Introduction
Cholangiocarcinoma (CCA) is a malignant tumor
arising from bile duct epithelial cells, which is the most common
form of cancer in Thailand and a major public health problem in the
northeast region (1). Several
epidemiological and experimental studies have provided support that
liver fluke (Opisthorchis viverrini; Ov) infection is the
etiological factor underlying the development of CCA (2–4). The
mechanistic link between host-parasite interaction and CCA
carcinogenesis has been reviewed previously (5). Our previous studies in a hamster animal
model revealed that chronic inflammation induced by repeated Ov
infection was a risk factor for CCA. Increased frequency Ov
infections can induce the accumulation of inducible nitric oxide
synthase (iNOS) in inflammatory cells and in the epithelium of bile
ducts, and subsequently causes nitrative and oxidative damage to
nucleic acids. In addition, NO may be involved in carcinogenesis
through heme oxygenase-1-derived iron accumulation, which may be
involved in the enhancement of oxidative DNA damage in the
epithelium of small bile ducts (6–9). It can be
concluded that repeated infection with Ov mediates oxidative and
nitrative DNA damage, and may be involved in the initiation and/or
promotion of CCA carcinogenesis through chronic inflammation.
Inflammatory cell infiltration triggered by repeated infections has
been shown to occur earlier than in single infections, and is
associated with alterations in liver enzymes and the severity of
periductal fibrosis (7). Furthermore,
Thanan et al (10) reported
that humans infected with Ov had an accumulation of urinary
8-oxo-7, 8-dihydro-2′-deoxyguanosine, which was positively
associated with plasma levels of nitrate/nitrite and aspartate
transaminase, indicating that Ov infection can induce oxidative and
nitrative stress via the production of reactive oxygen and reactive
nitrogen species during liver injury in humans. In addition, DNA
damage due to Ov infection-mediated lipid peroxidation has been
reported in animal and human studies (11–13).
Several cellular proteins in CCA tissue have been found to exist in
carbonylated forms via oxidative damage and subsequent dysfunction,
leading to the progression of CCA (14). The evidence to date suggests an
association between Ov infection, chronic inflammation and
subsequent CCA development.
The adaptive response of the genome protection
mechanism occurs in cells exposed to stress due to the
overproduction of free radicals via the inflammatory process and
infection. In such circumstances, cells maintain health via several
genome protection mechanisms. However, there is increasing evidence
that this adaptive response may have deleterious effects, for
example if there is a reduction of antioxidant enzymes and/or
imbalance in the DNA repair system, as this generates
microsatellite instability (MSI), which has procarcinogenic
implications. Evidence to support these effects have been shown in
studies by Guner et al (15)
and Hofseth et al (16). These
studies reported that low levels of antioxidant enzymes in
cancerous tissues were in the carcinogenic process and that the
adaptive imbalance in basal levels of excision-repair enzymes
generates MSI in chronic inflammation. Our previous study (17) revealed impairment of antioxidant
enzyme activity and increased expression of DNA repair enzymes in
hamster liver tissues, which were associated with the development
of cholangiocarcinoma in a hamster model.
Therefore, the present study hypothesized that MSI,
caused by imbalanced responses of antioxidant enzymes and/or DNA
repair enzymes due to oxidative/nitrative stress from the
inflammatory response, is involved in Ov-associated
cholangiocarcinogenesis. To confirm this hypothesis, the present
study aimed to identify the expression patterns of the antioxidant
enzymes, superoxide dismutase type 2 (SOD2) and catalase (CAT), and
the DNA repair enzymes, alkyladenine DNA glycosylase (AAG),
apurinic endonuclease (APE) and DNA polymerase β (DNA pol β), in
addition to determining the activites of the SOD2 and CAT
antioxidant enzymes in human and hamster CCA tissues. MSI was
investigated in human CCA tissues only. The results may provide
additional knowledge to explain the procarcinogenic effect of
cholangiocarcinoma caused by OV infection.
Materials and methods
White blood cells (WBCs) and
cholangiocarcinoma tissues
A total of 13 surgically resected liver specimens of
CCA patients and their WBCs were selected from the tissue bank of
the Liver Fluke and Cholangiocarcinoma Research Center at Khon Kaen
University (Khon Kaen, Thailand). Of the 13 CCA patients studied,
69% were male, resulting in a male to female ratio of 2.25:1. The
mean age was 53.9±8.9 years (range, 37–64 years). The control group
(n=5) comprised liver tissue obtained from cadaveric donors who had
succumbed to a car accident, and their kidneys were harvested for
transplantation. Informed consent was obtained from individual
patients and donors' relatives. The protocols for collection and
experiments were approved by the Ethics Committee for Human
Research, Khon Kaen University (approval no. HE571283).
Antibodies
The antibodies used for immunohistochemical staining
were as follows: Rabbit anti-SOD2 polyclonal antibody (dilution,
1:200; EMD Millipore, Billerica, MA, USA), rabbit anti-CAT
polyclonal antibody (dilution, 1:1,000; Abcam, Cambridge, MA, USA),
goat anti-AAG polyclonal antibody (dilution 1:100; Abcam), rabbit
anti-DNA pol β antibody (dilution 1:50; Abcam) and mouse anti-APE1
monoclonal antibody (dilution 1:50, Santa Cruz Biotechnology, Inc.,
Dallas, TX, USA).
Immunohistochemical investigation of
antioxidant and DNA repair enzymes
An immunohistochemical method was used to determine
the expression patterns of antioxidant enzymes and DNA repair
enzymes in human CCA tissues. Sections (size, 1.5×2 cm; thickness,
4 µm) of human liver tissues were deparaffinized and rehydrated in
a stepwise manner with decreasing concentrations of ethanol.
Antigen retrieval was performed via microwave treatment in 10 mM
citrate buffer (pH 6.0) at high power for 10 min, following which
the sections were immersed for 20 min in 3% (v/v) hydrogen peroxide
in PBS to inhibit endogenous hydrogen peroxide activity.
Nonspecific binding was blocked by skim milk in PBS for 30 min. The
sections were incubated with the primary antibodies in a moisture
chamber for 1 h at room temperature and at 4°C overnight.
Subsequently, the sections were washed in PBS and incubated with
peroxidase-conjugated Envision™ secondary antibody (Dako, Glostrup,
Denmark) in a moisture chamber for 1 hat room temperature.
