Introduction
Cervical cancer is the fourth most common cancer
among women worldwide and is considered an important public health
problem (1). According to the World
Health Organization, this type of tumor is the second most common
among women in less developed countries (1). In 2012, it was estimated the diagnosis
of 528,000 new cases and 270,000 mortalities of women by this type
of tumor in the world (1).
Cervical cancer develops from premalignant precursor
lesions known as cervical intraepithelial neoplasia (CIN).
According to the cytological degree of commitment, CINs are
classified as low-grade squamous intraepithelial lesions (LSILs) or
high-grade squamous intraepithelial lesions (HSILs) (2). Studies have shown that the immune system
serves a critical role in the elimination of these lesions, and
defects in the function of this system appear to be associated with
the mechanism of tumor escape from immune control (3,4).
Evidence suggests that dysfunction can be reversed
by stimulating the immune system with antigen-presenting cells and
cytokines such as interleukin (IL)-2 (5,6). Dendritic
cells (DCs) are known as professional antigen-presenting cells and
are part of the innate immune system. These cells can be detected
in the majority of peripheral tissues, where they act on the
initiation and modulation of the immune response during infections
by pathogens and on the development of antitumor immune responses
(7,8).
DCs are differentiated from pluripotent precursors
located in the bone marrow. They have two distinct pathways of
differentiation: i) The myeloid pathway, which generates myeloid
DCs, including Langerhans cells, dermal DCs and interstitial DCs,
which are characterized by the expression of the myeloid marker
cluster of differentiation (CD)11c; and ii) the lymphoid pathway,
which generates plasmacytoid DCs, which are characterized by the
expression of the cell marker CD123 (7,9,10).
These cells are transported from the bloodstream to
peripheral tissues in an immature form known as immature DCs
(iDCs), which are characterized by little or no expression of
co-stimulatory molecules such as CD40, CD80 and CD86, which are
important for the T lymphocyte activation process (11). In this stage of differentiation, these
immature cells have a reduced ability to activate the immune system
(11,12).
The recognition and processing of antigens results
in phenotypic and functional changes of iDCs, which leads to the
maturation of these cells. During the maturation process, iDCs lose
the molecules associated with the recognition of antigens and start
to present on their surface molecules of the major
histocompatibility complex, adhesion and co-stimulatory molecules,
including CD209, CD80 and CD86 (13).
Additionally, these cells become able to synthesize cytokines such
as IL-1, IL-12, IL-18 and IL-23, which are important in the T
lymphocyte activation process (13–15).
Thus, depending on the activating stimulus and the
extent of maturation, DCs can modulate the differentiation of T
helper (Th) lymphocytes (CD4+) in distinct
subpopulations (16,17). Th1 lymphocytes are characterized by
the synthesis of inflammatory cytokines such as IL-2 and interferon
(IFN)-γ, which activate the cellular immune response (18). The main function of IL-2 is the
self-regulation of the proliferation of Th1 lymphocytes (19). In addition, both IL-2 and IFN-γ
regulate the activation of other leucocytes, including cytotoxic T
lymphocytes (CD8+), natural killer (NK) cells and
macrophages, which are key cells involved in the elimination of
tumor cells (18,19).
By contrast, Th2 and regulatory T (Treg) cells are
associated with inhibition of the differentiation of
CD8+ T lymphocytes, NK cells and macrophages. This
occurs by inducing an immune condition that favors tumor growth via
the production of cytokines such as TGF-β by Treg cells and IL-4 by
Th2 lymphocytes (20,21).
Based on the ability of DCs to initiate an specific
and intense antitumor immune response through the induction of
T-cell clones, Th1, CD8+ and NK cells, the study of new
antitumor therapies has been supported by the use of this cell type
as a therapeutic tool (22). Previous
studies have focused on the development of immunotherapies with DCs
using new differentiation and maturation protocols for the
induction of an effective antitumor immune response (23,24).
Therefore, the purpose of the present study is to
aid the development of new therapeutic strategies based on the
differentiation and maturation of DCs in order to increase the
effectiveness of antitumor vaccines. However, since it is a
customized process, as it is performed with the patient's own
cells, the degree of maturation reached by DCs following the same
stimuli may be different from one patient to another. The present
study intends to verify if there is any influence of the degree of
malignancy on the maturation process and to evaluate the efficacy
of different protocols. Accordingly, it was determined whether the
type of cervical injury influences the differentiation of DCs in
vitro and whether the use of other protocols of differentiation
could result into mature DCs, which could be used as vaccines to
induce an antitumor response.
Materials and methods
Patients and controls
Eighty-three patients aged 36.5±11.5 years were
recruited from the Clinical Hospital of the Federal University of
Triângulo Mineiro (Uberaba, MG, Brazil) between 2013 and 2015.
Patients were selected through histopathological and cytological
diagnosis, and were grouped in LSIL (n=30), HSIL (n=22), cervical
cancer (n=10) and healthy patients (control, n=21) groups. The
number of cervical cancer patients was not sufficient for the
statistical analysis of the differentiation of DCs using protocols
II and III. Patients with immunosuppressive diseases or autoimmune
diseases, and those who were using immunosuppressive or antitumor
drugs or were pregnant, were excluded from the study.
All the patients and healthy controls involved in
the present study were counseled regarding its aims, and provided
written consent for participation. The study was approved by the
Ethics Committee of the Federal University of the Triângulo Mineiro
of Uberaba (Uberaba, MG, Brazil; approval no. 683-2006).
