Introduction
The incidence of gastric cancer (GC) ranks as the
fifth most frequent among all types of cancer worldwide (1). Nearly 40% of all GC cases occur in
China, and are often diagnosed in advanced stages (2). The median overall survival (OS) for GC
patients remains <12 months with first-line oxaliplatin,
5-fluorouracil (5-FU) and folinic acid treatment (3). Of all GC patients, ~1/2 could be
candidates for second-line treatment at the time of failure of
first-line chemotherapy (4).
Docetaxel is among the most frequently used agents for GC
second-line treatment (5). In a
previous study by the present authors, the median OS was 25.8
months for patients with high messenger RNA (mRNA) expression
levels of breast cancer susceptibility gene 1 (BRCA1) treated with
second-line docetaxel-based chemotherapy (6). Recent evidence also suggests that an
underlying cause of drug resistance may be the failure of
drug-induced apoptosis (7–9). Platinum treatment initiates apoptosis
through the formation of DNA adducts, which primarily form
intrastrand crosslinks that activate the apoptotic pathway,
eventually resulting in cell death (10,11). The
most recognized mechanism of docetaxel-based regimen is the binding
to microtubules, which arrests the cell cycle in G2/M and
eventually leads to cell death (12).
B-cell lymphoma 2 (BCL-2) interacting mediator of
cell death (BIM) belongs to the BCL-2 protein family, and is also a
member of the BCL-2-homology 3-only (BH3-only) family (13). BIM is expressed in a wide variety of
tissues, including GC, and acts as a pivotal regulator of the
mitochondrial apoptosis pathway (14). Abnormal levels of BIM have been
recognized to affect the chemotherapy response (15). Platinum-resistant cancer cells
conserved sensitivity to BH3-induced mitochondrial apoptosis
(16). In line with that, BH3-mimetic
compounds such as ABT-737 were able to sensitize cancer cells to
platinum (17). In addition,
overexpression of BIM enhanced the in vitro sensitivity to
docetaxel of non-small cell lung cancer (NSCLC) (18). Consistent with this finding,
downregulation of BIM by small interfering RNA (siRNA) delayed
paclitaxel-mediated apoptosis, indicating that low BIM expression
levels were responsible for resistance to paclitaxel (19). Notably, pre-treatment mRNA levels of
BIM strongly predicted the capacity of epidermal growth factor
receptor (EGFR), human EGFR 2 (HER2) and phosphoinositide 3-kinase
(PI3K) inhibitors to induce apoptosis in EGFR-mutant,
HER2-amplified and phosphatidylinositol-4,5-bisphosphate 3-kinase
catalytic subunit alpha-mutant tumors, respectively (20). In a previous study, the present
authors observed that patients with high BIM expression achieved
longer survival in EGFR-mutant NSCLC treated with erlotinib or
chemotherapy (21).
Astrocyte elevated gene-1 (AEG-1) was originally
identified as a novel gene induced by human fetal astrocytes
following infection with human immunodeficiency virus 1 (22). AEG-1 does not impact the uptake or
retention of chemotherapy drugs; instead, AEG-1 increases
chemoresistance by enhancing cell survival (23). Overexpression of AEG-1 suppresses
apoptosis through phosphorylation of substrates of the
anti-apoptotic protein kinase B (also known as AKT) (24), and is important in promoting cancer
malignant behavior (25). In previous
studies, AEG-1 overexpression correlated with poor prognosis in GC
(25) and NSCLC (26). It has been confirmed that AEG-1
contributed to resistance to chemotherapeutic drugs such as 5-FU in
hepatocellular carcinoma cell lines (27). Furthermore, knockdown of AEG-1
sensitized breast cancer cell lines to paclitaxel in vitro
and in vivo (23). Low AEG-1
expression was associated with longer progression-free survival in
platinum-based chemotherapy in NSCLC (28). In addition, AEG-1 mRNA expression
correlated with BRCA1 expression (28), which induced sensitivity to docetaxel
(6).
AXL receptor tyrosine kinase (AXL) belongs to the
Tyro3, AXL and Mer family (29).
Growth arrest-specific gene 6 (Gas6) is the ligand of AXL (30). In conjunction with each other,
Gas6/AXL signaling may enhance cell survival (31). Activation of Gas6/AXL signaling
induced the activation of the PI3K signaling pathway, which
increased the expression of anti-apoptotic proteins such as BCL-2
and BCL-extra large (BCL-XL) (32).
Overexpression of AXL was responsible for tumor growth in
mesothelioma (33), lung cancer
(34) and breast cancer (35). Furthermore, increased AXL activation
has been linked with cisplatin resistance in ovarian cancer
(36).
In the present study, the mRNA expression levels of
BIM, AEG-1 and AXL were examined in 131 advanced GC samples. In
addition, the expression levels of the above genes were correlated
with patients' clinicopathological features and OS to first-line
FOLFOX combination chemotherapy with folinic acid and 5-FU, with or
without second-line docetaxel-based chemotherapy.
Patients and methods
Study population
A total of 131 advanced GC samples in which BRCA1
mRNA expression levels had been previously determined (6) were included in the present study.
Patients' clinical characteristics are indicated in Table I. All patients received a combination
of oxaliplatin, 5-fluorouracil (FU) and folinic acid (FOLFOX) as
first-line therapy (85 mg/m2 oxaliplatin plus 200
mg/m2 folinic acid and 600 mg/m2 5-FU every
for 2 weeks until disease progression) for a median of 3 cycles
(range, 1–8 cycles). A total of 34 patients received single-agent
docetaxel (35 mg/m2), and the remaining 22 patients were
treated with docetaxel-based doublets (6 patients received 35
mg/m2 docetaxel plus 100 mg/m2 irinotecan
weekly for 3 weeks, every 4 weeks until disease progression; 11
patients received 35 mg/m2 docetaxel weekly for 3 weeks
plus 1,000 mg/m2 capecitabine daily for 2 weeks, every 4
weeks until disease progression; and 5 patients received 35
mg/m2 docetaxel weekly for 3 weeks plus 6
mg/m2 hydroxycamptothecin on days 1 and 5, every 4 weeks
until disease progression) for a median of 3 cycles (range, 1–7
cycles). Following progression, 56 patients further received
docetaxel-based second-line chemotherapy. A total of 34 patients
received single-agent docetaxel, and the remaining 22 patients were
treated with docetaxel-based doublets, based on their response to
first-line chemotherapy, Eastern Cooperative Oncology Group (ECOG)
performance status (PS) and patient consent. Survival was
calculated from the starting date of first-line treatment to the
date of last follow-up or mortality from any cause. Approval was
obtained from the patients and from the ethics committee of Drum
Tower Hospital (Nanjing, China).
 | Table I.Patient characteristics. |
Table I.
