Introduction
Hepatocellular carcinoma (HCC) is the second leading
cause of cancer-associated mortality worldwide (1). Population-based studies have indicated
that the incidence rate continues to approximate the mortality
rate, indicating that HCC accounts for the mortality of a high
proportion of patients who develop it (1). Excessive proliferation of cells is
responsible for the initiation and progression of malignant tumors,
including HCC (2). Therefore,
identifying molecules that suppress proliferation and enhance
apoptosis may provide novel targets for HCC therapies.
MicroRNAs (miRNAs) represent a large family of
endogenous, non-protein-coding RNA molecules that are able to
negatively regulate gene expression at the post-transcriptional
level through translational repression and/or mRNA cleavage
(3). miRNAs are involved in a variety
of biological processes, including proliferation, differentiation,
apoptosis and tumor metastasis, and may function as oncogenes or
tumor suppressor genes by binding to the 3′ untranslated region
(3′-UTR) of target mRNAs (3). Altered
expression of various miRNAs, including miR-122, miR-375, miR-101
and miR-199a-3p, have been observed in cases of HCC, which
indicates an involvement of miRNAs in the pathogenesis of HCC
(4–7).
More extensive investigations are required to elucidate the precise
functions of miRNAs in hepatocarcinogenesis.
Previously, we identified a specific aberrant miRNA
expression profile in HCC by comparison of miRNA expression
profiles in cancerous hepatocytes and normal human hepatocytes.
Among the aberrantly expressed miRNAs in HCC, miR-195 was of
particular interest as its putative targets include astrocyte
elevated gene 1 (AEG-1), which has been reported to serve critical
roles in hepatocarcinogenesis (8).
The aim of the present study was to clarify the functional
significance of miR-195 in HCC and identify cancer-associated
pathways that are regulated by miR-195. The results indicate that
miR-195 is pathologically downregulated in HCC, and that
restoration of miR-195 expression is able to inhibit proliferation
as well as initiate apoptosis of liver cancer cells by targeting
AEG-1, in vitro and in vivo.
Materials and methods
Tissue specimens and cell lines
In total, 36 pairs of HCC and adjacent non-tumor
liver tissues were obtained from patients who were pathologically
diagnosed with HCC and underwent hepatectomy at Tongji Hospital of
Tongji Medical College, Huazhong University of Science and
Technology (Wuhan, China) between September 2009 and September
2012. Tissue samples were immediately snap frozen in liquid
nitrogen until use. The study was approved by the local Research
Ethics Committee at Tongji Hospital of Huazhong University of
Science and Technology, and written informed consent was obtained
from the patients. The characteristics of the included patients are
described in Table I.
 | Table I.Clinical characteristics of patients
with hepatocellular carcinoma included in the present study
(n=36). |
Table I.
Clinical characteristics of patients
with hepatocellular carcinoma included in the present study
(n=36).
| Clinicopathological
parameters | n (%) |
|---|
| Sex |
|
Male | 33 (91.7) |
|
Female | 3 (8.3) |
| Age, years |
|
|
≥40 | 33 (91.7) |
|
<40 | 3 (8.3) |
| Serum alanine
aminotransferase level, U/l |
|
|
<50 | 21 (58.3) |
|
≥50 | 15 (41.7) |
| Hepatitis B virus
surface antigen |
|
|
Positive | 31 (86.1) |
|
Negative | 5 (13.9) |
| Serum α-fetoprotein
level, ng/ml |
|
|
<400 | 20 (55.6) |
|
≥400 | 16 (44.4) |
| Length of tumor,
cm |
|
|
<5 | 3 (8.3) |
| ≥5 | 33 (91.7) |
| New American Joint
Committee on Cancer stage |
|
| Early
(I–II) | 31 (86.1) |
| Late
(IIIA-IIIB) | 5 (13.9) |
The cell lines used in this study included the
immortalized human fetal liver cell line L02, and eight
immortalized human liver cancer cell lines (HepG2, Huh7, PLC/PRF/5,
SMMC-7721, SK-HEP-1, MHCC-97H, MHCC-97L and Hep 3B). All were
maintained in Gibco Dulbecco's modified Eagle's medium (DMEM;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) supplemented with
10% fetal bovine serum (FBS; Gibco; Thermo Fisher Scientific,
Inc.), 100 IU/ml penicillin and 100 mg/ml streptomycin, and
incubated at 37°C with 5% CO2.
RNA isolation
Total RNA was isolated using TRIzol reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol without any prior purification step. RNA
concentrations were determined spectrophotometrically.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Complementary DNA (cDNA) was synthesized from 1 µg
of total RNA using the First Strand cDNA Synthesis kit (Fermentas;
Thermo Fisher Scientific, Inc., Pittsburgh, PA, USA). qPCR was
performed in an Applied Biosystems® 7500 Real-Time PCR
System (Thermo Fisher Scientific, Inc.) using the Platinum SYBR
Green qPCR SuperMix-UDG (Invitrogen; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) according to the manufacturer's protocol.
The following thermocycling conditions were used: For RT, 37°C for
60 min and 70°C for 5 min; and for qPCR, 50°C for 2 min (UDG
incubation), 95°C for 2 min, and 40 cycles of 95°C for 3 sec and
60°C for 30 sec. β-actin was used as an internal control. The
primer sequences for AEG-1 and β-actin are listed in Table II. The expression levels of miR-195
were also analyzed by RT-qPCR using a Bulge-Loop™ miRNA qRT-PCR
Primer Set (Guangzhou RiboBio Co., Ltd., Guangzhou, China) and
normalized to U6. The catalog numbers of the primers for miR-195
and U6 were miRQ0000461-1-2 and MQP-0202 (both from Guangzhou
RiboBio Co., Ltd.). The 2−ΔΔCq method was used for
quantification (9). All reactions
were performed in triplicate.
| Table II.Sequences of the primers used in SYBR
Green reverse transcription-quantitative polymerase chain
reaction. |
Table II.
Sequences of the primers used in SYBR
Green reverse transcription-quantitative polymerase chain
reaction.
| Gene | Primer sequence
(5′-3′) |
|---|
| β-actin |
|
Forward |
CATGTACGTTGCTATCCAGGC |
|
Reverse |
CTCCTTAATGTCACGCACGAT |
| AEG-1 |
|
Forward |
AAATGGGCGGACTGTTGAAGT |
|
Reverse |
CTGTTTTGCACTGCTTTAGCAT |
miRNA mimics and plasmid
transfection
An miR-195 mimic and its matched negative control
(miR-NC) were purchased from Guangzhou RiboBio Co., Ltd. An AEG-1
expressing vector, pcDNA3.1-AEG-1 (which lacked the 3′-UTR of
AEG-1) was prepared as previously described (6). Lipofectamine 2000 reagent (Invitrogen;
Thermo Fisher Scientific, Inc.) was used for cell transfections
according to the manufacturer's protocol. For each transfection, 50
nM miRNA mimics or siRNA were used, unless otherwise noted.
