Introduction
Hepatocellular carcinoma (HCC) is a major malignancy
worldwide and its incidence is increasing annually; it is the
second most common cause of cancer-associated mortality (1). The majority of patients have a low
survival rate as a result of locally advanced or metastatic
diseases, and surgery is feasible for only a small percentage of
patients with HCC. Therefore, chemotherapy is the optimal
therapeutic strategy for inoperable HCC (2). Oxaliplatin has been widely used in
chemotherapy to reduce tumor recurrence and prolong survival in
patients with HCC because of its fewer side effects compared with
other platinum drugs (3). However,
chemoresistance to oxaliplatin in the form of suppressed HCC
apoptosis is commonly observed (4).
Interleukin-17 (IL-17) is predominantly secreted by
interleukin-17-producing T-helper (Th17) cells, which participate
in the progression and pathogenesis of inflammatory diseases
(3). The IL-17 receptor (IL-17R) is
expressed on the surface of numerous cells, including macrophages,
dendritic cells, epithelial cells, fibroblasts and T lymphocytes
(5,6).
Previous studies reported that IL-17-producing cells accumulated in
tumors (7,8), and that patients with malignant serum
effusions (9) or multiple myeloma
(10) showed significantly higher
serum levels of IL-17. Furthermore, patients with persistently
higher levels of IL-17 demonstrated the requirement for longer
courses of chemotherapy, since these patients comprised a
significant proportion of all cases of recurrence (11). Typically, IL-17 does not engage with
Toll/IL-1 receptor (TIR) domain-containing adaptors, such as MyD88,
TIR domain-containing adapter protein inducing interferon-β or IL-1
receptor-associated kinases (12).
Rather, IL-17 signals through nuclear factor (NF)-κB (13), mitogen-activated protein kinase (MAPK)
(14) and phosphoinositide 3-kinase
(PI3K) (14) signaling pathways. The
janus kinase 2 (JAK2)/signal transducer and activator of
transcription 3 (STAT3) signaling pathway plays an important role
in regulating a number of pathways associated with tumorigenesis,
including cell cycle progression, apoptosis and tumor cell evasion
of the immune system (15,16). In a previous study, phosphorylation of
STAT3 was markedly increased as early as 3 h following IL-17
treatment and lasted for 24 h (17),
which indicated that JAK2/STAT3 signaling may have important roles
in tumor progression associated with IL-17.
Autophagy involves lysosomal-mediated degradation of
cellular organelles and has been closely related to tumor
occurrence and progression (18). A
previous study reported that resistance to oxaliplatin in HepG2
cells could be recovered by inhibition of autophagy (19), which suggested opportunities for
exploitation of autophagy as a therapeutic target in cancer.
Although the association between IL-17 and tumour
chemotherapy has been previously investigated (20), the underlying mechanism remains
unclear. Therefore, the present study aimed to elucidate the role
of IL-17/IL-17R-induced autophagy in the resistance of HCC cells to
oxaliplatin, and to determine the potential underlying
mechanism.
Materials and methods
Patient samples and tissue
processing
A series of HCC specimens were obtained from 30
patients with pathologically confirmed HCC at the Affiliated
Hospital of North China University of Science and Technology
(Tangshan, China). No patients received adjuvant chemotherapy,
radiotherapy or surgery prior to admission. All patients were
administered one course (2 weeks) of oxaliplatin, after which the
concentration of IL-17 in sera and IL-17R mRNA levels were
detected. In addition, matched normal hepatic tissues were obtained
from 28 patients who were admitted to hospital due to wounds
obtained in a fall or traffic accident. Peripheral blood samples (3
ml) were collected from all patients. HCC biopsy specimens for the
detection of IL-17R mRNA expression levels by reverse
transcription-quantitative polymerase chain reaction (RT-qPCR) were
collected via paracentesis per cutem prior to and following
oxaliplatin treatment or surgical resection. Serum IL-17 levels
were determined using an ELISA. The present study was approved by
the Institutional Review Board of North China University of Science
and Technology. Informed consent was obtained from all patients
prior to specimen collection.
Cell lines and culture conditions
The human SMMC-7721, L02 and HepG2 cells lines were
maintained at 37°C in a humidified atmosphere containing 5%
CO2 in high-glucose Dulbecco's modified Eagle's medium
(Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
supplemented with 10% heat-inactivated fetal bovine serum (HyClone;
GE Healthcare Life Sciences, Logan, UT, USA), 100 units/ml
penicillin and 100 mg/ml streptomycin.
ELISA assays
Serum levels of IL-17 (pg/ml) were measured using a
solid phase sandwich ELISA assay according to the manufacturer's
protocols (R&D Systems, Inc., Minneapolis, MN, USA).
RNA isolation and RT-qPCR
To examine IL-17R expression in HCC patients prior
to and following oxaliplatin therapy, IL-17R mRNA expression levels
in the tumor tissues were compared with matched normal tissues by
RT-qPCR. Total RNA was extracted and reverse transcribed using an
RNeasy kit (Thermo Fisher Scientific, Inc., Pittsburgh, PA, USA),
according to the manufacturer's protocol. RNA (1 µg), along with
10X DNase I reaction buffer and 1 µg DNase I RNase-free was
transferred to a 1.5 ml tube where the volume was adjusted to 10 µl
using RNase-free water. After incubating for 30 min at 37°C, the
DNase I was inactivated by the addition of 1 µl 25 mM EDTA. The
mixture was subsequently heated for 10 min at 65°C. qPCR was
performed in 20 µl reaction volumes containing 2.0 µl cDNA, 0.4 µl
of each primer, 6.0 µl ddH2O, 0.4 µl ROX reference dye
and 10 µl fluorescent SYBR Green (Takara Bio, Inc., Otsu, Japan).
Amplification was performed in 96-well optical plates on a 7300
Real-Time PCR system (Applied Biosystems; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) with 30 sec incubation at 95°C, followed by
45 cycles of 95°C for 5 sec and 60°C for 60 sec. The primers used
were as follows: IL-17R forward, 5′-CACTCACTCTACGCAACCTTAA-3′,
reverse, 5′-AGATGCCCGTGATGAACC-3′; and GAPDH forward,
5′-GCACCGTCAAGGCTGAGAAC-3′ and reverse,
5′-ATGGTGGTGAAGACGCCAGT-3′.
Each sample was analyzed in
triplicate
The 2−ΔΔCq method of relative
quantification was performed to calculate relative changes in the
mRNA expression levels of target genes.