Patient characteristics.
|
Characteristics | All patients N
(%) | Patients receiving
only first-line chemotherapy N (%) | Patients receiving
second-line chemotherapy N (%) | P-value |
|---|
| Total, N (%) | 131 (100.0) | 75 (100.0) | 56 (100.0) |
|
| Age, years |
|
|
| 0.330 |
|
<60 | 63 (48.1) | 32 (42.7) | 31 (55.4) |
|
|
≥60 | 68 (51.9) | 43 (57.3) | 25 (44.6) |
|
| Gender |
|
|
| 0.410 |
|
Female | 31 (23.7) | 20 (26.7) | 11 (19.6) |
|
|
Male | 100 (76.3) | 55 (73.3) | 45 (80.4) |
| Tumor site |
|
|
| 0.270 |
| Distal
stomach | 50 (38.2) | 25 (33.3) | 25 (44.6) |
|
|
Proximal stomach | 38 (29.0) | 24 (32.0) | 14 (25.0) |
|
| Whole
stomach | 42 (32.1) | 25 (33.3) | 17 (30.4) |
|
|
Unknown | 1 (0.8) | 1 (1.4) | 0 (0.0) |
|
| Stage |
|
|
| 0.550 |
|
III | 79 (60.3) | 44 (58.7) | 35 (62.5) |
|
| IV | 52 (39.7) | 31 (41.3) | 21 (37.5) |
|
| ECOG PS |
|
|
| 0.390 |
|
0–1 | 119 (90.8) | 66 (88.0) | 53 (94.6) |
|
| 2 | 12 (9.2) | 9 (12.0) | 3 (5.4) |
|
| Histological
grade |
|
|
| 0.070 |
| G2 | 35 (26.7) | 20 (26.7) | 15 (26.8) |
|
|
G2-3 | 35 (26.7) | 17 (22.7) | 18 (32.1) |
|
| G3 | 59 (45.0) | 37 (49.3) | 22 (39.3) |
|
|
Uknown | 2 (1.6) | 1 (1.3) | 1 (1.3) |
|
Gene expression analysis
Gene expression profiling was performed on RNA
isolated from macrodissected tumor tissues containing ≥80% of tumor
cells, in accordance with a proprietary procedure (European patent
publication no. EP1945764-B1). Primers and probes for gene
expression analysis of BIM, AEG-1 and AXL are indicated in Table II. The mRNA levels of BIM, AEG-1 and
AXL were measured by reverse transcription-quantitative polymerase
chain reaction (RT-qPCR) using Taqman® Universal PCR
Master Mix (Applied Biosystems; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA), according to the comparative Cq method (37). β-actin was used as an endogenous
control, and commercial RNA controls (Stratagene; Agilent
Technologies, Inc., Santa Clara, CA, USA) were used as calibrators.
RT-qPCR was conducted in a 7900HT Fast Real-Time PCR System
(Applied Biosystems; Thermo Fisher Scientific, Inc.). The reactions
were initiated by heating to 50°C for 2 min and then to 95°C for 2
min, followed by 40 cycles of 95°C for 15 sec and 60°C for 60
sec.
| Table II.Sequences of primers and probes. |
Table II.
Sequences of primers and probes.
| Gene | Primers | Probes |
|---|
| β-actin | F
5′-TGAGCGCGGCTACAGCTT-3′ | 6-FAM
5′-ACCACCACGGCCGAGCGG-3′ TAMRA |
|
| R
5′-TCCTTAATGTCACGCACGATTT-3′ |
|
| AEG-1 | F
5′-GGGGAAGGAGTTGGAGTGAC-3′ | 6-FAM
5′-AATATTTTCTGGCATTGGGTCTA-3′ MGB |
|
| R
5′-GTAGACTGAGAAACTGGCTCAGCAG-3′ |
|
| AXLb | F
5′-CAGCGCAGCCTGCATGT-3′ | 6-FAM
5′-CAGGGCTGAACAAGAC-3′ MGB |
|
| R
5′-GCGTTATGGGCTTCGCAG-3′ |
|
| BIM | Assay-on-Demand BIM
(ID#Hs00708019_s1)a |
|
Statistical analysis
Gene expression levels were analyzed as categorical
variables by terciles. Correlations between gene expression and
clinicopathological parameters were analyzed with the χ2
test. Correlations among different genes were conducted using
Spearman's correlation coefficient analysis. Stratified log-rank
tests were used to assess the median OS. Estimation of survival
curves was performed with the Kaplan-Meier method. A multivariate
analysis was performed using the Cox proportional hazards
regression model. All analyses were performed with SPSS version
17.0 software (SPSS, Inc.,Chicago, IL, USA). Two-sided P<0.05
was considered to indicate a statistically significant
difference.
Results
Distribution and clinicopathological
features of all patients
A total of 131 advanced GC samples were included in
the study, of which, 100 were males and 31 females. The median age
of all patients enrolled was 59.6 years (range, 22–84 years). All
patients were pathologically confirmed as adenocarcinoma, of which,
79 patients (60.3%) were confirmed with stage III and 52 patients
(39.7%) with stage IV disease (Table
I).
Correlations among different
genes
A correlation was observed between BIM and AEG-1
mRNA expression (rs=0.30; P=0.002). However, no
associations were observed between BIM, AXL and AEG-1 mRNA
expression (P=0.100 and 0.140 respectively).