Cell proliferation assays
Huh7 or PLC/PRF/5 cells were plated in 96-well
plates at a density of 3×103 cells per well and
transfected with 50 nM miRNA mimics. In other words, complete
growth medium with 3×103 cells was added to each well
containing miRNA mimics-Lipofectamine 2000 complexes. Cell
proliferation was determined using a Cell Counting Kit-8 (CCK-8;
Promoter Biological Co., Ltd., Wuhan, China) at 0, 24, 48 and 72 h
according to the manufacturer's recommendations. In each treatment
group, at least three wells were measured for cell viability.
Cell apoptosis assays
Huh7 or PLC/PRF/5 cells were transiently transfected
with miRNA-NC or miR-195 mimics and were harvested after 48 h by
trypsinization. Double staining with fluorescein isothiocyanate
(FITC)-annexin V and propidium iodide (PI) was carried out using an
Annexin V-FITC/PI kit (Nanjing KeyGen Biotech Co., Ltd., Nanjing,
China) according to the manufacturer's instructions. Cells were
analyzed by flow cytometry (FACScan; BD Biosciences, Franklin
Lakes, NJ, USA) equipped with CellQuest software version 3.3 (BD
Biosciences). Cells were categorized as viable, dead, early
apoptotic and late apoptotic, and the ratio of total apoptotic
cells in the miR-195-transfected group relative to the total
apoptotic cells in the miR-NC-transfected group was compared.
Experiments were conducted in triplicate.
Analysis of clonogenicity in
vitro
For the colony formation assay, 24 h after
transfection, 1×102 transfected Huh7 cells were plated
in each well of a fresh 24-well plate and maintained in DMEM
containing 10% FBS for 2 weeks. Subsequently, the formed colonies
were fixed with methanol and stained with 0.1% crystal violet in
20% methanol for 10 min.
For the anchorage-independent growth assay,
transfected cells were trypsinized and suspended in complete medium
plus 0.3% low-melting-temperature agarose (Sigma-Aldrich; Merck
Millipore, Darmstadt, Germany). The mixture of cells and agar was
then seeded onto a 0.5% agar base layer with complete medium at a
density of 5×102 cells per well in 24-well plates. After
2 weeks, colonies containing >50 cells were counted. Triplicate
wells were prepared for each group of transfectants and the
experiment was repeated twice.
Target gene prediction for
miR-195
Putative miR-195 target genes and their target
miRNA-binding site seed regions were predicted using the target
prediction programs TargetScan (http://www.targetscan.org/) and microRNA.org (http://www.microrna.org/microrna/home.do).
Western blot
Huh7 cells were harvested 48 h after transfection
and protein lysates were prepared according to the manufacturer's
recommendations. Cell lysates were prepared with 1 ml RIPA Lysis
Buffer (P0013B; Beyotime Institute of Biotechnology, Haimen, China)
with 10 µl phenylmethylsulfonyl fluoride (100 mM) (ST506; Beyotime
Institute of Biotechnology). From each lysate, 30 µg proteins were
separated by 10% SDS-polyacrylamide gel electrophoresis and then
electrophoretically transferred to polyvinylidene fluoride
membranes. Following blocking with 5% nonfat dry milk for 1 h at
room temperature, immunoblotting was performed at 4°C overnight
with a diluted (1:2,000) polyclonal anti-AEG-1 antibody
(#13860-1-AP; Proteintech, Wuhan, China), with a diluted (1:5,000)
anti-GAPDH antibody (#10494-1-AP; Proteintech) used as an internal
control. Subsequently, the membranes were incubated with secondary
antibodies (1:4,000 dilution; #SA00001-2; Proteintech) for 1 h at
room temperature. Band signals were visualized using an enhanced
chemiluminescence kit (Pierce ECL Western Blotting Substrate;
Thermo Fisher Scientific, Inc.).
Luciferase assays
A pMIR-REPORT miRNA Expression Reporter Vector
System (Thermo Fisher Scientific, Inc.) was used to verify the
precise target of miR-195. The wild-type sequences of the AEG-1
3′-UTR (wt-AEG-1) and those with mutated miR-195 target sites
(mut-AEG-1) were synthesized and ligated between the SpeI
and HindIII restriction sites of the pMIR-REPORT Luciferase
vector to establish the pLUC-wt-AEG-1 and pLUC-mut-AEG-1 vectors.
The sequences of oligonucleotides are described in Table III. In 96-well plates, Huh7 cells
were transiently cotransfected with 100 ng of pLUC-wt-AEG-1 or
pLUC-mut-AEG-1, 13 ng of internal transfection efficiency control
pMIR-REPORT β-galactosidase plasmid, as well as miR-195 mimics or
miR-NC to a 50 nM final concentration, using 0.3 µl Lipofectamine
2000. At 24 h after transfection, luciferase and β-galactosidase
activities were measured using a Dual-Light System (Applied
Biosystems; Thermo Fisher Scientific, Inc.).
| Table III.Sequences of 55-mer double-stranded
oligonucleotides containing the predicted miRNA binding sites. |
Table III.
Sequences of 55-mer double-stranded
oligonucleotides containing the predicted miRNA binding sites.
| Targeted gene | Sequence
(5′-3′) |
|---|
| wt-AEG-1 |
|
Forward |
CTAGTGAGCAGAAATTTGGAAGGCTATTCAGTGCTGCTTAGTGTAGCAGCTAATA |
|
Reverse |
AGCTTATTAGCTGCTACACTAAGCAGCACTGAATAGCCTTCCAAATTTCTGCTCA |
| mut-AEG-1 |
|
Forward |
CTAGTGAGCAGAAATTTGGAAGGCTATTCATATTCTTGTAGTGTAGCAGCTAATA |
|
Reverse |
AGCTTATTAGCTGCTACACTACAAGAATATGAATAGCCTTCCAAATTTCTGCTCA |
Lentivirus-based miR-195
overexpression
In order to elucidate the role of miR-195 in
vivo, a recombinant lentivirus (LV-miR-195) was constructed to
generate stable gain-of-function of miR-195 in hepatoma cells. The
recombinant lentivirus LV-miR-195 was prepared as previously
described (10). The negative control
lentivirus was designated LV-miR-NC.
Tumorigenesis assay in nude mice
BALB/c athymic nude mice (male; age, 4–6 weeks;
weight, 16–20 g) were purchased from Beijing HFK Bioscience Co.,
Ltd. (Beijing, China) and were bred in pathogen-free conditions.
