Introduction
Angiogenesis, the process of the formation of new
blood vessels from pre-existing ones, plays a key role in the
progression of solid tumors and hematological malignancies, such as
lymphoma (1). Anti-angiogenesis can
inhibit tumor growth and metastasis, and has become an appealing
strategy for tumor therapy.
Traditional Chinese medicine (TCM) has been used for
several thousand years, and has been suggested to aid the
prevention and treatment of diseases such as cancer. Over the past
40 years, a number of TCM herbs have become a focus of interest
globally; anticancer components have been isolated from medicinal
herbs, which have been demonstrated to serve as modifiers of
biological responses and as inhibitors of angiogenesis (2).
The stem of Marsdenia tenacissima (MT), also
known as Marsdeniae tenacissimae (Roxb) Wight et Arn (family
Asclepiadaceae), is well known as ‘Tong-guan-teng’ in TCM. Over the
past 30 years, the antitumor effects of MT have aroused attention
and passion from researchers. Previous studies have indicated that
MT could inhibit proliferation and induce apoptosis in various
cancer cell lines, such as HepG2/Dox, SGC 7901, Lewis lung
carcinoma, U937, HL60, Jurkat, Raji and RPMI8226 (3–7). Studies
in vitro revealed that MT exerts antitumor activity through
different mechanisms, such as direct inhibition, inducing apoptosis
and differentiation, and immunity adjustments (8). However, to date, few studies have
focused on its antitumor effect in vivo, particularly in
lymphoma cells. It is also unclear whether MT acts through other
specific target(s), such as anti-angiogenesis.
The present study reports the growth inhibitory
effects of MT extract (MTE) against murine B-cell lymphoma in
vivo. Furthermore, the feasibility of MTE for treating lymphoma
via anti-angiogenesis is also investigated.
Materials and methods
MTE (Xiaoiping injection; code, 200709121) was
supplied by Nanjing Sanhome Pharmaceutical Co., Ltd., (Nanjing,
Jiangsu, China). The preparation of plant extracts was performed
according to the Drug Standards of the Ministry of Public Health of
the People's Republic of China (9)
and according to previously reported literature (10). MT bark and kibe powder (20 g with 40
ml 75% ethanol) were refluxed twice for 1.5 h in a hot water bath.
Following vacuum concentration, the filtrate was degreased with 40
ml petroleum ether (60–90°C). The final extracts were partitioned
with n-butanol twice to give n-butanol extract.
Cell lines and culture
Endothelial cells (ECs) were obtained from 10
newborn female C57BL/6 mice (1 week old; ~1.5 g) and were prepared
as previously described (11). A20
cells, the mouse B-cell lymphoma cell line, were obtained from the
Molecular Biology Laboratory of Jiangsu Province Hospital of TCM
(Nanjing, Jiangsu, China). The two cell types were maintained in
RPMI 1640 medium (Gibco; Thermo Fisher Scientific, Inc., Waltham,
MA, USA) containing 10% fetal bovine serum (FBS; Hangzhou Sijiqing
Biological Engineering Materials Co., Ltd., Hangzhou, China) and 1%
penicillin/streptomycin at 37°C in a humidified air atmosphere with
5% CO2.
Animal models and ethics
A total of 20 female BALB/c mice (weighing 18–22 g;
4–6 weeks old), purchased from the College of Veterinary Medicine
of Yangzhou University (Yangzhou, Jiangsu, China), were maintained
in a pathogen-free environment in the Center of Laboratory Animals,
Jiangsu Province Hospital of TCM and were provided with free access
to food and water. All animal experiments were approved by the
Animal Ethics Committee of the Chinese Medicine Hospital of Jiangsu
province, and all mice were handled strictly in accordance with the
National Institutes of Health Guidelines for the Care and Use of
Laboratory Animals.
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide
(MTT) assay
The cytotoxicity of MTE against ECs was determined
by MTT assay. Briefly, the cells were seeded in 96-well plates,
with 1×104 cells/well in 200 µl of RPMI 1640 medium
supplemented with 10% FBS at 37°C in a humidified atmosphere with
5% CO2, and then were either left untreated or were
treated with MTE (0, 6.25, 12.5, 25 and 50 µl/ml). Following
co-incubation for 48 h, 20 µl of MTT solution [10 mg/ml in
phosphate-buffered saline (PBS)] was added to each well for an
additional 4-h incubation. Subsequently, the medium was discarded
and the formed dark-blue formazan crystals in intact cells were
dissolved in 200 µl dimethyl sulfoxide. Absorbance was measured at
490 nm with a microplate reader (Spectra Max 340PC; Bio-Rad
Laboratories, Hercules, CA, USA).
Acridine orange/propidium iodide
(AO/PI) staining
ECs (5×104) treated with indicated
concentrations (0, 6.25, 12.5 and 25 µl/ml) of MTE were incubated
at 37°C with 5% CO2 for 48 h. At the end time point,
each cell culture was centrifuged at 800 × g and 4°C and
rinsed twice with normal saline (NS), and then resuspended in NS
and stained with AO/PI solution (100 µg/ml AO and 100 µg/ml PI).