Cytokine inhibitor treatment
Cells were cultured in 6-well plates and treated
with 20 µg/ml Oxa in the presence or absence of an anti-IL-17R
antibody (10 µg/ml) or IL-17 (200 ng/ml) for 18 h. The
apoptosis-related proteins BCL-2 and BAX were measured by western
blotting. LY294002 (Beyotime Institute of Biotechnology, Haimen,
China), a PI3K-specific inhibitor, AG490 (Beyotime Institute of
Biotechnology), a JAK2 inhibitor, and 3-MA (Sigma-Aldrich; Merck
Millipore, Darmstadt, Germany), an inhibitor of autophagy, were
dissolved in dimethyl sulfoxide prior to use. These inhibitors were
added to the culture medium 1 h prior to oxaliplatin treatment,
with AG490 added at a dose of 15 µg/ml and LY294002 at 7.5 µg/ml.
Cells were treated with 20 µg/ml oxaliplatin for 18 h. No cell
cytotoxicity of these inhibitors, as assessed using a nuclear dye
exclusion assay (21), was observed
at the doses used in this study (data not shown).
Western blotting
Following treatment of the cells with oxaliplatin
(20 µg/ml), the cells were lysed in whole-cell lysate (Wuhan Boster
Biological Technology, Ltd., Wuhan, China) containing
phenylmethylsulfonyl fluoride and a phosphatase inhibitor. Equal
quantities of cell lysate (60 µg) were separated by 10% SDS-PAGE
and transferred onto polyvinylidene difluoride membranes. After
blocking in 5% evaporated milk for 1 h at 37°C, the membranes were
incubated with the following primary antibodies: Anti-IL-17R
(#D1Y4C), anti-B-cell lymphoma (BCL)-2 (#D55G8),
anti-BCL-2-associated X protein (BAX; #D2E11),
anti-microtubule-associated protein 1 light chain 3β (LC3B; #D11),
anti-JAK2 (#D2E12), anti-phosphorylated (p)-JAK2 (#D15E2),
anti-STAT3 (#D3Z2G), anti-p-STAT3 (#6E4) (all 1:1,000 dilution;
Cell Signaling Technology, Inc., Danvers, MA, USA) and
anti-Beclin-1 (dilution, 1:500; #B6061; Sigma-Aldrich; Merck
Millipore). GAPDH was used as a loading control and was detected
using an anti-GAPDH antibody (dilution, 1:5,000; #AP0066; Bioworld
Technology, Inc., St. Louis Park, MN, USA). The membrane was
incubated for 1 h at 37°C with goat anti-mouse immunoglobulin (Ig)G
(dilution, 1:10,000; #ab6785) and goat anti-rabbit IgG (dilution,
1:10,000; #ab6721) (Abcam, Cambridge, UK). After washing the
membrane for 45 min with cleaning solution, proteins were detected
using an enhanced chemiluminescence system and graphs were analyzed
using Image Lab software v2.5.1 (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA). Each experiment was repeated three times.
Statistical analysis
Statistical analyses were performed using SPSS 17.0
software (SPSS, Inc., Chicago, IL, USA). Graphs were analyzed using
the Image Lab system. Data are expressed as the mean ± standard
deviation of the values from three independent experiments.
Statistical analyses were conducted using either the Student's
t-test or one-way analysis of variance in comparison with
corresponding controls. P<0.05 was considered to indicate a
statistically significant difference.
Results
Expression of IL-17 and IL-17R in HCC
patients
IL-17 has a role in numerous autoimmune and
inflammatory conditions, including rheumatoid arthritis, multiple
sclerosis, psoriasis, Crohn's disease and systemic lupus
erythematosus, through combining with IL-17R (22). However, evidence has shown that IL-17
may also contribute to disease progression and treatment response
in patients with tumors (23). In the
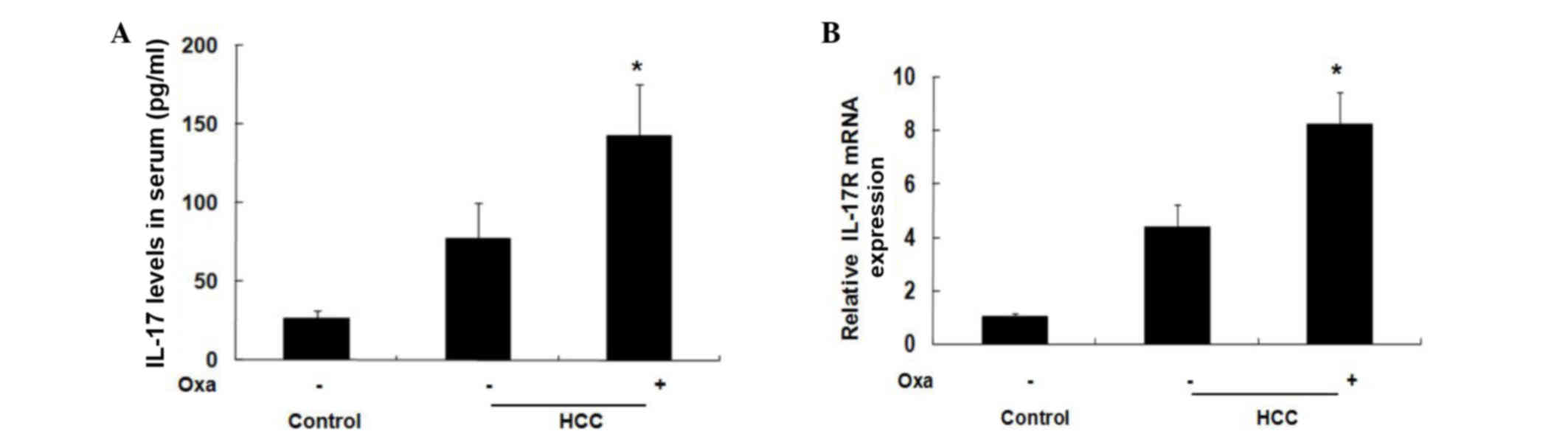
present study, the levels of IL-17 were significantly increased in
oxaliplatin-treated HCC patients, as compared with untreated HCC
patients. Prior to treatment, the serum levels of IL-17 were
77.36±22.90 pg/ml, but were significantly increased up to
142.41±33.25 pg/ml after one course of treatment (2 weeks)
(P<0.05; Fig. 1A). Furthermore,
the mRNA expression levels of IL-17R in oxaliplatin-treated HCC
biopsy specimens were significantly higher compared with untreated
HCC biopsy specimens (P<0.05; Fig.
1B). These results suggest that oxaliplatin increases the
expression of IL-17/IL-17R in patients with HCC, and that there is
an association between oxaliplatin-induced apoptosis and
IL-17/IL-17R.
