Introduction
Hepatocellular carcinoma (HCC) is the fifth most
common type of malignancy worldwide and is the third leading cause
of cancer-associated mortality worldwide (1). The majority of patients with HCC are
diagnosed when the disease is at an advanced stage, and potentially
curative therapeutic options for HCC are limited (2). Thus, there is an urgent need to identify
and establish alternative approaches for the treatment of HCC.
Immunotherapy has gained attention in previous years as a promising
adjuvant therapy for HCC patients. A complex role of the immune
system in the development and progression of HCC has been
highlighted by a growing number of studies (3–5). To
improve the clinical efficacy of cancer immunotherapy, a detailed
understanding of the immunological mechanism of tumorigenesis and
progression is required.
The tumor microenvironment is a systematic concept
that defines the behavior of cancer not only by the genetics of the
tumor cells alone but also by the surrounding milieu. Tumor
infiltrating lymphocytes (TILs), an important part of the tumor
surveillance system, have been detected in a number of patients
with HCC. In particular, cluster of differentiation (CD)
8+ cytotoxic T lymphocytes (CTLs) were found to be
linked to an improved patient survival (6,7). However,
the spontaneous clearance of established HCC lesions by endogenous
immune mechanisms is rare, despite spontaneous humoral and cellular
immune responses having been detected in a significant proportion
of patients with HCC against a number of tumor-associated antigens
(TAAs) (8–10). As a number of TAAs are derived from
normal self-constituents, anti-tumor immunity against these TAAs
was not sufficient to control the growth of the tumors.
Additionally, in the pressure of a host immune system, HCC has
evolved multiple passive and active mechanisms that generate a
suppressive network in order to evade the host's anti-tumor immune
response (11,12). Such mechanisms include an accumulation
of immunosuppression by regulatory T cells (Tregs),
tumor-associated macrophages and dysfunctional dendritic cells (DC)
(13–15). Tregs are the most extensively studied
mechanism, and an increased frequency of Tregs in the peripheral
blood and tumor tissues of HCC has been studied previously
(16,17). Furthermore, the upregulation of Tregs
is notably coupled with a significant decrease and impaired
effector function of cytotoxic CD8+ T cells, and a worse
predicted survival rate for patients with HCC (18,19).
However, cells with a potential suppressive function against
anti-tumor immunity are not confined to this cell subset.
Regulatory γδ T cells and natural killer (NK) T cells had been
gradually identified as having a potential suppressive function by
several studies (20–22). The distribution of forkhead box P3
(FOXP3)+ cells in various cell subsets of HCC has been
presented in our previous study (23).
The range of lymphocytes present in the liver
differs markedly from that in the peripheral blood and other organs
(24–26). Lymphocytes in the liver are
characterized by a high proportion of NK cells and significant
numbers of several unconventional T cell subsets, such as T cells
that co-express CD3 and CD56, which can also be referred to as
natural T cells (27). Considering
the high prevalence of NK and natural T cells in the liver, it is
suggested that they perform a critical role in immune homeostasis
and pathogenesis of the liver. The present study was interested in
the implication of NK and natural T cells in the pathology of HCC.
As reported by Kawarabayashi et al (28) a decreased abundance of NK cells and
natural T cells in liver diseases may be involved in their
susceptibility to HCC. Therefore, the principal objective of the
present study was to determine the representation of NK cells and
natural T cells in local tumor tissues and in peripheral blood of
patients with HCC. The present study observed a marked reduction in
NK cells and natural T cells in TILs vs. non-tumor infiltrating
lymphocytes (NILs). However, this pattern was not observed in
peripheral blood mononuclear cells (PBMCs) of patients with HCC vs.
healthy donors. In addition, a new type of FOXP3-expressing natural
T cell was identified in the tumorous tissues of HCC, and several
phenotypic and functional tests were then performed on this special
cell subset. It is considered that this discovery may provide a new
mechanistic explanation for HCC induced immunosuppression, and
outline a previously unrecognized potential target for the
immunotherapy of HCC.
Materials and methods
Patients and healthy donors
This study was approved by the hospital ethics
review committee at the Peking University People's Hospital
(Beijing, China), and written informed consent was obtained from
all patients prior to the start of the study. A total of 16 paired
tumorous and adjacent non-tumorous tissue samples were collected
from patients with HCC at the time of surgery. Surgery was
performed at the Center of Hepatobiliary Surgery, Tianjin Medical
University Cancer Institute and Hospital (Tianjin, China). Blood
samples from 11 of these patients were also collected, and blood
samples from 11 healthy volunteers were obtained from the blood
bank of Beijing Red Cross to use as controls. HCC was diagnosed
according to the diagnostic guidelines of the European Association
for the Study of the Liver (Geneva, Switzerland). Patient
characteristics and demographic data are shown in Table I.
 | Table I.Clinical characteristics of 16
patients. |
Table I.
Clinical characteristics of 16
patients.
| Variables | Results |
|---|
| Gender
(male/female) | 10/6 |
| Age (year, range of
median) | 60 (38–74) |
| α-fetoprotein level
(ng/ml, range of median) | 366 (0–1,210) |
| Hepatitis B surface
antigen positivity (%) | 68.75 |
| Hepatitis C virus
antibody positivity (%) | 6.25 |
|
Tumor-node-metastasis stage
(I/II/III/IV) | 3/6/5/2 |
| Differentiation
(high/medium/low) | 2/12/2 |
Cell isolation and purification
Tumorous and non-tumorous (>5 cm from the tumor
margin) tissue infiltrating lymphocytes (TILs and NILs) were
obtained using a method based on mechanical dissociation and
collagenase treatment as described previously (23). Freshly obtained blood samples were
immediately centrifuged (800 × g, 25°C) for 20 min on Ficoll-Paque
density gradients (Beijing Solarbio Science & Technology,
Beijing, China) to collect PBMCs. All isolated cells were either
used immediately in experiments, or cryopreserved for future use.
TILs or NILs were stained with the appropriate combinations of
fluorescence labeled antibodies for fluorescence activated cell
sorting (FACS) analysis or sorted using a BD FACSAria cell sorter
(BD Biosciences, San Diego, CA, USA) for purification of
CD25+CD3+CD56+,
CD4+CD25high and
CD4+CD25− cell subsets with a purity of
95–98%.