Images were acquired using a confocal laser scanning microscopy
(LSM510; Zeiss GmbH, Jena, Germany).
Quantification of apoptosis by flow
cytometry
The percentage of apoptotic cells was examined by
double staining with Annexin-V-fluorescein isothiocyanate (FITC)
and PI. Briefly, ~1×105 cells/well, untreated or treated
with MTE (0, 6.25, 12.5 and 25 µl/ml), were seeded into 6-well
plates and incubated at 37°C for 48 h. Subsequently, whole cells
were harvested and treated with 5 µl Annexin V-FITC for 30 min.
Prior to being analyzed by fluorescence-activated cell sorting
(Beckman Coulter EPICS Altra; Beckman Coulter, Inc., Brea, USA), 5
µl of PI was added to each tube.
Animal tumor models and treatment
In total, ~6.0×105 viable A20 cells in
100 µl PBS were used to establish solid tumor models after
subcutaneous injection into the right forelimbs of the BALB/c mice.
At 24 h post-injection, 20 mice were randomly divided into two
groups (10 mice per group), namely the control and MTE groups.
Following the onset of a visible solid tumor, the mice in the MTE
group were administered daily intraperitoneal injections of 300 µl
MTE for 14 consecutive days, and the mice in the control group were
administered 300 µl NS in the same manner. During administration,
subcutaneous injection tumors were measured with Vernier callipers
every 3 days. The tumor volume (V) was calculated according to the
following formula: V = 0.52 × length × width2. A total
of 5 mice from each group were sacrificed by cervical dislocation
24 h after the last administration. The eyeball was removed and
0.8–1.0 ml blood was collected and then centrifuged at 800 ×
g for 15 min at 4°C. The supernatant was collected and
stored at −80°C in a refrigerator for the enzyme-linked
immunosorbent assay (ELISA) of vascular endothelial growth factor,
matrix metalloproteinase (MMP)-2 and MMP-9 suing commercial kits
(cat. nos. EK0955 and EK1302; Elabscience Biotechnology Co., Ltd.,
Wuhan, China). The tumor tissues were dissected and weighed, then
collected for further analysis.
In order to further investigate the anti-lymphoma
activity of MTE in the murine model, the survival times of another
10 mice were recorded (5 mice per group). Side effects during the
experiment, including anepithymia, ruffling of fur, altered mental
state, behavioral changes, reaction to stimulation and weight loss,
were also observed.
For the tumor peritumorous angiogenesis experiments,
the animal tumor model was developed as aforementioned. The
experimental animals were grouped separately and administered with
300 µl MTE or vehicle for 7 days. When a tumor mass was detectable
visually, the mice were sacrificed by cervical dislocation. Tumor
tissues were excised and images of peritumorous blood vessels were
captured.
Microvessel density (MVD)
analysis
Tumor tissues fixed with 10% neutral-buffered
formalin were processed and subjected to histopathological analysis
with hematoxylin and eosin (HE) staining for the measurement of MVD
using a Lawrence and Mayo Lynx Reg microscope, as previously
described (12).
ELISA
The level of serum VEGF was determined by the ELISA,
using specific commercial kits (cat. no. EK0327; Elabscience
Biotechnology Co., Ltd.) according to the manufacturer's protocols.
Briefly, the plasma samples obtained by centrifugation from the
control or MTE-treated mice were pipetted into ELISA plates coated
with polyclonal antibody specific for mouse VEGF (cat. no.
500-P131; 1:250; PeproTech, Inc., Rocky Hill, NJ, USA). The
captured antibody was then detected by a biotinylated monoclonal
antibody against mouse VEGF (cat. no. PL03F1637M; PL Laboratories
Inc., Vancouver, BC, Canada). Bound anti-VEGF was quantified at 450
nm after addition of streptavidin peroxidase and substrate. The
expression of MMP-2 and MMP-9 was detected using the ELISA kit
(Wuhan Boster Biological Technology, Ltd., Wuhan, China) following
the manufacturer's protocols.
Chick chorioallantoic membrane (CAM)
assay
The anti-angiogenesis activity of MTE was further
verified by the CAM assay, which is another reliable model for
angiogenesis studies (13). Embryos
(27 days old) were randomized into two groups: The control and MTE
groups (10 embryos per group). In brief, a pore (~1 mm in diameter)
was punctured in the shell concealing the air sac and 100 µl MTE or
NS was carefully infused into the CAM. Next, the eggs were resealed
with transparent film and incubated for a further 2 days at 37°C in
60% humidity. Subsequent to incubation and treatment with MTE,
images of the blood vessels on the surface of the CAM were captured
using a canon digital camera (Canon, Inc., Tokyo, Japan) in the
treated and untreated eggs.
