Introduction
Lung cancer is one of the most common malignant
tumors in the world. Its incidence rate the highest of all human
tumors and there are ~1.2 million novel cases each year (1). In addition, lung cancer also has the
highest mortality rate of all human cancers, with 15,600 people
succumbing to the disease each year, which accounts for 19.4% of
the total number of cancer-associated mortalities (1,2). For
>80% of lung cancer patients admitted to hospital, the ideal
time to receive surgery and multidisciplinary radical cure has
already passed (2,3). Although the concept and means of lung
cancer treatment in recent years has made considerable progress,
the overall effect is not satisfactory.
An association between carcinogenesis and chronic
inflammation has long been suspected (4). The accepted hypothesis is that chronic
inflammation more often stimulates than inhibits tumor development
(5–9).
As a rate-limiting enzyme, cyclooxygenase-2 (COX-2) is involved in
the conversion of arachidonic acid into prostaglandin (PG) and
other bioactive lipids. With the exception of being involved in
inflammation, COX-2 produces large amounts of PGE2 in tumor tissues
(10–13), and is extremely important in the
development of tumors (10,14–19). COX-2
inhibitors have produced encouraging results in preventing and
treating certain digestive system cancers (20–22). The
present study hypothesized that COX-2 is associated with malignant
cell phenotype in lung cancer. Therefore, the present study
investigated whether silencing of COX-2 and the use of celecoxib,
an inhibitor of COX-2, affects lung cancer cell proliferation and
invasiveness (21,22). The present results demonstrated that
silencing of COX-2 and use of celecoxib inhibited the growth of
lung cancer cells and decreased their invasive abilities. In
addition, the present results revealed that the inhibition of a
malignant cell phenotype may be associated with an alteration in
vascular endothelial growth factor (VEGF), matrix metalloproteinase
(MMP)-2 and endothelial growth factor receptor (EGFR)
expression.
Materials and methods
Cell culture
Human lung adenocarcinoma A549 and LTEP-A2 cells
were purchased from the Cell Center of Peking Union Medical College
(Beijing, China). The cells were cultured in RPMI-1640 medium
(Gibco®; Thermo Fisher Scientific, Inc., Waltham, MA,
USA) supplemented with 10% fetal bovine serum (FBS;
Gibco®), 100 U/ml of penicillin and 100 U/ml of
streptomycin in a humidified 37°C incubator with 5%
CO2.
RNA interference (RNAi) vector
transfection
The specific RNAi vector psi10 and empty vector pU6
were constructed in a previous study, in which the construction
process and the interference effect of the vectors were observed
(23). In total, 3×105
cells were seeded into 35 mm culture plates. When the cells were
70–80% confluent, the cells were transfected with plasmids (pU6 and
psi10) using Lipofectamine 2000 (Invitrogen™; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol.
Complete RPMI-1640 medium supplemented with geneticin (800 µg/ml)
was used to screen the clones. The transfected A549 and LTEP-A2
cell strains were named A549-pU6, A549-si10, LTEP-A2-pU6 and
LTEP-A2-si10.
Western blotting
Protein was extracted using conventional protocols
(9 M urea, 4%
3-[(3-cholamidopropyl)dimethylammonio]-1-propanesulfonate, 1%
dithiothreitol, 0.5% carrier ampholytes and cocktail of protease
inhibitors). Protein concentrations were determined using Pierce™
BCA Protein Assay kit (Thermo Fisher Scientific, Inc.) with bovine
serum album as a standard. Subsequently, proteins were separated on
12% polyacrylamide gels using standard sodium dodecyl
sulfate-polyacrylamide gel electrophoresis techniques and then
transferred to nitrocellulose membranes. The membranes were blocked
by 5% milk (Invitrogen; Thermo Fisher Scientific, Inc.) for 1 h and
probed with specific antibodies for 4°C overnight. Mouse anti-COX-2
polyclonal antibodies (dilution, 1 µg/ml; catalog no., 3362R-100;
BioVision, Inc., Milpitas, CA, USA) were used, and mouse
anti-glyceraldehyde 3-phosphate dehydrogenase monoclonal antibodies
(dilution, 0.3 ng/µl; catalog no., 60004–1-Ig; ProteinTech Group,
Rosemont, IL, USA) were used as an internal reference. After three
washing steps of 15 min each with Tris-buffered saline and Tween
(TBST), the membranes were probed with horseradish
peroxidase-conjugated goat anti-mouse IgG antibodies (dilution,
1:5,000; catalog no., sc-2005; Santa Cruz Biotechnology, Inc., USA)
at 37°C for 1 h. After three washing steps of 15 min each with
TBST, the protein was tested by chemiluminescence detection
(catalog no., 29100; Engreen Biosystem Co., Ltd., Beijing, China).
The protein expression was analyzed by Image-Pro Plus 6.0 software
(Media Cybernetics, Inc., Rockville, MD, USA).
Cell growth curve assays
Tumor cells (3,000 cells/well) were seeded in
flat-bottom 96-well plates. Cell activity was assessed by a
3-(4,5-dimethyl-thiazol-2yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium
(MTS) assay (Promega, Madison, WI, USA). The growth of cells was
detected for 5 days. Subsequently, 20 µl MTS was added into each
well and then incubated with the cells for 3 h. The absorbance was
recorded at 490 nm with an ELx800 Absorbance Reader (Bio-Tek
Instruments, Winooski, VT, USA). This experiment was repeated three
times.
Colony formation assays
A total of 300 cells were seeded in 6-well plates.
The cells were cultured for 10 days. On the 10th day, the number of
colonies with >50 cells was recorded using the trypan blue
exclusion method. The experiment was repeated three times in
triplicate.
Cell migration assay
Cell motility was evaluated by two experiments. In
the wound healing experiment, channels were created by making a
scratch in a 6-well plate. Subsequently, 8×105 cells per
channel were seeded into the 6-well plates and cultured for 24 h.
At 0, 12 and 24 h, the wounds were observed and pictures were taken
using a phase-contrast microscope. The areas of the scratch were
calculated using Alpha View Analysis Tools version 1.0 software
(ProteinSimple, San Jose, CA, USA), and this was used to calculate
the percentage of wound closure.
In addition, a migration experiment was performed in
a 24-well Transwell unit (Corning, Inc., Corning, NY, USA), as
previously reported (24). The cells
were starved for 12 h and 8×105 cells were seeded in the
upper compartment of the Transwell unit with serum-free medium. The
lower compartment was filled with medium containing 10% FBS.
Subsequent to 24 or 48 h, the cells in the upper chamber were
removed by gentle swabbing. The number of cells migrating to the
lower surface of the membrane was determined using crystal violet.
Five microscopic fields were randomly selected and viewed at ×200
magnification. Three repeats were performed.
Invasion assay
Cell invasion was analyzed using Boyden chambers
polyvinylpyrrolidone-free polycarbonate filters (8 µm) coated with
5 µg/ml Matrigel (BD Biosciences, Franklin Lakes, NJ, USA). The
cells were starved for 12 h and 2×106 cells were seeded
in the upper compartment of the Transwell unit with serum free
medium. The lower compartment was filled with medium containing 10%
FBS. The filters were stained with crystal violet solution. Crystal
violet stained invading cells were viewed at ×400 magnification.
For each filter, cells in 10 randomly chosen fields were counted
and expressed as the number of invading cells per high-power
field.
COX-2 inhibitor celecoxib functional
studies
Cell proliferation, migration and invasion assays
were also evaluated in the presence of celecoxib (Selleck, Houston,
TX, USA). Briefly, A549 and LTEP-A2 cells were incubated in 96-well
plates or 6-well plates with celecoxib (45 µg/ml) for 3 h. Dimethyl
sulfoxide (0.3 µl/ml) was used as a control.