Phenotypic and functional flow
cytometric analysis
Multicolor FACS analysis was performed to determine
the frequency and phenotype of natural T/NK cells in TILs, NILs and
PBMCs using a series of fluorescence labeled monoclonal antibodies
in 1:10 dilutions, including peridinin-chlorophyll-protein
complex-Cy5.5-anti-human CD3 (catalog no. 340949),
phycoerythrin-anti-human CD56 (catalog no. 555516), fluorescein
isothiocyanate (FITC)-anti-human CD4 (catalog no. 555346),
allophycocyanin (APC)-anti-human CD8 (catalog no. 555349),
FITC-anti-human CD25 (catalog no. 555431), APC-anti-human CD25
(catalog no. 555434), FITC-anti-human CD45RO (catalog no. 11-0457),
and FITC-anti-human cytotoxic T-lymphocyte-associated protein 4
(CTLA-4; catalog no. 11-1529), with appropriate isotype-matched
controls. Antibodies against CD45RO and CTLA-4 were obtained from
eBioscience, Inc. (San Diego, CA, USA), and the rest were from BD
Pharmingen (San Diego, CA, USA). For intracellular staining of
interferon (IFN)-γ and perforin, FITC-anti-human IFN-γ (1:10;
catalog no. 552887) and Alexa 488-anti-human perforin (1:10;
catalog no. 563764; BD Pharmingen) antibodies were used. The cells
were stimulated with 50 ng/ml phorbol-12-myristate-13-acetate (PMA,
Sigma-Aldrich; Merck Millipore, Darmstadt, Germany), and 10 µg/ml
ionomycine (Sigma-Aldrich; Merck Millipore) for 4 h at 37°C. For
the remaining 2 h of this incubation, 1 µg/ml brefeldin A
(Sigma-Aldrich; Merck Millipore) was added into the culture. The
culture was subsequently incubated with previously mentioned
fluorescent-labeled antibodies at 4°C for 30 min and
CytoFix/Cytoperm (BD Pharmingen) as previously mentioned. Staining
for the FOXP3 protein was performed using a FOXP3 kit (1:20;
APC-anti-human FOXP3; catalog no. 77-5774; eBioscience Inc.)
according to the manufacturer's protocol. Cells were analyzed using
the FACSCalibur flow cytometer (BD Biosciences) and analyzed using
CellQuest software (BD Biosciences).
T cell proliferation and suppression
assay
CD25+CD3+CD56+
cells from the tumor tissues of 3 patients with HCC were isolated
and purified using FACSAria (BD Biosciences). These cells were used
to represent FOXP3+ natural T cells (NTreg), since
FOXP3, which is an intracellular molecule, could not be used as a
marker in cell sorting to keep cells alive for the functional
assay. To detect the anti-proliferative effect of NTreg, the
present study performed a co-culture test using purified autologous
CD4+CD25− cells as responder cells
(5×104 cells/well). These cells were preloaded with
carboxyfluorescein diacetate succinimidyl ester (CFSE) with NTreg
at a ratio of 1:1 in 96-well round-bottomed plates (Corning Costar
Incorporated, NY, USA) in triplicate.
Cellular proliferation was induced by stimulation
with the anti-CD3 antibody (1 µg/ml) in the presence of irradiated
allogeneic PBMCs as antigen-presenting cells (2×105
cells/well, irradiated at 35 Gy) for 4 days at 37°C. Proliferation
was measured by monitoring CFSE dilution and the results were shown
in histograms subsequent to gating on CFSE+ cells.
Statistical analysis
Data are presented as the mean ± standard error of
the mean for percentages. The significance of the difference
between group means was determined using the Student's t-test by
using GraphPad Prism (version5.0; GraphPad Software, Inc., La
Jolla, CA, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
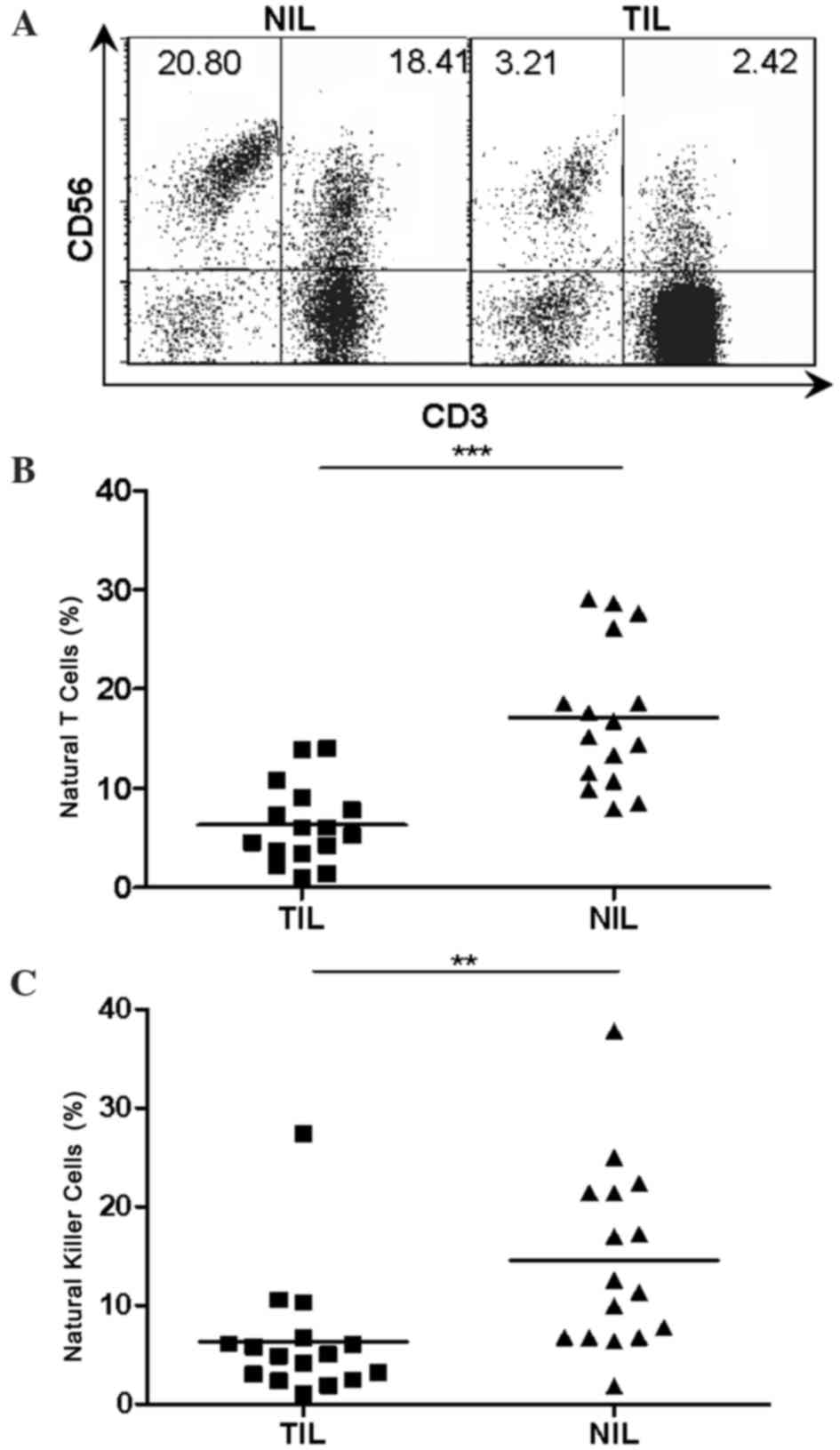
Reduced natural T and NK cells in TILs
of patients with HCC
Considering the abundance of natural T cells and NK
cells in the human liver, it was necessary to investigate the
representation of natural T cells and NK cells in local tumor
tissues of human HCC. TILs and NILs were obtained as described
previously (23). Flow cytometry was
then performed on a total of 16 paired TILs and NILs using
fluorescence labeled monoclonal antibodies specific for CD3 and
CD56. The percentages of natural T cells (6.315±1.002 vs.
