette tips to generate a straight line scratch. Then, the wells
were washed and replaced with serum-free medium in the presence of
paclitaxel. The cells in the scratch were imaged under a microscope
(DP70; Olympus Corporation, Tokyo, Japan) after 12 h, and the
images were used to calculate the distances between the two edges
of the scratch.
Luciferase assay
The miR-24 response element (wild type or mutated)
in the 3′-UTR of ABCB9 was transfected into the pMIR-reporter
plasmid (Ambion, Austin, TX, USA), which contained the luciferase
reporter gene. MCF-7 cells were co-transfected with luciferase
reporter constructs containing wild type or mutated type ABCB9
3′-UTR and miR-24 mimic in the presence/absence of miR-24
inhibitor. A luciferase assay kit (Promega Corporation, Madison,
WI, USA) was used to analyze the luciferase activities according to
the manufacturer's protocol.
Study population
A total of 20 human breast cancer tissues were
selected from breast cancer patients that exhibited poor
chemo-response after undergoing surgery at The Affiliated Cancer
Hospital of Nanjing Medical University (Nanjing, China) between
July 2010 and June 2014. Another group of patients (n=20) with a
good chemo-response was recruited as control. Tumor tissues were
washed and stored in liquid nitrogen prior to the experiments. In
combination with the pathological diagnosis, >90% necrosis was
considered drug-sensitive, while tumor necrosis <90% was
considered poor responder. The total protein and total RNA from
tissues were isolated using TRIzol reagent (Takara Bio Inc., Otsu,
Japan), and detected by western blotting and qPCR analysis as
previously described (19,20). The present study was approved by the
Medical Ethics Committee of Jiangsu Cancer Hospital (Nanjing,
China), and written informed consent was obtained from all
participants.
Animal models
A total of 40 female 8-week-old nude BALB/c mice
(20–22 g) were purchased from the Model Animal Research Center of
Nanjing University (Nanjing, China). Mice were maintained at 25°C
and exposed to 12 h light/dark cycles with free access to food and
water. Prior to the experiment, mice were allowed to acclimate for
5 days. A cell suspension of MCF-7, MCF-7/PR or miR-24
mimic-pretreated MCF-7/PR cells was prepared (108
cells/ml). Then, the cell suspension was subcutaneously injected
into the right upper flank of the 40 nude mice. When tumors mass
reached 50 mm in diameter, 10 mg/kg paclitaxel was
intraperitoneally injected once every 2 days. Tumor volume was
recorded and calculated as (r2xl)/2, where r and l are,
respectively, the short and long diameter of the tumors. After 14
days of administration, the animals were sacrificed and tumor
tissue was removed. All animal studies were carried out under the
supervision of the Committee for Animal Experiments of Nanjing
Medical University (Nanjing, China).
Terminal deoxynucleotidyl
transferase-mediated dUTP nick end labeling (TUNEL) staining
Tumor tissues were fixed in 4% paraformaldehyde for
paraffin sections. Then, a TUNEL kit (Roche Diagnostics,
Indianapolis, IN, USA) was used to stain the apoptotic cells
according to the manufacturer's protocol. The apoptotic ratio was
calculated as the number of TUNEL-positive cells divided the by
total number of cells.
Statistical analysis
Data were expressed as the mean ± standard
deviation. All statistical analysis was performed using PASW
Statistics 18.0 software (SPSS, Inc., Chicago, IL, USA).
Statistical comparisons between groups were analyzed using the t
test and one-way analysis of variance. P<0.05 was considered to
indicate a statistically significant difference.
Results
Overexpression of miR-24 increases the
sensitivity to paclitaxel of breast cancer cell lines
After the MCF-7 breast cancer cells were treated for
60 days with 1 µM paclitaxel, the IC50 of paclitaxel in
MCF-7/PR cells increased by 16-fold compared with that in normal
MCF-7 cells (Fig. 1A). In this PR
cell line, the expression of miR-24 decreased in a time-dependent
manner (Fig. 1B). Thus, to further
investigate the effect of miR-24 on the cytotoxicity of paclitaxel,
a microRNA mimic was used to overexpress miR-24 in MCF-7 (Fig. 1C) and other breast cancer cell lines
(data not shown). It was observed that overexpression of miR-24 did
not affect the IC50 of paclitaxel in normal MCF-7 cells
(IC50 changed from 2.627 to 3.429 µM, Fig. 1D). On the contrary, using a microRNA
mimic to increase the miR-24 expression level in MCF-7/PR cells
could obviously improve the sensitivity toward paclitaxel of this
drug-resistant cell model (IC50 changed from 8.944 to
48.760 µM, Fig. 1D). Similar results
were also observed in SKBR3 breast cancer cells (Fig. 1E and F).