The mice were provided with sterile water and food, and were kept
in the following environment: Room temperature, 26–28°C; relative
humidity, 40–60%; and daily light cycle, 10 h light:14 h dark. All
animal experiments were approved by the Tongji Medical College
Institutional Animal Care and Use Committee. Nude mice were
randomly divided into two groups (n=6 for the LV-miR-NC group, and
n=6 for the LV-miR-195 group), and 6×106 cells in
phosphate-buffered saline were inoculated subcutaneously into the
flanks of the nude mice. From the 10th day after inoculation, the
length (L) and width (W) of the tumors were measured with vernier
calipers every 4 days. Tumor volume (V) was calculated with the
formula V = (L × W2) × 0.5. Mice were sacrificed at the
30th day after inoculation.
Statistical analysis
Data are presented as the mean ± standard error from
three separate experiments performed in triplicate, unless
otherwise noted. Error bars indicate standard error. A Student's
t-test was employed to evaluate the differences between groups, and
P<0.05 was considered to indicate a statistically significant
difference.
Results
miR-195 is downregulated in HCC cell
lines and tissues
Previously, we identified a specific aberrant miRNA
expression profile in HCC by comparison of miRNA expression
profiles in cancerous hepatocytes with normal primary human
hepatocytes, and identified miR-195 as one of the most
downregulated miRNAs in HCC (6). In
the present study, RT-qPCR was performed to further verify this
result. First, the expression levels of miR-195 were detected in
eight human hepatoma cell lines, including HepG2, Huh7, PLC/PRF/5,
SMMC-7721, SK-HEP-1, MHCC-97H, MHCC-97L and Hep 3B, as well as in
the normal human liver cell line L02. Generally, a marked reduction
in the expression of miR-195 was observed in the cancer cell lines
compared with the normal liver cell line (Fig. 1A). In addition, among the eight HCC
cell lines, the expression levels of miR-195 in HepG2, Huh7,
PLC/PRF/5, MHCC-97H, MHCC-97L, SK-HEP-1 and Hep 3B were markedly
lower than that of SMMC-7721 (Fig.
1A). This result informs the selection of cell model for use in
further research of miR-195 in HCC.
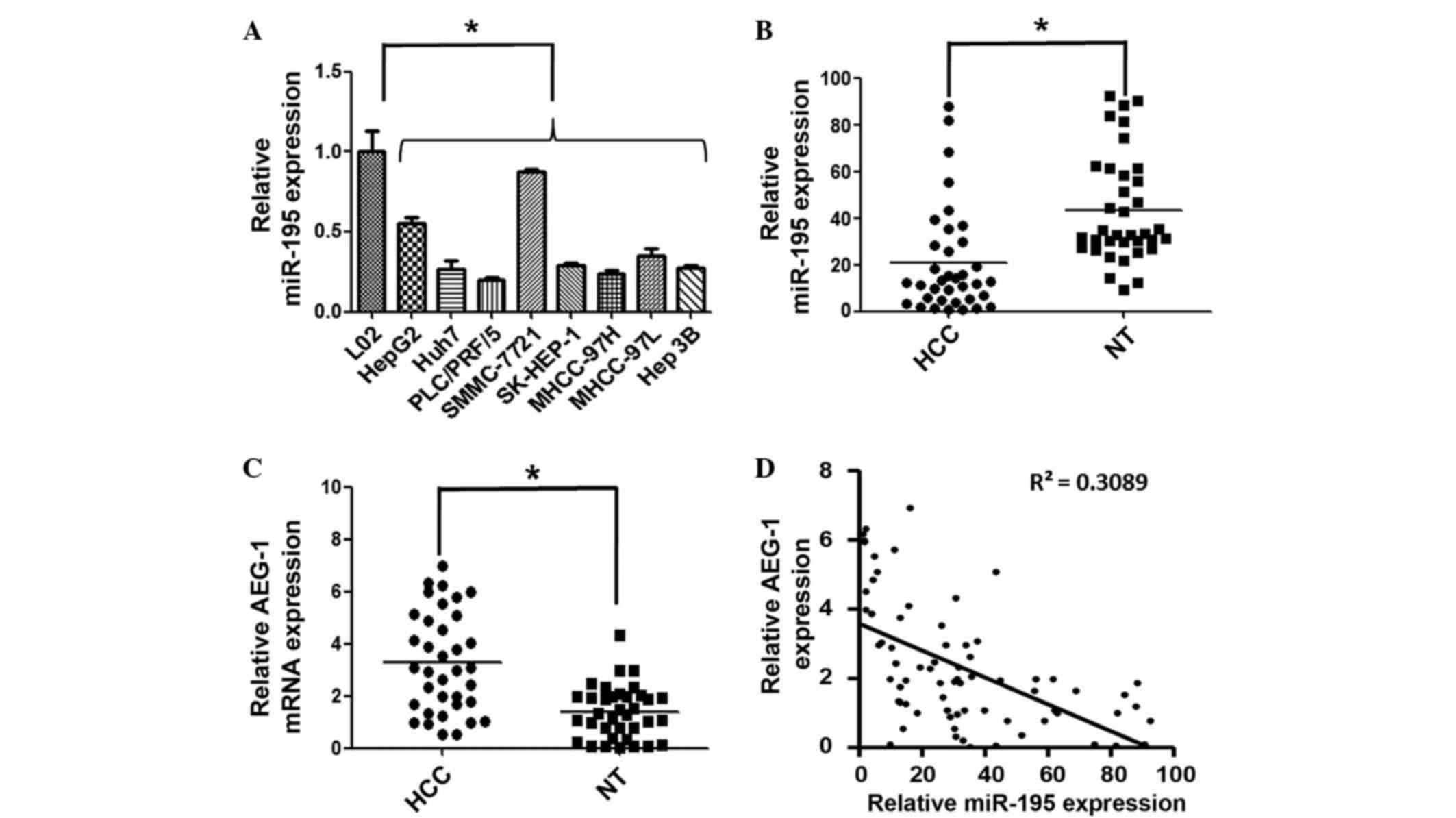 | Figure 1.miR-195 expression is downregulated
and is inversely correlated with AEG-1 expression in HCC. (A)
RT-qPCR was used to measure the expression levels of miR-195 in
eight human liver cancer cell lines (HepG2, Huh7, PLC/PRF/5,
SMMC-7721, SK-HEP-1, MHCC-97H, MHCC-97L and Hep 3B) and a normal
human hepatic cell line (L02). U6 was used as an internal control
and the mean miR-195 expression level in L02 liver cells was
designated as 1. (B) The expression of miR-195 was detected in 36
paired HCC and tumor-adjacent NT samples by RT-qPCR. miR-195 was
downregulated in HCC tissues compared with the adjacent NTs. The
central horizontal line represents the mean value. (C) AEG-1 mRNA
was detected in 36 paired HCC and NT samples by RT-qPCR. β-actin
was used as internal control and the central horizontal line
represents the mean value. (D) miR-195 expression is inversely
correlated with AEG-1 expression (R2=0.308; P<0.01,
Pearson's correlation). *P<0.05. miR-195, microRNA-195; AEG-1,
astrocyte elevated gene 1; HCC, hepatocellular carcinoma; RT-qPCR,
reverse transcription-quantitative polymerase chain reaction; NT,
normal tissue. |
The expression level of miR-195 was further examined
in 36 paired HCC and adjacent noncancerous liver tissues. Compared
with the corresponding noncancerous tissues, the mean relative
expression level of miR-195 was significantly decreased in HCC
samples (P<0.05; Fig. 1B).