Tumor growth in nude mice
The mice used in the present study were 4-week-o1d
male BALB/c nude mice (Vital River Laboratories Co., Ltd, Beijing,
China). The average weight of the mice was 17 g. Each group
included 6 mice. Nude mice were raised in a specific pathogen-free
level room and all feed was sterilized. The housing was maintained
at a temperature of 26–28°C. The relative humidity was maintained
at 40–60%. The daily light/dark cycle consisted of 10 h of light
and 14 h of dark. All animal experiments and maintenance were
approved by the Ethics Committee of the Beijing Chest Hospital,
Capital Medical University, Beijing, China, and conformed to the
guidelines of the Animal Care and Use Committee and the Chinese
Association of Laboratory Animal Care.
A549, A549-pU6 and A549-si10 cells were collected
and resuspended in 10% RMPI-1640 at a density of 1.5×107
cells/ml. In total, 3×106 cells were injected
subcutaneously into the flanks of 4-week-o1d male BALB/c nude mice.
The date at which a palpable tumor first arose was recorded. On the
60th day following tumor injection, the mice were euthanized using
a standard carbon dioxide method, and tumors were harvested and
weighed.
Immunohistochemistry
For immunohistochemistry, 4-µm-thick tumor sections
were immersed in 0.3% hydrogen peroxide for 10 min, microwaved in
citrate phosphate buffer (pH 6.0) and incubated with 10% normal
goat serum (Maixin Biotech. Co., Ltd., Fuzhou, China) for 30 min.
The tumor slices were incubated overnight at 4°C with the following
primary monoclonal antibodies (Maixin Biotech. Co., Ltd.): Rabbit
anti-COX-2 monoclonal antibodies (catalog no., RMA-0549); mouse
anti-MMP-2 monoclonal antibodies (catalog no., MAB-0244); and mouse
anti-VEGF monoclonal antibodies (catalog no., MAB-0243). Secondary
anti-mouse/rabbit antibodies from Kit-5030 (Maixin Biotech. Co.,
Ltd.) were then incubated with the samples at 37°C for 15 min.
Primary and secondary antibodies were working solution and were not
diluted. A standard staining procedure was finished by using a
3,3′-diaminobenzidine kit (Kit-0014; Maixin Biotech. Co., Ltd.).
Immunostaining was evaluated blindly by a board-certified
pathologist (Capital Medical University), who assigned the
intensity and prevalence score as described previously (25). Five microscopic fields were randomly
selected and viewed at a magnification of ×100. Briefly, the
intensity of staining was scored as follows: 0, negative; 1, weak
staining; 2, moderate staining; and 3, strong staining.
Statistical analysis
All statistical analyses were performed using SPSS
version 13.0 software (SPSS, Inc., Chicago, IL, USA). Quantitative
variables were compared using the one-way analysis of variance. The
χ2 test was used to assess qualitative variables.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Reduced expression of COX-2 in RNAi
transfected cell lines
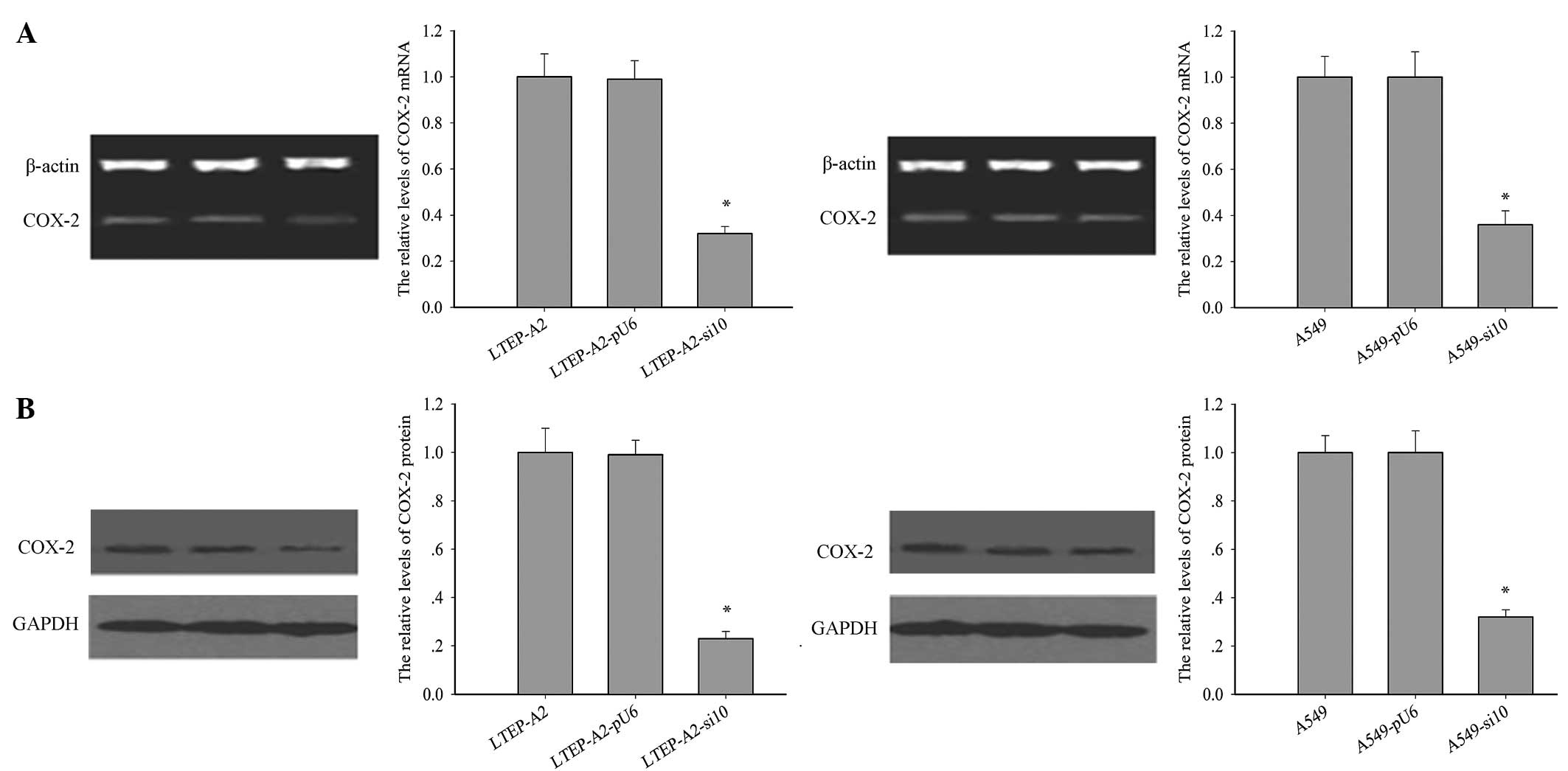
The levels of COX-2 protein and mRNA were examined
in the transfected lines (Fig. 1A and
B). Following RNAi transfection, the expression level of COX-2
was significantly reduced in LTEP-A2-si10 and A549-si10 cells.
Compared with the parental cells, the levels of COX-2 mRNA were
reduced by 60.0% (P=0.005) and 62.1% (P=0.004) and COX-2 protein by
68.2% (P=0.002) and 75.3% (P=0.001) in A549-si10 and LTEP-A2-si10
cells, respectively.
