Introduction
Cutaneous hemangioma is one of the most common types
of benign tumor (1). The majority of
types of hemangioma go through three distinct stages, the
proliferation stage, the involuting phase and the involuted phase.
Hemangioma typically presents itself during the proliferative
phase. During the proliferative phase, excessive proliferation of
endothelial cells and nuclear hypertrophy occurs, forming lumens or
lumps without cavities that are similar in size to a small
capillary. The proliferative phase is characterized by rapid
proliferation of capillary endothelial cells and the appearance of
a vascular anomaly. Subsequently, hemangiomas enter an involuting
phase during which the endothelial cells undergo apoptosis, and
extracellular matrix fibers and adipose tissue replace the
hemangioma. This phase can last between a number of months and
several years, following which the hemangioma enters the involuted
phase (2,3).
There are two processes that enable the formation of
new blood vessels, vasculogenesis and angiogenesis. Vasculogenesis
refers to the formation of early blood vessels through the
induction, differentiation and aggregation of endothelial
progenitor cells (4). Angiogenesis
refers to the process of forming new vessels from pre-existing
vessels following birth (5). The
first vessels in the embryo form through vasculogenesis and
following birth angiogenesis becomes the predominant pathway used
to develop blood vessels (6).
Abnormalities in vasculogenesis and angiogenesis can lead to
vascular diseases, of which hemangioma is a common type (7,8).
Folkman (8) reported
that an imbalance in the regulation between angiogenic factors and
angiogenic inhibitors that causes uncontrolled angiogenesis may
result in the occurrence of hemangioma. Therefore, the mechanism of
regulation of angiogenesis serves an important role in the
pathogenesis of hemangioma. A large number of factors that
positively and negatively regulate angiogenesis have different
levels of expression and roles in different stages of the
development of hemangioma (9–12). Previous studies have demonstrated that
multiple angiogenic factors, such as vascular endothelial growth
factor (VEGF), basic fibroblast growth factor (bFGF) and the
estrogen receptor, are highly expressed in the proliferative stage
of hemangioma development (2,13). The number of mast cells is increased
in proliferative hemangioma tissue compared with healthy tissue,
which decreases in the involuting stage and can be restored to
normal levels in the involuted stage (14).
A previous study demonstrated that the proliferation
and apoptosis of endothelial cells serves an important role in the
pathogenesis of hemangioma; the onset and involution of hemangioma
were identified to be associated with endothelial cell
proliferation and apoptosis (15,16). It
has been suggested that the proliferation and apoptosis of vascular
endothelial cells is associated with the onset and spontaneous
regression of hemangioma (17).
Specifically, it is thought that when an increased level of
apoptosis occurs in endothelial cells during the second year of a
child's life, cellular proliferation is offset which results in the
initiation of the spontaneous regression of hemangioma. Activation
of oncogenes and inactivation of tumor suppressor genes can cause
uncontrolled cell proliferation and apoptosis, and induce tumor
formation. The results of a previous study revealed that the
proliferation and apoptosis of vascular endothelial cells were
associated with the regulation of VEGF, transforming growth
factor-β, angiostatin and other cytokines (18,19).
B-cell lymphoma 2 (Bcl-2) belongs to the Bcl-2
protein family and serves an important role in regulating cell
survival and apoptosis (20,21). Tumor protein p53 (p53) is a common
tumor suppressor protein that is frequently activated when the cell
is damaged, and which serves a role in various signaling pathways
to regulate cell survival and death. p53 can affect DNA repair and
cell cycle progression, and serves an important role in the
inhibition of tumorigenesis (22–25). If a
mutation of the gene encoding p53 occurs, the mutated p53 protein
produced cannot trigger apoptosis, resulting in tumorigenesis
(26–28). Although Bcl-2 and p53 have been
studied in multiple tumor types, few studies investigating their
expression in tumor angiogenesis, particularly in the different
stages of hemangioma, have been performed. In the present study,
immunohistochemical analysis and quantum dot technology were used
to measure the expression of Bcl-2 and p53 in the proliferative and
involuting stages of hemangioma, and the blood vessels of healthy
skin, in order to investigate the role of Bcl-2 and p53 during the
proliferative and involuting stages of hemangioma.