Oxaliplatin induces the expression of
IL-17R in HCC cells
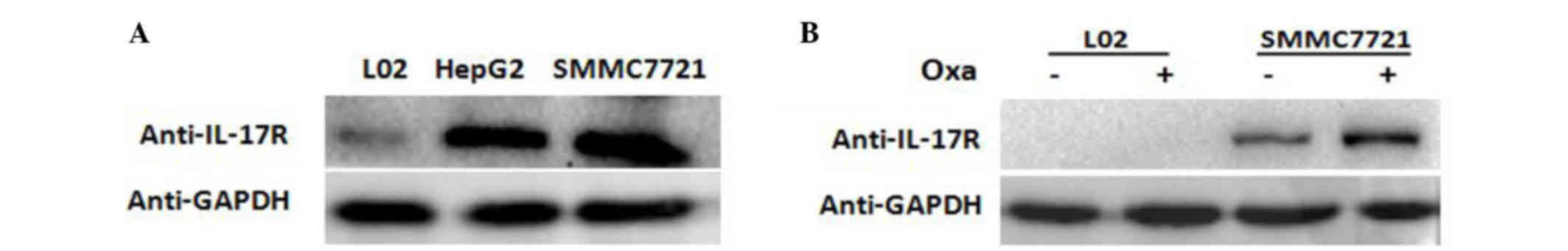
L02, HepG2 and SMMC-7721 cell lines were cultured in
the presence of oxaliplatin and western blot analysis was performed
to measure the amount of IL-17R protein in each cell line. The
results showed that IL-17R was expressed in HepG2 and SMMC-7721
cells, but not in L02 cells (Fig.
2A).
In order to confirm the changes in IL-17R expression
following oxaliplatin treatment of SMMC-7721 cells, oxaliplatin (20
µg/ml) was added to SMMC-7721 and L02 cells for 18 h. Western
blotting showed that the expression of IL-17R increased markedly in
oxaliplatin-treated SMMC-7721 cells compared with the untreated
cells (Fig. 2B). These results
suggest that IL-17 is expressed in HCC cells and is increased
following oxaliplatin treatment.
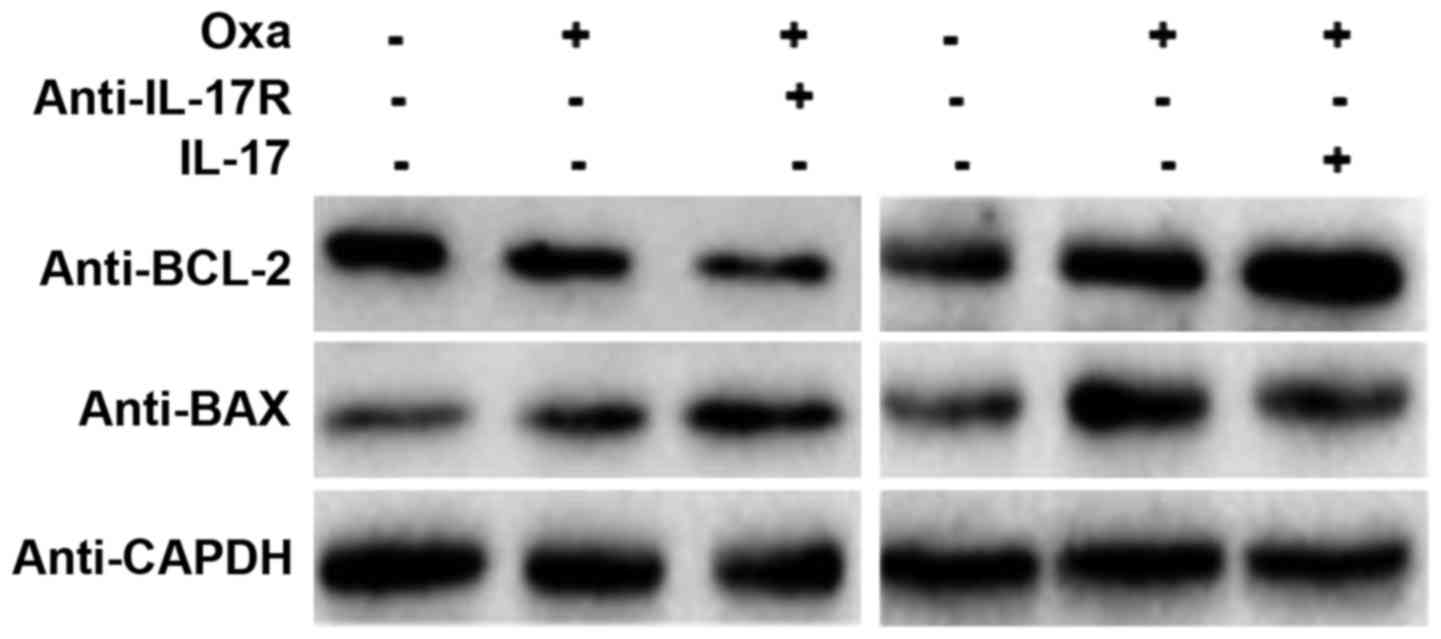
IL-17/IL-17R inhibits
oxaliplatin-induced apoptosis of SMMC-7721 cells
As shown in Fig. 2,
IL-17R was expressed in SMMC-7721 cells. Furthermore, expression of
IL-17R in SMMC-7721 cells increased markedly following oxaliplatin
treatment. However, the role of IL-17R oxaliplatin-induced in these
cells is unknown. As a mechanism for programmed cell death,
apoptosis is regulated by the BCL-2 family proteins, including BAX,
which control the sensitivity of cells to apoptotic stresses. BCL-2
is an anti-apoptosis gene, whereas BAX is an apoptosis-promoting
matrix gene (24). In the present
study, IL-17R-blocking and IL-17-promoting assays were designed to
observe the effect of IL-17/IL-17R on oxaliplatin-induced
apoptosis. Western blotting showed that BCL-2 protein expression
was decreased and BAX protein expression was increased in
oxaliplatin-treated SMMC-7721 cells, as compared with the untreated
cells, which indicated that apoptosis was induced by oxaliplatin
treatment (Fig. 3). When the IL-17R
on the surface of SMMC-7721 cells was blocked by a neutralizing
anti-IL-17R antibody (10 µg/ml; #MAB177; R&D Systems, Inc.),
the expression of BCL-2 was decreased and that of BAX was increased
(Fig. 3). Conversely, the expression
of BCL-2 was increased and that of BAX was decreased in
IL-17-promoting SMMC7721 cells. These results suggest that
IL-17/IL-17R may inhibit HCC apoptosis and that blocking IL-17R is
able to upregulate the susceptibility of HCC cells to
oxaliplatin-induced apoptosis.