Additionally, the expression of miR-195 was inversely correlated
with the expression of AEG-1, which is a potential target for
miR-195 (Fig. 1C and D). These
results indicated that the aberrant expression of miR-195 may be
involved in the process of hepatocarcinogenesis and may provide
insight into pathogenesis of HCC.
miR-195 decreases liver cancer cell
growth and induces apoptosis
Sustained cell growth and proliferation is
considered to contribute to cancer-associated mortality by
disrupting the balance of growth promotion and growth limitation.
To determine the function of miR-195 in HCC, a gain-of-function
analysis was performed using miR-195 mimics or miRNA-NC
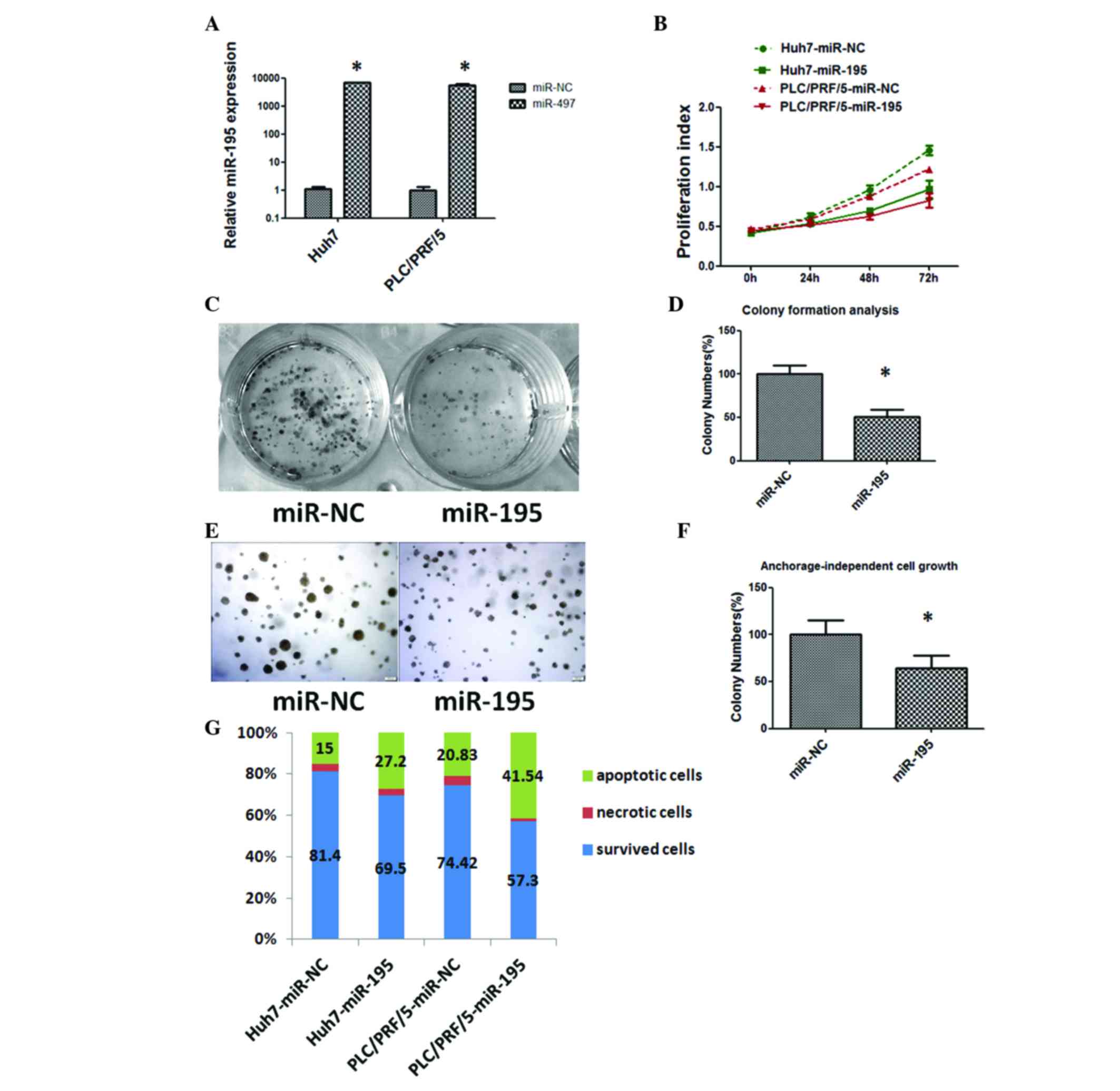
transfectants. As shown in Fig. 2A,
the increased expression of miR-195 was verified by RT-qPCR in Huh7
and PLC/PRF/5 cells transfected with miR-195 mimics.
Sustained cell growth is a notable hallmark of
cancer. The CCK-8 assay showed statistically significant inhibition
of HCC cell proliferation in miR-195 mimic transfectants in
comparison with miR-NC transfectants at 48 and 72 h after
transfection (Fig. 2B). Subsequently,
the capacity of the cells for colony formation and
anchorage-independent growth was evaluated in Huh7 cells
transfected with miR-195 mimics or miR-NC. As shown in Fig. 2C, D, E and F, miR-195
mimic-transfected cells formed markedly fewer and smaller colonies
compared with miR-NC transfectants.
Evading apoptosis is another property of tumor cells
that facilitates their limitless growth. Cell apoptosis in
miR-195-transfected Huh7 and PLC/PRF/5 cells was assessed by flow
cytometry, revealing that the fraction of apoptotic cells was
significantly increased in miR-195 transfectants compared with that
in miR-NC transfectants (P<0.05; Fig.
2G). These data indicate a growth-inhibitory role of miR-195 in
HCC.