Association between OS and
clinicopathological characteristics
The median OS was 11.6 months (95% CI, 9.8–13.6) in
all patients. Among the patients with stage III disease, the median
OS was 12.9 months, compared with 9.6 months among the patients
with stage IV disease (P=0.001). Notably, the median OS was ~12.5
months for patients with ECOG PS=0–1 vs. 6.3 months for patients
with ECOG PS=2 (P<0.001). There was no significant association
between OS and age (P=0.300), gender (P=0.630), tumor site
(P=0.110) or histological grade (P=0.070) (Table III).
| Table III.Association between gene expression
and clinicopathological characteristics in all patients. |
Table III.
Association between gene expression
and clinicopathological characteristics in all patients.
|
Characteristics | Patients, N
(%) | Median overall
survival, months (95% confidence interval) | P-value |
|---|
| Age, years |
|
| 0.300 |
|
<60 | 63 (48.1) | 12.5
(7.8–17.1) |
|
|
≥60 | 68 (51.9) | 10.9
(8.7–13.0) |
|
| Gender |
|
| 0.630 |
|
Female | 31 (23.7) | 11.7
(7.2–16.2) |
|
|
Male | 100 (76.3) | 11.3
(8.9–13.6) |
|
| Tumor site |
|
| 0.110 |
| Distal
stomach | 50 (38.2) | 11.3
(8.9–13.6) |
|
|
Proximal stomach | 38 (29.1) | 15.9
(11.0–20.9) |
|
| Whole
stomach | 42 (32.1) | 9.9 (9.0–10.7) |
|
|
Unknown | 1 (0.8) | – |
|
| Stage |
|
| 0.001 |
|
III | 79 (60.3) | 12.9
(9.1–16.7) |
|
| IV | 52 (39.7) | 9.6 (7.5–11.6) |
|
| ECOG PS |
|
| <0.001 |
|
0–1 | 119 (90.8) | 12.5
(9.9–15.1) |
|
| 2 | 12 (9.2) | 6.3 (0.7–11.9) |
|
| Histological
grade |
|
| 0.070 |
| G2 | 35 (26.7) | 12.6
(2.2–23.0) |
|
|
G2-3 | 35 (26.7) | 12.4
(7.2–17.6) |
|
| G3 | 59 (45.0) | 10.6
(9.1–11.1) |
|
|
Unknown | 2 (1.6) | – |
|
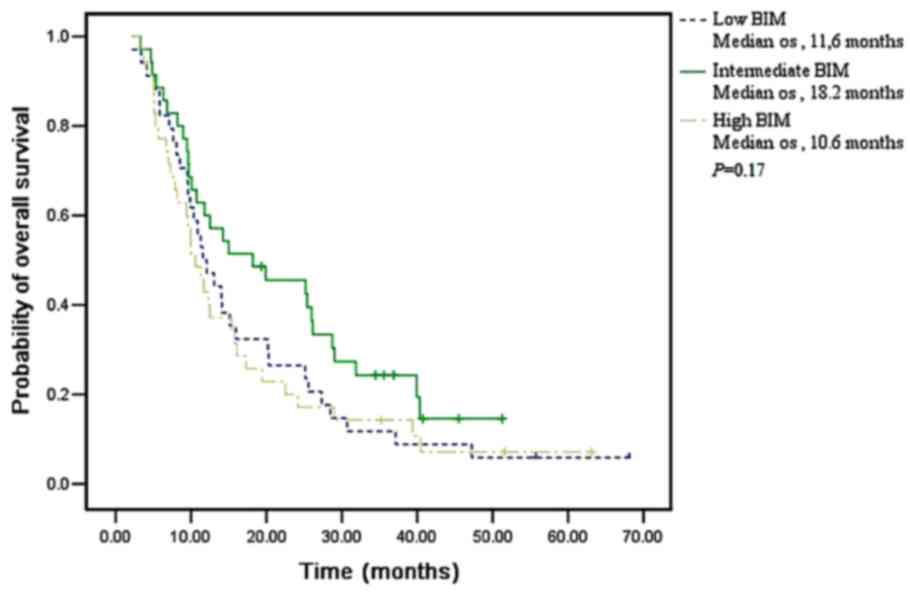
| BIM mRNA
levels |
|
| 0.170 |
|
Low | 34 (25.9) | 11.6
(8.4–14.7) |
|
|
Intermediate | 35 (26.7) | 18.2
(3.9–32.5) |
|
|
High | 35 (26.7) | 10.6
(8.1–13.0) |
|
| Not
detected | 27 (20.7) | – |
|
| AEG-1 mRNA
levels |
|
| 0.360 |
|
Low | 42 (32.1) | 10.9
(8.9–12.8) |
|
|
Intermediate | 40 (30.5) | 10.0
(6.9–13.1) |
|
|
High | 42 (32.1) | 12.5 (9.4
−15.7) |
|
| Not
detected | 7 (5.3) | – |
|
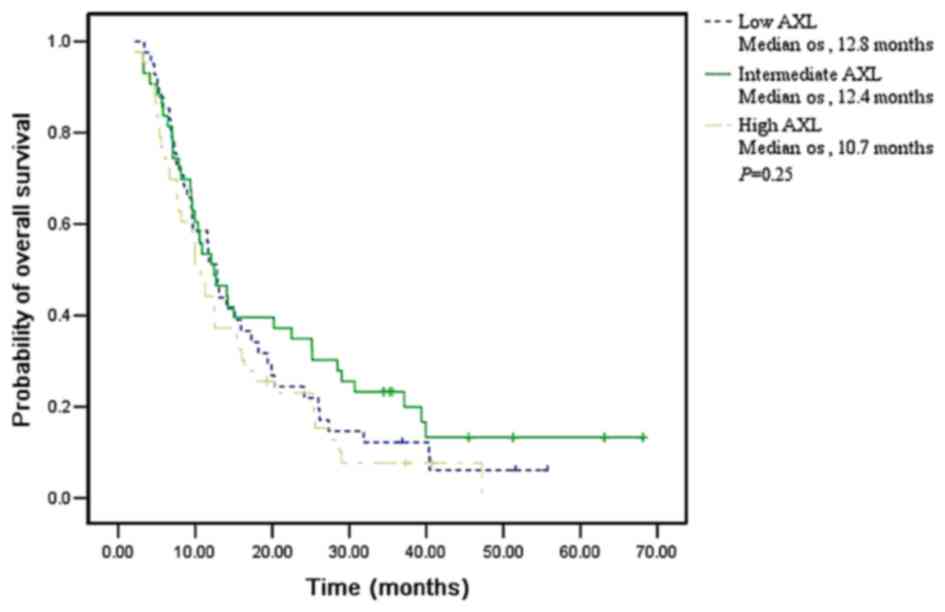
| AXL mRNA
levelsa |
|
| 0.250 |
|
Low | 41 (31.3) | 12.8 (10.9
−14.7) |
|
|
Intermediate | 43 (32.8) | 12.4
(7.9–17.0) |
|
|
High | 43 (32.8) | 10.7
(8.9–12.5) |
|
| Not
detected | 4 (3.1) | – |
|
| Second-line
chemotherapy |
|
| 0.060 |
|
Yes | 56 (42.7) | 15.0
(8.3–11.9) |
|
| No | 75 (57.3) | 10.1
(13.0–17.0) |
|
Survival for GC patients according to
mRNA expression levels
No association was observed between OS and the mRNA
expression levels of BIM (P=0.170), AEG-1 (P=0.360) and AXL
(P=0.250) in all 131 patients, respectively (Figs. 1–3).