Following washing in PBS, the color was developed using a 3,
3′diaminobenzidine tetrahydrochloride substrate kit for 5 min and
then counterstained with Mayer's haematoxylin. The sections were
rehydrated in a stepwise-manner with increasing concentrations of
ethanol, cleared with xylene and mounted with permount. The stained
sections were viewed under a microscope (Axio Scope.A1; Carl Zeiss
AG, Oberkochen, Germany). The protein expression levels were graded
by calculating the total immunostaining index as the product of the
frequency and intensity scores. The frequency score represented the
estimated fraction of positively-stained tumor cells (0=0%;
1=1–25%; 2=26–50%; 3=>50%). The intensity score represented the
estimated staining intensity (0, negative staining; 1, weak; 2,
moderate; 3, strong). The immunohistochemical index ranged between
0 and 9. Negative, low, moderate and high expression levels of SOD2
and CAT were defined as immunohistochemical indices 0, 1–2, 3–4 and
>4, respectively. Omission of the primary antibody served as a
negative control.
Investigation of enzymatic reactions
of antioxidant enzymes
To examine the activity of SOD2, the frozen human
CCA tissue was washed with PBS (pH 7.4) to remove any red blood
cells and clots. The tissue was homogenized in 1 ml of cold 20 mM
HEPES buffer (pH 7.2), which consisted of 1 mM EGTA, 210 mM
mannitol and 70 mM sucrose. The homogenate was centrifuged at 1,500
× g for 5 min at 4°C and the supernatant was removed to assess
total SOD activity (cytosolic and mitochondrial). To separate the
two compartments of the enzymes, 1,500 × g supernatant was
centrifuged at 10,000 × g for 15 min at 4°C, the supernatant
contained cytosolic SOD and the pellet contained mitochondrial SOD.
The mitochondrial pellet was homogenized in HEPES buffer to
determine mitochondrial SOD2 activity.
To determine the activity of CAT, the frozen human
CCA tissue was washed with PBS (pH 7.4) to remove any red blood
cells and clots. The tissue was homogenized in 1 ml of 50 mM
potassium phosphate (pH 7.0), containing 1 mM EDTA, and the
homogenate was centrifuged at 10,000 × g for 15 min at 4°C
to extract proteins. The supernatant was used to assess the
activity of CAT.
Subsequently, the extracted SOD2 and CAT proteins
were used for enzymatic reactions. SOD and CAT assay kits were
purchased from Cayman Chemical Company (Ann Arbor, MI, USA. The
principle of the SOD assay kit is that it utilizes a tetrazolium
salt for the detection of superoxide radicals generated by xanthine
oxidase and hypoxanthine. The absorbance was measured at 440–460 nm
on an ELISA reader (Tecan, Männedorf, Switzerland). An SOD level of
1 unit was considered the quantity of enzyme required to exhibit
50% dismutation of the superoxide radical. The CAT assay kit
utilizes the peroxidatic function of CAT to determine enzyme
activity. The method is based on the reaction of the enzyme with
methanol, in the presence of an optimal concentration of
H2O2. The formaldehyde produced is measured
spectrophotometrically with 4-amino-3-hydrazino-5-mercapto-1, 2,
4-trizazole as the chromogen. The absorbance was read at 540 nm
using an ELISA reader (Tecan). The activity of SOD is presented as
U/ml/mg protein, whereas the activity of CAT activity is presented
as µM/min/mg protein.
DNA extraction
The paraffinized sections of CCA tissue were
deparaffinized twice in xylene for 5 min to remove the paraffin
wax. Following deparaffinization, the slide was hydrated in
decreasing concentrations of ethanol (absolute alcohol for 3 min;
70% alcohol for 3 min) and rinsed in tap water. Following
hydration, the slide was stained with hematoxylin for 5 min, washed
in running tap water for 2 min, destained in acid alcohol (1% acid
in 70% alcohol), washed in running tap water again, and dehydrated
(70% alcohol for 2 min; absolute alcohol for 2 min) twice.
Successful staining of the nuclei was indicated by a blue color.
Following visualization under an Axio Scope.A1 microscope (Carl
Zeiss AG) and separation of tumorous and non-tumorous tissues,
extraction was performed using a QIAamp DNA formalin-fixed
paraffin-embedded tissue kit (Qiagen, Inc., Valencia, CA, USA).
Matched WBCs were prepared for DNA extraction by digestion in 1 ml
of lysis buffer, 0.2 µl of 10 mg/ml RNase and 10 µl of 17 mg/ml
proteinase K at 50°C for 30 min. Protein was precipitated by adding
300 µl of NaI solution. Isopropanol was added to the supernatant in
order to precipitate DNA. The DNA pellet was then resuspended in TE
buffer (pH 8.0).
MSI analysis
The DNA extracted from tumor tissues, non-tumor
tissues of the adjacent area and WBCs of the same patient were
analyzed for MSI using five microsatellite markers: BAT25, BAT26,
D5S346, D2S123 and D17S250 (18).
Oligonucleotide forward primers were fluorescent-5′-labeled.
Polymerase chain reaction (PCR) analysis was performed in a 20 ml
reaction mixture with 10–100 ng genomic DNA, 0.2 mM of each dNTP,
1X high fidelity PCR buffer, 0.2 µM of each primer and 1 unit Taq
DNA polymerase high fidelity (Invitrogen; Thermo Fisher Scientific,
Inc., Waltham, MA, USA). The sequences of each primer are shown in
Table I. The amplification conditions
for BAT25, BAT26 and D17S250 were as follows: 5 min initial
denaturation at 94°C, followed by 30 cycles of 30 sec at 94°C, 30
sec at 50°C and 1 min at 68°C, with a final extension of 10 min at
68°C. The amplification conditions for D2S123 and D5S346 were as
follows: 5 min initial denaturation at 94°C, followed by 30 cycles
of 30 sec at 94°C, 30 sec at 55°C and 1 min at 68°C, with a final
extension of 10 min at 72°C. The MSI was then analyzed in an ABI
Prism 3130 genetic analyzer (Applied Biosystems; Thermo Fisher
Scientific, Inc.) using GeneScan Analysis v3.2.1 software (Applied
Biosystems; Thermo Fisher Scientific, Inc). MSI was indicated as
alterations in the densities of bands observed in the tumor tissue
and non-tumor of adjacent area, which were not present in the
normal control WBCs from the same patient. Tumors exhibiting MSI at
two or more markers were defined as MSI-high (MSI-H). Tumors
showing instability at only one marker were defined as MSI-low
(MSI-L). Tumors in which no markers exhibited MSI were considered
to show microsatellite stability (MSS).
 | Table I.Specific primers for microsatellite
polymerase chain reaction analysis. |
Table I.