Statistical analysis
All statistical analyses were performed using the
SPSS 17.0 software package (SPSS, Inc., Chicago, IL, USA). All
data, with the exception of overall survival experiments are
expressed as the mean ± standard deviation. Survival was estimated
using the Kaplan-Meier product limit method. Comparison between
groups was assessed using a one-way analysis of variance followed
by Tukey's multiple comparison test. P<0.05 was considered to
indicate a statistically significant difference.
Results
Anti-proliferative and
apoptosis-inducing effect of MTE on ECs
The effect of MTE on EC viability was assessed using
the MTT assay. As shown in Fig. 1A,
the inhibitory effect of MTE on the ECs occurred in a
concentration-dependent manner. Compared with the control, cell
viability decreased 18.6, 47.5, 85.9 and 97.9%, respectively, in
the 6.25, 12.5, 25 and 50 µl/ml treatment groups. Calculated by the
Bliss method, the half maximal inhibitory concentration
(IC50) value of MTE was 11.91±0.24 µl/ml for the
ECs.
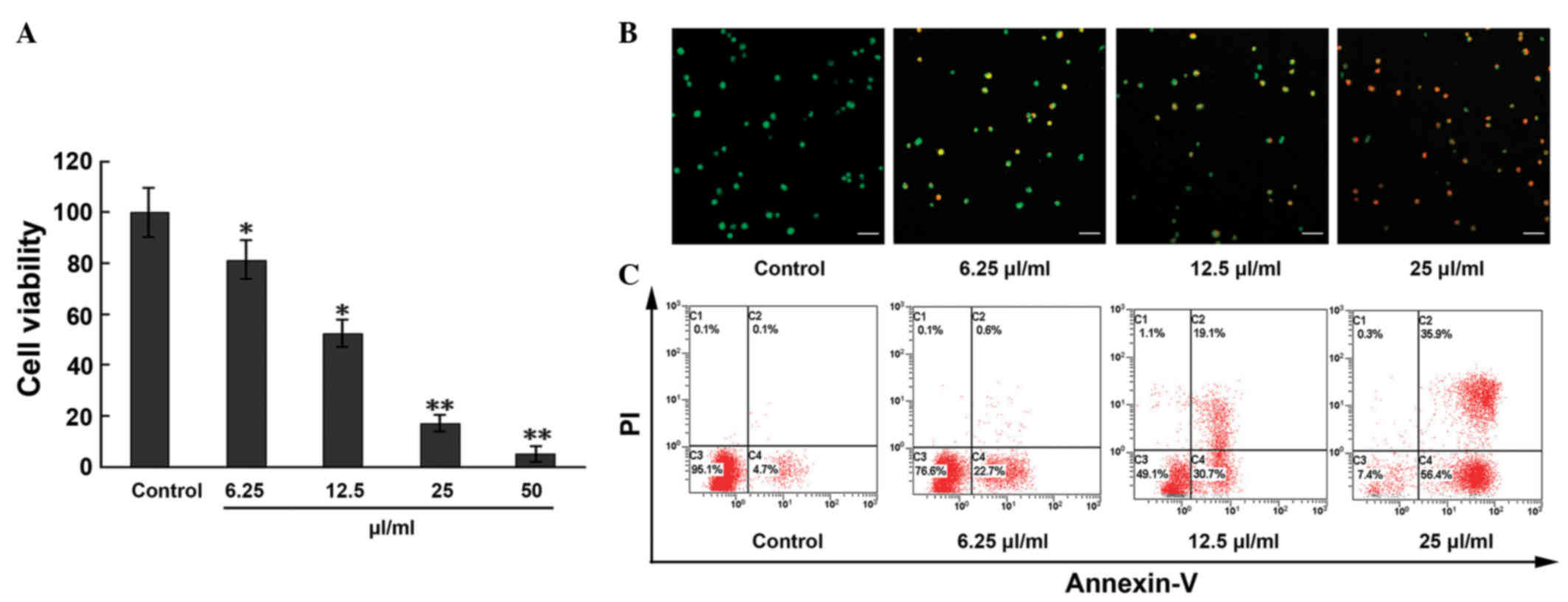 | Figure 1.Apoptosis and anti-proliferative
effects induced by MTE on ECs. (A) ECs were treated with MTE (0,
6.25, 12.5, 25 and 50 µl/ml) for 48 h. The cell viability was
analyzed by the
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide
assay. (B) Following incubation with different concentrations of
MTE (0, 6.25, 12.5, 25 µl/ml) for 48 h, the cells were stained with
acridine orange/PI solutions and observed under a confocal laser
scanning microscope. Green, jacinth and red spots represent living,
apoptotic and necrotic cells, respectively (x200 magnification).
Bar, 10 µm. (C) ECs were incubated with the indicated doses of MTE
for 48 h. After being stained with Annexin V-fluorescein
isothiocyanate and PI, apoptotic cell death (%) was determined with
flow cytometry. Lower left, Annexin V−/PI−
cells (normal); lower right, Annexin V+/PI−
cells (early apoptosis); upper right, Annexin
V+/PI+ cells (late apoptosis); and upper
left, Annexin V−/PI+ cells (necrosis).