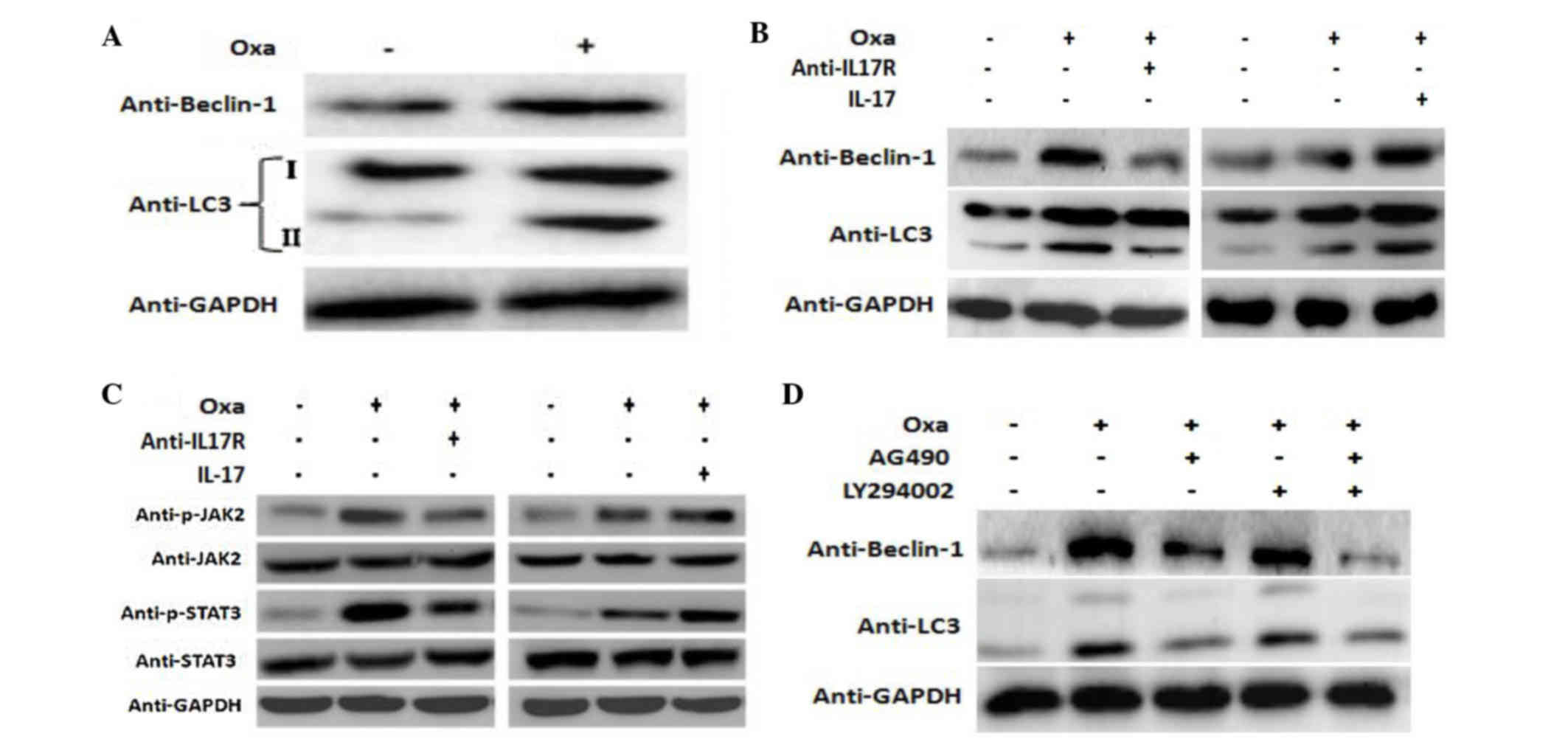
IL-17/IL-17R induces autophagy in
SMMC-7721 cells
Increasingly, studies have focused on the effect of
autophagy on tumor progression (25,26).
Autophagy is a reversible process that regulates tumor survival or
death; thus, it is closely associated with tumor progression
(27). Furthermore, autophagy during
chemotherapy has been shown to induce chemoresistance (28), while sensitivity to chemotherapy was
increased when autophagy was inhibited (29,30).
It is well known that decreased susceptibility to
apoptosis during chemotherapy is the main mechanism of
chemoresistance (31,32). LC3 is a specific marker of
autophagosomes in mammalian cells, and the conversion of the
soluble form of LC3 (LC3-I) to the autophagosome-associated form
(LC3-II) is a characteristic of autophagy (33–35).
Furthermore, Beclin-1 is another important autophagy gene. In the
present study, Beclin-1 and LC3-II expression in SMMC-7721 cells
was increased following treatment with oxaliplatin (Fig. 4A). In addition, the expression of
Beclin-1 and LC3-II was markedly attenuated following blocking of
IL-17R with a neutralizing antibody, and was increased accordingly
following exposure to IL-17 (Fig.
4B).
IL-17/IL-17R induces autophagy in
SMMC-7721 cells through the JAK2/STAT3 signaling pathway
IL-17 mediates cellular activities through various
signal transduction pathways, including NF-κB, MAPK and PI3K
(14). The present study investigated
the potential role of the JAK2/STAT3 signaling pathway in
IL-17-mediated autophagy. In the present study, the levels of
p-JAK2 and p-STAT3 were increased in oxaliplatin-stimulated cells
compared with the control cells, while they were decreased
following blocking of IL-17R. Conversely, treatment with IL-17
(Sigma-Aldrich; Merck Millipore) increased the levels of p-JAK2 and
p-STAT3 in SMMC-7721 cells (Fig. 4C).
These results suggest that IL-17/IL-17R mediate cellular responses
through the JAK2/STAT3 signaling pathway.
In order to investigate the relationship between
IL17/IL17R and JAK2/STAT3 and oxaliplatin-induced autophagy,
blocking experiments were performed. AG490, which is a JAK
inhibitor, is able to specifically block the activation of the
JAK2/STAT3 signaling pathway, as tyrosine phosphorylation of STAT3
is dependent on JAK activity (36).