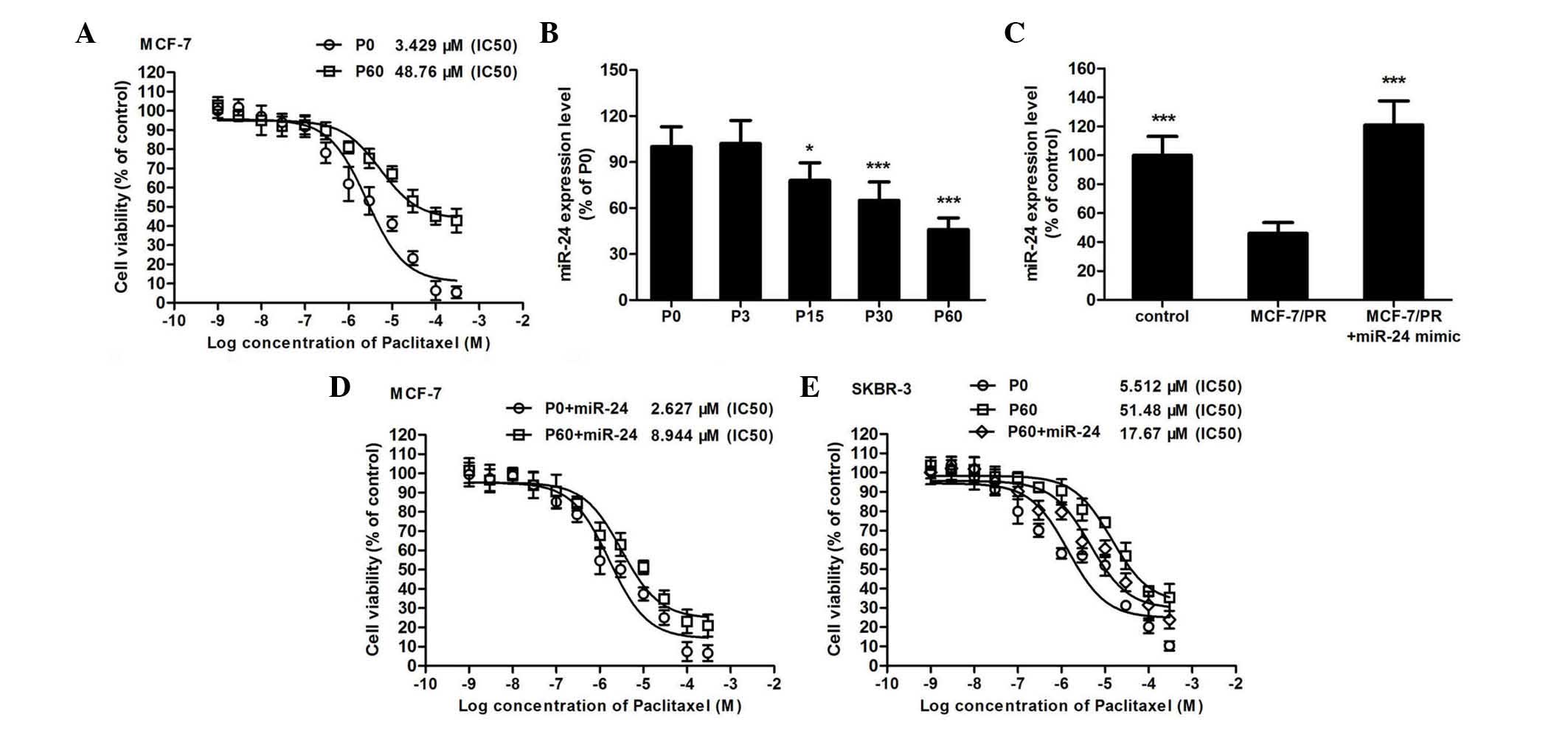 | Figure 1.Overexpression of miR-24 improves the
sensitivity toward paclitaxel of breast cancer cells. (A) The
cytotoxicity of paclitaxel on MCF-7 (P0) and MCF-7/PR (P60) cells
was determined by
3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2-H-tetrazolium bromide
assay (n=6). (B) Upon induction of paclitaxel resistance in MCF-7
cells for different days, the expression of miR-24 was determined
by qPCR (*P<0.05, ***P<0.001 compared with P0 cells, n=6).
(C) Using a microRNA mimic to overexpress miR-24 in MCF-7/PR cells,
the expression of miR-24 was detected by qPCR in normal MCF-7 and
MCF-7/PR cells (***P<0.001 compared with MCF-7/PR cells, n=6).
(D) The miR-24 mimic was added to MCF-7 or MCF-7/PR cells, and then
the IC50 of paclitaxel in each cell model was determined
(n=6). (E) The miR-24 mimic was added to SKBR3 or SKBR3/PR cells,
and then the IC50 of paclitaxel to each cell model was
determined (n=6). PR, paclitaxel-resistant; miR, microRNA;
IC50, half maximal inhibitory concentration; qPCR,
quantitative polymerase chain reaction; P, passage. |
Overexpression of miR-24 increases
paclitaxel-induced apoptosis and inhibition of migration in
MCF-7/PR cells via regulation of ABCB9 expression. As miR-24 could
increase the cytotoxic effect of paclitaxel on breast cancer cells,
the present study further investigated the effect of miR-24 on
paclitaxel-induced cell apoptosis and inhibition of migration.
Using Hoechst 33342 staining, paclitaxel-induced apoptosis was
determined. The apoptotic cells displayed highly bright and shrunk
nuclei (Fig. 2A). The apoptotic rate
in 10 µM paclitaxel-treated MCF-7 cells reached 40%, which was
4-fold higher than that in control cells (Fig. 2B). However, this cytotoxic effect was
not obvious in MCF-7/PR cells (Fig.