Specific primers for microsatellite
polymerase chain reaction analysis.
| Primer | Linked gene | Sequence |
|---|
| BAT25 | c-kit | Forward 5′-TCG CCT
CCA AGA ATG TAA GT-3′ |
|
|
| Reverse 5′-TCT GCA
TTT TAA CTA TGG CTC-3′ |
| BAT26 | hMSH2 | Forward 5′-TGA CTA
CTT TTG ACT TCA GCC-3′ |
|
|
| Reverse 5′-AAC CAT
TCA ACA TTT TTA ACC C-3′ |
| D2S123 | hMSH2 | Forward 5′-ACA TTG
CTG GAA GTT CTG GC-3′ |
|
|
| Reverse 5′-CCT TTC
TGA CTT GGA TAC CA-3′ |
| D5S346 | APC | Forward 5′-ACT CAC
TCT AGT GAT AAA TCG G-3′ |
|
|
| Reverse 5′-GTT TCC
ATT GTA GCA TCT TGA C-3′ |
| D17S250 | p53 | Forward 5′-GCT GGC
CAT ATA TAT ATT TAA ACC-3′ |
|
|
| Reverse 5′-GGA AGA
ATC AAA TAG ACA AT-3′ |
Statistical analysis
Statistical analysis was performed using GraphPad
Prism version 5 (GraphPad Software Inc., La Jolla, CA, USA). The
Mann-Whitney U test was used to compare differences between the
cadaveric donor and human CCA. Clinicopathological characteristics
and MSI (low and high) in human CCA tissues were compared using
Fisher's exact probability test. The Kaplan-Meier method was used
to calculate the survival curves, and the Log-rank test was
performed to compare differences in the survival rates of patients
who were subjected to curative surgery. P<0.05 was considered to
indicate a statistically significant difference.
Results
Protein expression of antioxidant
enzymes and DNA repair enzymes in human CCA tissues
Immunohistochemical analysis was performed to detect
the expression levels of SOD2 and CAT in human CCA tissues. The
results demonstrated that SOD2 and CAT were expressed in the
cytosolic bile duct CCA tissues. The majority of the normal bile
duct epithelia in the tissues adjacent to the tumor sections
stained positively for SOD2 and CAT. The expression of SOD2 and CAT
were also observed in hepatocytes. The numbers of samples with SOD2
at low, moderate and high levels of expression were 15 (55%), eight
(30%) and four (15%), respectively, whereas the number of samples
with CAT at negative, low, moderate and high levels of expression
were four (15%), 11 (40%), eight (30%) and four (15%),
respectively. The representative results of SOD2 and CAT staining
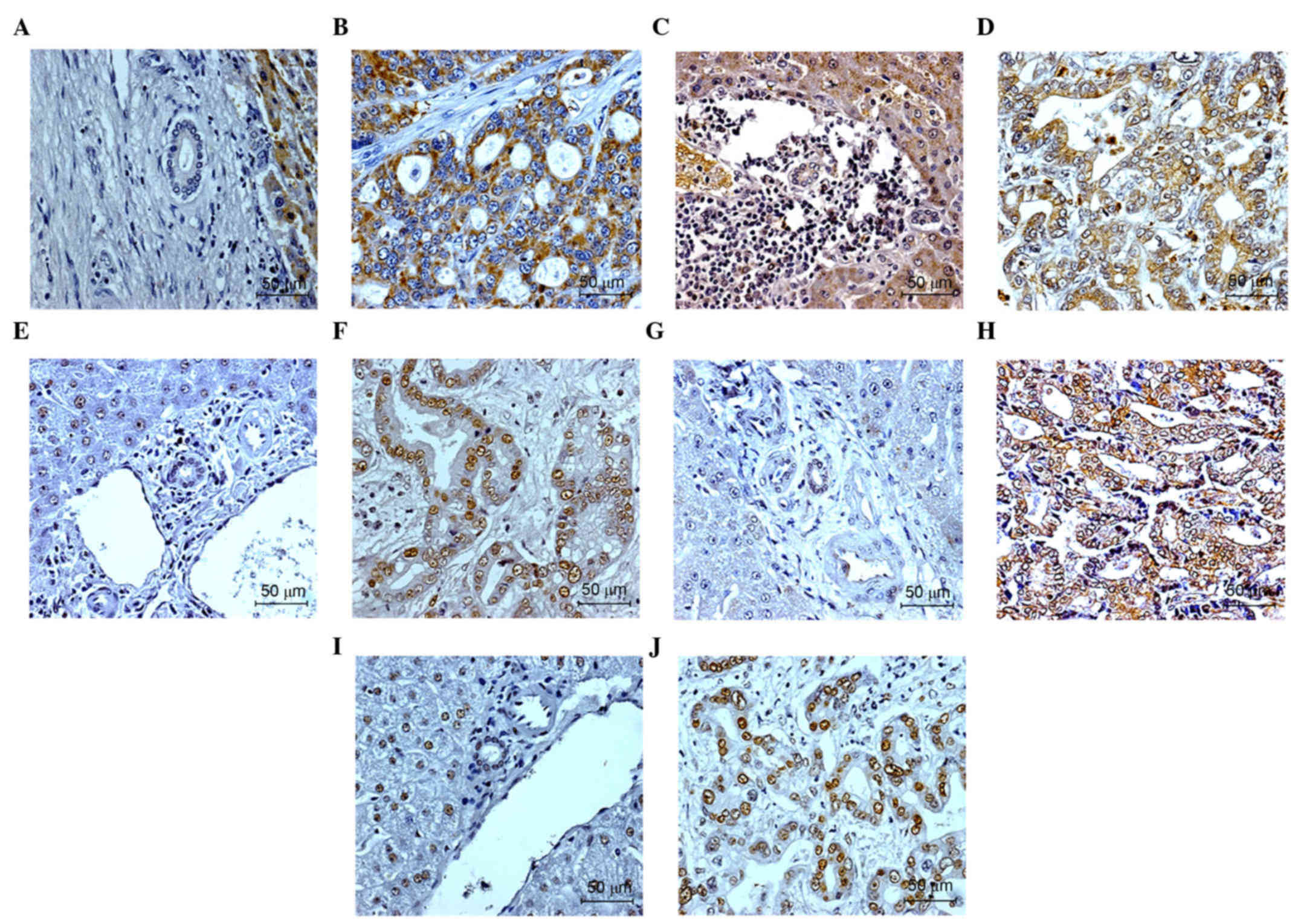
in the CCA tissue are shown in Fig.