Generation of mature DCs and
alternatively activated DCs
Peripheral blood mononuclear cells (PBMCs) were
isolated by density gradient centrifugation of peripheral blood
using Ficoll-Paque™ PLUS solution (GE Healthcare Life Sciences,
Chalfont, UK) at 600 × g for 30 min at 18°C. These cells were
subjected to three different protocols of differentiation and
activation. Mature DCs [protocol I (pI), pIDCs] were generated by
culture of PBMCs (5×106 cells) in the presence of
granulocyte-macrophage colony-stimulating factor (GM-CSF) and IL-4
(0.5 ng/ml each; BD Pharmingen, San Diego, CA, USA) followed by the
addition on day 6 of tumor necrosis factor (TNF)-α (0.5 ng/ml; BD
Pharmingen) for 48 h. Alternatively, activated DCs were generated
by culture of PBMCs (5×106 cells) in the presence of
GM-CSF and IL-4 (0.5 ng/ml each; BD Pharmingen) for 5 days in the
absence [protocol II (pII), pIIDCs] or presence [protocol III
(pIII), pIIIDCs] of non-adherent cells to obtain iDCs. These cells
were matured by the addition of TNF-α (0.5 ng/ml; BD Pharmingen) on
day 6 and of supernatant derived from lymphocytes stimulated with
lipopolysaccharide (LPS) (10 mg/ml for 48 h; Sigma-Aldrich; Merck
Millipore, Darmstadt, Germany) on day 7 for 24 h. DCs were cultured
in Iscove's modified Dulbecco's medium (Sigma-Aldrich; Merck
Millipore), while lymphocytes were cultured in RPMI-1640 medium
(Sigma-Aldrich; Merck Millipore) supplemented with 40 mg/ml
gentamycin, 200 mM L-glutamine and 10% fetal bovine serum
(Sigma-Aldrich; Merck Millipore) at 37°C with 5%
CO2.
Flow cytometry
On day 8 of culture, mature DCs and alternatively
activated DCs were incubated with the corresponding antibodies to
examine the expression of maturation markers. Antibodies used for
flow cytometry included anti-CD11c-allophycocyanin, anti-human
leukocyte antigen-antigen D related (HLA-DR)-peridinin chlorophyll
protein, anti-CD86-phycoerythrin and isotypic controls (BD
Pharmigen). Data were acquired in the flow cytometer FACSCalibur™
(BD Biosciences, Franklin Lakes, NJ, USA).
ELISA
Supernatants were harvested from DCs culture on day
8 and stored at −80°C. The procedure was performed according to the
manufacturer's protocol. Cytokines, including IL-12, p40, IL-4 and
TGF-β, were measured by sandwich ELISA (BD OptEIA™; BD
Biosciences).
Statistical analysis
Statistical analysis was performed with GraphPad
Prism 5.00 (GraphPad Software, Inc., La Jolla, CA, USA). The data
for each variable were tested to assess whether they were normally
distributed. For comparisons of non-normally distributed data,
Mann-Whitney test was used for comparison of paired groups, and
Kruskal-Wallis and post hoc Dunn's tests were used for comparisons
among three or more groups. The results are expressed as the median
and range. P<0.05 was considered to indicate a statistically
significant difference.
Results
pI results in a semi-mature phenotype
of DCs from cervical cancer patients
The expression of co-stimulatory molecules and
surface markers (CD11c, CD86 and HLA-DR) was evaluated by flow
cytometry to examine the maturity status of DCs differentiated
according to pI from healthy subjects and cervical cancer patients.
Compared with normal pIDCs, cervical cancer pIDCs expressed lower
levels of the surface marker CD11c and the co-stimulatory molecule
CD86, and significantly lower levels of the antigen-presenting
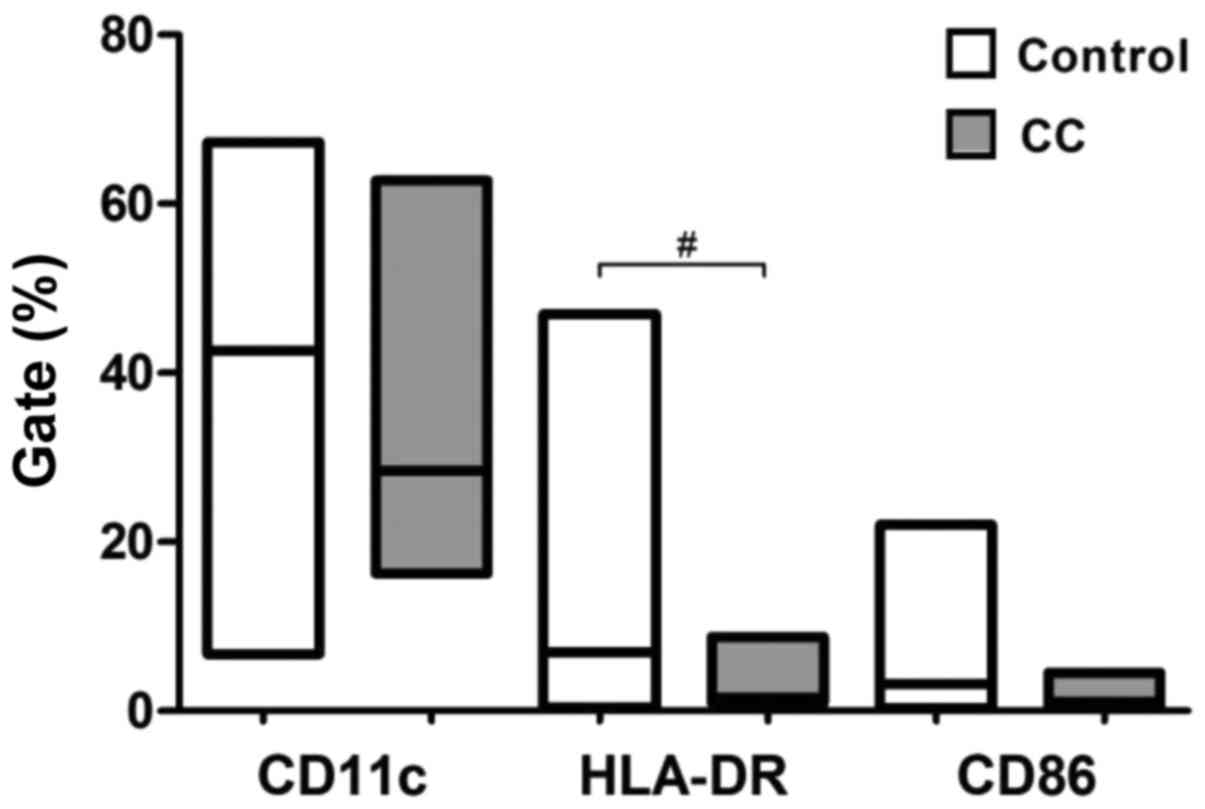
molecule HLA-DR (P=0.0113) (Fig. 1).