Materials and methods
Samples
Formalin-fixed paraffin-embedded samples of archived
cutaneous hemangioma tissue samples collected between January 2007
and December 2010 were obtained from the Department of Pathology of
Renmin Hospital of Wuhan University (Wuhan, China). There were
samples from 38 patients with hemangioma, including 16 males and 22
females, aged between 2 months and 18 years old. There were another
5 samples of healthy skin tissue surrounding the hemangioma. The
locations of the vascular tumors included the following: Scalp,
eyelids, forehead, back of the ears, neck, arms, back, legs, hands
and foot. The patients did not receive any adjuvant therapy prior
to surgery.
Reagents
The Ready-to-use anti-human mouse monoclonal
antibody directed against proliferating cell nuclear antigen
(PCNA), ready-to-use Ultra-Sensitive Immunohistochemical
Streptavidin-Peroxidase (S-P) kit, ready-to-use mouse anti-human
monoclonal antibody directed against p53, DAB Color Developing
Reagent kit and poly-lysine were purchased from Beijing Zhongshan
Golden Bridge Biotechnology Co., Ltd. (OriGene Technologies, Inc.,
Beijing, China). Mouse anti-human monoclonal antibody directed
against Bcl-2 was purchased from Santa Cruz Biotechnology, Inc.
(Dallas, TX, USA). The Quantum Dot Immunofluorescent
Double-Staining Reagent kit was purchased from Wuhan Jiayuan
Quantum Dot Technological Development Co., Ltd. (Wuhan, China; cat
no. K-3001-3) and diethylpyrocarbonate was purchased from Gibco
(Thermo Fisher Scientific, Inc., Waltham, MA, USA).
Classification of specimens
All specimens were stained through routine
hematoxylin and eosin (H&E) staining. H&E staining was
performed following conventional protocol and included dehydration,
permeabilization, paraffin-embedding, sectioning and H&E
staining (29). Neutral formalin
fixed buffer solution (10%) was used for fixing at room temperature
for 24 h. PCNA protein was detected using the ready-to-use
Ultra-Sensitive Immunohistochemical S-P kit, as described below.
Specimens were classified according to Mulliken's criteria
(30) combined with their expression
of PCNA. The staging of specimens was determined according to the
characteristics of their organizational structure. The involuting
stage was defined as hemangioma with fatty infiltration, fibrosis
or vascular lumen occlusion in >20% of the hemangioma. The
proliferative stage was defined as hemangioma without structural
changes. Positive reactions of PCNA were presented as
brownish-yellow particles in the nucleus, as determined by H&E
staining and the immunohistochemistry S-P method. PCNA participates
in DNA synthesis, and its expression has repeatedly been confirmed
to be closely related to cellular proliferation. As a result of its
involvement in cellular proliferation, PCNA can be used in the
diagnosis of malignant tumors such as hemangioma. In the present
study, PCNA expression was observed to be high in proliferative
stage hemangioma tissue, whilst its expression was low in
involuting stage hemangioma tissue. A significant difference was
found in the positive reaction of PCNA expression between
proliferative stage hemangioma tissue and involuting stage
hemangioma tissue.