1A-D.
The present study also examined the expression
levels of AAG, APE and DNA pol β in human CCA tissues using
immunohistochemistry. The results showed that AAG, APE and DNA pol
β were expressed in the nuclei of bile duct CCA tissues. In the
normal bile duct epithelia residing in tissue adjacent to the tumor
sections, weak staining for AAG, APE and DNA pol β were observed.
In the hepatocytes, positive staining was observed for AAG and DNA
pol β, but not for APE (Fig. 1). The
numbers of samples with low, moderate and high expression levels of
AAG were four (15%), eight (30%) and 15 (55%), respectively;
numbers of samples with negative, low, moderate and high expression
levels of APE were five (19%), 15 (55%), five (19%) and two (7%),
respectively. The number of samples with DNA pol β with moderate
and high levels of expression were eight (30%) and 19 (70%),
respectively. The representative results of AAG, APE and DNA pol β
in the patients with CCA are shown in Fig. 1E-J.
Activities of antioxidant enzymes in
human CCA tissues
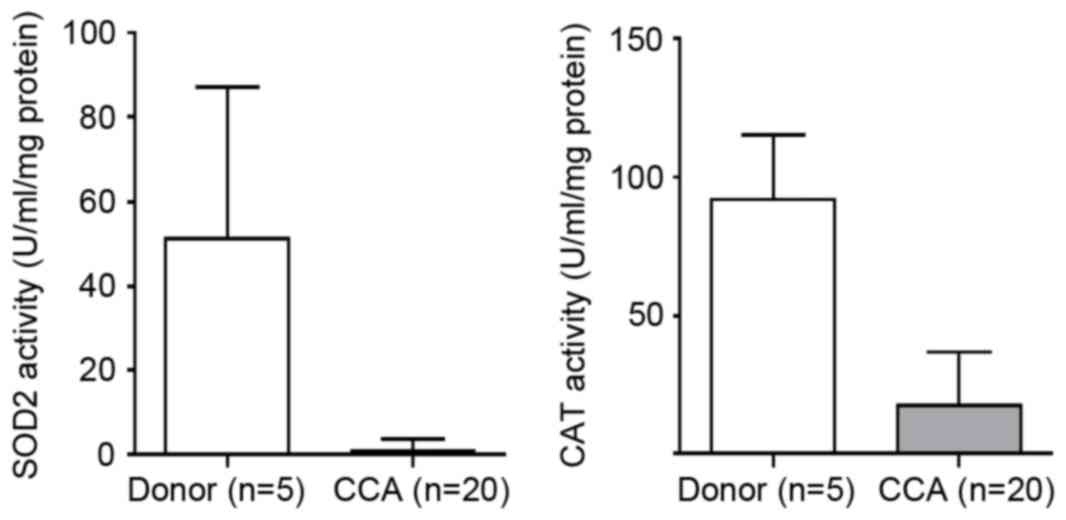
The activities of SOD2 and CAT were examined in
human CCA tissues. The results showed that the activities of SOD2
and CAT were significantly decreased in the CCA tissues, compared
with the tissues from the cadaveric donors (P=0.04 and P=0.004,
respectively), as shown in Fig.
2.
MSI analysis in human CCA tissues
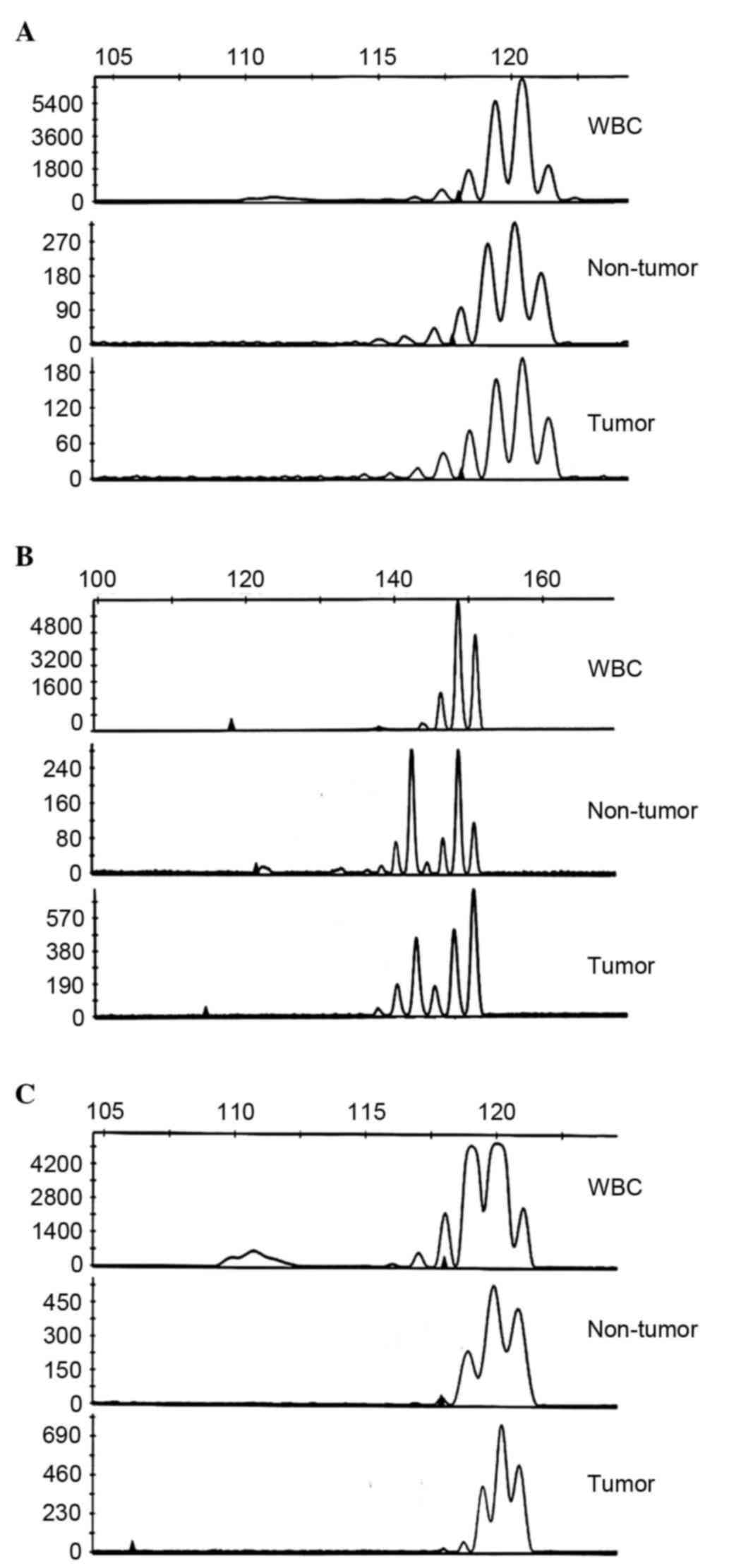
The present study examined MSI in 13 patients with
CCA using five microsatellite markers (18). MSI was indicated by altered band
density in the tumor tissue and adjacent non-tumor tissue, which
was not present in normal control WBCs from the same patient. The
results are presented in Table II.