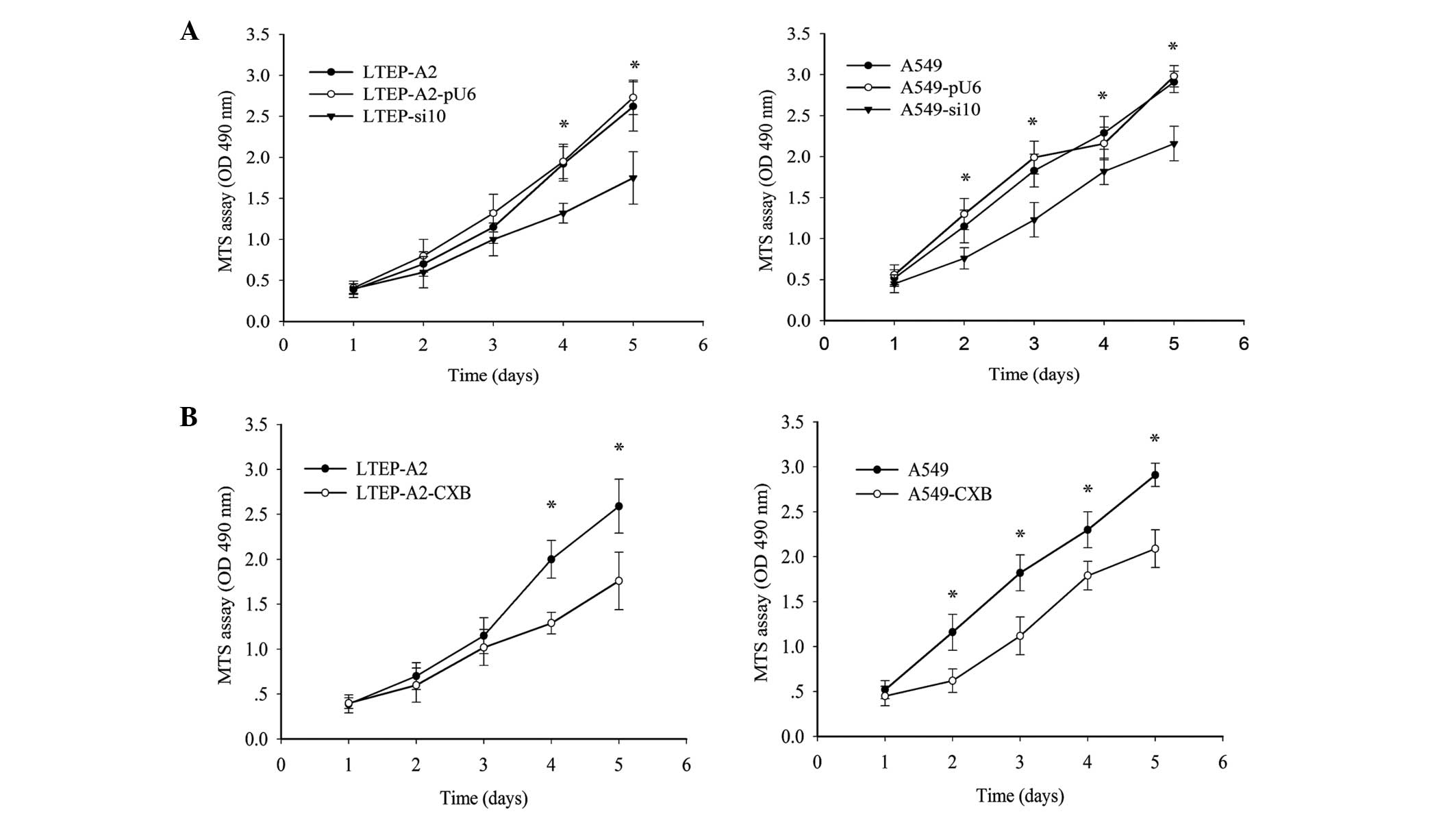
Altered proliferation rates of LTEP-A2
and A549 cells due to knockdown of COX-2 in vitro
To determine the result of silencing COX-2, cell
proliferation and colony formation assays were performed. As
indicated in Fig. 2A, the growth of
LTEP-A2-si10 cells was reduced on the third day. On the fourth and
fifth day, proliferation of LTEP-A2-si10 cells was considerably
reduced compared with parental cells (P=0.037). Similarly,
proliferation of A549-si10 cells was clearly reduced compared with
parental cells (P=0.041). To further confirm these results, a COX-2
inhibitor, celecoxib, was used to inhibit COX-2 expression and
revealed that COX-2 inhibition plays a role in cell proliferation.
COX-2 expression was clearly reduced by 89.5 (P=0.045) and 87.5%
(P=0.043) in A549 and LTEP-A2 cells, respectively, when 45 µg/ml
celecoxib was used (Fig. 2B).
Therefore, 45 µg/ml celecoxib was used in following experiments. An
MTS assay was performed, which demonstrated that the proliferation
ability of A549 and LTEP-A2 cells was similar to A549-si10 and
LTEP-A2-si10 cells following treatment with celecoxib (Fig. 2B).
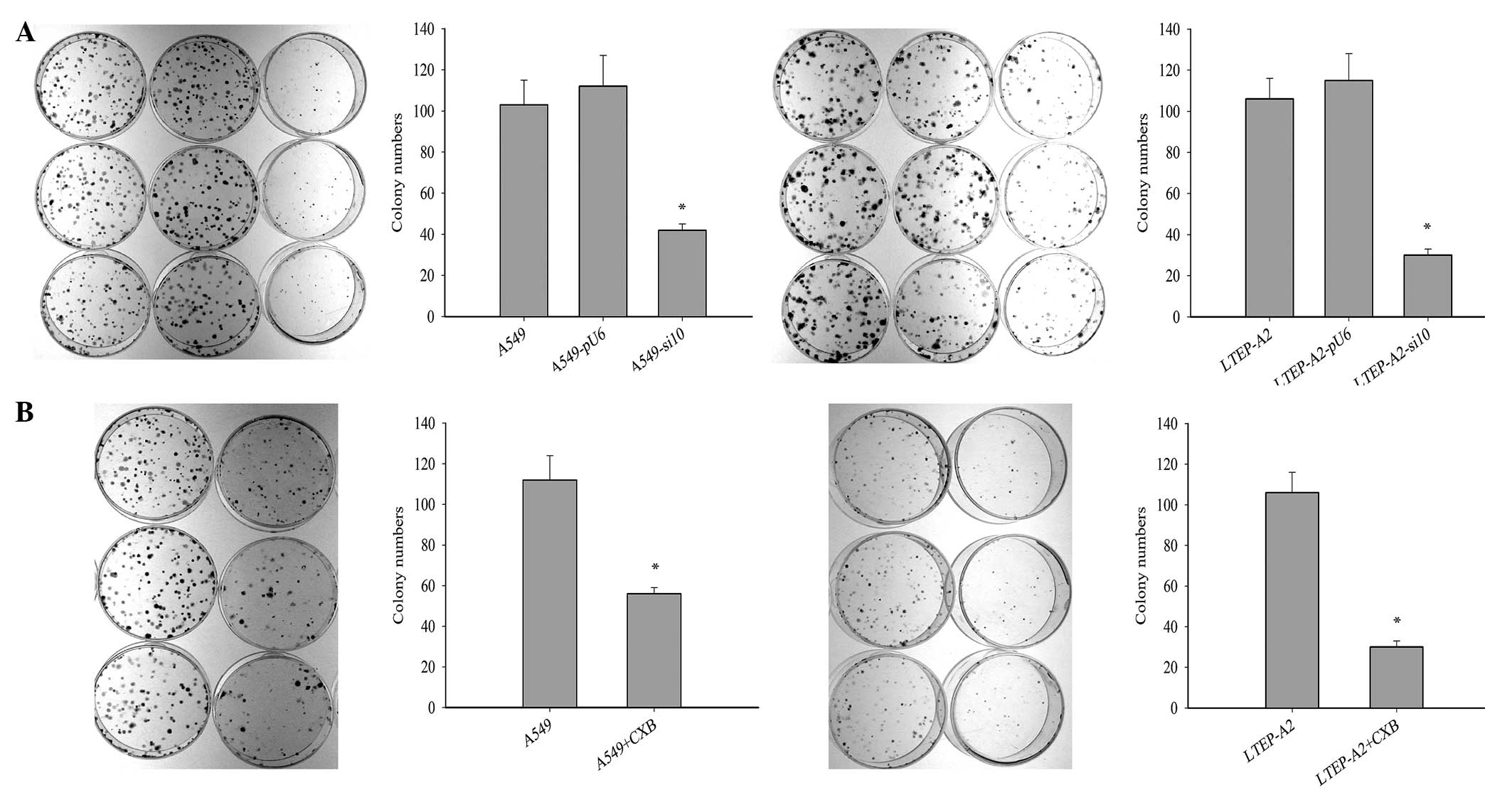
Cell tumorigenisis was inhibited
following suppression of COX-2
To investigate the effect of COX-2 on tumor growth,
cell tumorigenesis was evaluated by colony formation assay. The
colony formation rates of A549-si10 (P=0.021) and LTEP-A2-si10
(P=0.026) cells were significantly reduced compared with the
parental cells after 10 days (Fig.