These results suggest that cervical cancer monocyte-derived DCs
matured according to pI exhibit a lower maturation phenotype. There
were no significant changes for the other groups using this
maturation protocol (Table I).
 | Table I.Comparison between the different
protocols used for the activation profile and cytokine production
of monocyte-derived dendritic cells derived from cervical
intraepithelial lesions patients. |
Table I.
Comparison between the different
protocols used for the activation profile and cytokine production
of monocyte-derived dendritic cells derived from cervical
intraepithelial lesions patients.
| A, Gate (%) |
|---|
|
|---|
|
| Protocol I | Protocol II | Protocol III |
|---|
|
|
|
|
|
|---|
| Cytokine | Control | LSIL | HSIL | CC | Control | LSIL | HSIL | Control | LSIL | HSIL |
|---|
| CD11c | 42.65 | 30.91 | 39.54 | 33.87 | 2.19a | 1.57a | 7.26b,c | 30.17 | 46.36b,c | 32.76 |
| HLA-DR | 6.96 | 5.75 | 5.65 | 1.59c | 0.70a | 0.62b | 1.76 | 6.60 | 6.95 | 4.23 |
| CD86 | 3.25 | 2.05 | 2.56 | 1.12 | 0.57b | 0.67b | 1.11 | 1.91 | 1.59 | 3.44 |
|
| B, Cytokine
production (pg/ml) |
|
|
| Protocol I | Protocol II | Protocol III |
|
|
|
|
|
| Cytokine | Control | LSIL | HSIL | CC | Control | LSIL | HSIL | Control | LSIL | HSIL |
|
| IL-4 | 26.68 | 34.97 | 29.95 | 31.30 | 330.60b | 47.31b | 39.16 | 19.43 | 13.02b | 11.34b |
| IL-12 | 272.80 | 271.50 | 277.20 | 244.60 | 42.77a | 105.00 | 84.21 | 544.00b |
1,059.00b | 641.00 |
| TGF-β | 260.20 | 334.50 | 526.50 | 479.30 | 688.30b | 721.90b | 626.10 |
1,602.00a |
4,525.00a |
3,254.00b |
pII induces a significantly lower
maturation profile of DCs
The expression of co-stimulatory molecules and
surface markers (CD11c, CD86 and HLA-DR) was evaluated by flow
cytometry to determine if the use of different protocols of
differentiation could result into DCs with increased maturation
profile. Compared with pIDCs and pIIIDCs, pIIDCs exhibited
significantly lower expression of CD11c (P<0.0001), CD86
(P=0.0006) and HLA-DR (P<0.0001) (Fig.
2A-C). Furthermore, these cells displayed a significant
reduction in IL-12 (P<0.0001) production and a significant
increase in IL-4 production (P<0.0001) (Fig. 3A and B). These findings suggest that,
in the absence of non-adherent cells, monocyte-derived DCs show a
reduction in their differentiation and maturation processes.