17.16±1.804, P<0.001) and NK cells (6.324±1.559 vs. 14.52±2.336,
P<0.01) were markedly reduced in TILs compared with that in
NILs, as determined by flow cytometry analysis (Fig. 1).
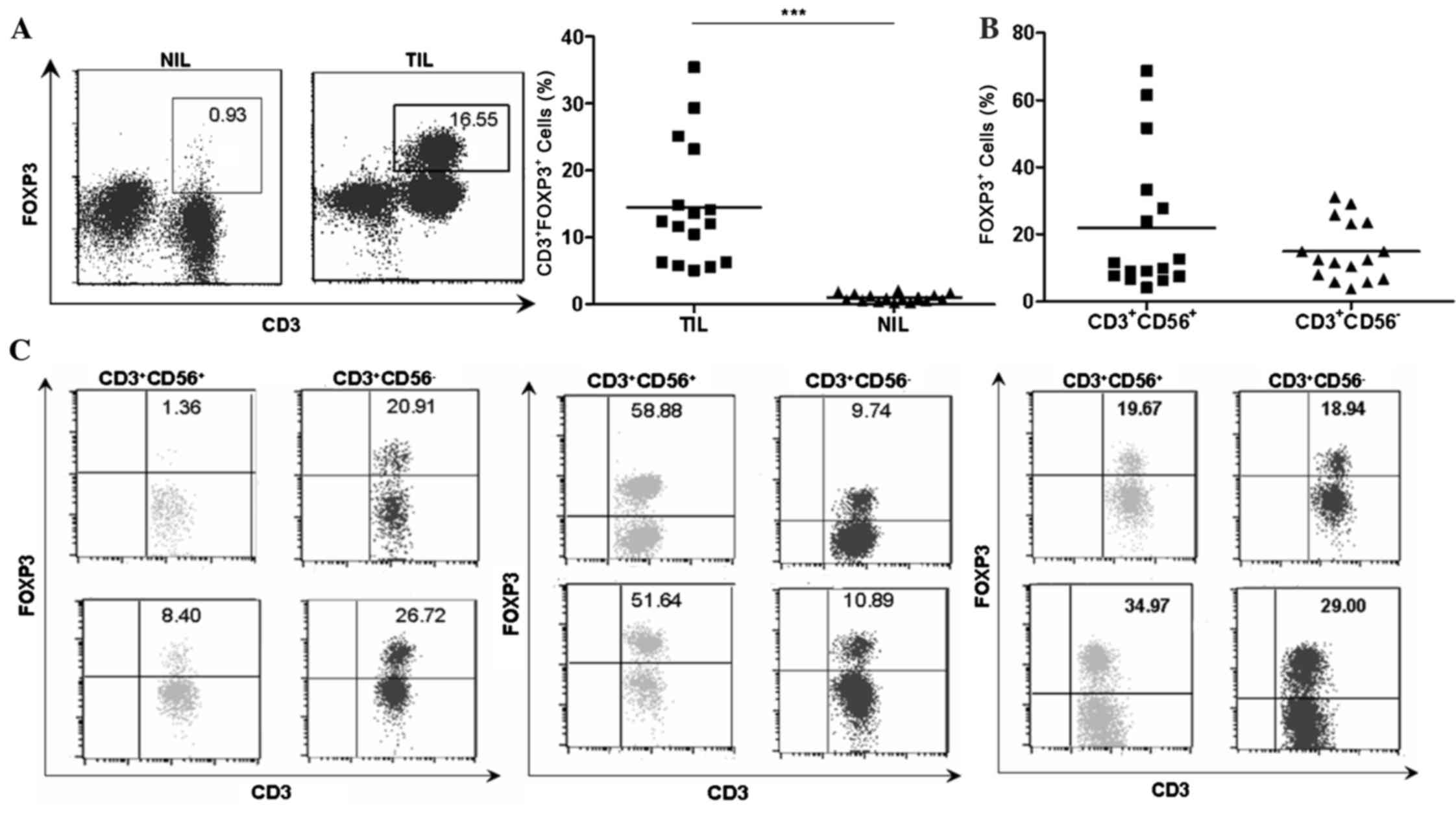
Identification of FOXP3+
natural T cells in TILs of patients with HCC
As shown in Fig. 2A,
the percentages of FOXP3+ T cells were higher in TILs
compared with NILs (14.48±2.297 vs. 1.023±0.1411, P<0.001)
despite a large variation among individual patients with HCC. The
present study subsequently analyzed the respective FOXP3 expression
in different cell subsets. Notably, FOXP3+ cells were
identified not in conventional T cells
(CD3+CD56−) and in natural T cells
(CD3+CD56+; Fig.
2B). The NK cells (CD3−CD56+) in TILs did
not acquire FOXP3 expression. As shown in in Fig. 2C, there were 3 different distribution
patterns of FOXP3+ cell subset in autologous natural T
cell and conventional T cell populations in TILs, which suggested
that there was no association between the level of FOXP3 and CD56
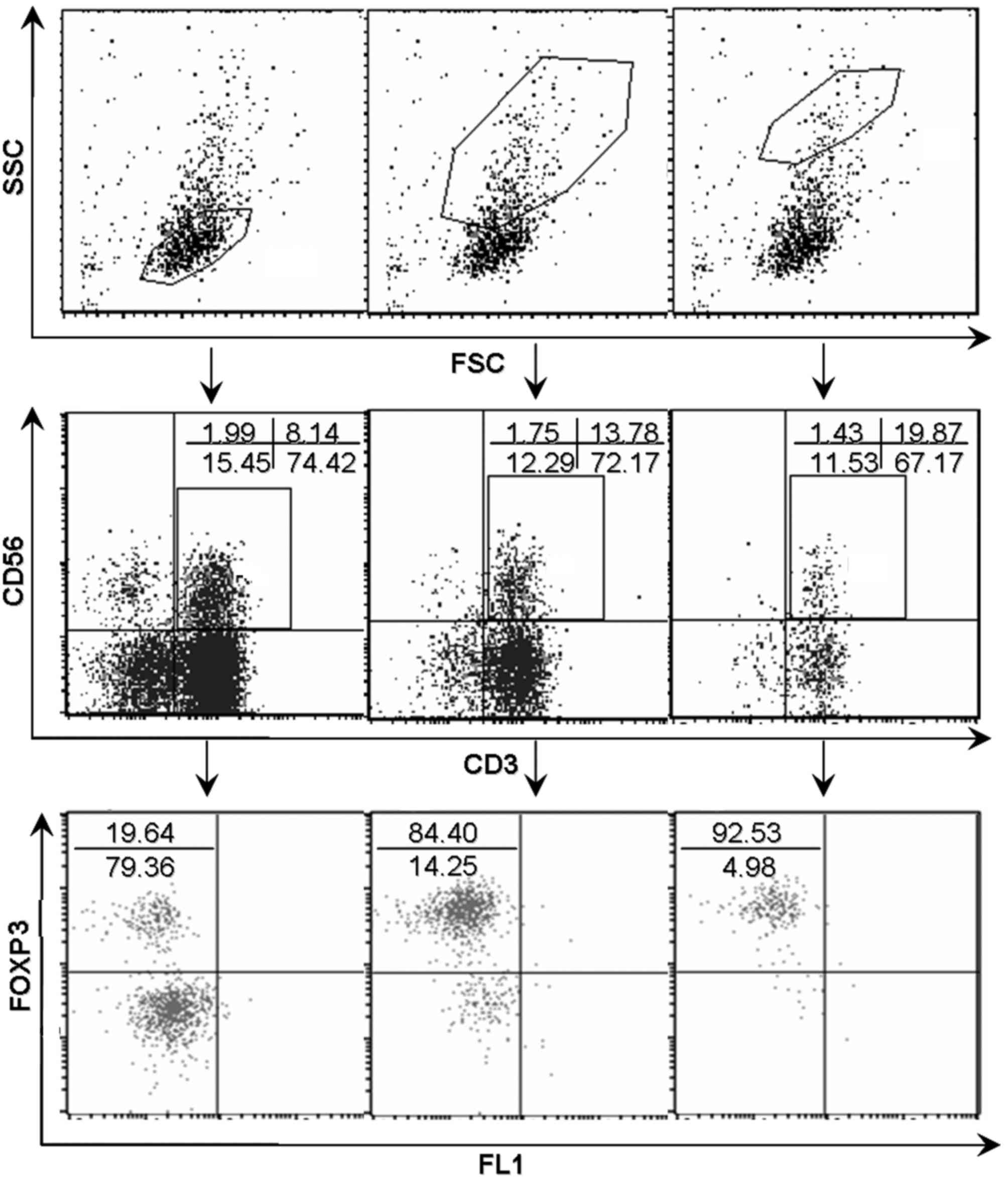
expression. By contrast, dot plots in Fig. 3 showed a positive association between
the level of FOXP3 expression and forward scatter/side scatter
gating in natural T cells.