3A). This suggests that COX-2 may overcome the
density-dependent inhibition of growth in tumor cells. When
celecoxib was used, the colony formation rates of parental A549
(P=0.019) and LTEP-A2 (P=0.043) cells were significantly reduced
compared with the control cells after 10 days (Fig. 3B).
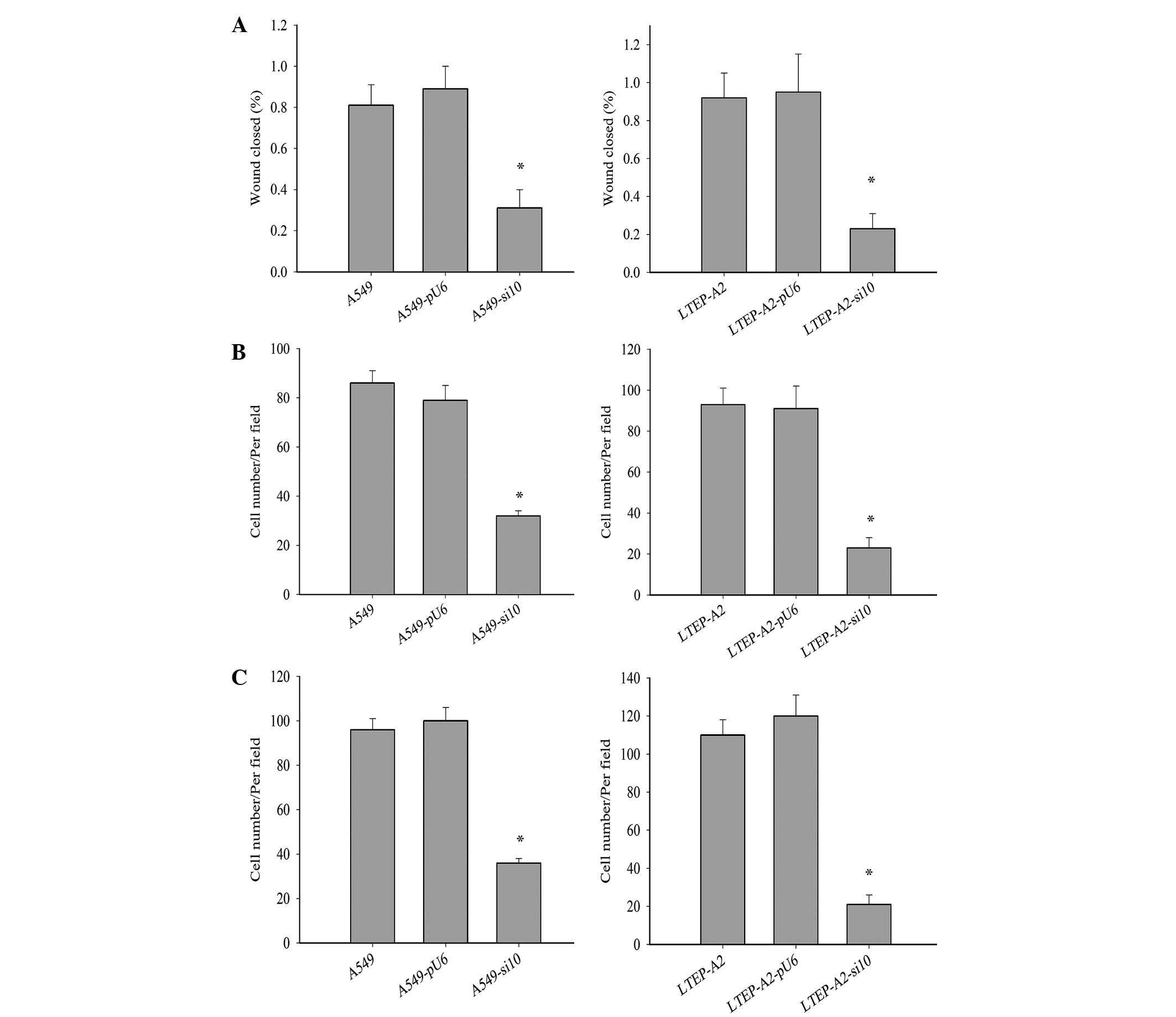
Cell metastasis was decreased by COX-2
suppression
Metastasis is a major characteristic of cancer
cells. To further study the effect of COX-2 on LTEP-A2 and A549
cell metastasis, the present study examined its effect on cell
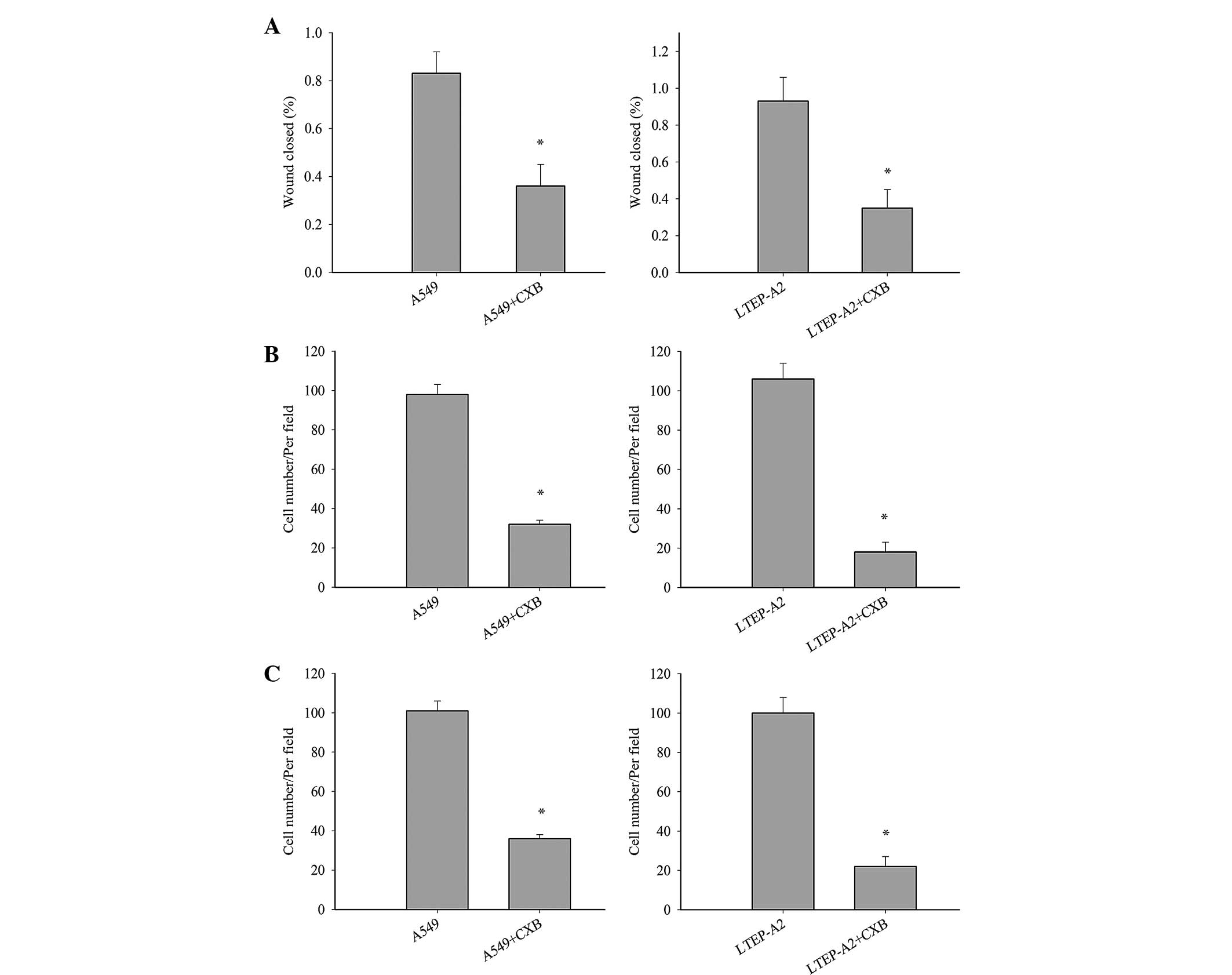
migration and invasion. Wound healing (Fig. 4A), invasion (Fig. 4B) and migration (Fig. 4C) were inhibited in A549-si10
(P=0.036, 0.033 and 0.032, respectively) and LTEP-A2-si10 (P=0.027,
0.019 and 0.028, respectively) cells compared with parental control
cells. When parental cells were treated with celecoxib, the cell
migration and invasion of A549 and LTEP-A2 cells were significantly
reduced compared with the control groups after 10 days (Fig. 5). These results reveal that COX-2 may
promote tumor cell metastasis.