Furthermore, since the PI3K pathway has a crucial role in autophagy
(37–39), the PI3K signaling pathway was used as
a control pathway to investigate the JAK2/STAT3 pathway. LY294002,
which is reported to inhibit AKT activation in a dose-dependent
manner (40), was used and compared
with the effect of AG490. SMMC-7721 cells were incubated with AG490
(15 µg/ml) or LY294002 (10 µg/ml) for 1 h prior to oxaliplatin
treatment, after which autophagy-related proteins were detected by
western blotting. Notably, a marked decrease in the expression of
both Beclin-1 and LC3 II proteins was observed in AG490-treated
cells, while only a negligible change was observed in
LY294002-treated cells (Fig. 4D).
These results suggest that inhibition of the JAK2/STAT3 signaling
pathway suppresses oxaliplatin-induced autophagy to a greater
extent than inhibition of the PI3K pathway, which further indicates
that the JAK2/STAT3 signaling pathway may have an important role in
oxaliplatin-induced autophagy.
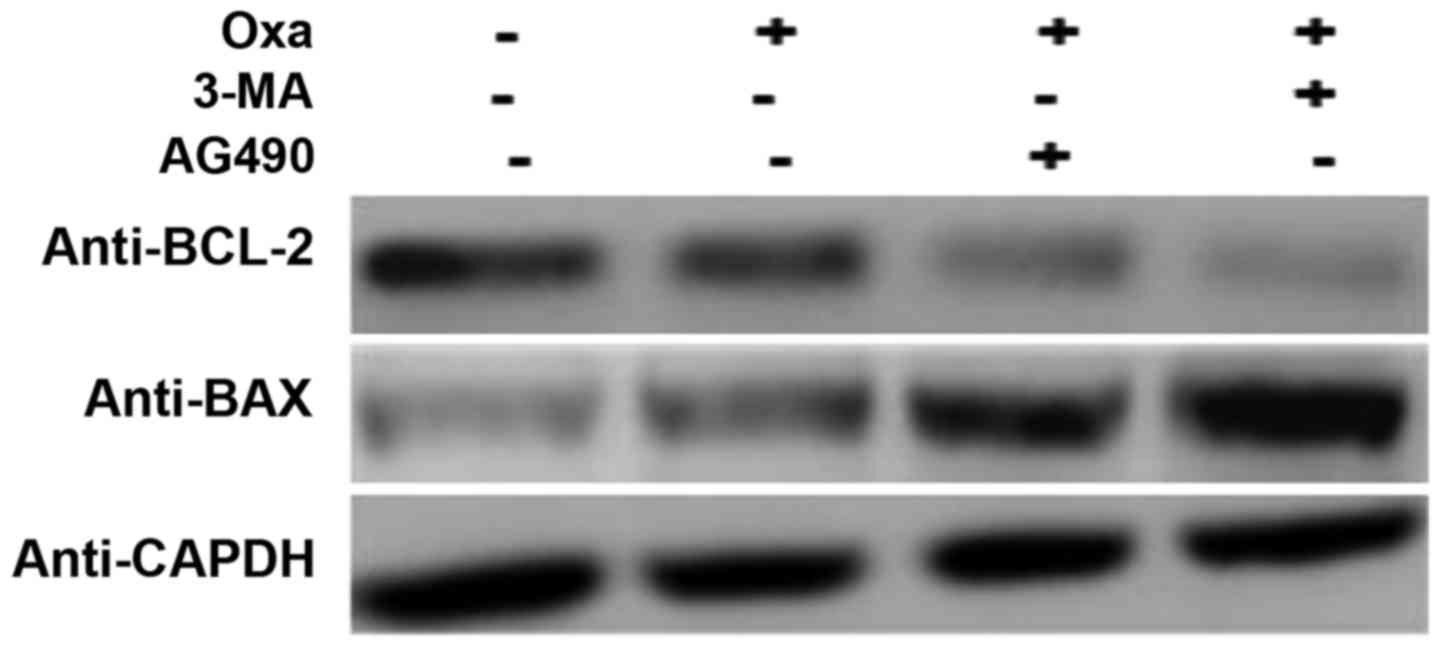
In order to further confirm the role of IL-17/IL-17R
and the JAK2/STAT3 signaling pathway in the induction of autophagy
and inhibition of apoptosis, SMMC-7721 cells were treated with
3-methyladenine (3-MA; 10 mM; Sigma-Aldrich; Merck Millipore),
which is a known inhibitor of autophagy (41), and AG490 to inhibit the JAK2/STAT3
signaling pathway. Compared with the cells treated with oxaliplatin
only, the AG490- and oxaliplatin-treated cells expressed less BCL-2
and more BAX, which indicated that apoptosis was induced to a
greater extent in these cells. There was no difference in the
expression of BCL-2 and BAX following treatment with AG490 and 3-MA
(Fig. 5). These results suggest that
autophagy in oxaliplatin-treated cells is induced through the
JAK2/STAT3 signaling pathway.
Together, the results of the present study suggest
that autophagy is activated in HCC cells following oxaliplatin
treatment via IL17/IL17R-JAK2/STAT3, and that this may be involved
in the chemoresistance of HCC to oxaliplatin.
Discussion
IL-17, which is the hallmark cytokine of the
newly-defined Th17 cell subset, serves an important role in
inflammatory diseases (42). Since
chronic inflammation has been associated with tumor invasion,
migration and metastasis (43), the
significance of IL-17 in tumor progression has received increasing
attention. The receptor of IL-17, IL-17R, is a common signaling
subunit used by multiple ligands (44). Researches have begun to investigate
the unusual functional motifs and novel proximal signaling
mediators employed by the IL-17R family to mediate downstream
events (7,45). There is evidence that IL-17 may emerge
as a novel prognostic marker in HCC.
In the present study, it was demonstrated that
autophagy downregulated oxaliplatin-induced HCC apoptosis through
the IL-17/IL-17R-JAK2/STAT3 signaling pathway. IL-17 and IL-17R
were markedly increased in oxaliplatin-treated HCC patients.
Previous studies reported that IL-17 may exert pro-tumor or
antitumor effects in various tumor contexts (46–48). The
explanation for this discrepancy remains unknown. The present study
aimed to determine the role that IL-17/IL-17R plays in the response
of HCC to chemotherapy. Western blot analysis demonstrated that the
expression of IL-17R was upregulated in HepG2 and SMMC-7721 cell
lines, which was consistent with the report that IL-17-producing
cells accumulate in various cancers, including HCC (8). In addition, IL-17/IL-17R expression was
shown to increase in oxaliplatin-treated SMMC-7721 cells, which was
associated with downregulation of apoptosis, as demonstrated by the
detection of BCL-2 and BAX expression by western blotting.