Results are represented as the mean ± standard deviation from three
separate experiments. *P<0.05;**P<0.001 vs. control. MTE.
Marsdenia tenacissima extract; EC, endothelial cell; PI,
propidium iodide. |
Based on the results of the cytotoxicity of MTE,
further studies were performed. The morphological examination of
ECs by the AO/PI staining method showed that the apoptosis rate
markedly increased with increasing MTE concentration, with
statistical significance among the indicated concentrations
(Fig. 1B). Similar results were
obtained from the Annexin-V/PI staining assay. In Fig. 1C, compared with the control, the
percentages of ECs undergoing early apoptosis
(Annexin-V+, PI−) and late apoptosis
(Annexin-V+, PI+) in the MTE-treated group
increased in a concentration-dependent manner to 4.8, 23.3, 49.8
and 92.3%, respectively.
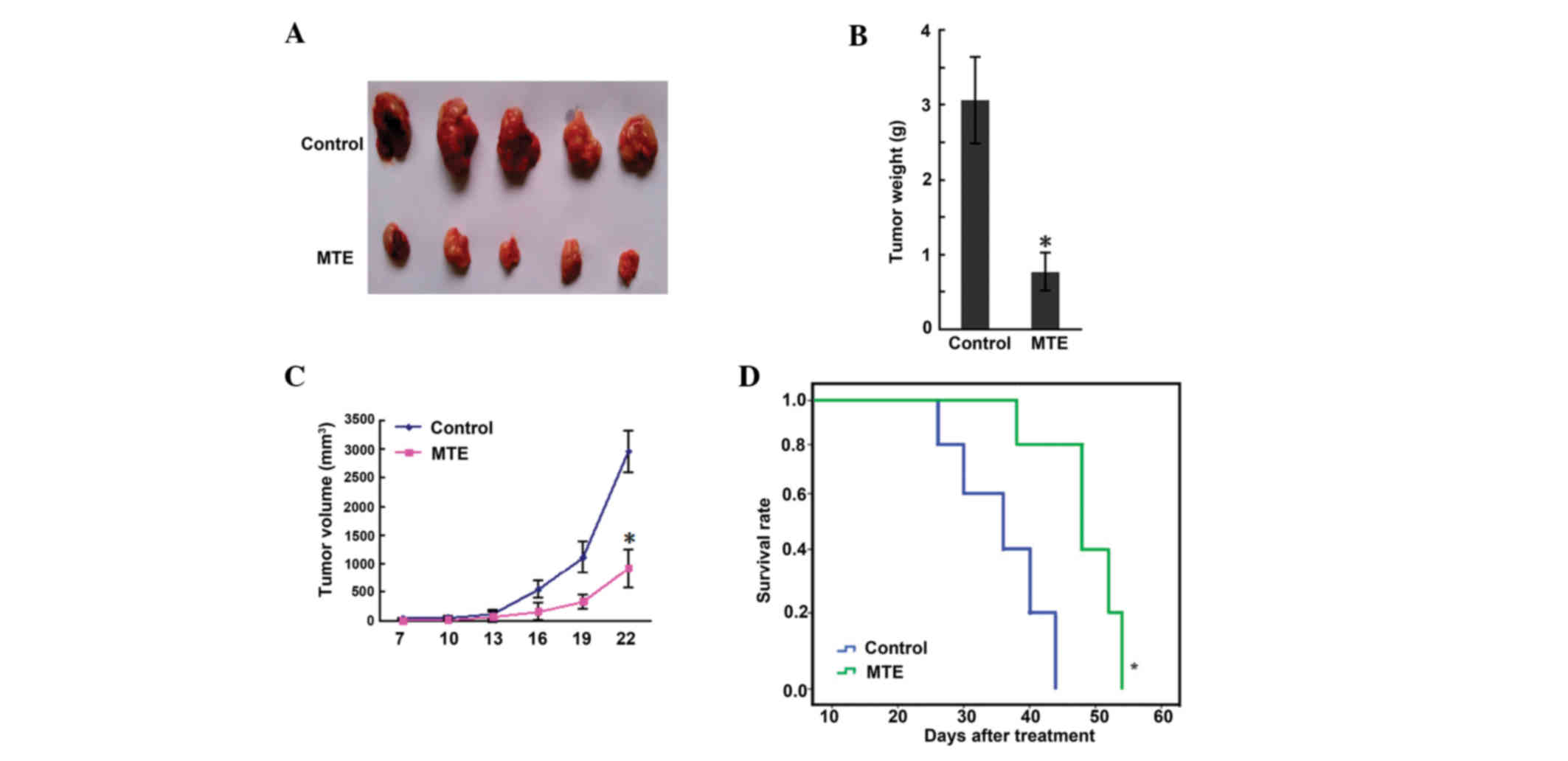
MTE inhibits murine A20 lymphoma
growth in vivo without any side effects
The results of a scatter graph of solid tumor weight
are presented in Fig. 2A. The tumor
weights of the control and MTE groups were 3.06±0.78 and 0.77±0.25
g, respectively, and a significant difference (P<0.01) was shown
between the two groups (Fig. 2B).