Representation of natural T cells and
NK cells in PBMCs of patients with HCC
In contrast with what was found in the local tumor
tissues, there was no clear change in the percentages of natural T
cells (Fig. 4A, P>0.05) and NK
cells (Fig. 4B, P>0.05) in the
peripheral blood from patients with HCC in comparison with that
from healthy donors. In addition, the prevalence of
FOXP3+ cells was not observed in natural T cell
populations in the peripheral blood of patients with HCC, despite a
significant accumulation of FOXP3+ cells in autologous
conventional T cell population in TILs having been detected at the
same time (Fig. 4C, P<0.001).
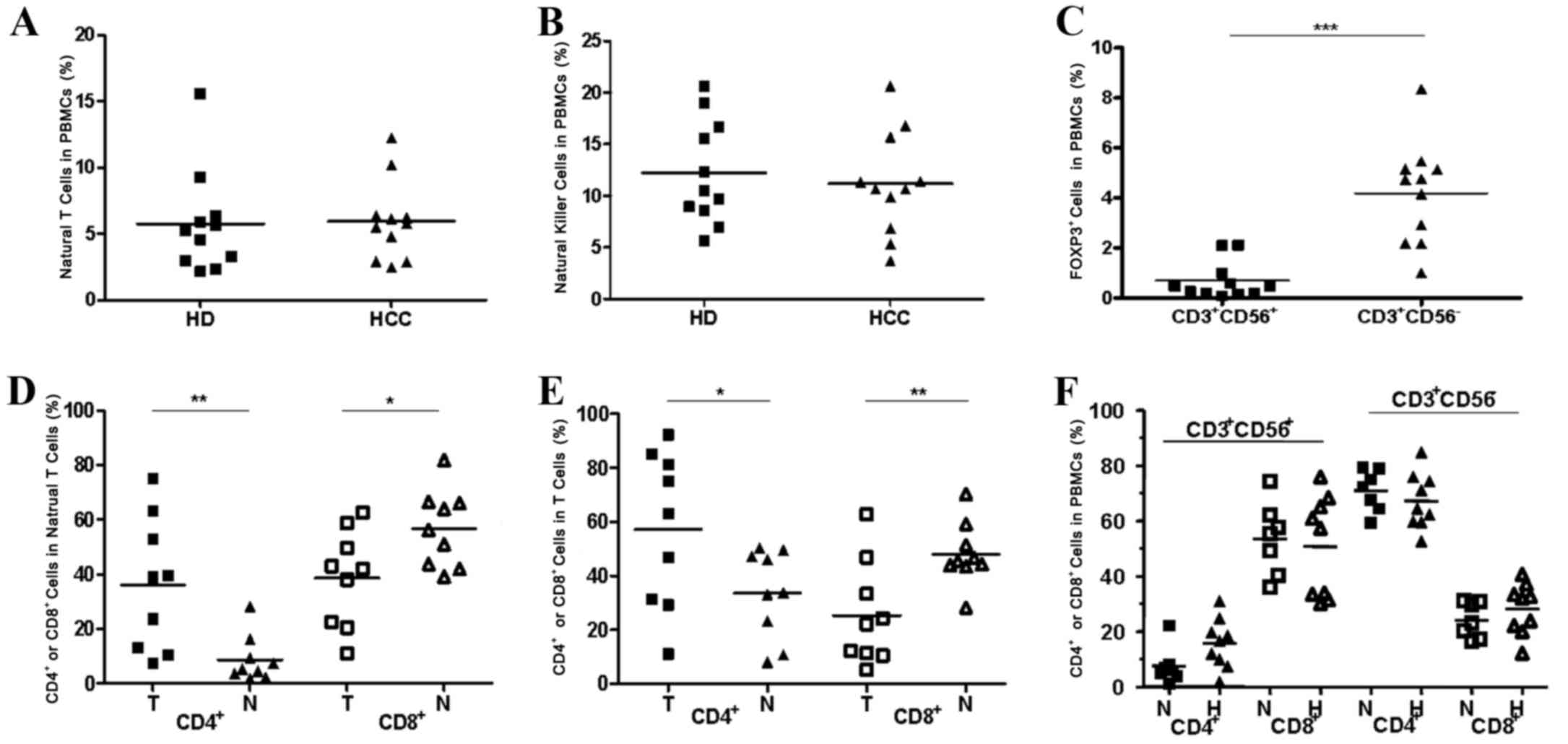 | Figure 4.Comparison of natural T cells and NK
cells in PBMCs between patients with HCC and healthy donors and an
altered CD4/CD8 distribution pattern in natural T cells and
conventional T cells in tumorous tissues compared with non-tumorous
tissues. (A) The percentages of natural T cells in PBMCs from
patients with HCC (n=11) and from healthy donors (n=11). (B) The
percentages of NK cells in PBMCs from patients with HCC (n=11) and
in healthy donors (n=11). (C) The percentages of FOXP3+
cells in natural T cells (CD3+CD56+) and
conventional T cells (CD3+CD56−) in PBMCs
from individual patients (n=11). (D) An altered CD4/CD8
distribution pattern in natural T cells in tumor tissues compared
with non-tumor tissues. (E) An altered CD4/CD8 distribution pattern
in conventional T cells in tumorous tissues compared with
non-tumorous tissues was observed. (F) An unaltered CD4/CD8
distribution pattern in natural T cells and T cells in PBMCs from
patients with HCC compared with normal donors. *P<0.05,
**P<0.01, ***P<0.001. PBMCs, peripheral blood mononuclear
cells; HCC, hepatocellular carcinoma; CD, cluster of
differentiation; HD, healthy donors; FOXP3, forkhead box P3; T,
tumor; H, patients with HCC; N, normal donors. |
One unique characteristic of resident human hepatic
lymphocytes is a reversed CD4/CD8 ratio (1:3.5) compared with that
in PBMCs (2:1) (24). The present
study found the percentage of CD4+ cells to be
observably higher in the natural T cell subset (Fig. 4D, P<0.01) and the conventional T
cell subset (Fig. 4E, P<0.05) in
TILs than in NILs. Conversely, the percentage of the
CD8+ cell subset was markedly reduced in TILs compared
with NILs (Fig. 4D, P<0.05;
Fig. 4E, P<0.01). Nevertheless,
there were no notable changes in the percentages of the
CD4+ cell population and the CD8+ cell
population in the natural T cell subset and the conventional T cell
subset when referring to the PBMCs from patients with HCC in
comparison with that from healthy donors (Fig. 4F, P>0.05).