Of the 13 CCA samples, four (31%) were identified as MSI-low and
nine (69%) were identified as MSI-high. None of the cases exhibited
MSS. A representative electropherograms from the assessment of MSI
are shown in Fig. 3A-C.
| Table II.Microsatellite marker analysis and
MSI status. |
Table II.
Microsatellite marker analysis and
MSI status.
|
| Microsatellite
marker |
|
|---|
|
|
|
|
|---|
| Patient ID | BAT25 | BAT26 | D2S123 | D5S346 | D17S250 | MSI diagnosis |
|---|
| R23 | + | − | + | − | + | MSI-high |
| R26 | + | − | + | − | + | MSI-high |
| R49 | + | − | − | − | − | MSI-low |
| R52 | + | − | − | − | − | MSI-low |
| R81 | − | − | − | + | + | MSI-high |
| R100 | + | − | + | + | − | MSI-high |
| R101 | + | − | − | + | − | MSI-high |
| R103 | − | − | − | + | − | MSI-low |
| R104 | − | − | + | + | + | MSI-high |
| R118 | − | − | + | + | − | MSI-high |
| R123 | + | − | + | + | − | MSI-high |
| R134 | − | − | − | + | − | MSI-low |
| R149 | + | − | + | − | − | MSI-high |
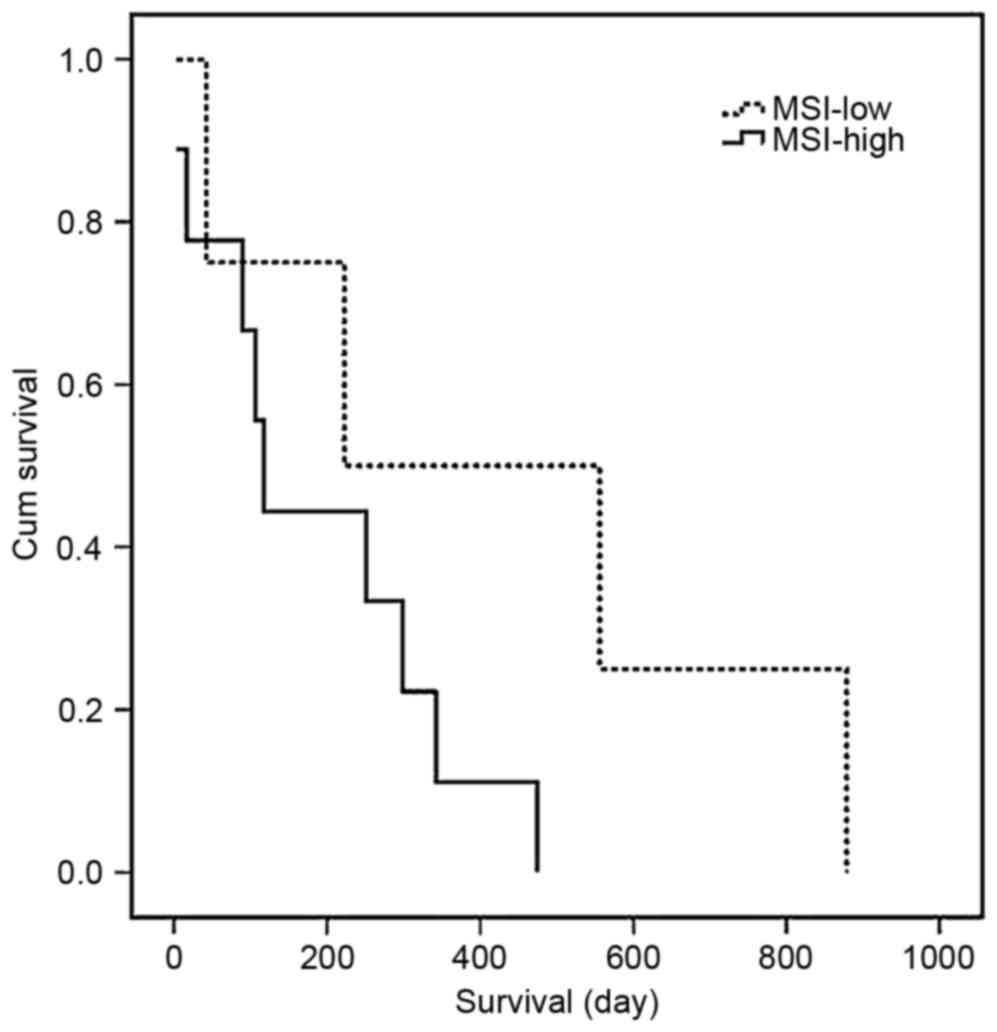
The present study compared the clinical and
histological tumor characteristics of the MSI-high tumors with
those of the MSI-low tumors. No differences were observed in age,
gender, histological type or metastasis. In comparing the patients
with MSI-low and patients with MSI-high, the 5-year survival rate
of the patients in the MSI-low (n=4) group was 31%, whereas that of
the patients in the MSI-high (n=9) group was 69%, indicating that
the survival rate was higher in the MSI-low group, as shown in
Fig. 4. However, no significant
difference between these two groups was found (P=0.14). Of note,
the expression of APE showed a significant negative correlation
with MSI status (Spearman's rank correlation test, −0.639;
P=0.019).