2B). Using a miR-24 mimic, the cytotoxicity of 10 µM paclitaxel
on MCF-7/PR cells was partially recovered (Fig. 2B). Furthermore, it was noticed that 10
µM paclitaxel could reduce the expression of the anti-apoptotic
protein Bcl-2, increase the expression of the apoptotic marker Bax
and activate caspase-3 in normal MCF-7 cells, while these effects
were suppressed in MCF-7/PR cells (Fig.
2C). However, increased miR-24 expression level restored the
pro-apoptotic effect of paclitaxel on drug-resistant cells
(Fig. 2C). In addition, the effect of
miR-24 on the paclitaxel-regulated migration of MCF-7 cells was
also detected. Scratch test revealed that the inhibitory effect of
10 µM paclitaxel on the migration of MCF-7/PR cells was not as
obvious as that observed in normal MCF-7 cells (Fig. 2C). However, in accordance with the
results of the apoptotic experiments, overexpression of miR-24
improved the inhibitory effect of paclitaxel on the migration of
MCF-7/PR cells (Fig. 2D).
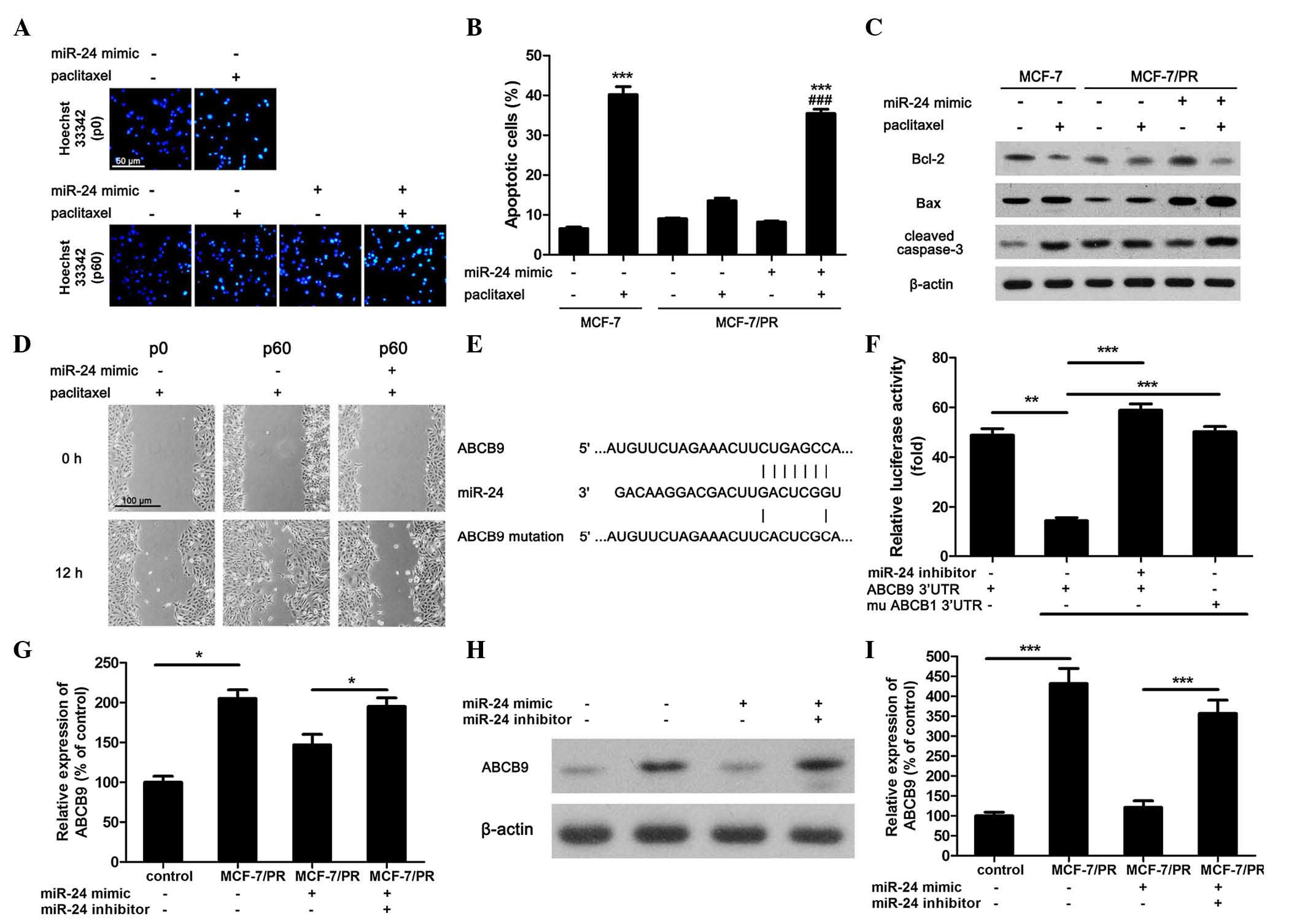 | Figure 2.Overexpression of miR-24 increases
paclitaxel-induced apoptosis and inhibition of migration in
MCF-7/PR cells via regulation of ABCB9 expression. (A) Hoechst
33342 staining was used to mark apoptotic cells, and images were
recorded by fluorescence microscopy (scale bar=50 µm). (B) The
apoptotic-positive cells were counted and divided by the total cell
number to calculate the apoptotic ratio. In total, 15 random images
from six parallel wells were selected (***P<0.001 compared with
control MCF-7 cells, ###P<0.001 compared with
paclitaxel-treated MCF-7/PR cells, n=6). (C) The expression of
apoptotic markers in different groups was determined by western
blotting. β-actin was used as internal reference. All blots were
repeated ≥3 times. (D) The migration of cells was investigated by
scratch test (scale bar=100 µm). (E) Prediction of the miR-24
binding site on ABCB9 and ABCB9 mutation site. (F) Luciferase assay
was performed to detect whether miR-24 could directly binding to
the 3′-UTR region of ABCB9 (**P<0.01, ***P<0.001, n=6). (G)
The messenger RNA expression of ABCB9 was determined by
quantitative polymerase chain reaction (*P<0.01, n=6). (H) The
protein expression of ABCB9 was determined by western blotting. (I)
The blot was analyzed by optical density (***P<0.001, n=3). PR,