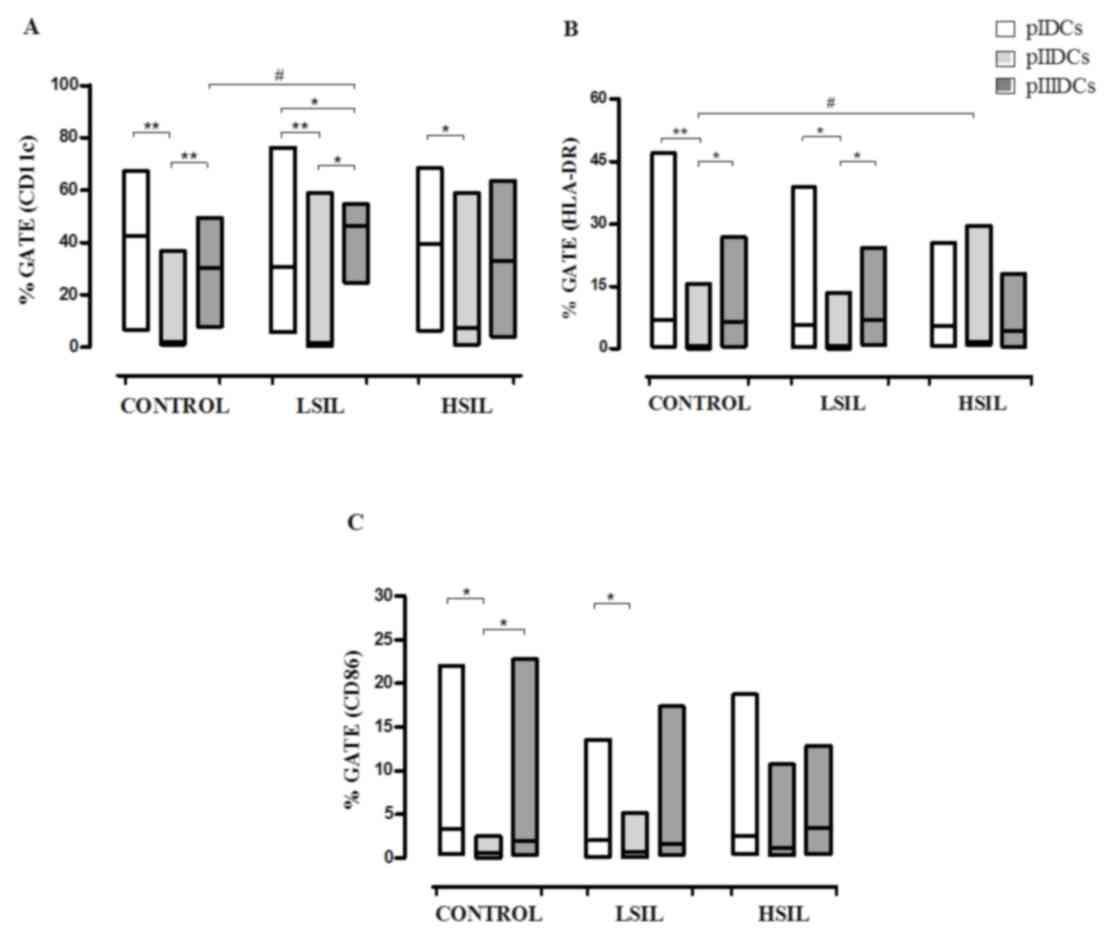 | Figure 2.Phenotypic profiles of
monocyte-derived pIDCs, pIIIDCs and pIIDCs. PBMCs were stimulated
with GM-CSF, IL-4 and TNF-α to generate pIDCs. The expression of
co-stimulatory molecules and surface markers (A) CD11c, (B) HLA-DR
and (C) CD86 and was evaluated by flow cytometry to evaluate the
effect of different protocols of differentiation on the maturation
profile. PBMCs were stimulated with GM-CSF, IL-4, TNF-α and
activated lymphocytes in the absence of non-adherent cells to
generate pIIDCs. PBMCs were stimulated with GM-CSF, IL-4, TNF-α and
activated lymphocytes in the presence of non-adherent cells to
generate pIIIDCs. Data are presented as the median and range
(Mann-Whitney and Kruskal-Wallis tests). #P<0.05
(comparison between patient groups); *P<0.05, **P<0.01
(comparison between protocols). Control, healthy patients. LSIL,
low-grade squamous intraepithelial lesion; HSIL, high-grade
squamous intraepithelial lesion; PBMC, peripheral blood mononuclear
cell; pI–III, protocol I–III; CD, cluster of differentiation; DC,
dendritic cell; HLA-DR, human leukocyte antigen-antigen D related;
GM-CSF, granulocyte-macrophage colony-stimulating factor; IL,
interleukin; TNF, tumor necrosis factor. |
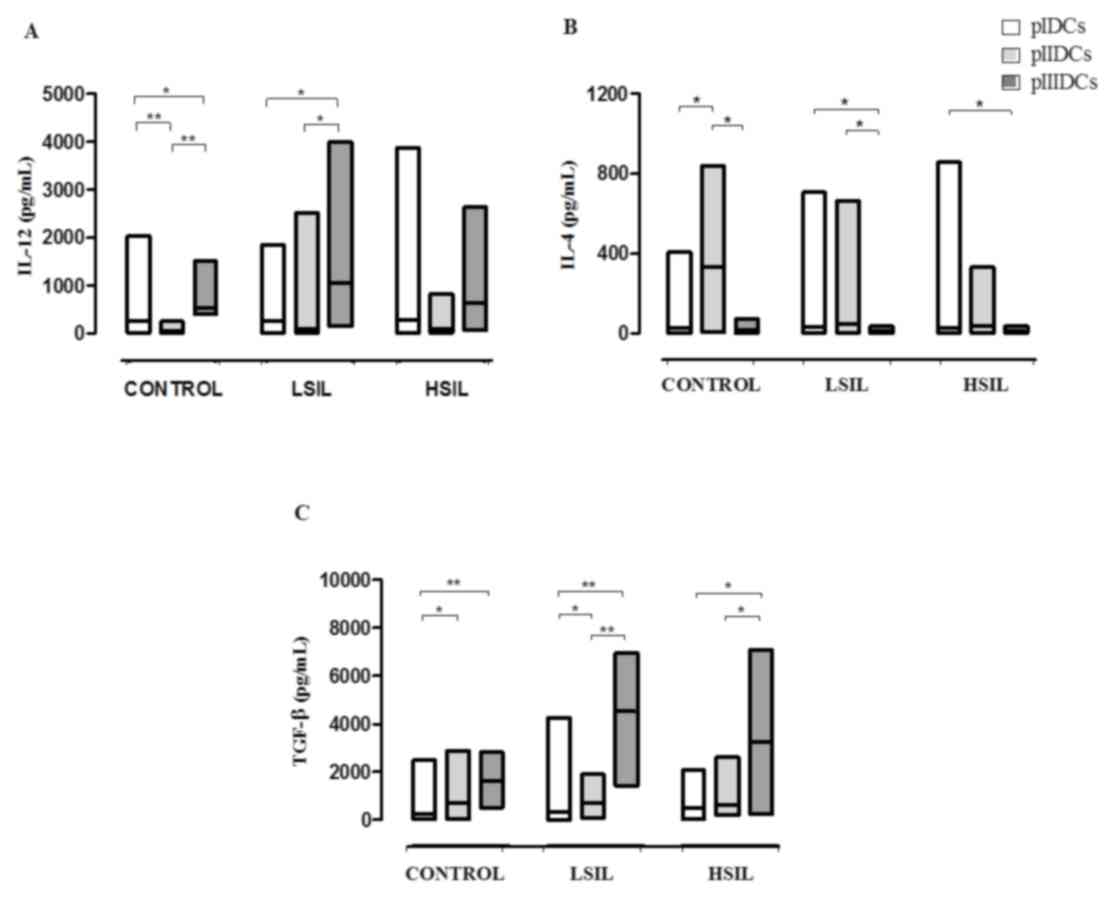 | Figure 3.Cytokine profile of monocyte-derived
pIDCs, pIIIDCs and pIIDCs. PBMCs were stimulated with GM-CSF, IL-4
and TNF-α to generate pIDCs. PBMCs were stimulated with GM-CSF,
IL-4, TNF-α and activated lymphocytes in the absence of
non-adherent cells to generate pIIDCs. PBMCs were stimulated with
GM-CSF, IL-4, TNF-α and activated lymphocytes in the presence of
non-adherent cells to generate pIIIDCs. LSIL pIIIDCs exhibited (A)
a significant increase in IL-12 production and (B) a significant
decrease in IL-4 production. (C) LSIL and HSIL pIDCs, pIIDCs and
pIIIDCs exhibited a significantly increased TGF-β production. Data
are presented as the median and range (Kruskal-Wallis test).
*P<0.05, **P<0.01. Control, healthy patients. LSIL, low-grade
squamous intraepithelial lesion; HSIL, high-grade squamous
intraepithelial lesion; PBMC, peripheral blood mononuclear cell;
pI–III, protocol I–III; DC, dendritic cell; TGF, transforming
growth factor; GM-CSF, granulocyte-macrophage colony-stimulating
factor; IL, interleukin; TNF, tumor necrosis factor. |
pII induces increased HLA-DR
expression in HSIL DCs
Compared with normal DCs, HSIL DCs differentiated
according to pII exhibited significantly increased expression of
HLA-DR (P=0.0113) (Fig. 2B). These
findings suggest that the presence of factors produced by
non-adherent cells can inhibit the expression of this molecule in