Detection of Bcl-2-, p53- and
PCNA-associated antigens through H&E staining and the
immunohistochemistry S-P method
Details of the protocol followed have been reported
previously (16). Tissue sections
were cut into 5-µm thick slices and conventionally dewaxed using
xylene for 20 min, anhydrous alcohol for 10 min, and a graded
ethanol series of 90, 80 and 70% alcohol (each for 5 min) and
finally soaked in distilled water twice for 10 min in order to
replace the paraffin in samples with water. Sections were then
washed and incubated with distilled 3% hydrogen peroxide for 10 min
at room temperature, in order to eliminate endogenous peroxidase
activity. Specimens were washed with distilled water and soaked in
PBS for 5 min. For antigen retrieval, slices were placed in 0.01 M
citrate buffer (pH 6.0), boiled (at 95°C for 15–20 min), and left
to cool for 20 min. Sections were then washed with cold water in
order to cool samples to room temperature, and finally washed with
PBS 3x for 5 min each. Following antigen retrieval, tissue sections
were blocked with 5–10% normal goat serum (Sigma-Aldrich; Merck
Millipore, Darmstadt, Germany; diluted in PBS), incubated at room
temperature for 10 min and washed with PBS 3 times (5 min/wash).
Following the discard of excessive serum, the following primary
antibodies were added: Bcl-2 (dilution, 1:100; cat. no. sc-7382;
Santa Cruz Biotechnology, Inc., Dallas, TX, USA), p53 (dilution,
1:100, cat.no. ZM-0408; Beijing Zhongshan Golden Bridge
Biotechnology Co., Ltd.; OriGene Technologies, Inc.) and PCNA
(dilution, 1:100; cat. co. ZM-0213; Beijing Zhongshan Golden Bridge
Biotechnology Co., Ltd.; OriGene Technologies, Inc.). Sections were
then incubated with these antibodies at 4°C overnight. The tissue
sections were then incubated with primary antibodies for 1 h at
37°C, and washed with TBS-Tween® 20 (TBST) 3 times for a
total of 5 min each. A total of 150 µl biotin-labeled secondary
antibody (cat. no. SPN-9001; Beijing Zhongshan Golden Bridge
Biotechnology Co., Ltd.; OriGene Technologies, Inc.) was diluted
with 1% bovine serum albumin (BSA; cat. no. ST023)-PBS and added to
each of the sections, which were then incubated at 37°C for 10–30
min. BSA was supplied by Beyotime Institute of Biotechnology
(Haimen, China). Sections were then washed 3 times with TBST for 5
min each time. A total of 150 µl streptavidin-labeled horseradish
peroxidase-conjugated secondary antibody (diluted with 1% PBS; cat.
no. V7023; Beijing Zhongshan Golden Bridge Biotechnology Co., Ltd.;
OriGene Technologies, Inc.) was then added to the first stain
solution (DAB Color Developing Reagent kit; cat. no. ZLI-9018;
Beijing Zhongshan Golden Bridge Biotechnology Co., Ltd.; OriGene
Technologies, Inc.) and incubated with the sections at 37°C for
10–30 min. Specimens were washed with tap water, counterstained
with hematoxylin and mounted using neutral gum/aqueous mounting
medium (cat. no. ZLI-9550; Beijing Zhongshan Golden Bridge
Biotechnology Co., Ltd.; OriGene Technologies, Inc.) onto the
slides.
Detection of Bcl-2, p53 and PCNA
through quantum dot double staining
Details of the protocol followed have been reported
previously (16). The
paraffin-embedded tissue sections were cut into 3–4-µm slices;
heated, dewaxed and hydrated. Following microwave antigen
retrieval, specimens were washed with TBS. Subsequently, tissue
sections were blocked with Immunol Staining blocking buffer (cat.
no. P0102; Beyotime Institute of Biotechnology, Shanghai, China)
and incubated in a wet box for 30 min at 37°C. The Bcl-2, P53 and
PCNA primary antibodies were incubated at 37°C for 2 h and
subsequently washed with TBST 3 times (5 min/wash). Specimens were
blocked with Immunol Staining blocking buffer (cat. no. P0102;
Beyotime Institute of Biotechnology) and incubated in a wet chamber
for 20 min at 37°C. Then biotin-labeled secondary antibody (cat.