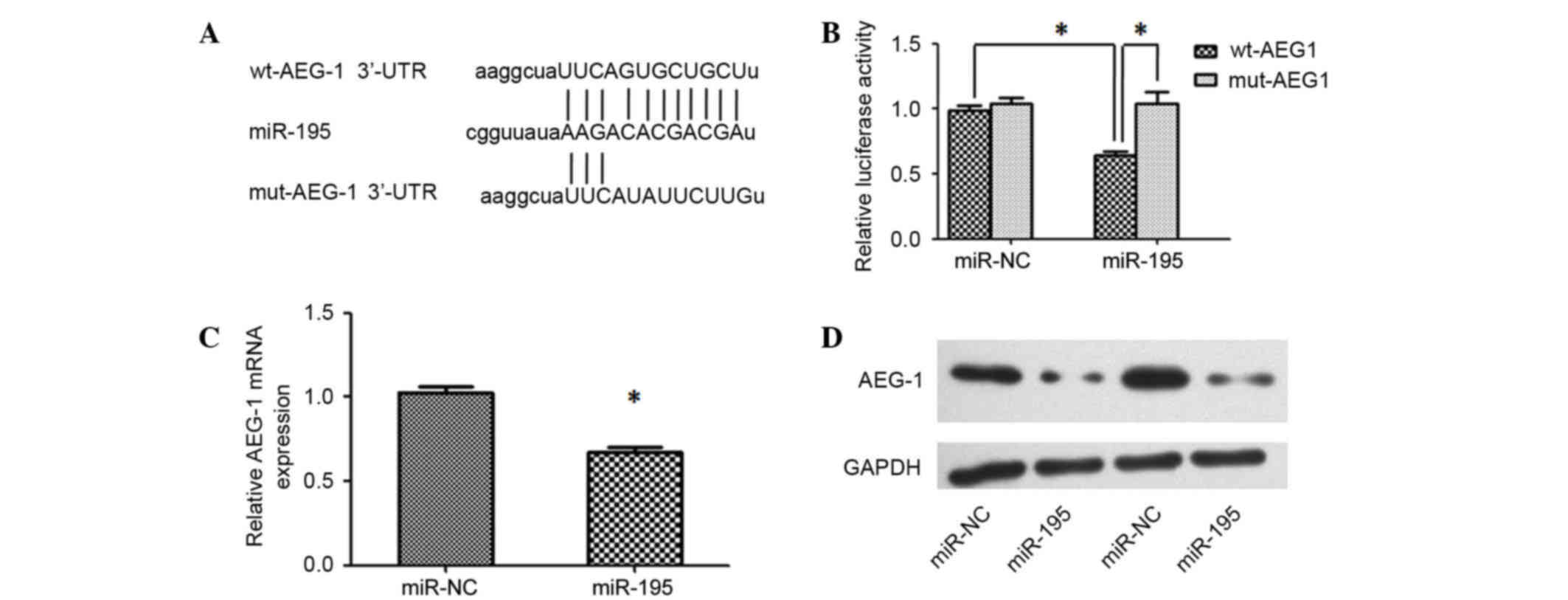
miR-195 directly targets AEG-1 and
blocks the carcinogenic effect of AEG-1 in HCC cells
To unravel the mechanism underlying miR-195-mediated
disruption of proliferation, positive regulators of carcinogenesis
were searched using miRNA target prediction software (TargetScan
and microRNA.org). Among the predicted target genes,
AEG-1 was determined to be of interest as it is reported to have
important roles in hepatocarcinogenesis (8). RT-qPCR was performed to analyze the
expression levels of AEG-1 in HCC tissues and adjacent noncancerous
liver tissues, and the results demonstrated that AEG-1 was
upregulated in HCC tissues (Fig. 1C).
Furthermore, an inverse correlation between the expression levels
of miR-195 and AEG-1 was identified (Fig.
1D). A luciferase reporter analysis, which was performed to
determine whether AEG-1 mRNA contained a target site for miR-195,
revealed that the cotransfection of miR-195 significantly inhibited
the activity of the luciferase reporter construct containing the
wild-type 3′-UTR of AEG-1, whereas this effect was abrogated when
the predicted 3′-UTR binding site was mutated (Fig. 3A and B). Additionally, the expression
levels of AEG-1 mRNA and protein were significantly reduced in Huh7
cells transfected with miR-195 mimics compared with those
transfected with miR-NC (Fig. 3C and
D). These results indicate that the effect of miR-195 is
partially due to its specific and direct interaction with the
putative binding sites in the 3′-UTRs of AEG-1.
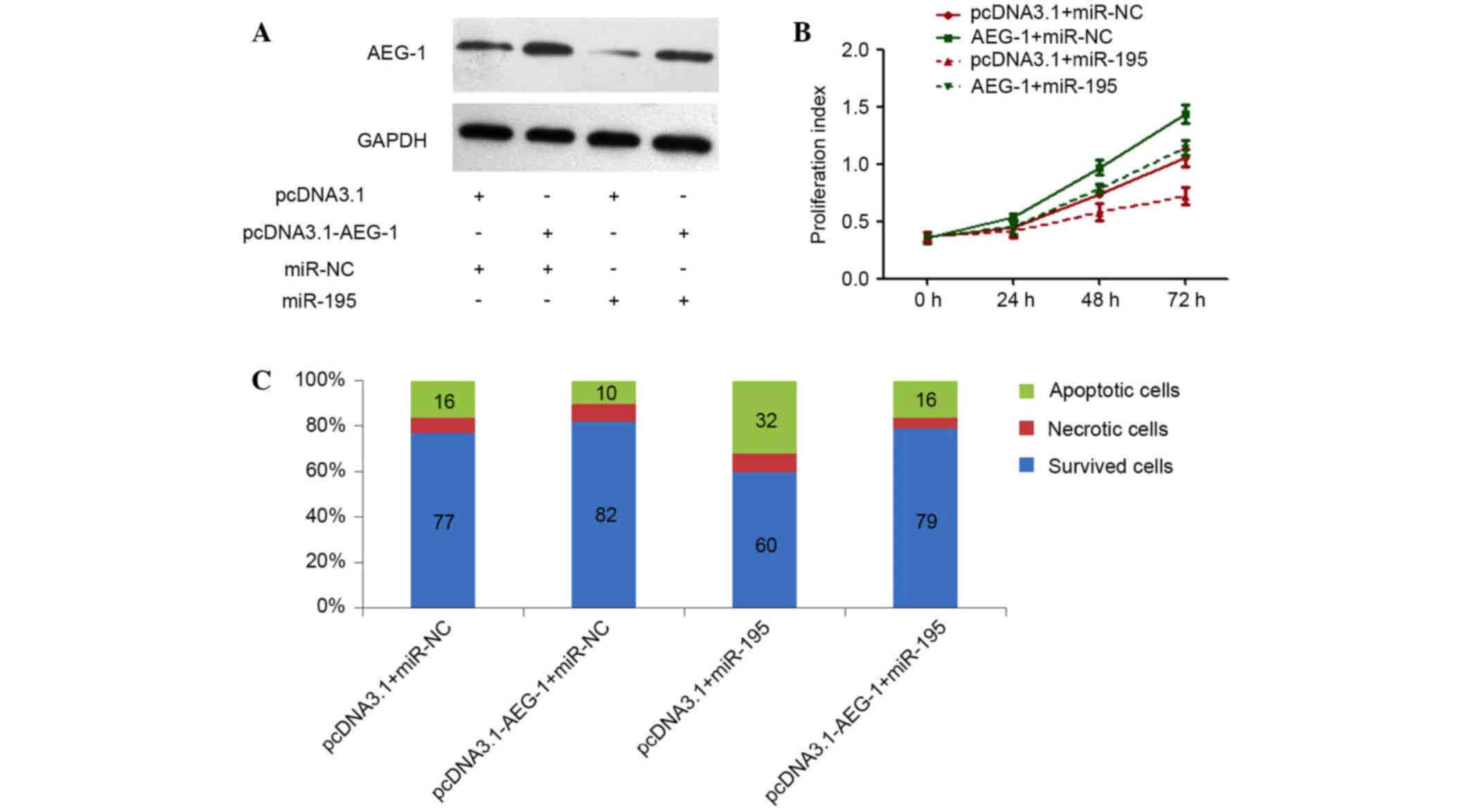
In our previous studies, we have silenced AEG-1 by
RNAi to explore the role of this gene in HCC (6), demonstrating that specific knockdown of
AEG-1 could inhibit cell proliferation and clonogenicity, and also