paclitaxel-resistant; miR, microRNA; Bcl-2, B-cell lymphoma-2; Bax,
Bcl-2-associated X protein; ABC, ATP-binding cassette; mu, mutant;
UTR, untranslated region. |
The above results demonstrated that upregulating
miR-24 expression did not affect normal MCF-7 cells survival
(Fig. 1D). Thus, it was speculated
that miR-24 may increase the sensitivity to paclitaxel of MCF-7/PR
cells through regulating efflux pumps or reducing the metabolism of
the compound. Using online microRNA target prediction tools
[including TargetScanHuman 6.2 (http://www.targetscan.org/vert_71/), miRBase
(http://www.mirbase.org/) and starBase (http://starbase.sysu.edu.cn/)], it was predicted that
the 3′-UTR region of ABCB9 has possible binding positions for
miR-24 (Fig. 2E). To confirm this
concept, a miR-24 mimic and a ABCB9 3′-UTR wild type or mutated
luciferase reporter were co-transfected into MCF-7/PR cells. It was
observed that the miR-24 mimic decreased the luciferase activity of
ABCB9 3′-UTR wild type-transfected cells, but not that of cells
transfected with the mutated version (Fig. 2F). In addition, using a designed
miR-24 inhibitor, ABCB9 3′-UTR wild type yielded similar results
compared with the mutated version (Fig.
2F). The expression of ABCB9 was next determined at both the
mRNA and protein level by qPCR and western blot analysis,
respectively. It was observed that the expression of ABCB9
increased significantly in MCF-7/PR cells in comparison with that
in normal MCF-7 cells, but miR-24 mimic reduced ABCB9 expression at
both the mRNA and protein level (Fig.
2F-H). Meanwhile, miR-24 inhibitor could abolish this
effect.
Expression of miR-24 and ABCB9 in
drug-resistant breast cancer patients
As miR-24 and ABCB9 are important in paclitaxel
resistance of breast cancer cells, the expression level of miR-24
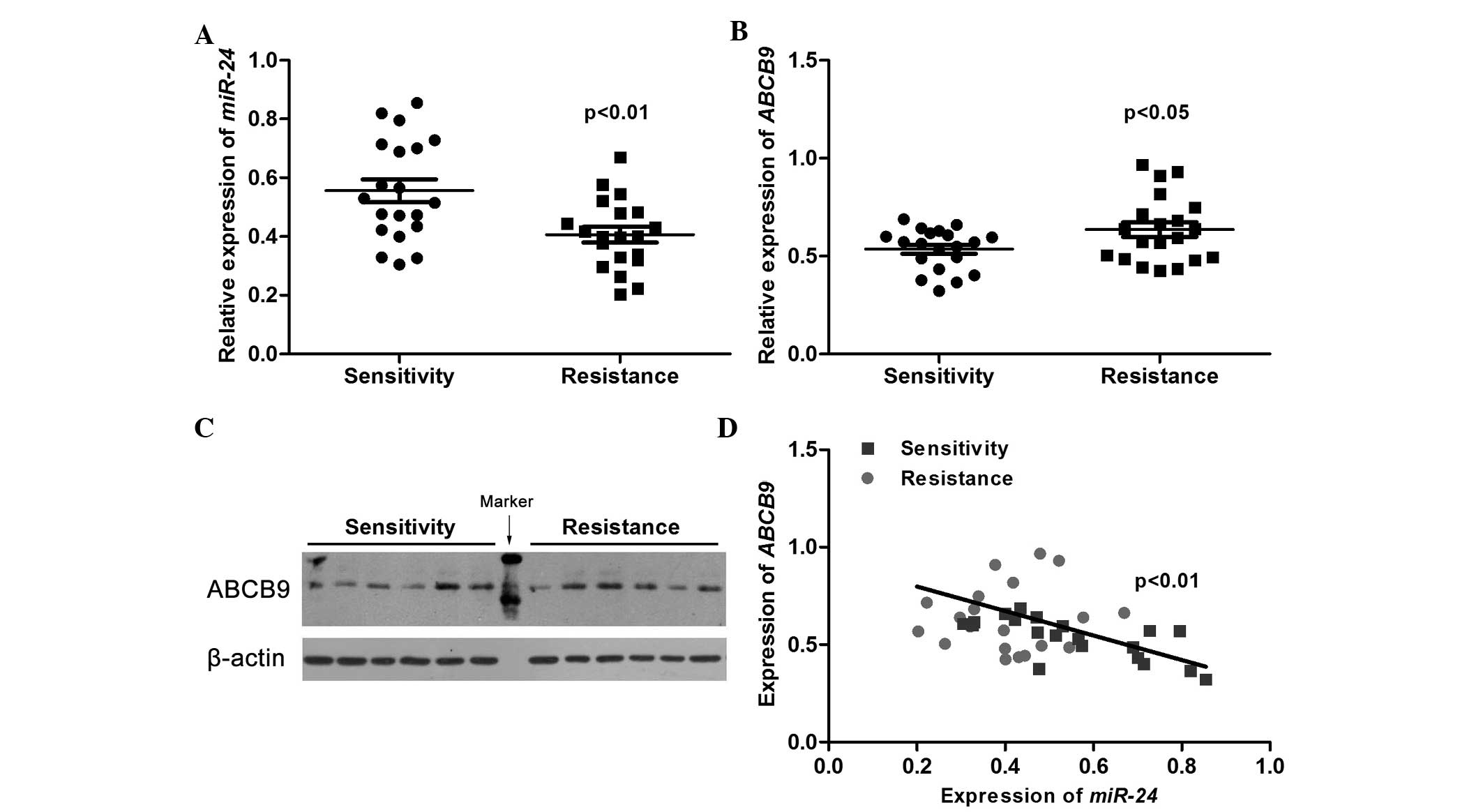
and ABCB9 in PR breast cancer patients was examined. Fig. 3A indicates that the miR-24 level was
significantly decreased in tumor tissues from breast cancer
patients with poor response to paclitaxel compared with that
observed in tumor tissues from patients with good drug sensitivity.
On the contrary, the expression of ABCB9 in the drug-resistant
group was significantly higher than that in patients with good drug
sensitivity (Fig. 3B). This result
was also verified by western blotting (Fig. 3C). Furthermore, correlation analysis