Previous studies have focused on the relationship
between autophagy and tumors (49,50). The
role of autophagy in tumors is complex, as it has been associated
with both tumor suppression and therapeutic resistance in advanced
tumors (51). Inhibition of autophagy
acted synergistically with chemotherapy in a mouse model of
lymphoma (52). Autophagy was
considered a temporary survival mechanism through the interaction
of autophagy-related Beclin-1 and anti-apoptotic BCL-2 (53); thus, autophagy may be induced upon
chemotherapy as a survival mechanism. In the present study, the
reduced sensitivity of HCC cells to apoptosis may have been related
to the induction of autophagy. Therefore, the expression of
autophagy-related proteins, including LC3B and Beclin-1, was
detected in oxaliplatin-treated SMMC-7721 cells. It was determined
that oxaliplatin was able to induce autophagy in HCC and that
autophagy was dependent on IL-17/IL-17R. To delineate the potential
mechanism underlying IL-17/IL-17R-induced autophagy, the activation
status of JAK2 and STAT3 under the interference of IL-17/IL-17R was
observed. It was found that IL-17/IL-17R increased the
phosphorylation of JAK2 and STAT3, which was consistent with the
fact that STAT3 controls Th17 cell differentiation (54). Previous studies have shown that the
PI3K signaling pathway has a crucial role in autophagy (55–57).
Therefore, to confirm the roles of the PI3K and/or the JAK2/STAT3
signaling pathway in autophagy activation in oxaliplatin-treated
HCC cells, SMMC-7721 cells were pre-treated with both LY294002 and
AG490 to block PI3K and JAK2/STAT3. According to the western blot
result, autophagy was inhibited to a greater extent in
AG490-treated cells, which suggested that the JAK2/STAT3 signaling
pathway was predominantly involved. 3-MA is a specific autophagy
inhibitor, and pre-treatment of SMMC-7721 cells with 3-MA induced
apoptosis to a greater extent than treatment with oxaliplatin alone
by increasing BAX and decreasing BCL-2 expression. Similarly, cells
treated with AG490 expressed less BCL-2 and more BAX, thus
confirming that autophagy was induced by the JAK2/STAT3 signaling
pathway.
Consistent with the results of the present study,
Huang et al (58) reported
that a JAK inhibitor was able to effectively suppress
IL-17A-induced gene expression in human bronchial epithelial cells.
Sun et al (59) suggested that
STAT3 signaling was a key pathway that mediates immune suppression
in the tumor microenvironment, and that aberrantly activated STAT3
in HCC cells resulted in the upregulation of cytokines, including
IL-17. Cross-Knorr et al (60)
showed that oxaliplatin was able to enhance the apoptosis of cancer
cells by disrupting survival signaling via the JAK/STAT pathway at
the receptor level in stage II colon cancer patients. Together,
these findings suggested the importance of the JAK2/STAT3 signaling
in the regulation of IL-17 signaling.
Previous reports have also indicated that
pharmacological inhibitors of JAKs are able to limit IL-17
signaling (61), although these
results should be interpreted with caution because of the
non-specific effects of such compounds (62). Furthermore, IL-17-induced activation
of STAT factors, which could promote cytokines secretion (including
IL-6), have not been satisfactorily disproved (7). Therefore, whether IL-17/IL-17R mediates
oxaliplatin-induced autophagy directly or through other pathways
requires further analysis.
In conclusion, the present study demonstrated that
autophagy inhibited oxaliplatin-induced HCC apoptosis via the
IL-17/IL-17R-JAK2/STAT3 signaling pathway. These results suggested
that blocking IL-17/IL-17R may be considered a novel therapy for
chemoresistant HCCs.
Acknowledgements
This study was supported by the Natural Science
Foundation of Hebei Province (grant no. 2016209007).
References
|
1
|
Zhao Y, Wang Q, Deng X, Shi P and Wang Z:
Quantitative assessment of the association between GSTP1 gene
Ile105Val polymorphism and susceptibility to hepatocellular
carcinoma. Tumor Biol. 34:2121–2126. 2013. View Article : Google Scholar
|
|
2
|
Louaf S, Boige V, Ducreux M, Bonyhay L,
Mansourbakht T, de Baere T, Asnacios A, Hannoun L, Poynard T and
Taïeb J: Gemcitabine plus oxaliplatin (GEMOX) in patients with
advanced hepatocellular carcinoma (HCC): Results of a phase II
study. Cancer. 109:1384–1390. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sun HQ, Zhang JY, Zhang H, Zou ZS, Wang FS
and Jia JH: Increased Th17 cells contribute to disease progression
in patients with HBV-associated liver cirrhosis. J Viral Hepat.
19:396–403. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Du H, Yang W, Chen L, Shi M, Seewoo V,
Wang J, Lin A, Liu Z and Qiu W: Role of autophagy in resistance to
oxaliplatin in hepatocellular carcinoma cells. Oncol Rep.
27:143–150. 2012.PubMed/NCBI
|
|
5
|
Shen F and Gaffen SL: Structure-function
relationships in the IL-17 receptor: Implications for signal
transduction and therapy. Cytokine. 41:92–104. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gaffen SL: Recent advances in the IL-17
cytokine family. Curr Opin Immunol. 23:613–619. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ma Y, Aymeric L, Locher C, Mattarollo SR,
Delahaye NF, Pereira P, Boucontet L, Apetoh L, Ghiringhelli F,
Casares N, et al: Contribution of IL-17-producing gamma delta T
cells to the ecacy of anticancer chemotherapy. J Exp Med.
208:491–503. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lv L, Pan K, Li XD, She KL, Zhao JJ, Wang
W, Chen JG, Chen YB, Yun JP and Xia JC: The accumulation and
prognosis value of tumor infiltrating IL-17 producing cells in
esophageal squamous cell carcinoma. PLoS One. 6:e182192011.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li Q, Han Y, Fei G, Guo Z, Ren T and Liu
Z: IL-17 promoted metastasis of non-small-cell lung cancer cells.
Immunol Lett. 148:144–150. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lemancewicz D, Bolkun L, Jablonska E,
Czeczuga-Semeniuk E, Kostur A, Kloczko J and Dzieciol J: The role
of interleukin-17A and interleukin-17E in multiple myeloma
patients. Med Sci Monit. 18:BR54–BR59. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Droeser RA, Güth U, Eppenberger-Castori S,
Stadlmann S, Hirt C, Terracciano L and Singer G: High
IL-17-positive tumor immune cell infiltretion is indicative for
chemosensitivity of ovarian carcinoma. J Cancer Res Clin Oncol.