Discussion
Impairment of the antioxidant system may favor the
accumulation of free radicals, as it has been found that low levels
of essential antioxidants are associated with an increased risk of
cancer (19). The results of the
present study demonstrated a significant decrease in the expression
levels of the SOD2 and CAT enzymes in human CCA tissue. This result
was consistent with a previous observation in prostatic
adenocarcinoma (20), which
demonstrated that the expression levels of SOD2 and CAT were
significantly lower in cancer tissue, compared with than that in
normal tissue. In addition, the present study found that the
activities of SOD2 and CAT in human CCA were significantly
decreased, compared with those in the cadaveric donor. Decreased
activities of SOD and CAT, and an increase in DNA lesions have been
suggested to be caused by free radicals in lung cancer tissues in
humans (21). In conjunction with
these results, Guner et al (15) found lower total activities of SOD and
CAT in lung carcinoma tissue. It has also been demonstrated that
levels of lipid peroxidation products and nitric oxide products are
significantly elevated, whereas the levels of SOD and CAT enzymes
are significantly lower in patients with oral cancer, compared with
normal healthy subjects (22).
The induction of an oxidant-generating system and
reduction of an antioxidant system are well-recognized causes of
oxidative stress. Oxidant-generating enzymes, including inducible
iNOS and cyclooxygenase 2 have been found to be upregulated in
animals with Ov-induced CCA, in vitro cell lines and
clinical samples of CCA (23–25). Plasma levels of nitrate/nitrite, which
are by-products of reactive nitrogen species, are significantly
increased in Ov-infected subjects and patients with Ov-associated
CCA, compared with uninfected healthy subjects (10). In addition, inflammatory cytokines
induce the formation of NO, and suppress the redox ratios of
glutathione and glutathione disulfide in CCA cells (26). Previously, a significant decrease in
the protein expression of manganese-SOD was reported in the cancer
cells of CCA tissues, compared with individual normal bile duct
cells located in regions adjacent to the tumor (14). Furthermore, the results of our
previous study showed that the decreased gene expression of CAT in
the peripheral blood leukocytes of patients with opisthorchiasis is
induced by Ov antigen (27).
Additionally CAT was found to be downregulated in an animal model
of opisthorchaiasis-associated CCA, detected via kinetic analysis
using the cDNA microarray technique (28).
These results are in agreement with those on an
associated liver fluke, Fasciola hepatica, in which
antioxidant potential was also shown to decrease in rat livers
following infection (29). This
reduction of antioxidant systems may increase oxidative damage to a
wide range of biomolecules, including DNA, lipids, carbohydrates
and proteins. The oxidation of lipids by excess levels of reactive
oxygen species/reactive nitrogen species generates by-products
including trans-4-hydroxy-2- nonenal, malondialdehyde and
crotonaldehyde, which can react with DNA bases to form a variety of
exocyclic DNA adducts (30–32). An increase in the formation of
1,N(6)-etheno-2′-deoxyadenosine and
3,N(4)-etheno-2′-deoxycytidine in CCA
tissues has been revealed (11,12). These
etheno modified DNA bases are miscoding lesions, which are
considered to contribute to the initiation of carcinogenesis
through the induction of specific point mutations in DNA.
Therefore, base excision repair (BER) is a key pathway used in the
protection of free radical attack on DNA (33). The present study revealed increased
expression levels of the AAG and DNA pol β enzymes, whereas the
expression of APE decreased.
The present study also found MSI-low (31%) and
MSI-high (69%) in human CCA tissues. MSI-high was correlated with
poor prognosis in patients, as indicated by shorter survival rates,
compared with the group of MSI-low patients; however, this was not
statistically significant, which may be due to the small sample
size. Liengswangwong et al reported MSI-low and MSS in liver
fluke infection-associated intrahepatic CCA (34). In addition, the prognostic and
predictive impacts of MSI status in patients with colon cancer have
been reported (35,36). Previously, it was demonstrated the
generation of a mutator phenotype in yeast was caused by imbalanced
BER (37). The data obtained
suggested a mechanism by which spontaneous mutation rates may be
modulated. An imbalance between apurinic/apyrimidinic (AP)
endonuclease and one particular DNA glycosylase produced a mutator
phenotype in yeast. Imbalance in the glycosylase to AP endonuclease
ratio may arise by promoter mutations, which affect the expression
of DNA repair genes. Imbalances in BER enzymes may represent a
previously unrecognized source of increased cancer risk.
Furthermore, imbalanced BER increases spontaneous mutation and
alkylation sensitivity in Escherichia coli (38). In a previous study of human colon
adenoma showing dysregulation of BER, a correlation was found
between the expression of AAG and iNOS. Nitration in the AAG active
site of tyrosine 162 in vitro was found to impair
εA-excision activity, whereas nitrosation of cysteine 167 increased
εA-excision activity. Therefore, the post-translational
modification of AAG by reactive nitrogen species via the
overproduction of NO appears to be one mechanism of BER
dysregulation leading to malignancy (39). In ulcerative colitis, it has been
shown that the adaptive imbalance in BER enzymes generates MSI in
chronic inflammation.
Mechanistic studies using yeast and human cell
models overexpressing AAG and/or APE1 were associated with
frameshift mutations and MSI, indicating that the adaptive
imbalanced increase in BER enzymes may have DNA-damaging effects
and may contribute to carcinogenesis in chronic inflammation
(16). Furthermore, studies have
shown that the overexpression of DNA pol β may be associated with
MSI, which can lead to malignancy. DNA pol β has a high level of
infidelity in replicating DNA in vitro, due to the lack of
associated proofreading activity and poor discrimination of
nucleotides. This may be due to error-prone features, whereas the
expression is tightly regulated in vivo, and constant and
low throughout the cell-cycle (40).
The overexpression of DNA pol β in mammalian cells has been
demonstrated to significantly increase mutation rate (41), which can consequently induce
chromosomal instability and enhance the tumorigenicity of
xenografts (42). Yamada and Farber
(43) reported the induction of a low
level of MSI by the overexpression of DNA pol β, and a mechanism
was identified for the induction of microsatellite mutations by
using a selection-based cell culture system with high sensitivity
to relatively small increases in mutation rates. These results
suggested that the overexpression of DNA pol β may lead to
increased genetic instability.
The results of the present study showed that a
reduction in the expression of antioxidant enzymes and the adaptive
imbalance of BER enzymes may cause MSI in human CCA. In addition,
the results showed that patients with MSI-low had improved survival
rates, compared with patients with MSI-high, which may increase
susceptibility to tumorigenesis.