Among the 75 patients receiving only first-line
FOLFOX chemotherapy, a trend towards longer survival was observed
in those with low BIM levels (P=0.080). However, there was no
difference in survival according to their AEG-1 (P=0.810) or AXL
mRNA expression levels (P=0.350).
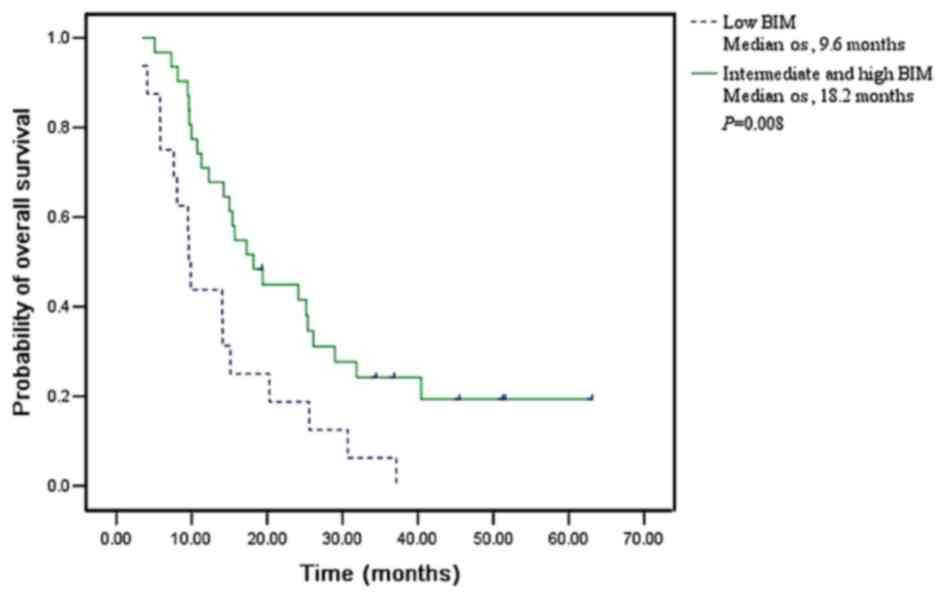
Among the 56 patients receiving additional
docetaxel-based second-line chemotherapy, the median OS was 9.6
months (95% CI, 8.9–10.3) for patients with low levels of BIM, 25.2
months (95% CI, 12.5–37.9) for those with intermediate BIM levels
and 15.7 months (95% CI, 9.4–22.0) for those with high BIM levels
(P=0.021). Considering the obvious trend of a longer OS in patients
with higher BIM expression, high and intermediate expression groups
were merged into a whole group for further analysis (Fig. 4). Patients with high or intermediate
levels of BIM exhibited a median OS of 18.2 months (95% CI,
12.8–23.6), while patients with low BIM exhibited a median OS of
just 9.6 months (95% CI, 8.9–10.3; P=0.008). Longer survival was
also observed in patients with high levels of AEG-1 (P=0.080),
although the difference was not significant. There was no
difference in OS according to the expression levels of AXL
(P=0.600).
To further understand the role of BIM as a
predictive biomarker, multivariate analysis of OS was performed.
Patients with low BIM mRNA levels had higher mortality than those
with high or intermediate BIM mRNA levels (HR of mortality, 2.61;
95% CI, 1.21–5.62; P=0.010). Lower risk of mortality was observed
in patients with ECOG PS=0–1 compared with those with ECOG PS=2
(HR, 0.17; 95% CI, 0.04–0.65; P=0.010) in patients with stage III
tumors, compared with patients with stage IV tumors (HR, 0.37; 95%
CI, 0.17–0.82; P=0.010) (Table
IV).
| Table IV.Median OS and HRs for risk of
mortality in all patients. |
Table IV.