monocyte-derived DCs.
pIII induces high CD11c+
IL-12-producing LSIL DCs
Compared with control pIIIDCs (P=0.0058), LSIL pIDCs
(P=0.0411) and LSIL pIIDCs (P=0.0003), LSIL pIIIDCs expressed
significantly increased levels of the surface marker CD11c
(Fig. 2A). Furthermore, LSIL pIIIDCs
also show a significant increase in IL-12 production (P=0.0007) and
a significant decrease in IL-4 production (P=0.0038) (Fig. 3A and B and Table I). Together, these findings suggest
that, in the presence of non-adherent cells, monocyte-derived DCs
stimulated with activated lymphocytes may be more effective in
modulating the antitumor response during immunotherapy for patients
with low-grade cervical lesions.
pIIDCs and pIIIDCs show increased
TGF-β production
Compared with control, LSIL and HSIL pIDCs, pIIDCs
(P=0.0189) and pIIIDCs (P=0.0002) showed a significantly increased
TGF-β production (Fig. 3C). These
findings suggest that stimulation of non-adherent cells with LPS
may induce an increased production of this cytokine by Treg
cells.
Discussion
The identification of specific immune responses in
cancer patients stimulated the development of numerous
immunotherapies for tumors (25). In
1994, Sallusto and Lanzavecchia (26)
developed a technique capable of inducing the differentiation of
mature DCs from peripheral blood monocytes using a combination of
the cytokines IL-4, GM-CSF and TNF-α. Since then, several protocols
have been developed in order to improve the function of these
cells, since their ability to activate naive T lymphocytes and to
initiate a specific immune response depends on their state of
maturity (26–28).
Among the protocols used, it is worth citing
stimulation with TNF-α, IL-1β, bacterial products such as LPS, IFNs
and prostaglandins (29–31). Conventional maturation protocols use a
combination of the cytokines IL-1β, IL-6, TNF-α and prostaglandin
E2, which are capable of inducing the differentiation of mature DCs
with great co-stimulatory capacity and migration ability, but with
reduced capacity of production of IL-12 (32,33).
The present study verified the differentiation
profile of DCs from patients with cervical lesions, and assessed
whether different protocols of maturation could result in DCs in a
better state of maturity. Studies have shown that cancer patients
have alterations in the DCs differentiation and maturation
processes, which can be associated with the mechanism of escaping
immune surveillance (34). No studies
were identified in the literature correlating the influence of the
degree of malignancy of cervical intraepithelial lesions or the
influence of non-adherent cells with the maturation of DCs.