revealed that the expression of miR-24 was negatively correlated
with the expression of ABCB9 (Fig.
3D), which supported the evidence that miR-24 downregulated the
expression of ABCB9, thereby increasing drug sensitivity.
Overexpression of miR-24 increases the
sensitivity of paclitaxel to MCF-7/PR cells in vivo
The effect of miR-24 on the sensitivity toward
paclitaxel of MCF-7/PR cells in vivo was next investigated
using a nude mouse model. Using a miR-24 mimic, a high expression
level of miR-24 could be sustained for >2 weeks (data not
shown). It was observed that 10 mg/kg paclitaxel could
significantly reduce tumor growth in MCF-7-implanted nude mice
(Fig. 4A). However, paclitaxel at
this dose was not able to affect the growth of tumors derived from
MCF-7/PR cells (Fig. 4A). On the
contrary, when MCF-7/PR cells were pre-transfected with a miR-24
mimic, the anti-tumor effect of 10 mg/kg paclitaxel was partially
recovered (Fig. 4A). Apoptosis in
tumor tissues was detected by TUNEL assay. Following 2 weeks of
treatment, 10 mg/kg paclitaxel could markedly induce tumor cell
apoptosis (Fig. 4B and C). In
accordance with the results of the experiment performed in
vitro, paclitaxel induced less apoptosis in MCF-7/PR
cell-implanted tumor tissue than that in the tissues of normal
MCF-7 cell-implanted nude mice (Fig. 4B
and C). Furthermore, pre-transfection with the miR-24 mimic
could recover the pro-apoptotic effect of paclitaxel in
vivo.
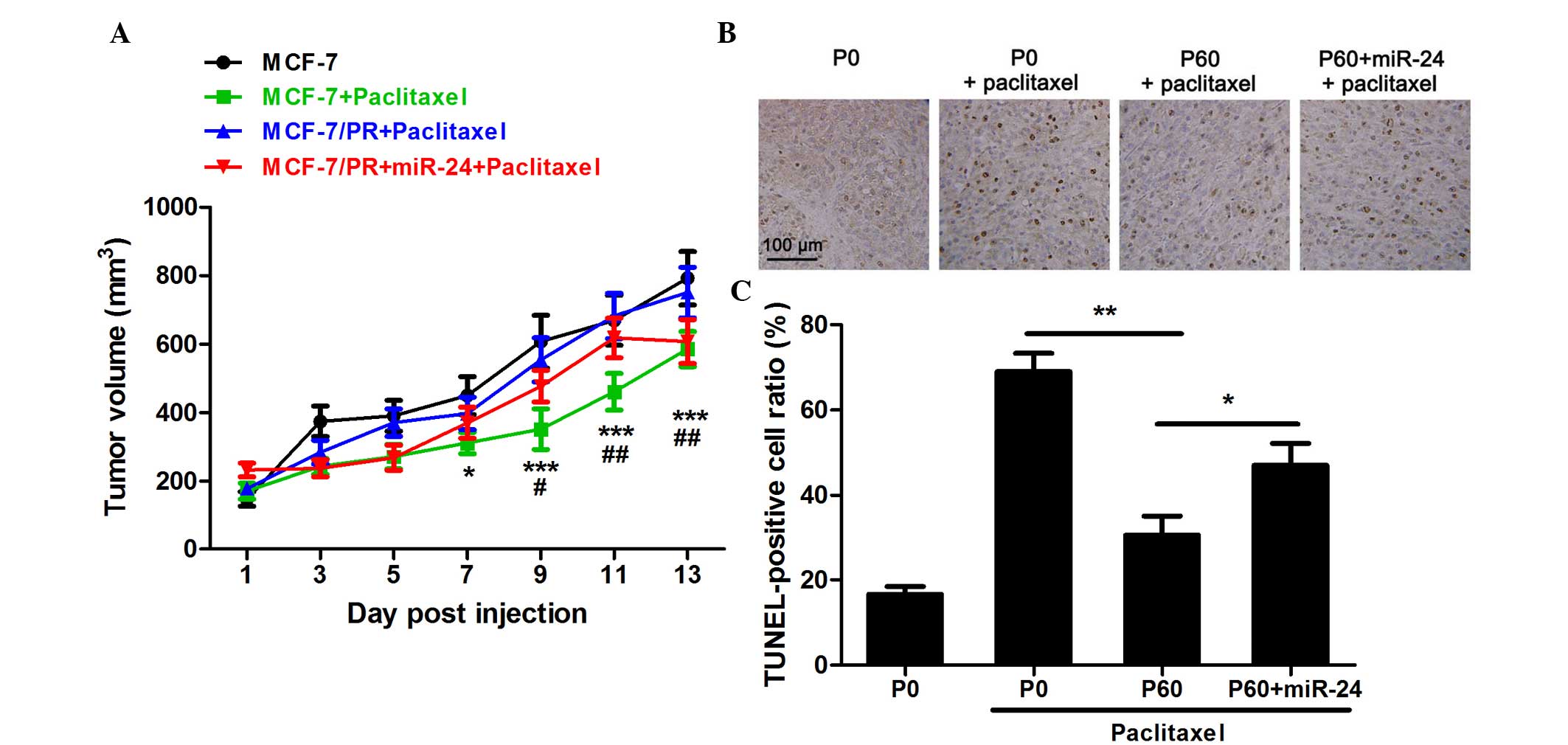 | Figure 4.Effect of paclitaxel on
miR-24-overexpressing MCF-7/PR cells in implanted nude mice. (A)
Tumor growth in nude mice was determined every 2 days. *P<0.05,
***P<0.001 in the MCF-7/PR-miR-24 group treated with paclitaxel
compared with the control MCF-7 group; #P<0.05,
##P<0.01 in the MCF-7/PR-miR-24 group treated with
paclitaxel compared with the MCF-7/PR group treated with
paclitaxel, n=8. (B) TUNEL assay was performed to observe the
apoptotic cells. Apoptotic nuclei exhibited brown staining, while
all nuclei were stained by hematoxylin with blue dye. Scale bar=100
µm. (C) The apoptotic ratio was calculated as the number of
TUNEL-positive cells divided by the total cell number. *P<0.05,
**P<0.01, n=8. PR, paclitaxel-resistant; miR, microRNA; P,
passage; TUNEL, terminal deoxynucleotidyl transferase-mediated dUTP