Phenotypic and functional
characterization of FOXP3+ natural T cells in TILs
Similar to conventional Tregs, the majority of the
FOXP3+ cell population in natural T cells exhibited
positive expression of CD25 and CTLA-4 (Fig. 5A).
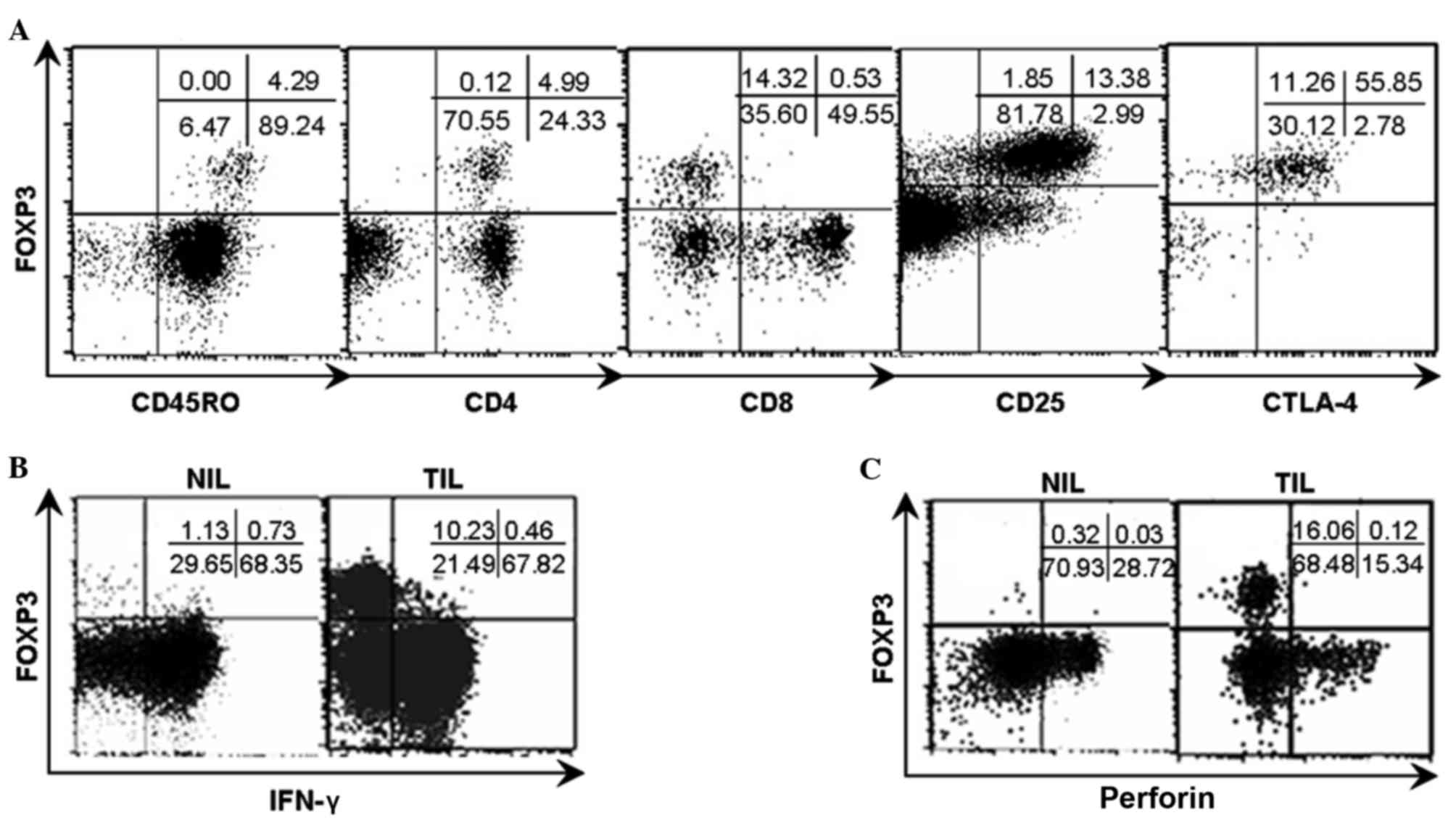 | Figure 5.Phenotypic characteristics of
FOXP3+ natural T cells and the FOXP3/IFN-γ and
FOXP3/perforin patterns in TILs and NILs from representative
patients. Analyses were repeated with samples from 8–10 HCC
patients with similar results. (A) The phenotypic analysis
indicated that the FOXP3+ natural T cells were
restricted to CD45RO+, CD4+,
CD25+, CTLA-4+ and CD8−
populations. (B) TILs and NILs were stimulated with PMA and
ionomycine for 4 h respectively and subsequently analyzed for
FOXP3/IFN-γ pattern subsequent to gating on natural T cells by flow
cytometry. (C) TILs and NILs were analyzed for FOXP3/perforin
pattern subsequent to gating on natural T cells by flow cytometry.
The numbers indicate the percentage of corresponding subsets.
FOXP3, forkhead box P3; IFN-γ, interferon-γ; TILs, tumor
infiltrating lymphocytes; NILs, non-tumor infiltrating lymphocytes;
HCC, hepatocellular carcinoma; CD, cluster of differentiation;
CTLA-4+, cytotoxic T-lymphocyte-associated protein 4;
PMA, phorbol-12-myristate-13-acetate. |
IFN-γ and perforin are important
effector molecules for natural T cells
TILs and NILs were stimulated with PMA (50 ng/ml)
and ionomycine (1 µg/ml) for 4 h, respectively, and subsequently
examined for the expression of IFN-γ and perforin in natural T
cells in the context of FOXP3 expression by flow cytometry. The
present results demonstrate that the acquisition of FOXP3 appeared
to be accompanied with the loss of expression of IFN-γ and
perforin, due to the mutually exclusive expression between IFN-γ
and perforin and FOXP3 as shown in the dot plots (Fig. 5B and C).