Acknowledgements
This study was supported by the Invitation Research
Grant (grant no. I56130) and the Research Assistantship Grant of
the Faculty of Medicine, Khon Kaen University (grant no. AS57202)
to Dr Watcharin Loilomewl, the Scholarship of the Liver Fluke and
Cholangiocarcinoma Research Center to Miss Sasithorn Kadsanitsk and
grants from the Higher Education Research Promotion and National
Research University Project of Thailand, Office of the Higher
Education Commission, through the Center of Excellence in Specific
Health Problems in Greater Mekong Sub-region cluster, Khon Kaen
Universitym, allocated to Dr Watcharin Loilomewl. The authors would
like to thank Professor Ross H Andrews for editing of the
manuscript via Publication Clinic KKU, Thailand.
References
|
1
|
Khuntikeo N, Chamadol N, Yongvanit P, et
al: CASCAP investigators: Cohort profile: Cholangiocarcinoma
screening and care program (CASCAP). BMC Cancer. 15:4592015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Flavell DJ and Lucas SB: Promotion of
N-nitrosodimethylamine-initiated bile duct carcinogenesis in the
hamster by the human liver fluke, Opisthorchis viverrini.
Carcinogenesis. 4:927–930. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ohshima H and Bartsch H: Chronic
infections and inflammatory processes as cancer risk factors:
Possible role of nitric oxide in carcinogenesis. Mutat Res.
305:253–264. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Thamavit W, Bhamarapravati N, Sahaphong S,
et al: Effects of dimethylnitrosamine on induction of
cholangiocarcinoma in Opisthorchis viverrini-infected Syrian golden
hamsters. Cancer Res. 38:4634–4639. 1978.PubMed/NCBI
|
|
5
|
Yongvanit P, Pinlaor S and Bartsch H:
Oxidative and nitrative DNA damage: Key events in
opisthorchiasis-induced carcinogenesis. Parasitol Int. 61:130–135.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Pinlaor S, Hiraku Y, Ma N, Yongvanit P,
Semba R, Oikawa S, Murata M, Sripa B, Sithithaworn P and Kawanishi
S: Mechanism of NO-mediated oxidative and nitrative DNA damage in
hamsters infected with Opisthorchis viverrini: A model of
inflammation-mediated carcinogenesis. Nitric Oxide. 11:175–183.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Pinlaor S, Ma N, Hiraku Y, Yongvanit P,
Semba R, Oikawa S, Murata M, Sripa B, Sithithaworn P and Kawanishi
S: Repeated infection with Opisthorchis viverrini induces
accumulation of 8-nitroguanine and
8-oxo-7,8-dihydro-2′-deoxyguanine in the bile duct of hamsters via
inducible nitric oxide synthase. Carcinogenesis. 25:1535–1542.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Pinlaor S, Sripa B, Sithithaworn P and
Yongvanit P: Hepatobiliary changes, antibody response, and
alteration of liver enzymes in hamsters re-infected with
Opisthorchis viverrini. Exp Parasitol. 108:32–39. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Pinlaor S, Yongvanit P, Hiraku Y, Ma N,
Semba R, Oikawa S, Murata M, Sripa B, Sithithaworn P and Kawanishi
S: 8-nitroguanine formation in the liver of hamsters infected with
Opisthorchis viverrini. Biochem Biophys Res Commun. 309:567–571.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Thanan R, Murata M, Pinlaor S,
Sithithaworn P, Khuntikeo N, Tangkanakul W, Hiraku Y, Oikawa S,
Yongvanit P and Kawanishi S: Urinary
8-oxo-7,8-dihydro-2′-deoxyguanosine in patients with parasite
infection and effect of antiparasitic drug in relation to
cholangiocarcinogenesis. Cancer Epidemiol Biomarkers Prev.
17:518–524. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Dechakhamphu S, Pinlaor S, Sitthithaworn
P, Bartsch H and Yongvanit P: Accumulation of miscoding etheno-DNA
adducts and highly expressed DNA repair during liver fluke-induced
cholangiocarcinogenesis in hamsters. Mutat Res. 691:9–16. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Dechakhamphu S, Pinlaor S, Sitthithaworn
P, Nair J, Bartsch H and Yongvanit P: Lipid peroxidation and etheno
DNA adducts in white blood cells of liver fluke-infected patients:
Protection by plasma alpha-tocopherol and praziquantel. Cancer
Epidemiol Biomarkers Prev. 19:310–318. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Dechakhamphu S, Yongvanit P, Nair J,
Pinlaor S, Sitthithaworn P and Bartsch H: High excretion of etheno
adducts in liver fluke-infected patients: Protection by
praziquantel against DNA damage. Cancer Epidemiol Biomarkers Prev.
17:1658–1664. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Thanan R, Oikawa S, Yongvanit P, Hiraku Y,
Ma N, Pinlaor S, Pairojkul C, Wongkham C, Sripa B, Khuntikeo N, et
al: Inflammation-induced protein carbonylation contributes to poor
prognosis for cholangiocarcinoma. Free Radic Biol Med.
52:1465–1472. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Guner G, Islekel H, Oto O, Hazan E and
Açikel U: Evaluation of some antioxidant enzymes in lung carcinoma
tissue. Cancer Lett. 103:233–239. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hofseth LJ, Khan MA, Ambrose M, Nikolayeva
O, Xu-Welliver M, Kartalou M, Hussain SP, Roth RB, Zhou X, Mechanic
LE, et al: The adaptive imbalance in base excision-repair enzymes
generates microsatellite instability in chronic inflammation. J
Clin Invest. 112:1887–1894. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Loilome W, Kadsanit S, Namwat N, Techasen
A, Puapairoj A, Dechakhamphu A, Pinitsoontorn C and Yongvanit P:
Impaired antioxidant enzyme activity and increased DNA repair
enzyme expression in hamster liver tissues related to
cholangiocarcinoma development. Asian Pac J Cancer Prev. (13
Suppl). S59–S64. 2012.
|
|
18
|
Boland CR, Thibodeau SN, Hamilton SR,
Sidransky D, Eshleman JR, Burt RW, Meltzer SJ, Rodriguez-Bigas MA,
Fodde R, Ranzani GN and Srivastava S: A National Cancer Institute
Workshop on Microsatellite Instability for cancer detection and
familial predisposition: Development of international criteria for
the determination of microsatellite instability in colorectal
cancer. Cancer Res. 58:5248–5257. 1998.PubMed/NCBI
|
|
19
|
Scibior D, Skrzycki M, Podsiad M and
Czeczot H: Glutathione level and glutathione-dependent enzyme
activities in blood serum of patients with gastrointestinal tract
tumors. Clin Biochem. 41:852–858. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Baker AM, Oberley LW and Cohen MB:
Expression of antioxidant enzymes in human prostatic
adenocarcinoma. Prostate. 32:229–233. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jaruga P, Zastawny TH, Skokowski J,
Dizdaroglu M and Olinski R: Oxidative DNA base damage and
antioxidant enzyme activities in human lung cancer. FEBS Lett.