Tumor growth in vivo was slowed by
COX-2 suppression
Tumor growth of mouse xenografts was substantially
slowed following COX-2 treatment compared with control groups
(Table I). Tumors developed three
weeks following inoculation: 6 tumors in A549 group; 4 tumors in
A549-pU6 group; 1 tumor in A549-si10 group. In the fourth week,
there were 6 tumors in A549 group, 6 in A549-pU6 group and 3 in the
A549-si10 group. By the 45th day, there were 6 tumors in A549
group, 6 in A549-pU6 group, and 5 in A549-si10 group.
 | Table I.Growth of mouse xenografts from human
lung adenocarcinoma A549 and LTEP-A2 cells transfected with pU6 and
si10. |
Table I.
Growth of mouse xenografts from human
lung adenocarcinoma A549 and LTEP-A2 cells transfected with pU6 and
si10.
| Groups |
|
| Tumor weights,
g |
|
| Average tumor
weight, g |
|---|
| A549 | 0.55 | 0.30 | 0.30 | 0.10 | 0.55 | 0.40 | 0.37±0.17 |
| A549-pU6 | 0.35 | 0.60 | 0.20 | 0.15 | 0.15 | 0.35 | 0.30±0.17 |
| A549-si10 | 0.20 | 0.30 | 0.05 | 0.05 | 0.20 | 0.00 |
0.13±0.11a |
VEGF, MMP-2 and EGFR expression was
downregulated following COX-2 suppression in vivo
The expression levels of COX-2 were primarily
evaluated in tumor tissues excised from the mice xenografts. The
findings were consistent with those in vitro (data not
shown). COX-2 expression in A549 and A549-pU6 xenografts was
moderately positive and scored 2, but this was clearly reduced in
A549-si10 xenografts, which were weakly positive and scored 1
(Fig. 6A). The expression of VEGF,
MMP-2 and EGFR in A549-si10 xenografts was clearly reduced compared
with A549 and A549-pU6 (Figs. 6B, C
and 7A). The expression of
hypoxia-inducible factor-α and MMP-9 in A549-si10 xenografts did
not clearly alter compared with A549 and A549-pU6 (Fig. 7B and C).
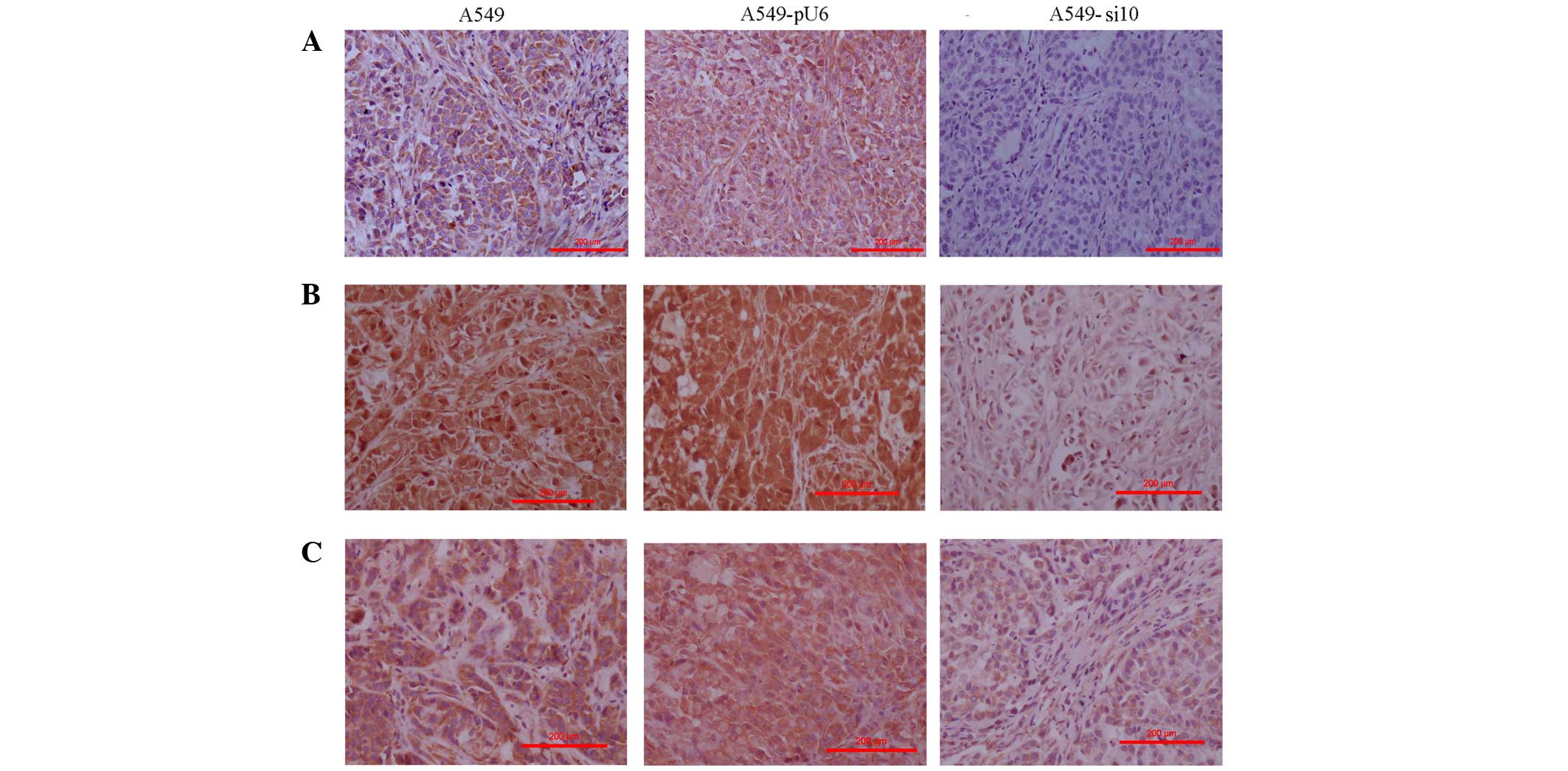 | Figure 6.Immunohistochemistry of mouse
xenografts from human lung adenocarcinoma A459 cells transfected
with si10 and pU6 (magnification, ×100). (A) Intensity of COX-2
expression staining was scored in the xenografts as follows: A549,
1; A549-pU6s, 1; A549-si10, 0. (B) Intensity of matrix
metalloproteinase-2 expression staining was scored in the
xenografts as follows: A549, 3; A549-pU6, 3; A549-si10, 1. (C)
Intensity of vascular endothelial growth factor expression staining
was scored in the xenografts as follows: A549, 2; A549-pU6, 2;
A549-si10, 1. Scale bar, 200 µm. COX-2, cyclooxygenase-2; si10,
small interfering RNA against COX-2. |
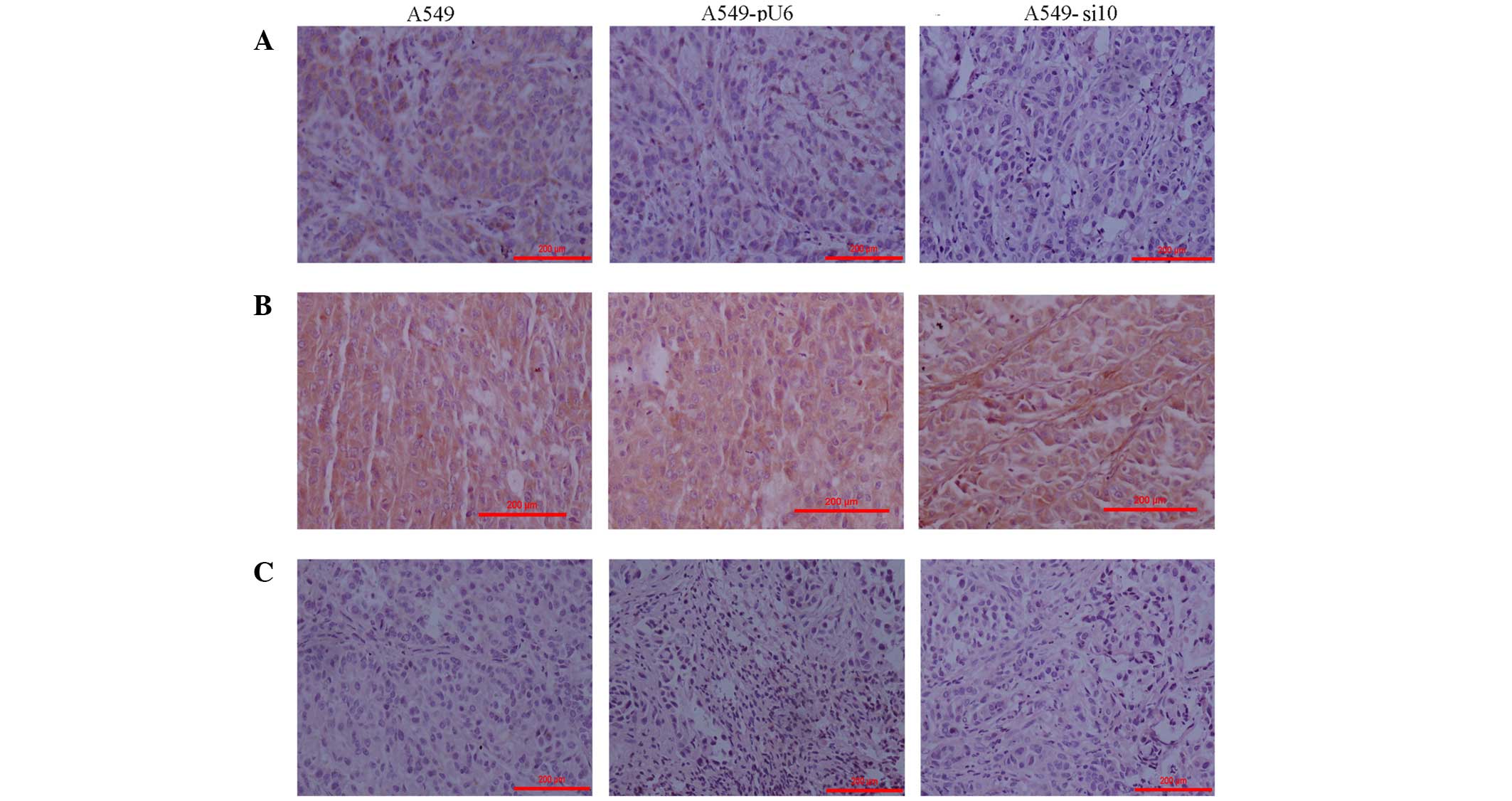 | Figure 7.Immunohistochemistry of mouse
xenografts from human lung adenocarcinoma A459 cells transfected
with si10 and pU6 (magnification, ×100). (A) Intensity of epidermal
growth factor receptor expression staining was scored in the
xenografts as follows: A459, 1; A459-pU6, 1; A459-si10, 0. (B)
Intensity of matrix metalloproteinase-9 expression staining was
scored in the xenografts as follows: A549, 2; A549-pU6, 2;
A549-si10, 2. (C) Intensity of hypoxia-inducible factor-α
expression staining was scored in the xenografts as follows: A549,
0; A549-pU6, 0; A549-si10, 0. Scale bar, 200 µm. si10, small
interfering RNA against cyclooxygenase-2. |
Discussion
Recently, numerous studies have demonstrated that