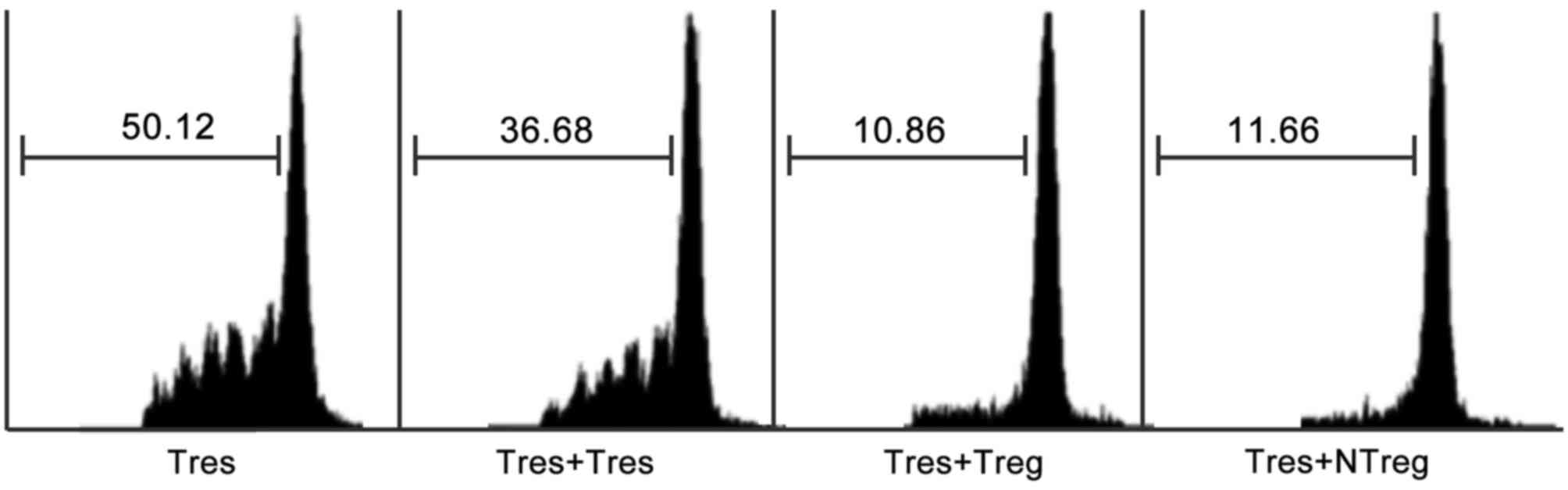
Immunosuppressive function of
FOXP3+ natural T cells in TILs of patients with HCC
It was important to distinguish whether the
expression of FOXP3 in natural T cells represented the
immunosuppressive function acquisition. As an intracellular
molecule, FOXP3 could not be used as a marker to obtain live cells
for functional analysis.
CD25+CD3+CD56+ cells were purified
from TILs of patients with HCC to represent FOXP3+
natural T cells, since the majority of FOXP3+ natural T
cells were CD25+, whereas CD25 expression was hardly
detected in FOXP3− natural T cells as indicated in
Fig. 5A. In order to test the
immunosuppressive function of FOXP3+ natural T cells in
TILs, a CFSE assay through co-culture of
CD25+CD3+CD56+ cells with CFSE
labeled autologous CD4+CD25− responder T
cells was performed as previously described. As expected, results
from the CFSE dilution assay indicated that FOXP3+
natural T cells exerted a comparable inhibitory effect on the
proliferation of responder T cells to that mediated by conventional
Tregs (Fig. 6).
Discussion
A number of studies have documented HCC progress in
the presence of tumor-specific immune responses in a majority of
patients with HCC, indicating that HCC has evolved multiple
mechanisms to evade host anti-tumor immunity (3–5,11,12). To
improve the efficacy of immunotherapy for HCC, it is critical to
improve understanding of the mechanisms behind this immune
suppression.
Although it is well established that reduced
CD8+ CTLs and increased FOXP3+ cells in HCC
are coupled with poor prognosis (18,19), it is
now also being recognized that the innate arm of the immune system
may also perform a potentially important role in immune responses
in liver diseases (25,28–32).
Suppressive innate immunocytes including regulatory DCs, γδ T cells
and NKT cells have gained attention in previous years (15,20–22).
The present study describes a different
representation of natural T cells and NK cells between TILs and
peripheral lymphocytes of human HCC. Natural T cells and NK cells
were markedly decreased in TILs vs. NILs, whilst the proportions
were not altered in PBMCs from patients with HCC in comparison with
that from healthy donors. By contrast, several studies have
indicated a reduction in the proportion of NK cells in vPBMCs from
patients with HCC (33,34). Conversely, a previous investigation
from 28 patients with HCC in tumor node metastasis stage I
demonstrated that the proportion of NK cells in PBMCs did not
differ from the proportion of NK cells in PBMCs from healthy
controls (35). These conflicting
studies may be in part due to the relatively low proportion of NK
cells in PBMCs of the included healthy donors in the present
study.
The present study identified a novel
FOXP3+ natural T cell population
(CD3+CD56+FOXP3+) with a similar
phenotype and comparable suppressive capacity to conventional Tregs
in TILs of HCC. Due to the resemblance between FOXP3+
natural T cells and conventional FOXP3+ Tregs, it is
necessary to determine whether or not the FOXP3+ natural
T cells are a special subset of Tregs that acquire CD56 expression
at the same time. A correlation analysis was firstly performed
between the proportion of FOXP3-expressing cell population in the
natural T cell subset (CD3+CD56+) and that in
the conventional T cell subset (CD3+CD56−) in
TILs. These results however suggested that there is no significant
correlation between these factors. The mean proportion of FOXP3
expressing cells was higher in the natural T cell subset compared
with that in the conventional T cell subset in TILs (Fig. 2B). In individuals, the proportions of
FOXP3+ cells in the natural T cell subset and in the
conventional T cell subset in TILs were not always equivalent
(Fig. 2C). Regarding the PBMCs from
patients with HCC, an unaltered representation of natural T cells
and NK cells was observed in comparison with that from healthy
donors, and a significant proportion of the FOXP3+ cell
population only existed in the conventional T cell subset but not
in the natural T cell subset. In NK cells, no FOXP3-expressing
cells were detected in either TILs or PBMCs. In summary, it was
identified that FOXP3+ natural T cells spontaneously
existed in TILs of HCC as a new regulatory cell subset, which were
mostly derived from natural T cells infiltrating in the local
microenvironment of HCC but not recruited from the peripheral
blood.