no. YM005; dilution, 1:100; Quantum Dot Immuno-Fluorescent Double
Staining reagent kit) was added to the sections, which were
incubated at 37°C for 30 min. Drops of undiluted
streptavidin-coated quantum dots (QDs-SA; dilution, 1:50-200; final
concentration, 5–20 nmol/l; Quantum Dot Immuno-Fluorescent Double
Staining reagent kit) were added to specimens, which were then
incubated in a wet box at 37°C for 30 min and washed with TBST 3
times, (5 min/wash). Following air-drying, tissue samples were
mounted on slides using 90% glycerin buffer as the mounting medium.
Specimens were visualized using an Olympus IX71 fluorescence
microscope (Olympus Corporation, Tokyo, Japan). A wavelength of 545
nm was used to excite the QDs with ultraviolet light, and a
wavelength of 605 nm was used to excite QDs with blue light.
Analysis of the results of
immunohistochemical and quantum dot staining
Cells were considered Bcl-2-positive if brown-yellow
granules were identified in the cytoplasm and/or the cell membrane.
Cells were considered PCNA- or p53-positive if brown-yellow
granules appeared in the nucleus. No brown-yellow granules were
identified in the cytoplasm or nucleus of the negative control
group.
Bcl-2 and p53 expression was quantitatively analyzed
through a Nuance FX Multispectral Imaging system (Cambridge
Research & Instrumentation, Inc., Woburn, MA, USA). For each
section, five complete and non-overlapping high magnification
(x400) fields were randomly selected for each sample. The average
optical density, area of positive staining and total area of all
cells in each field was measured, and used to calculate the
positive area rate. Positive area rate was calculated as follows:
(Total area of positive staining/the total area of cells)
×100%.
Statistical analysis
Quantum dot staining results are illustrated as the