COX-2 is not only involved in tumorigenesis and tumor development,
but also plays a role in inflammation (26–28). In
tumor tissues, overexpression of COX-2 is a common phenomenon.
Previously published results of the associations between COX-2 and
clinicopathological factors are not consistent for lung cancer
(19,29–31). Our
previous results demonstrated that COX-2 was associated with
malignant pathology, tumor-node-metastasis stage, lymph node
metastasis, degree of differentiation and smoking (32). These results are also supported by the
literature (29–31).
Since COX-2 is associated with numerous
clinicopathological parameters, it may be extremely important in
the occurrence and development of lung cancer. To confirm this
hypothesis, the present study used RNAi technology to investigate
whether interfering with COX-2 expression affects lung cancer cell
proliferation and invasion. The present results revealed that COX-2
knockdown significantly slowed the proliferation and invasion of
LTEP-A2 and A549 lung cancer cells. In mouse xenografts, tumor
growth was slowed following silencing of COX-2, which further
confirmed our hypothesis. In addition, the present results
demonstrated that silencing COX-2 in lung cancer cells inhibited
cell proliferation and increased cell sensitivity to
density-dependent inhibition. This is consistent with previous
studies (33,34). Notably, COX-2 knockdown clearly
inhibited cell migration and invasion, suggesting that COX-2 has an
important effect on the metastasis of non-small cell lung cancer
(NSCLC) cells (35–37). The present results indicate that COX-2
plays a key role in the proliferation, motility and invasion of
lung cancer cells. In order to further confirm the role of COX-2 in
tumorigenesis and development, a COX-2 inhibitor, celecoxib, was
used by the present study in migration, invasion and proliferation
assays, which had similar results compared with COX-2
interference.
Subsequently, the present study investigated the
molecular mechanisms by which COX-2 is key in tumor malignancy. The
expression of COX-2 was primarily detected in mouse xenografts to
verify the changes following the silencing of COX-2. Since, COX-2
is associated with proliferation, migration and vascular lumen
formation of endothelial cells (23,28), VEGF,
MMP-2, MMP-9 and EGFR were investigated by the present study. The
expression of MMP-2, VEGF and EGFR was clearly reduced following
silencing of COX-2. MMP is a protein that degrades the
extracellular matrix (38,39), and previous studies have verified the
involvement of MMPs in NSCLC (40–42). In
addition, MMP-2 is associated with the lymphatic and vascular
invasion of NSCLC, and its expression may predict a poor prognosis
of early-stage patients with NSCLC (43). In addition, studies have reported that
COX-2 inhibitors inhibit the expression of MMP-2 and MMP-9 in
prostate cancer (44). In the present
study, silencing of COX-2 resulted in decreased levels of MMP-2;
however, there was no alteration in MMP-9 expression in
COX-2-silenced cells (A549). These results are consistent with a
previous study (45).
It is well known that VEGF is important in normal
and abnormal angiogenesis, since it is involved in almost every
step in the angiogenic process (46).