Median OS and HRs for risk of
mortality in all patients.
|
| Only first-line
FOLFOX chemotherapy | First-line FOLFOX
plus second-line docetaxel-based chemotherapy |
|---|
|
|
|
|
|---|
| Variable | Median OS (95%
CI) | P-value | Risk of mortality,
HR (95% CI) | P-value | Median OS (95%
CI) | P-value | Risk of mortality,
HR (95% CI) | P-value |
|---|
| Stage |
| 0.180 |
|
|
| <0.001 |
|
|
|
III | 10.9
(8.6–13.1) |
| 0.66
(0.34–1.26) | 0.210 | 20.3
(12.5–28.1) |
| 0.37
(0.17–0.82) | 0.010 |
| IV | 8.9 (6.3–11.5) |
| 1.00
(reference) |
| 9.7 (7.8–11.6) |
| 1.00
(reference) |
|
| ECOG PS |
| 0.001 |
|
|
| 0.110 |
|
|
|
0–1 | 10.6
(8.4–12.7) |
| 0.34
(0.14–0.82) | 0.020 | 15.1
(11.8–18.4) |
| 0.17
(0.04–0.65) | 0.010 |
| 2 | 6.3 (0.0–15.3) |
| 1.00
(reference) |
| 8.1 (0.6–15.7) |
| 1.00
(reference) |
|
| Histological
grade |
| 0.240 |
|
|
| 0.140 |
|
|
| G2 | 12.1
(10.3–13.9) |
| 0.65
(0.30–1.41) | 0.270 | 20.2
(1.9–38.5) |
| 0.90
(0.33–2.45) | 0.840 |
|
G2-3 | 7.5 (6.4–8.7) |
| 1.57
(0.73–3.40) | 0.250 | 17.3
(12.1–22.4) |
| 0.60
(0.26–1.33) | 0.200 |
| G3 | 10.0
(9.0–11.0) |
| 1.00
(reference) |
| 11.3
(6.3–16.3) |
| 1.00
(reference) |
|
| BIM levels |
| 0.180 |
|
|
| 0.008 |
|
|
|
Low | 12.1
(9.0–15.2) |
| 0.79
(0.42–1.47) | 0.450 | 9.6 (8.9–10.3) |
| 2.61
(1.21–5.62) | 0.010 |
|
Intermediate and high | 9.6 (7.3–11.9) |
| 1.00
(reference) |
| 18.2
(12.8–23.6) |
| 1.00
(reference) |
|
Discussion
Following the failure of first-line chemotherapy,
several drugs are recommended for second-line regimens, including
paclitaxel, docetaxel and irinotecan (38). The median OS of patients receiving
second-line docetaxel-based regimens ranges from 3.5 to 10.9
months, which is still dismal (5).
The present authors previously observed that GC patients with high
BRCA1 levels could benefit from receiving second-line
docetaxel-based chemotherapy (6). In
addition, the median OS was further prolonged for patients with
high levels of BRCA1 and multiple myeloma SET domain (39).
The DNA damage caused by chemotherapy leads to cell
cycle arrest, DNA repair or commitment to apoptosis (40). Failure of drug-induced apoptosis is a
vital reason for chemoresistance (41). A previous study identified that
overexpression of genes involved in apoptosis appeared to
contribute to docetaxel sensitivity in breast cancer through
high-throughput screening of thousands of genes (42). It is commonly known that taxanes
interfere with the dynamics of the microtubules and induce
apoptosis through the mitochondrial apoptotic pathway (43).
In the present study, a marked difference in OS
(18.2 vs. 9.6 months) was observed in patients receiving
second-line docetaxel-based chemotherapy, according to their BIM
mRNA expression levels in univariate analysis (P=0.008). In
addition, this association was also significant in multivariate
analysis, which further confirmed the role of BIM as a predictive
biomarker. Patients with low BIM mRNA levels exhibited higher
mortality than those with high or intermediate BIM mRNA levels (HR,
2.61; 95% CI, 1.21–5.62; P=0.010). These results were consistent
with previously published data suggesting that overexpression of
BIM was accompanied by a collateral increase in sensitivity to
taxanes (19), which may translate
into prolonged OS.
BIM is an important mediator of tumor cell death
(15). Other studies have previously
demonstrated that several kinase-driven tumors, including chronic
myelogenous leukemia and NSCLC, maintain a survival advantage by
suppressing BIM transcription and by targeting BIM protein for
proteasomal degradation (44–46). Numerous studies have clearly
demonstrated that activation of the PI3K/AKT signaling pathway
could regulate BIM expression (44,47). The
PI3K/AKT signaling pathway triggers a cascade of cell responses,
including cell cycle progression, programmed cell death and DNA
damage repair in cancer (48).
Furthermore, the PI3K/AKT signaling pathway is closely associated
with the development and recurrence of cancer (49). The class O of forkhead box (FOXO)
transcription factors are downstream effectors of the PI3K/AKT
signaling pathway (50). When active,
FOXOs induce cell cycle arrest and apoptosis, acting as
anti-proliferative factors (51). BIM
is mainly regulated by FOXO3a, a member of the FOXO family
(52). Following an apoptotic-stress
event, BIM translocates to the mitochondria, and is essential to
mediate the release of cytochrome c from the mitochondria,
which in turn activates the effector caspase-9 and the formation of
the apoptosome (53). Previous
studies reported that the PI3K inhibitor LY294002 could increase
BIM expression and cell death, which partly demonstrated a
modulating role of PI3K/AKT signaling on BIM expression (47). The aforementioned results are in
agreement with the pattern of FOXO3a dephosphorylation and nuclear
translocation. The dephosphorylation status of AKT inhibits the
nuclear export of its substrate FOXO3a to the cytoplasm, which
transactivates the main target gene, BIM, to cause cell cycle
arrest and cell death (52). In a
previous study, BIM mRNA levels could be increased by upregulation
of FOXO3a following paclitaxel treatment, leading to apoptosis in
breast cancer cells and contributing to tumor sensitivity to
paclitaxel (54). Thus, the impact of
BIM and other BH3-only proteins in GC patients through apoptosis
pathways should be further investigated.
AEG-1 was identified as an oncogene that caused
detrimental effects to patients' OS through preventing cancer cells
from undergoing apoptosis (55).