139:1295–1302. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Maitra A, Shen F, Hanel W, Mossman K,
Tocker J, Swart D and Gaffen SL: Distinct functional motifs within
the IL-17 receptor regulate signal transduction and target gene
expression. Proc Nat Acad Sci USA. 104:7506–7511. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Korn T: Th17 cells: Effector T cells with
inflammatory properties. Semin Immunol. 19:362–371. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gaffen SL: Structure and signaling in the
IL-17 receptor family. Nat Rev Immunol. 9:556–567. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Okinaga T, Ariyoshi W and Akifusa S:
Essential role of JAK/STAT pathway in the induction of cell cycle
arrest in macrophages infected with periodontopathic bacterium
Aggregatibacter actinomycetemcomitans. Med Microbiol Immunol.
202:167–74. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chen Y, Jiao B, Yao M, Shi X, Zheng Z, Li
S and Chen L: ISG12a inhibits HCV replication and potentiates the
anti-HCV activity of IFN-α through activation of the Jak/STAT
signaling pathway independent of autophagy and apoptosis. Virus
Res. 10:S0168–S01702. 2016.
|
|
17
|
Gu FM, Li QL, Gao Q, Jiang JH, Zhu K,
Huang XY, Pan JF, Yan J, Hu JH, Wang Z, et al: IL-17 induces
AKT-dependent IL-6/JAK2/STAT3 activation and tumor progression in
hepatocellular carcinoma. Mol Cancer. 10:1502011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xu Y, An Y, Wang Y, Zhang C, Zhang H,
Huang C, Jiang H, Wang X and Li X: miR-101 inhibits autophagy and
enhances cisplatin-induced apoptosis in hepatocellular carcinoma
cells. Oncol Rep. 29:2019–2024. 2013.PubMed/NCBI
|
|
19
|
Harris SJ, Ciuclan L, Finan PM, Wymann MP,
Walker C, Westwick J, Ward SG and Thomas MJ: Genetic ablation of
PI3Kγ results in defective IL-17RA signalling in T lymphocytes and
increased IL-17 levels. Eur J Immunol. 42:3394–3404. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Murugaiyan G and Saha B: Protumor vs
antitumor functions of IL-17. J Immunol. 183:4169–4175. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Fortmüller K, Alt K, Gierschner D, Wolf P,
Baum V, Freudenberg N, Wetterauer U, Elsässer-Beile U and Bühler P:
Effective targeting of prostate cancer by lymphocytes redirected by
a PSMAxCD3 bispecific single-chain diabody. Prostate. 71:588–596.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tesmer LA, Lundy SK, Sarkar S and Fox DA:
Th17 cells in human disease. Immunol Rev. 223:87–113. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Martin F, Apetoh L and Ghiringhelli F:
Controversies on the role of Th17 in cancer: A TGF-β-dependent
immunosuppressive activity? Trends Mol Med. 18:742–749. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Anilkumar U and Prehn JHM: Anti-apoptotic
BCL-2 family proteins in acute neural injury. Front Cell Neurosci.
30:2812014.
|
|
25
|
Xu Y and Lv SX: The effect of JAK2
knockout on inhibition of liver tumor growth by inducing apoptosis,
autophagy and anti-proliferation via STATs and PI3K/AKT signaling
pathways. Biomed Pharmacother. 84:1202–1212. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Tan YQ, Zhang J and Zhou G: Autophagy and
its implication in human oral diseases. Autophagy. 20:02016.
|
|
27
|
Cui J, Gong Z and Shen HM: The role of
autophagy in liver cancer: Molecular mechanisms and potential
therapeutic targets. Biochim Biophys Acta. 1836:15–26.
2013.PubMed/NCBI
|
|
28
|
Liu D, Yang Y, Liu Q and Wang J:
Inhibition of autophagy by 3-MA potentiates cisplatin-induced
apoptosis in esophageal squamous cell carcinoma cells. Med Oncol.
28:105–111. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Golden EB, Cho HY, Jahanian A, Hofman FM,
Louie SG, Schönthal AH and Chen TC: Chloroquine enhances
temozolomide cytotoxicity in malignant gliomas by blocking
autophagy. Neurosurg Focus. 37:E122014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wu W, Li W, Zhou Y and Zhang C: Inhibition
of beclin1 affects the chemotherapeutic sensitivity of
osteosarcoma. Int J Clin Exp Pathol. 7:7114–7122. 2014.PubMed/NCBI
|
|
31
|
Moore N, Houghton J and Lyle S:
Slow-cycling therapy-resistant cancer cells. Stem Cells Dev.
21:1822–1830. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Jungwirth U, Xanthos DN, Gojo J, Bytzek
AK, Körner W, Heffeter P, Abramkin SA, Jakupec MA, Hartinger CG,
Windberger U, et al: Anticancer activity of methyl-substituted
oxaliplatin analogs. Mol Pharmacol. 81:719–728. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Shpilka T, Weidberg H, Pietrokovski S and
Elazar Z: Atg8: An autophagy-related ubiquitin-like protein family.
Genome Biol. 12:2262011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kabeya Y, Mizushima N, Ueno T, Yamamoto A,
Kirisako T, Noda T, Kominami E, Ohsumi Y and Yoshimori T: LC3, a
mammalian homologue of yeast Apg8p, is localized in autophagosome
membranes after processing. EMBO J. 19:5720–5728. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhou YY, Li Y, Jiang WQ and Zhou LF:
MAPK/JNK signalling: a potential autophagy regulation pathway.
Biosci Rep. 35:e01992015.
|
|
36
|
Xiong H, Zhang ZG, Tian XQ, Sun DF, Liang
QC, Zhang YJ, Lu R, Chen YX and Fang JY: Inhibition of JAK1,
2/STAT3 signaling induces apoptosis, cell cycle arrest, and reduces
tumor cell invasion in colorectal cancer cells. Neoplasia.
10:287–297. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yuan L, Wei S, Wang J and Liu X:
Isoorientin induces apoptosis and autophagy simultaneously by
reactive oxygen species (ROS)-related p53, PI3K/Akt, JNK, and p38
signaling pathways in HepG2 cancer cells. J Agric Food Chem.
62:5390–5400. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Jaber N, Dou Z, Chen JS, Catanzaro J,
Jiang YP, Ballou LM, Selinger E, Ouyang X, Lin RZ, Zhang J and Zong
WX: Class III PI3K Vps34 plays an essential role in autophagy and
in heart and liver function. Proc Natl Acad Sci USA. 109:2003–2008.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhang DM, Liu JS, Deng LJ, Chen MF, Yiu A,
Cao HH, Tian HY, Fung KP, Kurihara H, Pan JX and Ye WC:
Arenobufagin, a natural bufadienolide from toad venom, induces
apoptosis and autophagy in human hepatocellular carcinoma cells
through inhibition of PI3K/Akt/mTOR pathway. Carcinogenesis.