Fig. 2C shows that the tumor volume
of the MTE group was smaller than that of the control group
(P<0.05). Furthermore, the survival time in the MTE group was
markedly prolonged compared with that in the control group
(P<0.05) (Fig. 2D). During
administration, no side effects, such as bad appetite, reduced
activity, stunted response, colorless fur or weight loss, were
observed in the MTE group.
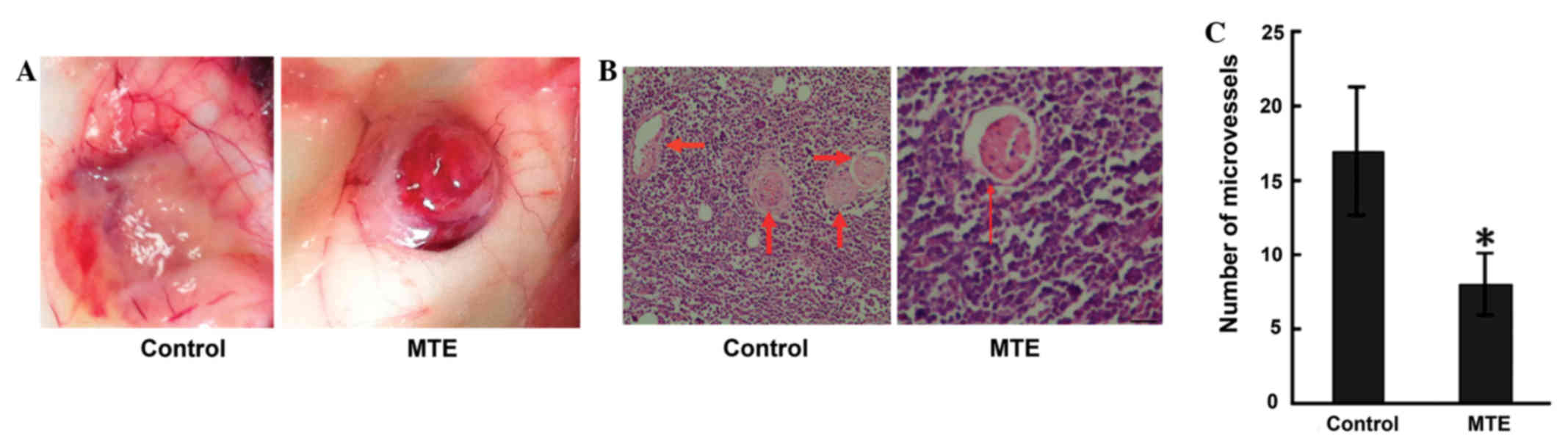
MTE inhibits angiogenesis
Angiogenesis is required for growth, as well as for
the expansion of solid tumors, particularly those at 1–2 mm in
diameter (14). In the present study,
the angiopreventative effect of MTE on the surface of mouse
subcutaneous tumors was verified. Unexpectedly, MTE significantly
reduced peritumorous angiogenesis compared with no treatment in the
tumor-bearing mice (Fig. 3A). Similar
results were obtained from HE staining (Fig. 3B). The MTE-treated mice exhibited a
significantly lower MVD than the control mice (Fig. 3C).
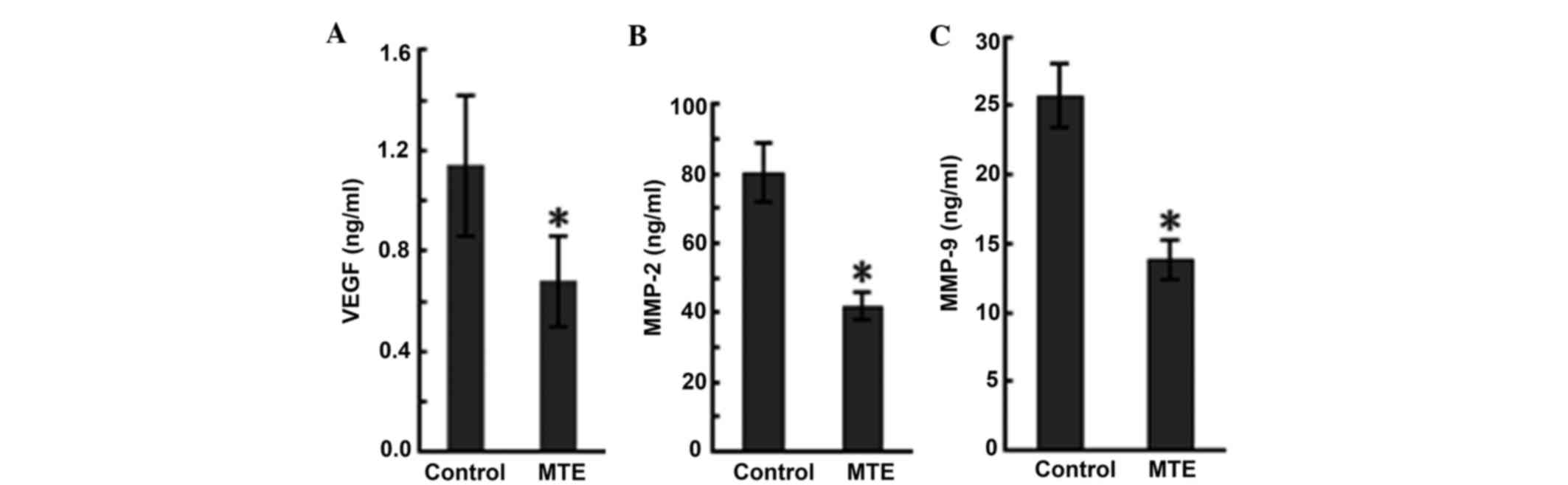
MTE reduces the production of VEGF,
MMP-2 and MMP-9
VEGF is one of the most potent and specific
promoters of angiogenesis, and it acts by stimulating ECs to form
new vessels (15). ELISA was
performed to evaluate the production of VEGF in the tumor-bearing
mice, and the results are presented in Fig. 4A. MTE reduced the VEGF level compared
with the control.