Overexpression of AEG-1 leads to the activation of the PI3K/AKT
pro-survival signaling pathway and the downregulation of BCL-2
associated agonist of cell death (BAD), p21, p27 and FOXO3a
(24). In addition, increasing
expression and activation of FOXO3a by AEG-1 knockdown further
confirms this mechanism (56). AXL, a
receptor tyrosine kinase, was originally cloned from cancer cells
(57). A crucial step in
AXL-dependent signal transduction is the activation of PI3K/AKT
(58). The activation of AXL protects
cells from apoptosis and increases the expression of the
anti-apoptotic proteins BCL-2 and BCL-XL (59), as well as the phosphorylation of BAD
(60). AXL is also implicated in
angiogenesis (61) and immune
response (62). Preclinical findings
and retrospective studies have illustrated that overexpression of
AEG-1 and AXL confers broad drug resistance to chemotherapeutic
agents, including paclitaxel (63),
cisplatin (64,65) and 5-FU (27). However, no correlations were observed
in the present study between AEG-1 and AXL mRNA expression levels
and patients' outcome to chemotherapy, either in first-line or
second-line chemotherapy. This reflects the complexity of tumor
drug response and the fact that single genes may not be sufficient
to predict the therapeutic effect.
The present study has certain limitations. First,
the number of patients included in the study is relatively small,
which may cause bias in data analysis. In addition, the study is
retrospective in nature. Furthermore, multiple gene models or
signatures may be more effective than single biomarkers, as gene
expression patterns associated with drug resistance and sensitivity
are complex.
In conclusion, based on the significantly prolonged
OS among patients with high or intermediate BIM mRNA expression in
the present study, BIM may act as a potential biomarker in
second-line docetaxel-based chemotherapy for GC. The findings in
the current study pave the way for personalized chemotherapy in
GC.
Acknowledgements
The present study was funded by grants from the
National Natural Science Foundation of China (Beijing, China; grant
no. 81000980, 81220108023 and 81370064), the Fundamental Research
Funds for the Central Universities (Beijing, China; grant no.
20620140729), the Jiangsu Provincial Program of Medical Science
(Nanjing, China; grant no. BL2012001) and the Distinguished Young
Investigator Project of Nanjing (Nanjing, China; grant no.
JQX12002).
References
|
1
|
Fock KM: Review article: The epidemiology
and prevention of gastric cancer. Aliment Pharmacol Ther.
40:250–260. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wadhwa R, Song S, Lee JS, Yao Y, Wei Q and
Ajani JA: Gastric cancer-molecular and clinical dimensions. Nat Rev
Clin Oncol. 10:643–655. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wagner AD, Grothe W, Haerting J, Kleber G,
Grothey A and Fleig WE: Chemotherapy in advanced gastric cancer: A
systematic review and meta-analysis based on aggregate data. J Clin
Oncol. 24:2903–2909. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ji SH, Lim DH, Yi SY, Kim HS, Jun HJ, Kim
KH, Chang MH, Park MJ, Uhm JE, Lee J, et al: A retrospective
analysis of second-line chemotherapy in patients with advanced
gastric cancer. BMC Cancer. 9:1102009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wesolowski R, Lee C and Kim R: Is there a
role for second-line chemotherapy in advanced gastric cancer?
Lancet Oncol. 10:903–912. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wei J, Costa C, Ding Y, Zou Z, Yu L,
Sanchez JJ, Qian X, Chen H, Gimenez-Capitan A, Meng F, et al: mRNA
expression of BRCA1, PIAS1, and PIAS4 and survival after
second-line docetaxel in advanced gastric cancer. J Natl Cancer
Inst. 103:1552–1556. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yang X, Fraser M, Moll UM, Basak A and
Tsang BK: Akt-mediated cisplatin resistance in ovarian cancer:
Modulation of p53 action on caspase-dependent mitochondrial death
pathway. Cancer Res. 66:3126–3136. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yuan Z, Wang F, Zhao Z, Zhao X, Qiu J, Nie
C and Wei Y: BIM-mediated AKT phosphorylation is a key modulator of
arsenic trioxide-induced apoptosis in cisplatin-sensitive
and-resistant ovarian cancer cells. PLoS One. 6:e205862011.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Asselin E, Mills GB and Tsang BK: XIAP
regulates Akt activity and caspase-3-dependent cleavage during
cisplatin-induced apoptosis in human ovarian epithelial cancer
cells. Cancer Res. 61:1862–1868. 2001.PubMed/NCBI
|
|
10
|
Jamieson ER and Lippard SJ: Structure,
recognition, and processing of cisplatin-DNA adducts. Chem Rev.
99:2467–2498. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kelland L: The resurgence of
platinum-based cancer chemotherapy. Nat Rev Cancer. 7:573–584.
2007. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Alberti C: Taxane- and epothilone-based
chemotherapy: from molecule cargo cytoskeletal logistics to
management of castration-resistant prostate carcinoma. Eur Rev Med
Pharmacol Sci. 17:1658–1664. 2013.PubMed/NCBI
|
|
13
|
Zheng JH, Follis A Viacava, Kriwacki RW
and Moldoveanu T: Discoveries and controversies in BCL-2
protein-mediated apoptosis. FEBS J. 283:2690–2700. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Correia C, Lee SH, Meng XW, Vincelette ND,
Knorr KL, Ding H, Nowakowski GS, Dai H and Kaufmann SH: Emerging
understanding of Bcl-2 biology: Implications for neoplastic
progression and treatment. Biochim Biophys Acta. 1853:1658–1671.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Iurlaro R and Muñoz-Pinedo C: Cell death
induced by endoplasmic reticulum stress. FEBS J. 283:2640–2652.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Crawford N, Chacko AD, Savage KI, McCoy F,
Redmond K, Longley DB and Fennell DA: Platinum resistant cancer
cells conserve sensitivity to BH3 domains and obatoclax induced
mitochondrial apoptosis. Apoptosis. 16:311–320. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Simonin K, N'Diaye M, Lheureux S,
Loussouarn C, Dutoit S, Briand M, Giffard F, Brotin E,
Blanc-Fournier C and Poulain L: Platinum compounds sensitize
ovarian carcinoma cells to ABT-737 by modulation of the Mcl-1/Noxa
axis. Apoptosis. 18:492–508. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Inoue Y, Gika M, Abiko T, Oyama T, Saitoh
Y, Yamazaki H, Nakamura M, Abe Y, Kawamura M and Kobayashi K: Bcl-2
overexpression enhances in vitro sensitivity against docetaxel in
non-small cell lung cancer. Oncol Rep. 13:259–264. 2005.PubMed/NCBI
|
|
19
|
Savry A, Carre M, Berges R, Rovini A,
Pobel I, Chacon C, Braguer D and Bourgarel-Rey V: Bcl-2-enhanced
efficacy of microtubule-targeting chemotherapy through Bim
overexpression: Implications for cancer treatment. Neoplasia.