induce apoptosis (6), which
phenocopied the effects of miR-195 overexpression in HCC cells. In
the present study, whether AEG-1 could counteract the
proliferation-inhibiting and apoptosis-promoting functions of
miR-195 was investigated. An AEG-1 recombinant plasmid was
constructed, which encoded the entire AEG-1 coding sequence but
lacked the 3′-UTR of AEG-1 mRNA. Cotransfection of the AEG-1
plasmid and miR-195 in Huh7 cells efficiently restored AEG-1
expression with enhanced expression of miR-195 (Fig. 4A), and partially reversed the
antitumor effects induced by miR-195 (Fig. 4B and C). Taken together, these results
suggest that miR-195 suppressed AEG-1 expression by binding
directly to the 3′-UTR of AEG-1, and that the negative regulation
of AEG-1 by miR-195 may contribute partially to the antitumor
effects of miR-195 involved in HCC.
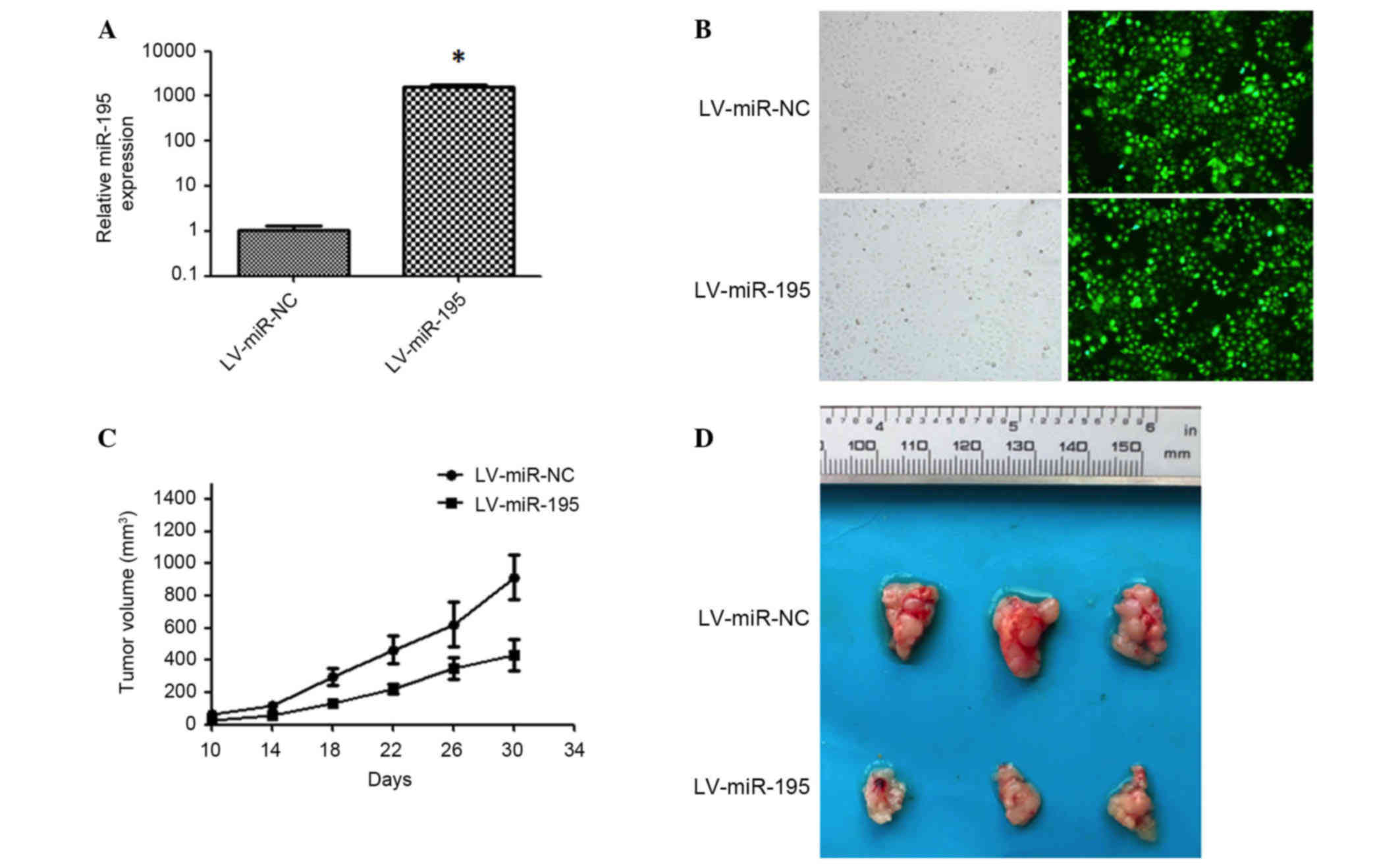
miR-195 inhibits tumor growth of
hepatoma xenografts in nude mice
To investigate the potential antitumor activity of
miR-195 in vivo, the recombinant lentiviral vector
LV-miR-195 and its matched negative control, LV-miR-NC, were
constructed and transduced into Huh7 cells to induce the expression
of miR-195 (Fig. 5A and B). These
lentiviral-transfected Huh7 cells were then injected subcutaneously
into athymic nude male mice (n=6 in each control and experimental
group). Tumor sizes in the two groups were measured every 4 days
from the 10th day after inoculation, and the results indicated that
tumor growth in the LV-miR-195-transfected group was significantly
reduced (Fig. 5C). At 30 days after
inoculation, all mice were sacrificed and the tumor volumes in the
miR-195 group were 47.4% of that of the control (P<0.05;
Fig. 5D). These results suggest that
miR-195 inhibits tumorigenesis in vivo.