In the present study, a reduction in the maturity of
monocyte-derived DCs from cervical cancer patients was observed.
Gabrilovich et al (4) observed
that monocyte-derived DCs from patients with breast cancer
exhibited a reduction in antigen presentation, which is consistent
with the findings of our study.
When exposed to lymphocytes activated with LPS, a
significant increase in CD11c+ pIIIDCs was observed when
comparing the LSIL group with the control group. Our results also
indicate a significant reduction in CD11c+ pIIDCs
compared with pIIIDCs and pIDCs in the LSIL and control groups. In
addition, a significant increase in CD11c+ pIIIDCs
compared with pIDCs was observed. These results suggest that the
presence of non-adherent cells in the supernatant of
monocyte-derived DCs culture may assist in their
differentiation.
In 2003, Wolf et al demonstrated an increased
number of Treg lymphocytes in peripheral blood from patients with
different types of cancer (35).
These cells can synthesize TGF-β, a cytokine able to assist in
tumor development (36). Luttmann
et al reported that this cytokine is able to inhibit HLA-DR
expression in peripheral blood cells (37). The significant increase in
HLA-DR+ pIIDCs in the HSIL group compared with the
control group observed in the present study may be associated with
the absence of Treg lymphocytes in the culture of monocyte-derived
pIIDCs.
When evaluating HLA-DR expression across the
different protocols used, lower HLA-DR+ pIIDCs were
observed in all the groups of patients. This reduction was
statistically significant when comparing pIIDCs with pIDCs and
pIIIDCs in the LSIL and control groups, suggesting that DCs
maturation is also influenced by the presence of non-adherent cells
in the culture supernatant.
Orsini et al observed that colorectal cancer
patients had deficiency in monocyte-derived DCs differentiation,
with reduced expression of co-stimulatory molecules and reduced
IL-12 and TNF-α synthesis (38). In
our study, no significant changes in the expression of the
co-stimulatory molecule CD86 were observed, suggesting that the
degree of the cervical lesion was not able to interfere on the
expression of this molecule.
Previous studies have shown that lymphocytes of the
innate immune response, including NK, NKT and γδ T cells, are able
to induce DCs maturation, which may be evidenced by increased CD86
expression and IL-12 production (39–41). In
the present study, it was noticed a lower expression of CD86 in
pIIDCs compared with DCs derived from the other protocols. This
reduction was significant when comparing CD86+ pIIDCs
with CD86+ pIDCs and pIIIDCs in the LSIL and control
groups, which suggested that lymphocytes present in the culture
supernatant not only stimulate HLA-DR expression, but also induce
CD86 expression in monocyte-derived DCs.
Human monocyte-derived CD11c+ DCs induce
predominantly the differentiation of naive T lymphocytes in Th1
cells by IL-12 production, whereas CD11c− cells induce
prevalently Th2 responses (13,42).
Furthermore, IL-12 produced by DCs is capable of stimulating
cytotoxicity and production of IFN-γ by CD8+ T
lymphocytes and NK cells (43,44).
In our study, no statistically significant
differences were observed for the production of this cytokine among
the LSIL, HSIL, cervical cancer or control groups. By contrast,
when comparing the different protocols used for differentiation,
lower IL-12 production was noticed in pIIDCs compared with pIDCs
and pIIIDCs. This reduction in IL-12 synthesis may be due to the
absence of non-adherent cells in the supernatants of DCs culture,
since these cells appear to be directly associated with the
maturation of DCs and the synthesis of IL-12, as described above.
In addition, increased IL-12 production and a significant reduction
in IL-4 production by pIIIDCs was observed, suggesting that it
could be more effective in inducing antitumor responses during
immunotherapy with DCs since IL-12 cytokine induces the
differentiation of Th1 lymphocytes, which is important in the
antitumor response.
In normal cells, TGF-β acts as a tumor suppressor
because it can inhibit cell proliferation and promote cell
differentiation and apoptosis (36).
However, in the early stages of tumorigenesis, cells lose the
inhibition of growth mediated by TGF-β as a result of mutation or
loss of expression of genes associated with the TGF-β signaling
pathway (36). Once they have become
resistant against growth inhibition by TGF-β, tumor cells and
stromal cells present in the tumor increase the production of this
cytokine, which stimulates angiogenesis, cell motility and
suppression of the immune system, thus promoting the invasion and
metastasis of tumor cells (36).
pIIIDCs have shown increased TGF-β production in
patients with SILs in the present study. When comparing the
different protocols used, it was observed that there was an
increase in TGF-β production in pIIDCs and pIIIDCs compared with
pIDCs for all the groups of patients evaluated. This increase was
significant when comparing pIIIDCs with pIDCs in the control group,
and when comparing pIIIDCs with pIDCs and pIIDCs in patients with
LSIL and HSIL. This may be associated with the presence of Treg
lymphocytes in the culture supernatant, which, according to
Caramalho et al, can be activated by stimulation with LPS
(45).