15:49–60. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Faber AC, Corcoran RB, Ebi H, Sequist LV,
Waltman BA, Chung E, Incio J, Digumarthy SR, Pollack SF, Song Y, et
al: BIM expression in treatment-naive cancers predicts
responsiveness to kinase inhibitors. Cancer Discov. 1:352–365.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Costa C, Molina MA, Drozdowskyj A,
Giménez-Capitán A, Bertran-Alamillo J, Karachaliou N, Gervais R,
Massuti B, Wei J, Moran T, et al: The impact of EGFR T790M
mutations and BIM mRNA expression on outcome in patients with
EGFR-mutant NSCLC treated with erlotinib or chemotherapy in the
randomized phase III EURTAC trial. Clin Cancer Res. 20:2001–2010.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Su ZZ, Kang DC, Chen Y, Pekarskaya O, Chao
W, Volsky DJ and Fisher PB: Identification and cloning of human
astrocyte genes displaying elevated expression after infection with
HIV-1 or exposure to HIV-1 envelope glycoprotein by rapid
subtraction hybridization, RaSH. Oncogene. 21:3592–3602. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hu G, Wei Y and Kang Y: The multifaceted
role of MTDH/AEG-1 in cancer progression. Clin Cancer Res.
15:5615–5620. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lee SG, Su ZZ, Emdad L, Sarkar D, Franke
TF and Fisher PB: Astrocyte elevated gene-1 activates cell survival
pathways through PI3K-Akt signaling. Oncogene. 27:1114–1121. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jian-bo X, Hui W, Yu-long H, Chang-hua Z,
Long-juan Z, Shi-rong C and Wen-hua Z: Astrocyte-elevated gene-1
overexpression is associated with poor prognosis in gastric cancer.
Med Oncol. 28:455–462. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Song L, Li W, Zhang H, Liao W, Dai T, Yu
C, Ding X, Zhang L and Li J: Over-expression of AEG-1 significantly
associates with tumour aggressiveness and poor prognosis in human
non-small cell lung cancer. J Pathol. 219:317–326. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yoo BK, Gredler R, Vozhilla N, Su ZZ, Chen
D, Forcier T, Shah K, Saxena U, Hansen U, Fisher PB and Sarkar D:
Identification of genes conferring resistance to 5-fluorouracil.
Proc Natl Acad Sci USA. 106:12938–12943. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Santarpia M, Magri I, Sanchez-Ronco M,
Costa C, Molina-Vila MA, Gimenez-Capitan A, Bertran-Alamillo J,
Mayo C, Benlloch S, Viteri S, et al: mRNA expression levels and
genetic status of genes involved in the EGFR and NF-.
|
|
29
|
B pathways in metastatic non-small-cell
lung cancer patients. J Transl Med. 9:1632011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Graham DK, DeRyckere D, Davies KD and Earp
HS: The TAM family: Phosphatidylserine sensing receptor tyrosine
kinases gone awry in cancer. Nat Rev Cancer. 14:769–785. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
van der Meer JH, van der Poll T and van 't
Veer C: TAM receptors, Gas6, and protein S: roles in inflammation
and hemostasis. Blood. 123:2460–2469. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Papadakis ES, Cichoń MA, Vyas JJ, Patel N,
Ghali L, Cerio R, Storey A and O'Toole EA: Axl promotes cutaneous
squamous cell carcinoma survival through negative regulation of
pro-apoptotic Bcl-2 family members. J Invest Dermatol. 131:509–517.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Verma A, Warner SL, Vankayalapati H,
Bearss DJ and Sharma S: Targeting Axl and Mer kinases in cancer.
Mol Cancer Ther. 10:1763–1773. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ou WB, Corson JM, Flynn DL, Lu WP, Wise
SC, Bueno R, Sugarbaker DJ and Fletcher JA: AXL regulates
mesothelioma proliferation and invasiveness. Oncogene.
30:1643–1652. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wimmel A, Glitz D, Kraus A, Roeder J and
Schuermann M: Axl receptor tyrosine kinase expression in human lung
cancer cell lines correlates with cellular adhesion. Eur J Cancer.
37:2264–2274. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Berclaz G, Altermatt HJ, Rohrbach V,
Kieffer I, Dreher E and Andres AC: Estrogen dependent expression of
the receptor tyrosine kinase axl in normal and malignant human
breast. Ann Oncol. 12:819–824. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Macleod K, Mullen P, Sewell J, Rabiasz G,
Lawrie S, Miller E, Smyth JF and Langdon SP: Altered ErbB receptor
signaling and gene expression in cisplatin-resistant ovarian
cancer. Cancer Res. 65:6789–6800. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Bubner B and Baldwin IT: Use of real-time
PCR for determining copy number and zygosity in transgenic plants.
Plant Cell Rep. 23:263–271. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Elimova E, Shiozaki H, Wadhwa R, Sudo K,
Chen Q, Estrella JS, Blum MA, Badgwell B, Das P, Song S and Ajani
JA: Medical management of gastric cancer: A 2014 update. World J
Gastroenterol. 20:13637–13647. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wei J, Costa C, Shen J, Yu L, Sanchez J,
Qian X, Sun X, Zou Z, Gimenez-Capitan A, Yue G, et al: Differential
effect of MMSET mRNA levels on survival to first-line FOLFOX and
second-line docetaxel in gastric cancer. Br J Cancer.
110:2662–2668. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Pan ST, Li ZL, He ZX, Qiu JX and Zhou SF:
Molecular mechanisms for tumour resistance to chemotherapy. Clin
Exp Pharmacol Physiol. 43:723–737. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Cree IA and Charlton P: Molecular chess?