MMPs, a family of at least 24 zinc-dependent
endopeptidases that degrade the basement membrane and all protein
components of the extra-cellular matrix, are hypothesized to play
an essential role in angiogenesis (16). In particular, MMP-2 and MMP-9 play
vital roles in cancer cell invasion and metastasis, as the two MMPs
are abundantly expressed in various malignant cancer cells
(17). In the present study, ELISA
data in serum showed MMP-2 and MMP-9 expression in the MTE group
(42.01±4.12 and 13.87±1.43 ng/ml, respectively), but higher
expression in the control group (80.56±8.64 and 25.73±2.31 ng/ml,
respectively). MTE significantly decreased the levels compared with
the control (Fig. 4B and C).
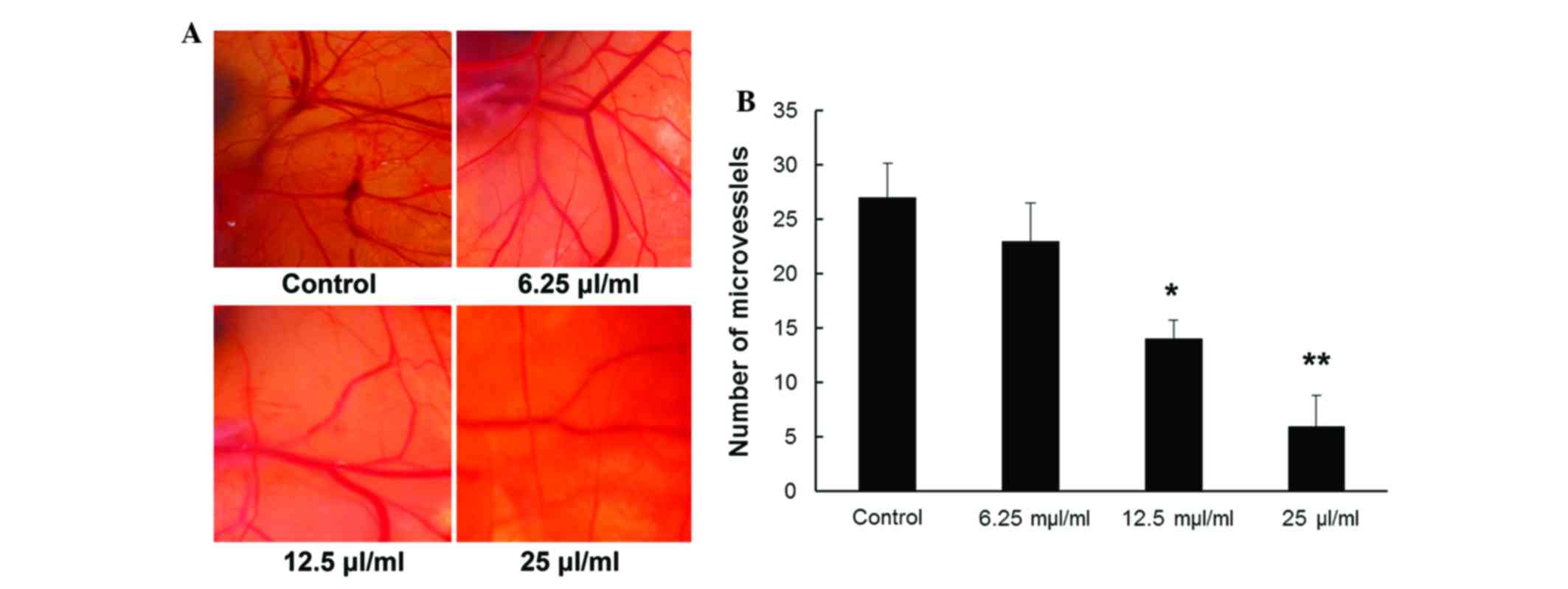
Inhibitory effects of MTE on
angiogenesis in an in vivo CAM assay
A CAM assay was performed to directly examine the
effect of MTE on angiogenesis and vascular development in
vivo. As indicated in Fig. 5A,
compared with the untreated vehicle control, CAM treated with MTE
showed markedly fewer newly formed blood vessels. For quantitative
analysis, new blood vessel branches in a circular area (5 mm in
diameter) surrounding the filter paper disc were counted. Fig. 5B shows that the MVD was
dose-dependently reduced with MTE concentration (P<0.05),
whereas a non-significance difference was observed between the
control and 6.25 µl/ml MTE (P>0.05).