34:1331–1342. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Bai R, Ding T, Zhao J, Liu S, Zhang L, Lan
X, Yu Y and Yin L: The effect of PI3K inhibitor LY294002 and
gemcitabine hydrochloride combined with ionizing radiation on the
formation of vasculogenic mimicry of Panc-1 cells in vitro and in
vivo. Neoplasma. 63:80–92. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Li J, Yang D, Wang W, Piao S, Zhou J,
Saiyin W, Zheng C, Sun H and Li Y: Inhibition of autophagy by 3-MA
enhances IL-24- induced apoptosis in human oral squamous cell
carcinoma cells. J Exp Clin Cancer Res. 34:972015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Yamada H: Th17 cells in human rheumatoid
arthritis. Nihon Rinsho Meneki Gakkai Kaishi. 32:249–255. 2009.(In
Japanese). View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zou W and Restifo NP: T(H)17 cells in
tumour immunity and immunotherapy. Nat Rev Immunol. 10:248–256.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Novatchkova M, Leibbrandt A, Werzowa J,
Neubüser A and Eisenhaberemail F: The STIR-domain superfamily in
signal transduction, development and immunity. Trends Biochem Sci.
28:226–229. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Schraml BU, Hildner K, Ise W, Lee WL,
Smith WA, Solomon B, Sahota G, Sim J, Mukasa R, Cemerski S, et al:
The AP-1 transcription factor Batf controls T(H)17 differentiation.
Nature. 460:405–409. 2009.PubMed/NCBI
|
|
46
|
Zhang X, Weng W, Xu W, Wang Y, Yu W, Tang
X, Ma L, Pan Q, Wang J and Sun F: Prognostic significance of
interleukin 17 in cancer: A meta-analysis. Int J Clin Exp Med.
7:3258–3269. 2014.PubMed/NCBI
|
|
47
|
Zhang JP, Yan J, Xu J, Pang XH, Chen MS,
Li L, Wu C, Li SP and Zheng L: Increased intratumoral
IL-17-producing cells correlate with poor survival in
hepatocellular carcinoma patients. J Hepatol. 50:980–989. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Kryczek I, Banerjee M, Cheng P, Vatan L,
Szeliga W, Wei S, Huang E, Finlayson E, Simeone D, Welling TH, et
al: Phenotype, distribution, generation, and functional and
clinical relevance of Th17 cells in the human tumor environments.
Blood. 114:1141–1149. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Hambright HG and Ghosh R: Autophagy: In
the cROSshairs of cancer. Biochem Pharmacol. (In Press). PubMed/NCBI
|
|
50
|
Marin JJ, Lozano E and Perez MJ: Lack of
mitochondrial DNA impairs chemical hypoxia-induced autophagy in
liver tumor cells through ROS-AMPK-ULK1 signaling dysregulation
independently of HIF-1α. Free Radic Biol Med. 101:71–84. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Weiland A, Roswall P, Hatzihristidis TC,
Pietras K, Ostman A and Strell C: Fibroblast-dependependent
regulation of the stem cell properties of cancer cells. Neoplasma.
59:719–727. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Amaravadi RK, Yu D, Lum JJ, Bui T,
Christophorou MA, Evan GI, Thomas-Tikhonenko A and Thompson CB:
Autophagy inhibition enhances therapy-induced apoptosis in a
Myc-induced model of lymphoma. J Clin Invest. 117:326–336. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Yuan J, Zhang Y, Sheng Y, Fu X, Cheng H
and Zhou R: MYBL2 guides autophagy suppressor VDAC2 in the
developing ovary to inhibit autophagy through a complex of
VDAC2-BECN1-BCL2L1 in mammals. Autophagy. 11:1081–1098. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Xu Y, Li Z, Yin Y, Lan H, Wang J, Zhao J,
Feng J, Li Y and Zhang W: Ghrelin inhibits the differentiation of T
helper 17 cells through mTOR/STAT3 signaling pathway. PLoS One.
10:e01170812015. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Li P, Shi J, He Q, Hu Q, Wang YY, Zhang
LJ, Chan WT and Chen WX: Streptococcus pneumoniae Induces autophagy
through the inhibition of the PI3K-I/Akt/mTOR pathway and ROS
hypergeneration in A549 cells. PLoS One. 10:e01227532015.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Aveleira CA, Botelho M, Carmo-Silva S,
Pascoal JF, Ferreira-Marques M, Nóbrega C, Cortes L, Valero J,
Sousa-Ferreira L, Álvaro AR, et al: Neuropeptide Y stimulates
autophagy in hypothalamic neurons. Proc Natl Acad Sci USA.
112:E1642–E1651. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Wang P, Guo QS, Wang ZW and Qian HX: HBx
induces HepG-2 cells autophagy through PI3K/Akt-mTOR pathway. Mol
Cell Biochem. 372:161–168. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Huang F, Kao CY, Wachi S, Thai P, Ryu J
and Wu R: Requirement for both JAK-mediated PI3K signaling and
ACT1/TRAF6/TAK1-dependent NF-κappaB activation by IL-17A in
enhancing cytokine expression in human airway epithelial cells. J
Immunol. 179:6504–6513. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Sun X, Zhang J, Hou Z, Han Q, Zhang C and
Tian Z: miR-146a is directly regulated by STAT3 in human
hepatocellular carcinoma cells and involved in anti-tumor immune
suppression. Cell Cycle. 14:243–252. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Cross-Knorr S, Lu S, Perez K, Guevara S,
Brilliant K, Pisano C, Quesenberry PJ, Resnick MB and Chatterjee D:
RKIP phosphorylation and STAT3 activation is inhibited by
oxaliplatin and camptothecin and are associated with poor prognosis
in stage II colon cancer patients. BMC Cancer. 13:4632013.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Kim KW, Cho ML, Park MK, Yoon CH, Park SH,
Lee SH and Kim HY: Increased interleukin-17 production via a
phosphoinositide 3-kinase/Akt and nuclear factor kappaB-dependent
pathway in patients with rheumatoid arthritis. Arthritis Res Ther.
7:R139–R148. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
62
|
Huang C, Yang G, Jiang T, Huang K, Cao J
and Qiu Z: Effects of IL-6 and AG490 on regulation of Stat3
signaling pathway and invasion of human pancreatic cancer cells in
vitro. J Exp Clin Cancer Res. 29:512010. View Article : Google Scholar : PubMed/NCBI
|