nick end labeling. |
Discussion
Drug resistance in breast cancer has been regarded
as one of the primary obstacles contributing to chemotherapy
failure (22). Thus, it is necessary
to explore novel therapeutic methods to reduce drug resistance for
the treatment of breast cancer. In the present study, it was
observed that the expression of miR-24 was significantly reduced in
tumor tissues from PR breast cancer patients and PR breast cancer
cell lines. Further in vitro experiments confirmed that
upregulating miR-24 expression could partially recover the
sensitivity to paclitaxel of breast cancer cells. Therefore, miR-24
may be a useful target to reduce drug resistance and a possible
marker to guide clinical medication.
miR-24 has been reported to be an oncogene in oral
carcinoma (23), prostate cancer
(24) and lung cancer (25). In addition, miR-24 could promote cell
proliferation in lung cancer (25)
and hepatocellular carcinoma (26).
However, miR-24 could inhibit cell proliferation in gastric cancer
(27,28), and acts as a tumor suppressor in colon
cancer (29). In breast cancer, the
role of miR-24 also remains controversial. The expression of miR-24
in serum from early breast cancer patients was decreased in a
previous study (30), but another
study using paraffin-embedded tissues observed that the expression
of miR-24 was upregulated (31). In
addition, Du et al reported that the expression of miR-24
was higher in breast cancer samples than that in benign breast
tissues, and speculated that miR-24 may enhance tumor invasion
(32). Therefore, as an important
microRNA associated with cancer development, the role of miR-24 in
breast and other cancers should be further investigated.
Using microRNA target prediction tools, the present
authors identified that ABCB9 could be one of the target genes of
miR-24. ABCB9 is a member of the ABC sub-family B, which has been
reported to be involved in drug intracellular trafficking, thereby
affecting chemotherapy-related multidrug resistance (MDR) (33,34). Thus,
it was speculated that miR-24 may affect the expression of ABCB9 to
regulate the sensitivity toward paclitaxel of breast cancer cells.
It was identified that miR-24 could directly bind to the 3′-UTR of
ABCB9, thereby inhibiting the translation of ABCB9. These results
were in accordance with those from previous studies (35,36).