From these results, it can be concluded that the
cell-to-cell contact of activated lymphocytes is able to induce a
better differentiation of monocyte-derived DCs. Furthermore, it was
also observed that activated non-adherent cells were capable of
inducing increased CD86 and HLA-DR expression, and probably capable
of inducing Th1 responses as well, which are notably important for
the antitumor response. This could be observed by the increase in
IL-12 production characteristic of this profile and by the
reduction in IL-4 synthesis, which is a cytokine produced by Th2
cells. The presence of non-adherent cells in the culture of DCs
appears to contribute to their differentiation process, since their
removal induced a significant reduction in the expression of the
surface markers CD11c, CD86 and HLA-DR, and induced a significant
reduction in IL-12 synthesis. In addition, it was also observed
that the extent of the cervical lesion can influence the
differentiation process of DCs, since a significant reduction in
HLA-DR+ DCs differentiated according to pI was obtained
from patients with cervical cancer. Therefore, future studies are
required to understand the effects of non-adherent cells on the
differentiation and maturation of DCs.
Acknowledgements
The authors acknowledge the funding received from
the Brazilian National Council for Scientific and Technological
Development (grant no. 302011/2015-3), the Uberaba Foundation for
Teaching and Research (grant no. 255/2012) and the Foundation for
Research Assistance of the State of Minas Gerais (grant no. Rede
11/14).
References
|
1
|
Stewart BW and Wild CP: World Cancer
Report 2014. World Health Organization, IARC; Lyon: 2016
|
|
2
|
Apgar BS, Zoschnick L and Wright TC Jr:
The 2001 bethesda system terminology. Am Fam Phisician.
68:1992–1998. 2003.
|
|
3
|
Parkin DM and Bray F: Chapter 2: The
burden of HPV-related cancers. Vaccine. 24:(Suppl 3). 11–25. 2006.
View Article : Google Scholar
|
|
4
|
Gabrilovich DI, Corak J, Ciernik IF,
Kavanaugh D and Carbone DP: Decreased antigen presentation by
dendritic cells in patients with breast cancer. Clin Cancer Res.
3:483–490. 1997.PubMed/NCBI
|
|
5
|
Vopenkova K, Mollova K, Buresova I and
Michalek J: Complex evaluation of human monocyte-derived dendritic
cells for cancer immunotherapy. J Cell Mol Med. 16:2827–2837. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ferrantini M, Capone I and Belardelli F:
Dendritic cells and cytokines in immune rejection of cancer.
Cytokine Growth Factor Rev. 19:93–107. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Banchereau J and Steinman RM: Dendritic
cells and the control of immunity. Nature. 392:245–252. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Steinman RM and Banchereau J: Taking
dendritic cells into medicine. Nature. 449:419–426. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
van Nierop K and de Groot C: Human
follicular dendritic cells: Function, origin and development. Semin
Immunol. 14:251–257. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pulendran B, Smith JL, Caspary G, Brasel
K, Pettit D, Maraskovsky E and Maliszewski CR: Distinct dendritic
cell subsets differentially regulate the class of immune response
in vivo. Proc Natl Acad Sci USA. 96:1036–1041. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kelleher P and Knight SC: IL-12 increases
CD80 expression and the stimulatory capacity of bone marrow-derived
dendritic cells. Int Immunol. 10:749–755. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Schwartz RH: A cell culture model for T
lymphocyte clonal anergy. Science. 248:1349–1356. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Banchereau J, Briere F, Caux C, Davoust J,
Lebecque S, Liu Y, Pulendran B and Palucka K: Immunobiology of
dendritic cells. Annu Rev Immunol. 18:767–811. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Randolph GJ, Sanchez-Schmitz G and Angeli
V: Factors and signals that govern the migration of dendritic cells
via lymphatics: Recent advances. Springer Semin Immunopathol.
26:273–287. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Palucka K and Banchereau J: Cancer
immunotherapy via dendritic cells. Nat Rev Cancer. 12:265–277.
2012. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu YJ: Dendritic cell subsets and
lineages, and their functions in innate and adaptive immunity.
Cell. 106:259–262. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bradley LM: Migration and T-lymphocyte
effector function. Curr Opin Immunol. 15:343–348. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Corthay A, Skovseth DK, Lundin KU, Røsjø
E, Omholt H, Hofgaard PO, Haraldsen G and Bogen B: Primary
antitumor immune response mediated by CD4+ T cells. Immunity.
22:371–383. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Pardoll DM and Topaliant SL: The role of
CD4+ T cell responses in antitumor immunity. Courrent Opin Immunol.
10:588–594. 1998. View Article : Google Scholar
|
|
20
|
Roncarolo MG, Bacchetta R, Bordignon C,
Narula S and Levings MK: Type 1 T regulatory cells. Immunol Rev.
182:68–79. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Fukaura H, Kent SC, Pietrusewicz MJ,
Khoury SJ, Weiner HL and Hafler DA: Induction of circulating myelin
basic protein and proteolipid protein-specific transforming growth
factor- beta 1-secreting Th3 T cells by oral administration of
myelin in multiple sclerosis patients. J Clin Invest. 98:70–77.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Koido S, Homma S, Takahara A, Namiki Y,
Tsukinaga S, Mitobe J, Odahara S, Yukawa T, Matsudaira H, Nagatsuma
K, et al: Current immunotherapeutic approaches in pancreatic
cancer. Clin Dev Immunol. 2011:2675392011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Rodrigues CM, Matias BF, Murta EF and
Michelin MA: The role of T lymphocytes in cancer patients
undergoing immunotherapy with autologous dendritic cells. Clin Med
Insights Oncol. 5:107–115. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Matias BF, de Oliveira TM, Rodrigues CM,
Abdalla DR, Montes L, Murta EF and Michelin MA: Influence of
immunotherapy with autologous dendritic cells on innate and
adaptive immune response in cancer. Clin Med Insights Oncol.