Discussion
The present study showed that MTE suppressed the
growth and induced the apoptosis of ECs in vitro. The in
vivo antitumor effect of MTE against A20 murine B-cell lymphoma
model system was evident by the extended survival times. In
addition, MTE resulted in decreased angiogenesis when compared with
the angiogenesis found on the surface of untreated tumor-bearing
mice. The MVD in the tumor sections of the treated animals was
decreased significantly compared with that of the control group.
Furthermore, a reduction in VEGF, MMP-2 and MMP-9 expression in the
serum was also observed in the treated group. The anti-angiogenesis
activity of MTE was further assessed by the CAM assay.
The solid tumor model system is the most reliable
and representative of major histological types of cancer, thereby
providing the rapid action of drug delivery (18). MVD measurements are widely used as a
surrogate marker in pathological specimens and tumor models in
order to determine the disease prognosis (10,19). VEGF
regulates vascular formation through the initiation of vessel
growth, the incorporation of hematopoietic and endothelial
progenitor cells into the developing vasculature, and the
inhibition of EC apoptosis (20).
VEGF is the well-studied initial factor of angiogenesis (21,22).
Increased MMP expression renders tumor cells capable of digesting
essential tissue barriers, particularly basement membranes that
line blood vessels, thereby promoting cell motility (23). MMP-2 and MMP-9 are also able to
regulate angiogenesis directly (24,25). In
the present study, MTE reduced the density of microvessels in the
implanted A20 tumors and inhibited VEGF expression in the
tumor-bearing mice. The results also indicated that MMP-2 and MMP-9
were important factors involved in the inhibition of angiogenesis
by MTE. This indicates that MTE has potent anti-lymphoma activity
and that the efficacy of anti-angiogenesis may be one of the
possible underlying mechanisms of tumor inhibition. To the best of
our knowledge, this is the first study in which the antitumor
activity and the anti-angiogenesis potency of MTE were evaluated in
a mouse tumor model system.
EC proliferation is the key step for the onset and
progression of angiogenesis (26),
and the induction of EC death could be a strategy for
anti-angiogenic therapies (27). In
the present study, MTE showed promising anti-proliferative potency
against ECs with an IC50 of 11.91±0.24 µl/ml in the MTT
assay. A similar result was found in a recent study, which
demonstrated that MTE was highly effective at inhibiting cell
proliferation in human umbilical vein ECs (HUVECs) by blocking the
cell cycle progression from G1 to S phase (28). In addition, MTE has been shown to
inhibit HUVEC migration and prevent vascular formation through VEGF
suppression in vitro (22).
Notably, the inhibition of EC proliferation in vitro and the
suppression of angiogenesis in vivo appeared to be
associated to the induction of EC apoptosis.
The process of angiogenesis is regulated by a series
of positive and negative factors. Further study is required to
investigate which step of VEGF signal transduction pathways in A20
cells can be taken as the acting target of the antagonists for MTE.
Additional animal experiments and further clinical trials are also
required to understand whether MTE inhibits angiogenesis through
other signaling pathways.
In conclusion, the present study showed that MTE
suppressed tumor growth and reduced angiogenesis in A20 mouse
lymphoma. MTE may represent a natural anti-angiogenic target for
lymphoma therapy.
Acknowledgements
The authors would like to thank Ms. Shuangshuang
Wang, Jiangsu Province Hospital of TCM, for assisting with the
histopathological experiments. In the process of a previous study
on the synergic effect and mechanism on Burrkitt lymphoma induced
by a combination of Xiaoaiping injection and chemotherapeutics
based on Aurora kinase A (batch no, 81603456) supported by the
National Natural Science Foundation, it was observed that decreased
angiogenesis around tumors was shown in MTE treated mice; therefore
the present study was subsequently carried out.
References
|
1
|
Ribatti D, Nico B, Crivellato E, Roccaro
AM and Vacca A: The history of the angiogenic switch concept.
Leukemia. 21:44–52. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sun Y: The role of Chinese medicine in
clinical oncology. Chin J Integr Med. 20:3–10. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hu YJ, Shen XL, Lu HL, Zhang YH, Huang XA,
Fu LC and Fong WF: Tenacigenin B derivatives reverse
P-glycoprotein-mediated multidrug resistance in HepG2/Dox cells. J
Nat Prod. 71:1049–1051. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Li MQ, Shen JH, Xu B, et al: The mechanism
of laboratory research for xiaoaiping treating SGC-7901 gastric
carcinoma cellular strains. J Interventional Radiology. 10:228–231.