341:59–64. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Beevi SS, Rasheed AM and Geetha A:
Evaluation of oxidative stress and nitric oxide levels in patients
with oral cavity cancer. Jpn J Clin Oncol. 34:379–385. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Itatsu K, Sasaki M, Yamaguchi J, Ohira S,
Ishikawa A, Ikeda H, Sato Y, Harada K, Zen Y, Sato H, et al:
Cyclooxygenase-2 is involved in the up-regulation of matrix
metalloproteinase-9 in cholangiocarcinoma induced by tumor necrosis
factor-alpha. Am J Pathol. 174:829–841. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Pinlaor S, Hiraku Y, Yongvanit P,
Tada-Oikawa S, Ma N, Pinlaor P, Sithithaworn P, Sripa B, Murata M,
Oikawa S and Kawanishi S: iNOS-dependent DNA damage via NF-kappaB
expression in hamsters infected with Opisthorchis viverrini and its
suppression by the antihelminthic drug praziquantel. Int J Cancer.
119:1067–1072. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wu T: Cyclooxygenase-2 and prostaglandin
signaling in cholangiocarcinoma. Biochim Biophys Acta.
1755:135–150. 2005.PubMed/NCBI
|
|
26
|
Prawan A, Buranrat B, Kukongviriyapan U,
Sripa B and Kukongviriyapan V: Inflammatory cytokines suppress NAD
(P)H: Quinone oxidoreductase-1 and induce oxidative stress in
cholangiocarcinoma cells. J Cancer Res Clin Oncol. 135:515–522.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yongvanit P, Thanan R, Pinlaor S,
Sithithaworn P, Loilome W, Namwat N, Techasen A and Dechakhamphu S:
Increased expression of TLR-2, COX-2 and SOD-2 genes in the
peripheral blood leukocytes of opisthorchiasis patients induced by
Opisthorchis viverrini antigen. Parasitol Res. 110:1969–1977. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wu Z, Boonmars T, Boonjaraspinyo S, Nagano
I, Pinlaor S, Puapairoj A, Yongvanit P and Takahashi Y: Candidate
genes involving in tumorigenesis of cholangiocarcinoma induced by
Opisthorchis viverrini infection. Parasitol Res. 109:657–673. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kolodziejczyk L, Siemieniuk E and
Skrzydlewska E: Antioxidant potential of rat liver in experimental
infection with Fasciola hepatica. Parasitol Res. 96:367–372. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
el Ghissassi F, Barbin A, Nair J and
Bartsch H: Formation of 1,N6-ethenoadenine and 3, N4-ethenocytosine
by lipid peroxidation products and nucleic acid bases. Chem Res
Toxicol. 8:278–283. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Nair J, De Flora S, Izzotti A and Bartsch
H: Lipid peroxidation-derived etheno-DNA adducts in human
atherosclerotic lesions. Mutat Res. 621:95–105. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Singer B and Hang B: Mammalian enzymatic
repair of etheno and para-benzoquinone exocyclic adducts derived
from the carcinogens vinyl chloride and benzene. IARC Sci Publ.
233–247. 1999.PubMed/NCBI
|
|
33
|
Bohr VA: Repair of oxidative DNA damage in
nuclear and mitochondrial DNA, and some changes with aging in
mammalian cells. Free Radic Biol Med. 32:804–812. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Liengswangwong U, Nitta T, Kashiwagi H,
Kikukawa H, Kawamoto T, Todoroki T, Uchida K, Khuhaprema T, Karalak
A, Srivatanakul P and Miwa M: Infrequent microsatellite instability
in liver fluke infection-associated intrahepatic
cholangiocarcinomas from Thailand. Int J Cancer. 107:375–380. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Losso GM, Rda S Moraes, Gentili AC and
Messias-Reason IT: Microsatellite instability-MSI markers (BAT26,
BAT25, D2S123, D5S346, D17S250) in rectal cancer. Arq Bras Cir Dig.
25:240–244. 2012.(In English, Portuguese). View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Saridaki Z, Souglakos J and Georgoulias V:
Prognostic and predictive significance of MSI in stages II/III
colon cancer. World J Gastroenterol. 20:6809–6814. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Glassner BJ, Rasmussen LJ, Najarian MT,
Posnick LM and Samson LD: Generation of a strong mutator phenotype
in yeast by imbalanced base excision repair. Proc Natl Acad Sci
USA. 95:9997–10002. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Posnick LM and Samson LD: Imbalanced base
excision repair increases spontaneous mutation and alkylation
sensitivity in Escherichia coli. J Bacteriol. 181:6763–6771.
1999.PubMed/NCBI
|
|
39
|
Jones LE Jr, Ying L, Hofseth AB, Jelezcova
E, Sobol RW, Ambs S, Harris CC, Espey MG, Hofseth LJ and Wyatt MD:
Differential effects of reactive nitrogen species on DNA base
excision repair initiated by the alkyladenine DNA glycosylase.
Carcinogenesis. 30:2123–2129. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zmudzka BZ, Fornace A, Collins J and
Wilson SH: Characterization of DNA polymerase beta mRNA: Cell-cycle
and growth response in cultured human cells. Nucleic Acids Res.
16:9587–9596. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Canitrot Y, Cazaux C, Fréchet M, Bouayadi
K, Lesca C, Salles B and Hoffmann JS: Overexpression of DNA
polymerase beta in cell results in a mutator phenotype and a
decreased sensitivity to anticancer drugs. Proc Natl Acad Sci USA.
95:12586–12590. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Bergoglio V, Pillaire MJ, Lacroix-Triki M,
Raynaud-Messina B, Canitrot Y, Bieth A, Garès M, Wright M, Delsol
G, Loeb LA, et al: Deregulated DNA polymerase beta induces
chromosome instability and tumorigenesis. Cancer Res. 62:3511–3514.
2002.PubMed/NCBI
|
|
43
|
Yamada NA and Farber RA: Induction of a
low level of microsatellite instability by overexpression of DNA
polymerase Beta. Cancer Res. 62:6061–6064. 2002.PubMed/NCBI
|