A previous study demonstrated that COX-2 inhibitors inhibit tumor
growth via an antiangiogenic mechanism (47). In addition, there is a close
association between COX-2 expression and tumor angiogenesis
(48). As a result, COX-2
overexpression may increase tumor blood supply and contribute to
tumor growth. The present data suggested that silencing COX-2
reduced VEGF expression in A549 cells. EGFR is a member of the
epidermal growth factor receptor family. Once activated, it leads
to cell division, proliferation, invasion and angiogenesis
(49–51). The present study observed that
silencing COX-2 lead to a reduction in EGFR expression in A549
cells.
VEGF, MMP-2 and EGFR are associated with tumor
angiogenesis (52–54). In the present study, the growth of
cancer cells was slowed and the migration and invasion ability
reduced following COX-2 silencing. These phenotypic changes may
have been caused by the alterations in the expression of VEGF,
MMP-2 and EGFR following COX-2 silencing. To the best of our
knowledge, this is the first study to report the direct association
between VEGF, MMP-2 and EGFR expression and COX-2 in lung cancer
cells.
In conclusion, the present study revealed that COX-2
is very important in lung tumor growth, infiltration and metastasis
via regulating VEGF, MMP-2 and EGFR expression. COX-2 may be a
potential target for lung cancer prevention and treatment.
Acknowledgements
This study was financed by the Beijing Natural
Science Foundation (Beijing, China; grant no. 7153162).
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tumino R, Capocaccia R, Traina A, Madeddu
A, Contrino ML and Zigon G: Estimates of cancer burden in Sicily.
Tumori. 99:399–407. 2013.PubMed/NCBI
|
|
3
|
Saver JL, Starkman S, Eckstein M, Stratton
SJ, Pratt FD, Hamilton S, Conwit R, Liebeskind DS, Sung G, Kramer
I, et al: FAST-MAG Investigators and Coordinators: Prehospital use
of magnesium sulfate as neuroprotection in acute stroke. N Engl J
Med. 372:528–536. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Balkwill F and Mantovani A: Inflammation
and cancer: Back to Virchow? Lancet. 357:539–545. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhang L, Conejo-Garcia JR, Katsaros D,
Gimotty PA, Massobrio M, Regnani G, Makrigiannakis A, Gray H,
Schlienger K, Liebman MN, et al: Intratumoral T cells, recurrence,
and survival in epithelial ovarian cancer. N Engl J Med.
348:203–213. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ding Y, Tong M, Liu S, Moscow JA and Tai
HH: NAD+-linked 15-hydroxyprostaglandin dehydrogenase (15-PGDH)
behaves as a tumor suppressor in lung cancer. Carcinogenesis.
26:65–72. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hussain SP and Harris CC: Inflammation and
cancer: An ancient link with novel potentials. Int J Cancer.
121:2373–2380. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Galon J, Costes A, Sanchez-Cabo F,
Kirilovsky A, Mlecnik B, Lagorce-Pagès C, Tosolini M, Camus M,
Berger A, Wind P, et al: Type, density, and location of immune
cells within human colorectal tumors predict clinical outcome.
Science. 313:1960–1964. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Whiteside TL: The tumor microenvironment
and its role in promoting tumor growth. Oncogene. 27:5904–5912.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Denkert C, Kobel M, Berger S, Siegert A,
Leclere A, Trefzer U and Hauptmann S: Expression of cyclooxygenase
2 in human malignant melanoma. Cancer Res. 61:303–308.
2001.PubMed/NCBI
|
|
11
|
Masferrer JL, Leahy KM, Koki AT, Zweifel
BS, Settle SL, Woerner BM, Edwards DA, Flickinger AG, Moore RJ and
Seibert K: Antiangiogenic and antitumor activities of
cyclooxygenase-2 inhibitors. Cancer Res. 60:1306–1311.
2000.PubMed/NCBI
|
|
12
|
Hold GL and El-Omar EM: Genetic aspects of
inflammation and cancer. Biochem J. 410:225–235. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kokawa A, Kondo H, Gotoda T, Ono H, Saito
D, Nakadaira S, Kosuge T and Yoshida S: Increased expression of
cyclooxygenase-2 in human pancreatic neoplasms and potential for
chemoprevention by cyclooxygenase inhibitors. Cancer. 91:333–338.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Khuri FR, Wu H, Lee JJ, Kemp BL, Lotan R,
Lippman SM, Feng L, Hong WK and Xu XC: Cyclooxygenase-2
overexpression is a marker of poor prognosis in stage I non-small
cell lung cancer. Clin Cancer Res. 7:861–867. 2001.PubMed/NCBI
|
|
15
|
Gupta S, Srivastava M, Ahmad N, Bostwick
DG and Mukhtar H: Over-expression of cyclooxygenase-2 in human
prostate adenocarcinoma. Prostate. 42:73–78. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chen YJ, Wang LS, Wang PH, Lai CR, Yen MS,
Ng HT and Yuan CC: High cyclooxygenase-2 expression in cervical
adenocarcinomas. Gynecol Oncol. 88:379–385. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sahin M, Sahin E and Gümüslü S:
Cyclooxygenase-2 in cancer and angiogenesis. Angiology. 60:242–253.
2009.PubMed/NCBI
|
|
18
|
Fidler MJ, Argiris A, Patel JD, Johnson
DH, Sandler A, Villaflor VM, Coon J IV, Buckingham L, Kaiser K,
Basu S and Bonomi P: The potential predictive value of
cyclooxygenase-2 expression and increased risk of gastrointestinal
hemorrhage in advanced non-small cell lung cancer patients treated
with erlotinib and celecoxib. Clin Cancer Res. 14:2088–2094. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Van Dyke AL, Cote ML, Prysak GM, Claeys
GB, Wenzlaff AS, Murphy VC, Lonardo F and Schwartz AG: COX-2/EGFR
expression and survival among women with adenocarcinoma of the
lung. Carcinogenesis. 29:1781–1787. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Banu N, Buda A, Chell S, Elder D, Moorghen
M, Paraskeva C, Qualtrough D and Pignatelli M: Inhibition of COX-2
with NS-398 decreases colon cancer cell motility through blocking
epidermal growth factor receptor transactivation: Possibilities for
combination therapy. Cell Prolif. 40:768–779. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Leahy KM, Ornberg RL, Wang Y, Zweifel BS,
Koki AT and Masferrer JL: Cyclooxygenase-2 inhibition by celecoxib
reduces proliferation and induces apoptosis in angiogenic
endothelial cells in vivo. Cancer Res. 62:625–631. 2002.PubMed/NCBI
|
|
22
|
Jiang MC, Liao CF and Lee PH: Aspirin
inhibits matrix metalloproteinase-2 activity, increases E-cadherin
production and inhibits in vitro invasion of tumor cells. Biochem
Biophys Res Commun. 282:671–677. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li W, Wang H, Lai B, Yang X and Zhang C:
The effects of interfering COX-2 gene expression on malignant
proliferation of human lung adenocarcinoma A2 cell in vitro.
Zhongguo Fei Ai Za Zhi. 12:100–105. 2009.(In Chinese). PubMed/NCBI
|
|
24
|
Jungi TW: Assay of chemotaxis by a
reversible Boyden chamber eliminating cell detachment. Int Arch
Allergy Appl Immunol. 48:341–352. 1975. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kurosumi M: Immunohistochemical assessment
of hormone receptor status using a new scoring system (J-Score) in
breast cancer. Breast Cancer. 14:189–193. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Harris RE, Casto BC and Harris ZM:
Cyclooxygenase-2 and the inflammogenesis of breast cancer. World J
Clin Oncol. 5:677–692. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Huang QC and Huang RY: The
cyclooxygenase-2/thromboxane A2 pathway: A bridge from rheumatoid
arthritis to lung cancer? Cancer Lett. 354:28–32. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Dubois RN: Role of inflammation and
inflammatory mediators in colorectal cancer. Trans Am Clin Climatol
Assoc. 125:358–372; discussion 372–373. 2014.PubMed/NCBI
|
|
29
|
Strazisar M, Mlakar V and Glavac D: The
expression of COX-2, hTERT, MDM2, LATS2 and S100A2 in different
types of non-small cell lung cancer (NSCLC). Cell Mol Biol Lett.