2001.(In Chinese).
|
|
5
|
Liang XH, Gao GH, Zhou XL, et al: The
anti-angiogenic effect of Xiaoaiping injection on Lewis lung
carcinoma in C57 mice. Chin Clin Oncol. 15:689–692. 2010.(In
Chinese).
|
|
6
|
Li D, Ou YJ, Li CP, et al: Marsdensia
tenacissima induces apoptosis of human U937, HL60 leukemic
cells. Chin J Biochem Pharm. 29:33–37. 2008.(In Chinese).
|
|
7
|
Chen B, Li CP, Chen JH, et al: Effect of
extract from Marsdenia tenacissima on Jurkat, Raji and
RPMI8226 cells in vitro. Chin J of Biochem Pharmc. 30:174–177.
2009.(In Chinese).
|
|
8
|
Fang YQ and Sun XM: Review on Marsdenia
tenacissima's chemical components and their antitumor
mechanism. Chin J of Biochem Pharmac. 32:165–167. 2011.(In
Chinese).
|
|
9
|
Editorial Committee of the pharmacopoeia
of People's Republic of China: The pharmacopoeia of People's
Republic of China, Part 1. Chemical Industry Press; Beijing, China:
pp. 442–443. 2005, (In Chinese).
|
|
10
|
Yao Q, Lu TL, Mao CQ, et al: Determination
of total saponins and tenac issoside-B in bark and kibe of
Marsdenia tenacissima. Anhui Med and Pharmac J. 14:36–38.
2010.(In Chinese).
|
|
11
|
Zhang M, Tang H, Guo Z, An H, Zhu X, Song
W, Guo J, Huang X, Chen T, Wang J and Cao X: Splenic stroma drives
mature dendritic cells to differentiate into regulatory dendritic
cells. Nat Immunol. 5:1124–1133. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Prabhakar BT, Khanum SA, Shashikanth S and
Salimath BP: Antiangiogenic effect of 2-benzoyl-phenoxy acetamide
in EAT cell is mediated by HIF-1alpha and down regulation of VEGF
of in-vivo. Invest New Drugs. 24:471–478. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Staton CA, Reed MW and Brown NJ: A
critical analysis of current in vitro and in vivo angiogenesis
assays. Int J Exp Pathol. 90:195–221. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Folkman J: Proceeding: Tumor angiogenesis
factor. Cancer Res. 34:2109–2113. 1974.PubMed/NCBI
|
|
15
|
Ferrara N, Gerber HP and LeCouter J: The
biology of VEGF and its receptors. Nat Med. 9:669–676. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jackson C: Matrix metalloproteinases and
angiogenesis. Curr Opin Nephrol Hypertens. 11:295–299. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kang MS, Oh JS, Kang IC, Hong SJ and Choi
CH: Inhibitory effect of methyl gallate and gallic acid on oral
bacteria. J Microbiol. 46:744–750. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Talmadge JE, Singh RK, Fidler IJ and Raz
A: Murine models to evaluate novel and conventional therapeutic
strategies for cancer. Am J Pathol. 170:793–804. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Prabhakar BT, Khanum SA, Jayashree K,
Salimath BP and Shashikanth S: Anti-tumor and proapoptotic effect
of novel synthetic benzophenone analogues in Ehrlich ascites tumor
cells. Bioorg Med Chem. 14:435–446. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ellis LM and Hicklin DJ: VEGF-targeted
therapy: Mechanisms of anti-tumor activity. Nat Rev Cancer.
8:579–591. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Folkman J: What is the evidence that
tumors are angiogenesis dependent? J Natl Cancer Inst. 82:4–6.
1990. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hanada D and Folkman J: Patterns and
emerging mechanisms of the angiogenic switch during tumorigenesis.
Cell. 86:353–364. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Egeblad M and Werb Z: New functions for
the matrix metalloproteinases in cancer progression. Nat Rev
Cancer. 2:161–174. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bergers G, Brekken R, McMahon G, Vu TH,
Itoh T, Tamaki K, Tanzawa K, Thorpe P, Itohara S, Werb Z and
Hanahan D: Matrix metalloproteinase-9 triggers the angiogenic
switch during carcinogenesis. Nat Cell Biol. 2:737–744. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Coussens LM, Fingleton B and Matrisian LM:
Matrix metalloproteinase inhibitors and cancer: Trials and
tribulations. Science. 295:2387–2392. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Risau W: Mechanisms of angiogenesis.
Nature. 386:671–674. 1997. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Folkman J: Angiogenesis and apoptosis.
Semin Cancer Biol. 13:159–167. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Huang Z, Lin H, Wang Y, Cao Z, Lin W and
Chen Q: Studies on the anti-angiogenic effect of Marsdenia
tenacissima extract in vitro and in vivo. Oncol Lett.
5:917–922. 2013.PubMed/NCBI
|