7:165–172. 2013.PubMed/NCBI
|
|
25
|
Baxevanis CN, Perez SA and Papamichail M:
Combinatorial treatments including vaccines, chemotherapy and
monoclonal antibodies for cancer therapy. Cancer Immunol
Immunother. 58:317–324. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sallusto BF and Lanzavecchia A: Efficient
presentation of solube antigen by cultured human dendritic cells is
maintained by granulocyte/macrophage colony-stimulating factor plus
interleukin 4 and downregulated by tumor necrosis factor alpha. J
Exp Med. 179:1109–1118. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Apostolopoulos V, Pietersz GA, Tsibanis A,
Tsikkinis A, Stojanovska L, McKenzie IF and Vassilaros S: Dendritic
cell immunotherapy: Clinical outcomes. Clin Transl Immunol.
3:e212014. View Article : Google Scholar
|
|
28
|
Figdor CG, de Vries IJ, Lesterhuis WJ and
Melief CJ: Dendritic cell immunotherapy: Mapping the way. Nat Med.
10:475–480. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Han TH, Jin P, Ren J, Slezak S, Marincola
FM and Stroncek DF: Evaluation of 3 clinical dendritic cell
maturation protocols containing lipopolysaccharide and
interferon-gamma. J Immunother. 32:399–407. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
ten Brinke A, Karsten ML, Dieker MC,
Zwaginga JJ and van Ham SM: The clinical grade maturation cocktail
monophosphoryl lipid A plus IFNgamma generates monocyte-derived
dendritic cells with the capacity to migrate and induce Th1
polarization. Vaccine. 25:7145–7152. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Snijders A, Kalinski P, Hilkens CM and
Kapsenberg ML: High-level IL-12 production by human dendritic cells
requires two signals. Int Immunol. 10:1593–1598. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Jonuleit H, Kühn U, Müller G, Steinbrink
K, Paragnik L, Schmitt E, Knop J and Enk AH: Pro-inflammatory
cytokines and prostaglandins induce maturation of potent
immunostimulatory dendritic cells under fetal calf serum-free
conditions. Eur J Immunol. 27:3135–3142. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Nicolette CA, Healey D, Tcherepanova I,
Whelton P, Monesmith T, Coombs L, Finke LH, Whiteside T and
Miesowicz F: Dendritic cells for active immunotherapy: Optimizing
design and manufacture in order to develop commercially and
clinically viable products. Vaccine. 25:(Suppl 2). B47–B60. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Pinzon-Charry A, Maxwell T and López JA:
Dendritic cell dysfunction in cancer: A mechanism for
immunosuppression. Immunol Cell Biol. 83:451–461. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wolf AM, Wolf D, Steurer M, Gastl G,
Gunsilius E and Grubeck-loebenstein B: Increase of regulatory T
Cells in the peripheral blood of cancer patients. Clin Cancer Res.
9:606–612. 2003.PubMed/NCBI
|
|
36
|
Blobe GC, Schiemann WP and Lodish HF: Role
of transforming growth factor beta in human disease. N Engl J Med.
342:1350–1308. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Luttmann W, Franz P, Schmidt S, Barth J,
Matthys H and Virchow JC Jr: Inhibition of HLA-DR expression on
activated human blood eosinophils by transforming growth
factor-beta1. Scand J Immunol. 48:667–671. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Orsini G, Legitimo A, Failli A, Ferrari P,
Nicolini A, Spisni R, Miccoli P and Consolini R: Defective
generation and maturation of dendritic cells from monocytes in
colorectal cancer patients during the course of disease. Int J Mol
Sci. 14:22022–22041. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Mocikat R, Braumüller H, Gumy A, Egeter O,
Ziegler H, Reusch U, Bubeck A, Louis J, Mailhammer R, Riethmüller
G, et al: Natural killer cells activated by MHC class I(low)
targets prime dendritic cells to induce protective CD8 T cell
responses. Immunity. 19:561–569. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Hermans IF, Silk JD, Gileadi U, Salio M,
Mathew B, Ritter G, Schmidt R, Harris AL, Old L and Cerundolo V:
NKT cells enhance CD4+ and CD8+ T cell responses to soluble antigen
in vivo through direct interaction with dendritic cells. J Immunol.
171:5140–5147. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Leslie DS, Vincent MS, Spada FM, Das H,
Sugita M, Morita CT and Brenner MB: CD1-mediated gamma/delta T cell
maturation of dendritic cells. J Exp Med. 196:1575–1584. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Maldonado-López R, De Smedt T, Michel P,
Godfroid J, Pajak B, Heirman C, Thielemans K, Leo O, Urbain J and
Moser M: CD8alpha+ and CD8alpha- subclasses of dendritic cells
direct the development of distinct T helper cells in vivo. J Exp
Med. 189:587–592. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Dorman SE and Holland SM:
Interferon-gamama and interleukin-12 pathway defects and human
disease. Cytokine Growth Factor Rev. 11:321–333. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Heiser A, Coleman D, Dannull J, Yancey D,
Maurice MA, Lallas CD, Dahm P, Niedzwiecki D, Gilboa E and Vieweg
J: Autologous dendritic cells transfected with prostate-specific
antigen RNA stimulate CTL responses against metastatic prostate
tumors. J Clin Invest. 109:409–417. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Caramalho I, Lopes-Carvalho T, Ostler D,
Zelenay S, Haury M and Demengeot J: Regulatory T cells selectively
express toll-like receptors and are activated by
lipopolysaccharide. J Exp Med. 197:403–411. 2003. View Article : Google Scholar : PubMed/NCBI
|