Hallmarks of anti-cancer drug resistance. BMC cancer. 17:102017.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Chang JC, Wooten EC, Tsimelzon A,
Hilsenbeck SG, Gutierrez MC, Elledge R, Mohsin S, Osborne CK,
Chamness GC, Allred DC and O'Connell P: Gene expression profiling
for the prediction of therapeutic response to docetaxel in patients
with breast cancer. Lancet. 362:362–369. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Kang BW, Kwon OK, Chung HY, Yu W and Kim
JG: Taxanes in the rreatment of advanced gastric cancer. Molecules.
21:E6512016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Costa DB, Halmos B, Kumar A, Schumer ST,
Huberman MS, Boggon TJ, Tenen DG and Kobayashi S: BIM mediates EGFR
tyrosine kinase inhibitor-induced apoptosis in lung cancers with
oncogenic EGFR mutations. PLoS Med. 4:1669–1680. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Gong Y, Somwar R, Politi K, Balak M,
Chmielecki J, Jiang X and Pao W: Induction of BIM is essential for
apoptosis triggered by EGFR kinase inhibitors in mutant
EGFR-dependent lung adenocarcinomas. PLoS Med. 4:e2942007.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Kuroda J, Puthalakath H, Cragg MS, Kelly
PN, Bouillet P, Huang DC, Kimura S, Ottmann OG, Druker BJ,
Villunger A, et al: Bim and Bad mediate imatinib-induced killing of
Bcr/Abl+ leukemic cells, and resistance due to their
loss is overcome by a BH3 mimetic. Proc Natl Acad Sci USA.
103:14907–14912. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Qi XJ, Wildey GM and Howe PH: Evidence
that Ser87 of BimEL is phosphorylated by Akt and regulates BimEL
apoptotic function. J Biol Chem. 281:813–823. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Brown JS and Banerji U: Maximising the
potential of AKT inhibitors as anti-cancer treatments. Pharmacol
Ther. December 2–2016.(Epub ahead of print). View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Sun Y, Tian H, Wang L and Yang H: The
effects of silencing of PI3K p85α on 5-FU-induced colorectal cancer
cells apoptosis. Med Oncol. 30:7042013. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Carbajo-Pescador S, Mauriz JL,
García-Palomo A and González-Gallego J: FoxO proteins: Regulation
and molecular targets in liver cancer. Curr Med Chem. 21:1231–1246.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
de Mattos S Fernández, Villalonga P,
Clardy J and Lam EW: FOXO3a mediates the cytotoxic effects of
cisplatin in colon cancer cells. Mol Cancer Ther. 7:3237–3246.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Vogiatzi P, De Falco G, Claudio PP and
Giordano A: How does the human RUNX3 gene induce apoptosis in
gastric cancer? Latest data, reflections and reactions. Cancer Biol
Ther. 5:371–374. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Piñon JD, Labi V, Egle A and Villunger A:
Bim and Bmf in tissue homeostasis and malignant disease. Oncogene.
27:(Suppl 1). S41–S52. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Sunters A, de Mattos S Fernández, Stahl M,
Brosens JJ, Zoumpoulidou G, Saunders CA, Coffer PJ, Medema RH,
Coombes RC and Lam EW: FoxO3a transcriptional regulation of Bim
controls apoptosis in paclitaxel-treated breast cancer cell lines.
J Biol Chem. 278:49795–49805. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Hu G, Wei Y and Kang Y: The multifaceted
role of MTDH/AEG-1 in cancer progression. Clin Cancer Res.
15:5615–5620. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Wilson MS, Brosens JJ, Schwenen HD and Lam
EW: FOXO and FOXM1 in cancer: The FOXO-FOXM1 axis shapes the
outcome of cancer chemotherapy. Curr Drug Targets. 12:1256–1266.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Lemke G: Biology of the TAM receptors.
Cold Spring Harb Perspect Biol. 5:a0090762013. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Li Y, Jia L, Ren D, Liu C, Gong Y, Wang N,
Zhang X and Zhao Y: Axl mediates tumor invasion and
chemosensitivity through PI3K/Akt signaling pathway and is
transcriptionally regulated by slug in breast carcinoma. IUBMB
Life. 66:507–518. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Hasanbasic I, Cuerquis J, Varnum B and
Blostein MD: Intracellular signaling pathways involved in
Gas6-Axl-mediated survival of endothelial cells. Am J Physiol Heart
Circ Physiol. 287:H1207–H1213. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Goruppi S, Ruaro E, Varnum B and Schneider
C: Gas6-mediated survival in NIH3T3 cells activates stress
signalling cascade and is independent of Ras. Oncogene.
18:4224–4236. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Holland SJ, Powell MJ, Franci C, Chan EW,
Friera AM, Atchison RE, McLaughlin J, Swift SE, Pali ES, Yam G, et
al: Multiple roles for the receptor tyrosine kinase axl in tumor
formation. Cancer Res. 65:9294–9303. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Tjwa M, Bellido-Martin L, Lin Y, Lutgens
E, Plaisance S, Bono F, Delesque-Touchard N, Hervé C, Moura R,
Billiau AD, et al: Gas6 promotes inflammation by enhancing
interactions between endothelial cells, platelets, and leukocytes.
Blood. 111:4096–4105. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Hu G, Chong RA, Yang Q, Wei Y, Blanco MA,
Li F, Reiss M, Au JL, Haffty BG and Kang Y: MTDH activation by 8q22
genomic gain promotes chemoresistance and metastasis of
poor-prognosis breast cancer. Cancer Cell. 15:9–20. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Li C, Li Y, Wang X, Wang Z, Cai J, Wang L,
Zhao Y, Song H, Meng X, Ning X, et al: Elevated expression of
astrocyte elevated gene-1 (AEG-1) is correlated with
cisplatin-based chemoresistance and shortened outcome in patients
with stages III–IV serous ovarian carcinoma. Histopathology.
60:953–963. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Hong J, Peng D, Chen Z, Sehdev V and
Belkhiri A: ABL regulation by AXL promotes cisplatin resistance in
esophageal cancer. Cancer Res. 73:331–340. 2013. View Article : Google Scholar : PubMed/NCBI
|