Overall, the above results indicated that downregulation of ABCB9
may contribute to the effect of miR-24 on the sensitivity toward
paclitaxel of breast cancer cells. Furthermore, the present study
confirmed that ABCB9 was also upregulated in chemotherapy-resistant
breast cancer patients, which was negatively correlated with the
level of miR-24, suggesting that miR-24 was associated with drug
resistance in breast cancer via regulating ABCB9 expression. Unlike
other members of the ABC transporter protein family, the regulatory
effect of ABCB9 on drug resistance has not been frequently reported
(34,36). The present study provided novel
evidence regarding the role of ABCB9 in chemotherapy-related MDR.
Furthermore, the role of miR-24 in PR breast cancer cells was
evaluated in vivo using a nude mouse model. It was observed
that miR-24 increased the sensitivity to paclitaxel of
MCF-7/PR-implanted nude mice. Although multiple mechanisms may be
involved, including p450-related drug metabolism and various
β-tubulin mutations, which require further investigation in
miR-24-associated drug resistance, the present results suggested
that miR-24 could be important in drug resistance of breast cancer
cells, and could improve the sensitivity toward paclitaxel both
in vitro and in vivo.
In conclusion, the present study demonstrated for
the first time that miR-24 improves the sensitivity toward
paclitaxel of breast cancer cells, suggested an important novel
function for miR-24, and provided a possible microRNA target and a
useful marker in the clinical treatment of breast cancer.
Acknowledgements
The present study was supported by a grant from
Jiangsu Cancer Hospital (Nanjing, China; grant no. ZM201105).
References
|
1
|
Bottos A and Hynes NE: Cancer: Staying
together on the road to metastasis. Nature. 514:309–310. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Xuan Q, Ji H, Tao X, Xu Y and Zhang Q:
Quantitative assessment of HER2 amplification in HER2-positive
breast cancer: Its association with clinical outcomes. Breast
Cancer Res Treat. 150:581–588. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wang Y, Zhang Y, Pan C, Ma F and Zhang S:
Prediction of poor prognosis in breast cancer patients based on
microRNA-21 expression: A meta-analysis. PLoS One. 10:e01186472015.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Grobosch T, Schwarze B, Stoecklein D and
Binscheck T: Fatal poisoning with Taxus baccata: Quantification of
paclitaxel (taxol A), 10-deacetyltaxol, baccatin III,
10-deacetylbaccatin III, cephalomannine (taxol B), and
3,5-dimethoxyphenol in body fluids by liquid chromatography-tandem
mass spectrometry. J Anal Toxicol. 36:36–43. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Johnson MT, Reichley R, Hoppe-Bauer J,
Dunne WM, Micek S and Kollef M: Impact of previous antibiotic
therapy on outcome of Gram-negative severe sepsis. Crit Care Med.
39:1859–1865. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ganoth A, Merimi KC and Peer D: Overcoming
multidrug resistance with nanomedicines. Expert Opin Drug Deliv.
12:223–238. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ohashi-Kobayashi A, Ohashi K, Du WB, Omote
H, Nakamoto R, Al-Shawi M and Maeda M: Examination of drug
resistance activity of human TAP-like (ABCB9) expressed in yeast.
Biochem Biophys Res Commun. 343:597–601. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kaur P, Garg T, Rath G, Murthy RS and
Goyal AK: Development, optimization and evaluation of
surfactant-based pulmonary nanolipid carrier system of paclitaxel
for the management of drug resistance lung cancer using Box-Behnken
design. Drug Deliv. 23:1912–1925. 2016.PubMed/NCBI
|
|
9
|
Zhu Z, Mu Y, Qi C, Wang J, Xi G, Guo J, Mi
R and Zhao F: CYP1B1 enhances the resistance of epithelial ovarian
cancer cells to paclitaxel in vivo and in vitro. Int J Mol Med.
35:340–348. 2015.PubMed/NCBI
|
|
10
|
Verma K and Ramanathan K: Investigation of
paclitaxel resistant R306C mutation in β-tubulin-a computational
approach. J Cell Biochem. 116:1318–1324. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Png KJ, Halberg N, Yoshida M and Tavazoie
SF: A microRNA regulon that mediates endothelial recruitment and
metastasis by cancer cells. Nature. 481:190–194. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Shibayama Y, Kondo T, Ohya H, Fujisawa SI,
Teshima T and Iseki K: Upregulation of microRNA-126-5p is
associated with drug resistance to cytarabine and poor prognosis in
AML patients. Oncol Rep. 33:2176–2182. 2015.PubMed/NCBI
|
|
13
|
Acunzo M, Romano G, Wernicke D and Croce
CM: MicroRNA and cancer-a brief overview. Adv Biol Regul. 57:1–9.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yu PN, Yan MD, Lai HC, Huang RL, Chou YC,
Lin WC, Yeh LT and Lin YW: Downregulation of miR-29 contributes to
cisplatin resistance of ovarian cancer cells. Int J Cancer.
134:542–551. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang Y, Duan G and Feng S: MicroRNA-301a
modulates doxorubicin resistance in osteosarcoma cells by targeting
AMP-activated protein kinase alpha 1. Biochem Biophys Res Commun.