As studied, apart from conventional
CD4+FOXP3+ regulatory cells, CD8+
and T cell receptor (TCR) γδ+ regulatory T cells have
been identified in prostate and breast cancer, respectively
(36,37). Although TCRγδ+ and
CD8+ cell populations were dominant in the natural T
cell subset in the human liver, the identified FOXP3+
natural T cells in TILs of HCC were confined to TCRγδ−
and CD8− cell populations. In addition, the acquisition
of FOXP3, appeared to be accompanied with the loss of classical
functions of natural T cells, since the expression of FOXP3, IFN-γ
and perforin in natural T cells was mutually exclusive on single
cell level through the flow cytometric scatter plots graphs
(Fig. 5B and C). By contrast,
granzyme B and perforin were found to be relevant for
Tregs-mediated suppression of tumor clearance in vivo
(38).
FOXP3 is essential for the programming, development,
differentiation and suppressive functioning of Tregs, and is a
definitive marker for Tregs (39). A
number of studies have demonstrated that overexpression of FOXP3 in
naïve T cells and/or T cell lines through lentiviral infection
resulted in acquisition of suppressive activity (40–45). The
authors of the present study speculate whether in vitro
FOXP3-lentiviral infection is sufficient for the acquisition of
immunosuppressive activity in natural T cells, and this will be
investigated in further studies. Although there was a high
percentage of natural T cells with FOXP3+ expression in
TILs, the prevalence of FOXP3+ natural T cells in TILs
was not as high as Tregs in TILs, due to a decrease in cell numbers
in the overall natural T cell subset in TILs.
In conclusion, the identification of the unique
FOXP3+ natural T cells may be helpful to improve
understanding of the immunosuppressive local microenvironment in
HCC, and it is hypothesized that it may also provide useful clues
for the improvement of therapeutic intervention of HCC in the
future.
Acknowledgements
The present study was supported in part by grants
from the National Science and Technology Major Project (grant no.
2013ZX09303001), the National Natural Science Foundation of China
(grant nos. 81501984, 81601377), the Tianjin Municipal Bureau of
Health Science & Technology Fund (grant no. 2013KZ088) and the
Tianjin Medical University Science Fund (grant no. 2013KYQ07).
References
|
1
|
Lafaro KJ, Demirjian AN and Pawlik TM:
Epidemiology of hepatocellular carcinoma. Surg Oncol Clin N Am.
24:1–17. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kulik LM and Chokechanachaisakul A:
Evaluation and management of hepatocellular carcinoma. Clin Liver
Dis. 19:23–43. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hui D, Qiang L, Jian W, Ti Z and Da-Lu K:
A randomized, controlled trial of postoperative adjuvant
cytokine-induced killer cells immunotherapy after radical resection
of hepatocellular carcinoma. Dig Liver Dis. 41:36–41. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wirth TC: Spontaneous and therapeutic
immune responses in hepatocellular carcinoma: Implications for
current and future immunotherapies. Expert Rev Gastroenterol
Hepatol. 8:101–110. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Breous E and Thimme R: Potential of
immunotherapy for hepatocellular carcinoma. J Hepatol. 54:830–834.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Unitt E, Marshall A, Gelson W, Rushbrook
SM, Davies S, Vowler SL, Morris LS, Coleman N and Alexander GJ:
Tumour lymphocytic infiltrate and recurrence of hepatocellular
carcinoma following liver transplantation. J Hepatol. 45:246–253.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Flecken T, Schmidt N, Hild S, Gostick E,
Drognitz O, Zeiser R, Schemmer P, Bruns H, Eiermann T, Price DA, et
al: Immunodominance and functional alterations of tumor-associated
antigen specific CD8+ T-cell responses in hepatocellular carcinoma.
Hepatology. 59:1415–1426. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang HG, Chen HS, Peng JR, Shang XY,
Zhang J, Xing Q, Pang XW, Qin LL, Fei R, Mei MH, et al: Specific
CD8(+) T cell responses to HLA-A2 restricted MAGE-A3 p271-279
peptide in hepatocellular carcinoma patients without vaccination.
Cancer Immunol Immunother. 56:1945–1954. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liu Y, Daley S, Evdokimova VN, Zdobinski
DD, Potter DM and Butterfield LH: Hierarchy of alpha fetoprotein
(AFP)-specific T cell responses in subjects with AFP-positive
hepatocellular cancer. J Immunol. 177:712–721. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Korangy F, Ormandy LA, Bleck JS,
Klempnauer J, Wilkens L, Manns MP and Greten TF: Spontaneous
tumor-specific humoral and cellular immune responses to NY-ESO-1 in
hepatocellular carcinoma. Clin Cancer Res. 10:4332–4341. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhao F, Korangy F and Greten TF: Cellular
immune suppressor mechanisms in patients with hepatocellular
carcinoma. Dig Dis. 30:477–482. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Unitt E, Rushbrook SM, Marshall A, Davies
S, Gibbs P, Morris LS, Coleman N and Alexander GJ: Compromised
lymphocytes infiltrate hepatocellular carcinoma: The role of
T-regulatory cells. Hepatology. 41:722–730. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hoechst B, Ormandy LA, Ballmaier M, Lehner
F, Krüger C, Manns MP, Greten TF and Korangy F: A new population of
myeloid-derived suppressor cells in hepatocellular carcinoma
patients induces CD4(+)CD25(+)Foxp3(+) T cells. Gastroenterology.
135:234–243. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Shirabe K, Mano Y, Muto J, Matono R,
Motomura T, Toshima T, Takeishi K, Uchiyama H, Yoshizumi T,
Taketomi A, et al: Role of tumor-associated macrophages in the
progression of hepatocellular carcinoma. Surg Today. 42:1–7. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Han YM, Chen ZB, Yang Y, Jiang Z, Gu Y,
Liu Y, Lin C, Pan Z, Yu Y, Jiang M, et al: Human CD14+CTLA-4+
regulatory dendritic cells suppress T-cell response by cytotoxic
T-Lymphocyte antigen-4-dependent IL-10 and
indoleamine-2,3-dioxygenase production in hepatocellular carcinoma.
Hepatology. 59:567–579. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ormandy LA, Hillemann T, Wedemeyer H,
Manns MP, Greten TF and Korangy F: Increased populations of
regulatory T cells in peripheral blood of patients with
hepatocellular carcinoma. Cancer Res. 65:2457–2464. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yang XH, Yamagiwa S, Ichida T, Matsuda Y,
Sugahara S, Watanabe H, Sato Y, Abo T, Horwitz DA and Aoyagi Y:
Increase of CD4+CD25+ regulatory T-cells in the liver of patients
with hepatocellular carcinoma. J Hepatol. 45:254–262. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Fu J, Xu DP, Liu Z, Shi M, Zhao P, Fu B,
Zhang Z, Yang H, Zhang H, Zhou C, et al: Increased regulatory T
cells correlate with CD8 T-cell impairment and poor survival in
hepatocellular carcinoma patients. Gastroenterology. 132:2328–2339.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Gao Q, Qiu SJ, Fan J, Zhou J, Wang XY,
Xiao YS, Xu Y, Li YW and Tang ZY: Intratumoral balance of
regulatory and cytotoxic T cells is associated with prognosis of
hepatocellular carcinoma after resection. J Clin Oncol.