14:442–456. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Grimminger PP, Stöhlmacher J, Vallböhmer
D, Schneider PM, Hölscher AH, Metzger R, Danenberg PV and Brabender
J: Prognostic significance and clinicopathological associations of
COX-2 SNP in patients with nonsmall cell lung cancer. J Oncol.
1395902009.PubMed/NCBI
|
|
31
|
Zhu C, Liu J and Wang X: Detection of EGFR
and COX-2 expression by immunohistochemical method on a tissue
microarray section in lung cancer and biological significance.
Zhongguo Fei Ai Za Zhi. 13:107–111. 2010.(In Chinese). PubMed/NCBI
|
|
32
|
Li W, Yue W, Niu N, Zhang L, Zhao X, Ma L,
Yang X, Zhang C, Wang Y and Gu M: Expression and significance of
cyclooxygenase-2 in human lung cancer. Chinese-German J Clin Oncol.
13:203–206. 2014.
|
|
33
|
Li S, Gu Z, Xiao Z, Zhou T, Li J and Sun
K: Anti-tumor effect and mechanism of cyclooxygenase-2 inhibitor
through matrix metalloproteinase 14 pathway in PANC-1 cells. Int J
Clin Exp Pathol. 8:1737–1742. 2015.PubMed/NCBI
|
|
34
|
Atari-Hajipirloo S, Nikanfar S, Heydari A,
Noori F and Kheradmand F: The effect of celecoxib and its
combination with imatinib on human HT-29 colorectal cancer cells:
Involvement of COX-2, Caspase-3, VEGF and NF-κB genes expression.
Cell Mol Biol (Noisy-le-grand). 62:68–74. 2016.PubMed/NCBI
|
|
35
|
Shao Y, Li P, Zhu ST, Yue JP, Ji XJ, Ma D,
Wang L, Wang YJ, Zong Y, Wu YD and Zhang ST: MiR-26a and miR-144
inhibit proliferation and metastasis of esophageal squamous cell
cancer by inhibiting cyclooxygenase-2. Oncotarget. 7:15173–15186.
2016.PubMed/NCBI
|
|
36
|
Kim KM, Im AR, Kim SH, Hyun JW and Chae S:
Timosaponin AIII inhibits melanoma cell migration by suppressing
COX-2 and in vivo tumor metastasis. Cancer Sci. 107:181–188. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ho MY, Hung SW, Liang CM and Liang SM:
Recombinant viral capsid protein VP1 suppresses lung cancer
metastasis by inhibiting COX-2/PGE2 and MIG-7. Oncotarget.
5:3931–3943. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Miyata Y, Koga S, Kanda S, Nishikido M,
Hayashi T and Kanetake H: Expression of cyclooxygenase-2 in renal
cell carcinoma: Correlation with tumor cell proliferation,
apoptosis, angiogenesis, expression of matrix metalloproteinase-2,
and survival. Clin Cancer Res. 9:1741–1749. 2003.PubMed/NCBI
|
|
39
|
Sivula A, Talvensaari-Mattila A, Lundin J,
Joensuu H, Haglund C, Ristimäki A and Turpeenniemi-Hujanen T:
Association of cyclooxygenase-2 and matrix metalloproteinase-2
expression in human breast cancer. Breast Cancer Res Treat.
89:215–220. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Passlick B, Sienel W, Seen-Hibler R,
Wöckel W, Thetter O, Mutschler W and Pantel K: Overexpression of
matrix metalloproteinase 2 predicts unfavorable outcome in
early-stage non-small cell lung cancer. Clin Cancer Res.
6:3944–3948. 2000.PubMed/NCBI
|
|
41
|
Gridelli C, Maione P, Airoma G and Rossi
A: Selective cyclooxygenase-2 inhibitors and non-small cell lung
cancer. Curr Med Chem. 9:1851–1858. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Dohadwala M, Batra RK, Luo J, Lin Y,
Krysan K, Pold M, Sharma S and Dubinett SM: Autocrine/paracrine
prostaglandin E2 production by non-small cell lung cancer cells
regulates matrix metalloproteinase-2 and CD44 in
cyclooxygenase-2-dependent invasion. J Biol Chem. 277:50828–50833.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Marrogi AJ, Travis WD, Welsh JA, Khan MA,
Rahim H, Tazelaar H, Pairolero P, Trastek V, Jett J, Caporaso NE,
et al: Nitric oxide synthase, cyclooxygenase 2, and vascular
endothelial growth factor in the angiogenesis of non-small cell
lung carcinoma. Clin Cancer Res. 6:4739–4744. 2000.PubMed/NCBI
|
|
44
|
Liekens S, De Clercq E and Neyts J:
Angiogenesis: Regulators and clinical applications. Biochem
Pharmacol. 61:253–270. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Wu GS, Zou SQ, Liu ZR, Tang ZH and Wang
JH: Celecoxib inhibits proliferation and induces apoptosis via
prostaglandin E2 pathway in human cholangiocarcinoma cell lines.
World J Gastroenterol. 9:1302–1306. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Ghosh N, Chaki R, Mandal V and Mandal SC:
COX-2 as a target for cancer chemotherapy. Pharmacol Rep.
62:233–244. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Menter DG, Schilsky RL and DuBois RN:
Cyclooxygenase-2 and cancer treatment: Understanding the risk
should be worth the reward. Clin Cancer Res. 16:1384–1390. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Harris RE: Cyclooxygenase-2 (cox-2)
blockade in the chemoprevention of cancers of the colon, breast,
prostate, and lung. Inflammopharmacology. 17:55–67. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Hsu JY, Chang KY, Chen SH, Lee CT, Chang
ST, Cheng HC, Chang WC and Chen BK: Epidermal growth factor-induced
cyclooxygenase-2 enhances head and neck squamous cell carcinoma
metastasis through fibronectin up-regulation. Oncotarget.
6:1723–1739. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Asting AG, Farivar A, Iresjö BM, Svensson
H, Gustavsson B and Lundholm K: EGF receptor and COX-1/COX-2 enzyme
proteins as related to corresponding mRNAs in human per-operative
biopsies of colorectal cancer. BMC Cancer. 13:5112013. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Choi S, Lim TG, Hwang MK, Kim YA, Kim J,
Kang NJ, Jang TS, Park JS, Yeom MH and Lee KW: Rutin inhibits B [a]
PDE-induced cyclooxygenase-2 expression by targeting EGFR kinase
activity. Biochem Pharmacol. 86:1468–1475. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Chang CH, Huang YL, Shyu MK, Chen SU, Lin
CH, Ju TK, Lu J and Lee H: Sphingosine-1-phosphate induces VEGF-C
expression through a MMP-2/FGF-1/FGFR-1-dependent pathway in
endothelial cells in vitro. Acta Pharmacol Sin. 34:360–366. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Kim D, Dai J, Park YH, Fai L Yenwong, Wang
L, Pratheeshkumar P, Son YO, Kondo K, Xu M, Luo J, Shi X and Zhang
Z: Activation of EGFR/p38/HIF-1α is pivotal for angiogenesis and
tumorigenesis of malignantly transformed cells induced by
hexavalent chromium. J Biol Chem. May 25–2016.(Epub ahead of
print). View Article : Google Scholar
|
|
54
|
Lee HC, Su MY, Lo HC, Wu CC, Hu JR, Lo DM,
Chao TY, Tsai HJ and Dai MS: Cancer metastasis and EGFR signaling
is suppressed by amiodarone-induced versican V2. Oncotarget.
6:42976–42987. 2015.PubMed/NCBI
|