Discussion
Accumulating evidence has indicated that miRNAs are
aberrantly expressed in numerous types of human cancer (11). Furthermore, miRNA expression
signatures of HCC have been reported by a number of researchers
(6,12,13). Our
group also reported 37 differentially expressed miRNAs between
cancerous hepatocytes and normal primary human hepatocytes based on
expression profiles (6). miR-195 was
one of the aberrantly expressed miRNAs that was significantly
downregulated in HCC. In the current study, miR-195 was further
confirmed to be significantly suppressed in HCC tissues. These
results are in agreement with those previously reported (14,15). The
detection of miR-195 expression in various cell lines in the
present study indicated that miR-195 was also significantly
suppressed in the 8 hepatoma cell lines compared with the normal
liver cell line. These results provide reference for the selection
of cell models to be used in further research of miR-195 in
HCC.
The mechanism underlying miR-195 dysregulation in
HCC remains elusive. In our previous study, the preliminary data
and bioinformatics analysis suggested that DNA methylation may play
an important and complex role in the regulation of miR-195
expression in HCC, which may provide insight into the mechanism
underlying miR-195 dysregulation in HCC (16).
Sustaining proliferation and evading apoptosis are
hallmarks of malignant tumors, and the aberrant expression of
miRNAs has been implicated in the regulation of various cellular
processes that are often dysregulated during the initiation and
progression of HCC (2,11). For instance, the upregulation of
miR-221 in HCC augments cell proliferation, colony formation and
invasion, and increases the number of cells in S phase, as well as
inhibiting cell apoptosis (17).
miR-199a/b-3p was reported to be downregulated in HCC and
restoration of its expression could target tumor-promoting P21
(RAC1)-activated kinase 4 (PAK4) to suppress HCC growth through
inhibiting the PAK4/Raf/MEK/ERK pathway, in vitro and in
vivo (4). In the present study,
it was demonstrated that miR-195 could inhibit proliferation and
colony-formation by HCC cells, as well as induce apoptosis in
vitro. Furthermore, miR-195 suppressed tumorigenesis of Huh7
subcutaneous xenografts in a nude mouse tumor model. This research
indicate that miR-195 functions as a tumor suppressor in HCC, in
vivo and in vitro.
AEG-1 gene is always amplified in patients with HCC
and serves key role in regulating hepatocarcinogenesis (8). The molecular mechanisms mediating the
oncogenic properties of AEG-1 involve proliferation, apoptosis,
metastasis, angiogenesis and chemoresistance. AEG-1 is a downstream
gene of the oncogenic Ha-Ras pathway, being transcriptionally
regulated by c-Myc upon Ha-Ras and phosphoinositide 3-kinase (PI3K)
activation (8). Overexpression of
AEG-1 activates the PI3K/Akt, nuclear factor-κB, and Wnt/β-catenin
signaling pathways (8). Our data also
documented that AEG-1 was overexpressed in HCC tissues compared
with the corresponding normal liver tissues. Furthermore, an
inverse correlation was observed between miR-195 and AEG-1
expression in HCC tissues for the first time. miR-195 was shown to
suppress AEG-1 expression by directly binding to the 3′-UTR of
AEG-1. To the best of our knowledge, AEG-1 as a direct target gene
of miR-195 has not been reported previously. Furthermore,
restoration of AEG-1 could partially reverse the antitumor effects
induced by miR-195. Theoretically, downregulation of miR-195
should, at least partially, contribute to the increased expression
of AEG-1 in HCC as an alternative mechanism.
The function of miR-195 has already been
investigated in several distinct cancer types, such as non-small
cell lung cancer (18,19), bladder cancer (20,21),
breast cancer (22,23), colorectal cancer (24) and HCC (25–27). To
date, 17 genes, including PCMT1, SRC-3, CCND1, CCNE1, CDC25A,
CCND3, CDK4, CDK6, E2F3, BTRC, IKKα, TAB3, VEGF, VAV2, CDC42, LATS2
and BCL-w, have been identified as targets of miR-195 in HCC
(14,15,25–30).
miR-195 has also been implicated in various biological processes of
HCC, including cell cycle (14,28),
proliferation (25,29), apoptosis (26), drug resistance (30), angiogenesis (15) and metastasis (15,25). The
reported targets of miR-195 and our observations suggest that
miR-195 may regulate multiple signaling pathways, and that loss of
miR-195 could lead to tumor progression in HCC.
In summary, the present study investigated the
potential role of miR-195 in HCC tumorigenesis and found that
miR-195 negatively regulates oncogenic AEG-1 and inhibits hepatoma
cell proliferation and tumor growth in vitro and in
vivo. These data suggest an important role of miR-195 in the
molecular etiology of HCC and indicate a potential application of
miR-195 in cancer therapy.
Acknowledgements
This work was supported by grants from the National
Natural Science Foundation of China (grant nos. 81472832, 81302112
and 81372663).
References
|
1
|
Maluccio M and Covey A: Recent progress in
understanding, diagnosing, and treating hepatocellular carcinoma.
CA Cancer J Clin. 62:394–399. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hung CH, Chiu YC, Chen CH and Hu TH:
MicroRNAs in hepatocellular carcinoma: Carcinogenesis, progression,
and therapeutic target. Biomed Res Int. 2014:4864072014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hou J, Lin L, Zhou W, Wang Z, Ding G, Dong
Q, Qin L, Wu X, Zheng Y, Yang Y, et al: Identification of miRNomes
in human liver and hepatocellular carcinoma reveals miR-199a/b-3p
as therapeutic target for hepatocellular carcinoma. Cancer Cell.