25:2586–2593. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Moreira-Teixeira L, Resende M, Devergne O,
Herbeuval JP, Hermine O, Schneider E, Dy M, Cordeiro-da-Silva A and
Leite-de-Moraes MC: Rapamycin combined with TGF-β converts human
invariant NKT cells into suppressive Foxp3+ regulatory cells. J
Immunol. 188:624–631. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Monteiro M, Almeida CF, Caridade M, Ribot
JC, Duarte J, Agua-Doce A, Wollenberg I, Silva-Santos B and Graca
L: Identification of regulatory Foxp3+ invariant NKT cells induced
by TGF-beta. J Immunol. 185:2157–2163. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Peng G, Wang HY, Peng W, Kiniwa Y, Seo KH
and Wang RF: Tumor infiltrating gammadelta T cells suppress T and
dendritic cell function via mechanisms controlled by a unique
Toll-like receptor signaling pathway. Immunity. 27:334–348. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Pang YL, Zhang HG, Peng JR, Pang XW, Yu S,
Xing Q, Yu X, Gong L, Yin YH, Zhang Y and Chen WF: The
immunosuppressive tumor microenvironment in hepatocellular
carcinoma. Cancer Immunol Immunother. 58:877–886. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Norris S, Collins C, Doherty DG, Smith F,
McEntee G, Traynor O, Nolan N, Hegarty J and O'Farrelly C: Resident
human hepatic lymphocytes are phenotypically different from
circulating lymphocytes. J Hepatol. 28:84–90. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Doherty DG and O'Farrelly C: Innate and
adaptive lymphoid cells in the human liver. Immunol Rev. 174:5–20.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Doherty DG, Norris S, Madrigal-Estebas L,
McEntee G, Traynor O, Hegarty JE and O'Farrelly C: The human liver
contains multiple populations of NK cells, T cells, and CD3+CD56+
natural T cells with distinct cytotoxic activities and Th1, Th2,
and Th0 cytokine secretion patterns. J Immunol. 163:2314–2321.
1999.PubMed/NCBI
|
|
27
|
Norris S, Doherty DG, Collins C, McEntee
G, Traynor O, Hegarty JE and O'Farrelly C: Natural T cells in the
human liver: Cytotoxic lymphocytes with dual T cell and natural
killer cell phenotype and function are phenotypically heterogenous
and include Valpha24-JalphaQ and gammadelta T cell receptor bearing
cells. Hum Immunol. 60:20–31. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kawarabayashi N, Seki S, Hatsuse K, Ohkawa
T, Koike Y, Aihara T, Habu Y, Nakagawa R, Ami K, Hiraide H and
Mochizuki H: Decrease of CD56(+) T Cells and natural killer cells
in cirrhotic livers with hepatitis C may be involved in their
susceptibility to hepatocellular carcinoma. Hepatology. 32:962–969.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Golden-Mason L, Castelblanco N, O'Farrelly
C and Rosen HR: Phenotypic and functional changes of cytotoxic
CD56pos natural T cells determine outcome of acute hepatitis C
virus infection. J Virol. 81:9292–9298. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ye L, Wang X, Wang S, Wang Y, Song L, Hou
W, Zhou L, Li H and Ho W: CD56+ T cells inhibit hepatitis C virus
replication in human hepatocytes. Hepatology. 49:753–762. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Gulubova M, Manolova I, Kyurkchiev D,
Julianov A and Altunkova I: Decrease in intrahepatic CD56+
lymphocytes in gastric and colorectal cancer patients with liver
metastases. APMIS. 117:870–879. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Doskali M, Tanaka Y, Ohira M, Ishiyama K,
Tashiro H, Chayama K and Ohdan H: Possibility of adoptive
immunotherapy with peripheral blood-derived CD3-CD56+ and CD3+CD56+
cells for inducing antihepatocellular carcinoma and antihepatitis C
virus activity. J Immunother. 34:129–138. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liao Y, Wang B, Huang ZL, Shi M, Yu XJ,
Zheng L, Li S and Li L: Increased circulating Th17 cells after
transarterial chemoembolization correlate with improved survival in
stage III hepatocellular carcinoma: A prospective study. PLoS One.
8:e604442013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cai L, Zhang Z, Zhou L, Wang H, Fu J,
Zhang S, Shi M, Zhang H, Yang Y, Wu H, et al: Functional impairment
in circulating and intrahepatic NK cells and relative mechanism in
hepatocellular carcinoma patients. Clin Immunol. 129:428–437. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Nakajima T, Mizushima N and Kanai K:
Relationship between natural killer activity and development of
hepatocellular carcinoma in patients with cirrhosis of the liver.
Jpn J Clin Oncol. 17:327–332. 1987.PubMed/NCBI
|
|
36
|
Kiniwa Y, Miyahara Y, Wang HY, Peng W,
Peng G, Wheeler TM, Thompson TC, Old LJ and Wang RF: CD8+Foxp3+
regulatory T cells mediate immunosuppression in prostate cancer.
Clin Cancer Res. 13:6947–6958. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Peng G, Wang HY, Peng W, Kiniwa Y, Seo KH
and Wang RF: Tumor-infiltrating gammadelta T cells suppress T and
dendritic cell function via mechanisms controlled by a unique
toll-like receptor signaling pathway. Immunity. 27:334–348. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Cao X, Cai SF, Fehniger TA, Song J,
Collins LI, Piwnica-Worms DR and Ley TJ: Granzyme B and perforin
are important for regulatory T cell-mediated suppression of tumor
clearance. Immunity. 27:635–646. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Gavin MA, Rasmussen JP, Fontenot JD, Vasta
V, Manganiello VC, Beavo JA and Rudensky AY: Foxp3-dependent
programme of regulatory T-cell differentiation. Nature.
445:771–775. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Fontenot JD, Gavin MA and Rudensky AY:
Pillars Article: Foxp3 programs the development and function of
CD4+CD25+ regulatory T cells. Nat Immunol 2003. 4: 330–336. J
Immunol. 198:986–992. 2017.PubMed/NCBI
|
|
41
|
Hori S, Nomura T and Sakaguchi S: Pillars
Article: Control of regulatory T cell development by the
transcription factor Foxp3. Science 2003. 299: 1057–1061. Science.
198:981–985. 2017.
|
|
42
|
Kim JY, Kim HJ, Hurt EM, Chen X, Howard OM
and Farrar WL: Functional and genomic analyses of FOXP3-transduced
Jurkat-T cells as regulatory T (Treg)-like cells. Biochem Biophys
Res Commun. 362:44–50. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ziegler SF: FOXP3: Of mice and men. Annu
Rev Immunol. 24:209–226. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Yaqi H, Nomura T, Nakamura K, Yamazaki S,
Kitawaki T, Hori S, Maeda M, Onodera M, Uchiyama T, Fujii S and
Sakaguchi S: Crucial role of FOXP3 in the development and function
of human CD25+CD4+ regulatory T cells. Int Immunol. 16:1643–1656.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Allan SE, Alstad AN, Merindol N, Crellin
NK, Amendola M, Bacchetta R, Naldini L, Roncarolo MG, Soudeyns H
and Levings MK: Generation of potent and stable human CD4+ T
regulatory cells by activation-independent expression of FOXP3. Mol
Ther. 16:194–202. 2008. View Article : Google Scholar : PubMed/NCBI
|