459:367–373. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chatterjee A, Chattopadhyay D and
Chakrabarti G: miR-17-5p downregulation contributes to paclitaxel
resistance of lung cancer cells through altering beclin1
expression. PLoS One. 9:e957162014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gao Y, Liu Y, Du L, Li J, Qu A, Zhang X,
Wang L and Wang C: Downregulation of miR-24-3p in colorectal cancer
is associated with malignant behavior. Med Oncol. 32:3622015.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Guo C, Deng Y, Liu J and Qian L:
Cardiomyocyte-specific role of miR-24 in promoting cell survival. J
Cell Mol Med. 19:103–112. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang T, Sun P, Chen L, Huang Q, Chen K,
Jia Q, Li Y and Wang H: Cinnamtannin D-1 protects pancreatic
β-cells from palmitic acid-induced apoptosis by attenuating
oxidative stress. J Agric Food Chem. 62:5038–5045. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wu J, Sun P, Zhang X, Liu H, Jiang H, Zhu
W and Wang H: Inhibition of GPR40 protects MIN6 β cells from
palmitate- induced ER stress and apoptosis. J Cell Biochem.
113:1152–1158. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang XW, Xi XQ, Wu J, Wan YY, Hui HX and
Cao XF: microRNA-206 attenuates tumor proliferation and migration
involving the downregulation of NOTCH3 in colorectal cancer. Oncol
Rep. 33:1402–1410. 2015.PubMed/NCBI
|
|
22
|
Huang J, Li H and Ren G:
Epithelial-mesenchymal transition and drug resistance in breast
cancer (Review). Int J Oncol. 47:840–848. 2015.PubMed/NCBI
|
|
23
|
Lin SC, Liu CJ, Lin JA, Chiang WF, Hung PS
and Chang KW: miR-24 up-regulation in oral carcinoma: Positive
association from clinical and in vitro analysis. Oral Oncol.
46:204–208. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Qin W, Shi Y, Zhao B, Yao C, Jin L, Ma J
and Jin Y: miR-24 regulates apoptosis by targeting the open reading
frame (ORF) region of FAF1 in cancer cells. PLoS One. 5:e94292010.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhao G, Liu L, Zhao T, Jin S, Jiang S, Cao
S, Han J, Xin Y, Dong Q, Liu X and Cui J: Upregulation of miR-24
promotes cell proliferation by targeting NAIF1 in non-small cell
lung cancer. Tumour Biol. 36:3693–3701. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu YX, Long XD, Xi ZF, Ma Y, Huang XY,
Yao JG, Wang C, Xing TY and Xia Q: MicroRNA-24 modulates aflatoxin
B1-related hepatocellular carcinoma prognosis and tumorigenesis.
Biomed Res Int. 2014:4829262014.PubMed/NCBI
|
|
27
|
Duan Y, Hu L, Liu B, Yu B, Li J, Yan M, Yu
Y, Li C, Su L, Zhu Z, et al: Tumor suppressor miR-24 restrains
gastric cancer progression by downregulating RegIV. Mol Cancer.
13:1272014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wu J, Zhang YC, Suo WH, Liu XB, Shen WW,
Tian H and Fu GH: Induction of anion exchanger-1 translation and
its opposite roles in the carcinogenesis of gastric cancer cells
and differentiation of K562 cells. Oncogene. 29:1987–1996. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Mishra PJ, Song B, Mishra PJ, Wang Y,
Humeniuk R, Banerjee D, Merlino G, Ju J and Bertino JR: MiR-24
tumor suppressor activity is regulated independent of p53 and
through a target site polymorphism. PLoS One. 4:e84452009.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sochor M, Basova P, Pesta M, Dusilkova N,
Bartos J, Burda P, Pospisil V and Stopka T: Oncogenic microRNAs:
miR-155, miR-19a, miR-181b and miR-24 enable monitoring of early
breast cancer in serum. BMC Cancer. 14:4482014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yin JY, Deng ZQ, Liu FQ, Qian J, Lin J,
Tang Q, Wen XM, Zhou JD, Zhang YY and Zhu XW: Association between
mir-24 and mir-378 in formalin-fixed paraffin-embedded tissues of
breast cancer. Int J Clin Exp Pathol. 7:4261–4267. 2014.PubMed/NCBI
|
|
32
|
Du WW, Fang L, Li M, Yang X, Liang Y, Peng
C, Qian W, O'Malley YQ, Askeland RW, Sugg SL, et al: MicroRNA
miR-24 enhances tumor invasion and metastasis by targeting PTPN9
and PTPRF to promote EGF signaling. J Cell Sci. 126:1440–1453.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Demirel O, Bangert I, Tampé R and Abele R:
Tuning the cellular trafficking of the lysosomal peptide
transporter TAPL by its N-terminal domain. Traffic. 11:383–393.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hlavata I, Mohelnikova-Duchonova B,
Vaclavikova R, Liska V, Pitule P, Novak P, Bruha J, Vycital O,
Holubec L, Treska V, et al: The role of ABC transporters in
progression and clinical outcome of colorectal cancer. Mutagenesis.
27:187–196. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Xu F, Wang F, Yang T, Sheng Y, Zhong T and
Chen Y: Differential drug resistance acquisition to doxorubicin and
paclitaxel in breast cancer cells. Cancer Cell Int. 14:5382014.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Dong Z, Zhong Z, Yang L, Wang S and Gong
Z: MicroRNA-31 inhibits cisplatin-induced apoptosis in non-small
cell lung cancer cells by regulating the drug transporter ABCB9.
Cancer Lett. 343:249–257. 2014. View Article : Google Scholar : PubMed/NCBI
|