19:232–243. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nakao K, Miyaaki H and Ichikawa T:
Antitumor function of microRNA-122 against hepatocellular
carcinoma. J Gastroenterol. 49:589–593. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
He XX, Chang Y, Meng FY, Wang MY, Xie QH,
Tang F, Li PY, Song YH and Lin JS: MicroRNA-375 targets AEG-1 in
hepatocellular carcinoma and suppresses liver cancer cell growth
in vitro and in vivo. Oncogene. 31:3357–3369. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Xu L, Beckebaum S, Iacob S, Wu G, Kaiser
GM, Radtke A, Liu C, Kabar I, Schmidt HH, Zhang X, et al:
MicroRNA-101 inhibits human hepatocellular carcinoma progression
through EZH2 downregulation and increased cytostatic drug
sensitivity. J Hepatol. 60:590–598. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sarkar D: AEG-1/MTDH/LYRIC in liver
cancer. Adv Cancer Res. 120:193–221. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Livak and Schmittgen: Analysis of relative
gene expression data using real-time quantitative PCR and the
2-ΔΔCt method. Methods. 25:402–408. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Scherr M, Venturini L, Battmer K,
Schaller-Schoenitz M, Schaefer D, Dallmann I, Ganser A and Eder M:
Lentivirus-mediated antagomir expression for specific inhibition of
miRNA function. Nucleic Acids Res. 35:e1492007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hata A and Lieberman J: Dysregulation of
microRNA biogenesis and gene silencing in cancer. Sci Signal.
8:re32015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tao J, Ji J, Li X, Ding N, Wu H, Liu Y,
Wang XW, Calvisi DF, Song G and Chen X: Distinct anti-oncogenic
effect of various microRNAs in different mouse models of liver
cancer. Oncotarget. 6:6977–6988. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Baek S, Cho KJ, Ju HL, Moon H, Choi SH,
Chung SI, Park JY, Choi KH, do Kim Y, Ahn SH, et al: Analysis of
miRNA expression patterns in human and mouse hepatocellular
carcinoma cells. Hepatol Res. 45:1331–1340. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xu T, Zhu Y, Xiong Y, Ge YY, Yun JP and
Zhuang SM: MicroRNA-195 suppresses tumorigenicity and regulates
G1/S transition of human hepatocellular carcinoma cells.
Hepatology. 50:113–121. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang R, Zhao N, Li S, Fang JH, Chen MX,
Yang J, Jia WH, Yuan Y and Zhuang SM: MicroRNA-195 suppresses
angiogenesis and metastasis of hepatocellular carcinoma by
inhibiting the expression of VEGF, VAV2 and CDC42. Hepatology.
58:642–653. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
He XX, Kuang SZ, Liao JZ, Xu CR, Chang Y,
Wu YL, Gong J, Tian DA, Guo AY and Lin JS: The regulation of
microRNA expression by DNA methylation in hepatocellular carcinoma.
Mol Biosyst. 11:532–539. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
He XX, Guo AY, Xu CR, Chang Y, Xiang GY,
Gong J, Dan ZL, Tian DA, Liao JZ and Lin JS: Bioinformatics
analysis identifies miR-221 as a core regulator in hepatocellular
carcinoma and its silencing suppresses tumor properties. Oncol Rep.
32:1200–1210. 2014.PubMed/NCBI
|
|
18
|
Guo H, Li W, Zheng T and Liu Z: MiR-195
targets HDGF to inhibit proliferation and invasion of NSCLC cells.
Tumour Biol. 35:8861–8866. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang X, Wang Y, Lan H and Li J: MiR-195
inhibits the growth and metastasis of NSCLC cells by targeting
IGF1R. Tumour Biol. 35:8765–8770. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Itesako T, Seki N, Yoshino H, Chiyomaru T,
Yamasaki T, Hidaka H, Yonezawa T, Nohata N, Kinoshita T, Nakagawa M
and Enokida H: The microRNA expression signature of bladder cancer
by deep sequencing: The functional significance of the miR-195/497
cluster. PLoS One. 9:e843112014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Fei X, Qi M, Wu B, Song Y, Wang Y and Li
T: MicroRNA-195-5p suppresses glucose uptake and proliferation of
human bladder cancer T24 cells by regulating GLUT3 expression. FEBS
Lett. 586:392–397. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yang G, Wu D, Zhu J, Jiang O, Shi Q, Tian
J and Weng Y: Upregulation of miR-195 increases the sensitivity of
breast cancer cells to Adriamycin treatment through inhibition of
Raf-1. Oncol Rep. 30:877–889. 2013.PubMed/NCBI
|
|
23
|
Li D, Zhao Y, Liu C, Chen X, Qi Y, Jiang
Y, Zou C, Zhang X, Liu S, Wang X, et al: Analysis of MiR-195 and
MiR-497 expression, regulation and role in breast cancer. Clin
Cancer Res. 17:1722–1730. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang L, Qian L, Li X and Yan J:
MicroRNA-195 inhibits colorectal cancer cell proliferation,
colony-formation and invasion through targeting CARMA3. Mol Med
Rep. 10:473–478. 2014.PubMed/NCBI
|
|
25
|
Ding J, Huang S, Wang Y, Tian Q, Zha R,
Shi H, Wang Q, Ge C, Chen T, Zhao Y, et al: Genome-wide screening
reveals that miR-195 targets the TNF-α/NF-κB pathway by
down-regulating IκB kinase alpha and TAB3 in hepatocellular
carcinoma. Hepatology. 58:654–666. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yang X, Yu J, Yin J, Xiang Q, Tang H and
Lei X: MiR-195 regulates cell apoptosis of human hepatocellular
carcinoma cells by targeting LATS2. Pharmazie. 67:645–651.
2012.PubMed/NCBI
|
|
27
|
Amer M, Elhefnawi M, El-Ahwany E, Awad AF,
Gawad NA, Zada S and Tawab FM: Hsa-miR-195 targets PCMT1 in
hepatocellular carcinoma that increases tumor life span. Tumour
Biol. 35:11301–11309. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Furuta M, Kozaki K, Tanimoto K, Tanaka S,
Arii S, Shimamura T, Niida A, Miyano S and Inazawa J: The
tumor-suppressive miR-497-195 cluster targets multiple cell-cycle
regulators in hepatocellular carcinoma. PLoS One. 8:e601552013.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jiang HL, Yu H, Ma X, Xu D, Lin GF, Ma DY
and Jin JZ: MicroRNA-195 regulates steroid receptor coactivator-3
protein expression in hepatocellular carcinoma cells. Tumour Biol.
34:6955–6960. 2014. View Article : Google Scholar
|
|
30
|
Yang X, Yin J, Yu J, Xiang Q, Liu Y, Tang
S, Liao D, Zhu B, Zu X, Tang H and Lei X: MiRNA-195 sensitizes
human hepatocellular carcinoma cells to 5-FU by targeting BCL-w.
Oncol Rep. 27:250–257. 2012.PubMed/NCBI
|