mean ± standard deviation. One-way analysis of variance and the
Student-Newman-Keuls q post hoc test were used to determine the
significance of differences in the average optical density and
positive area rate between groups. A significance level of α=0.05
was selected. Normality test and variance homogeneity test were
conducted prior to testing. Each group met the requirements of
normal distribution and homogeneity of variance. All tests were
performed using SPSS software (version 13.0; SPSS, Inc., Chicago,
IL, USA). P<0.05 was considered to indicate a statistically
significant difference.
Results
H&E staining
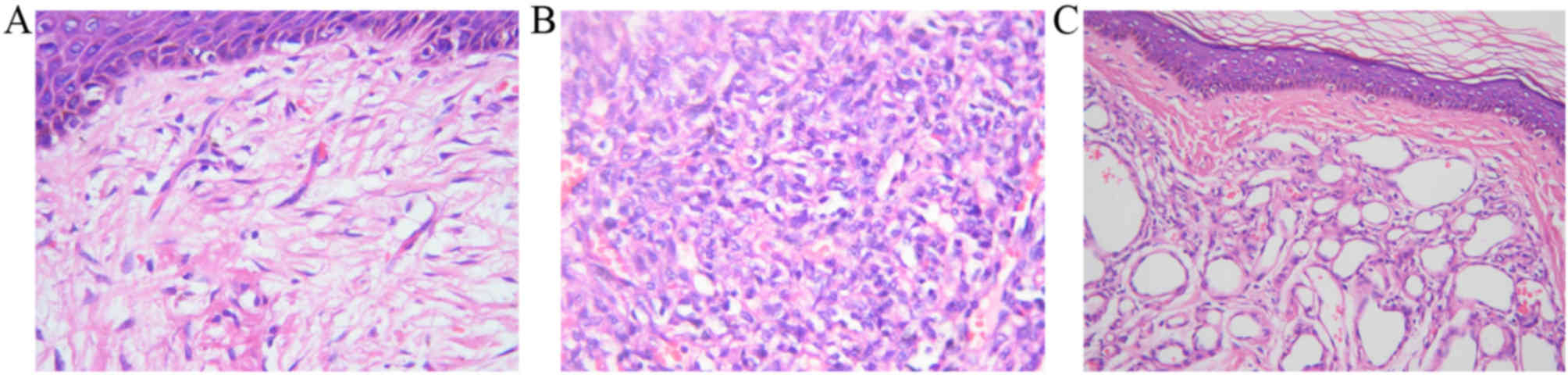
The capillaries in the healthy skin tissue samples
had thin walls and were surrounded by 1–2 endothelial cells, which
were primarily thin but were thicker around the nuclei (Fig. 1A). In addition, the nuclei of the
endothelial cells were flat compared with other cell types
(Fig. 1A). A large number of active
proliferative endothelial cells shaped like a cord or mass were
observed to be gathered in proliferative hemangioma tissue samples
(Fig. 1B). There were also irregular
gaps in the sinusoid capillaries between endothelial cells, the
vessel lumen and large weakly stained nuclei (Fig. 1B). In involuting hemangioma tissue
samples, the number of endothelial cells surrounding the
capillaries was decreased, vascular lumens were enlarged, and
fibrosis of the blood vessels, fatty deposits, lumen occlusion and
flat endothelial cell nuclei were present (Fig. 1C).
Detection of PCNA through the
immunohistochemistry S-P method
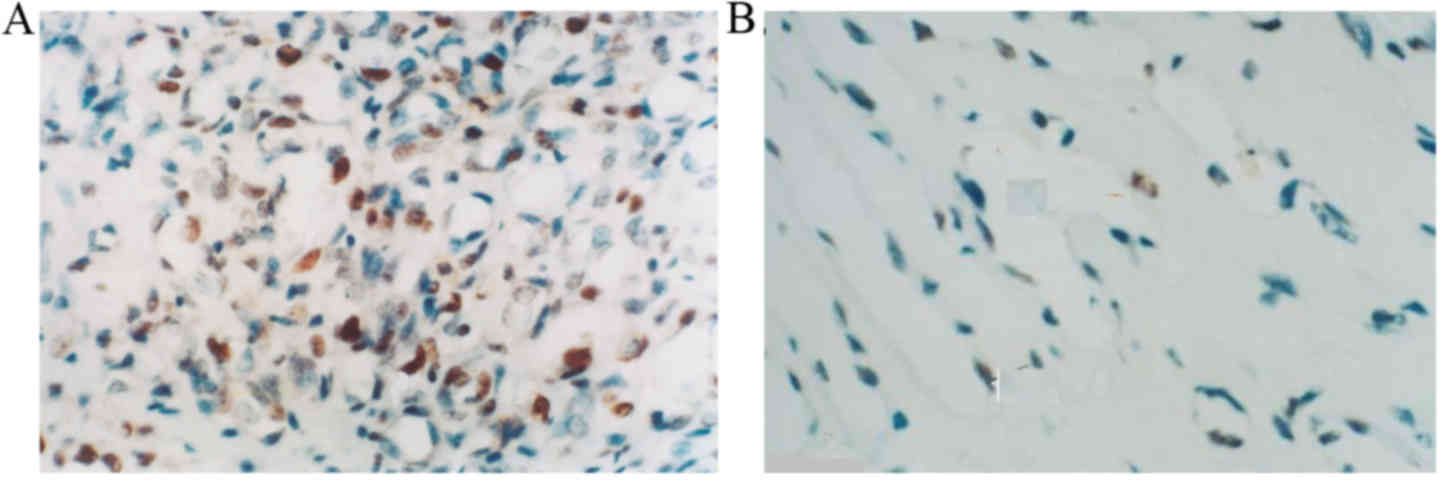
Cells were considered PCNA-positive if brown-yellow
granules were distributed diffusely in the nuclei of endothelial
cells. In proliferative hemangioma tissue samples, the nuclei of
endothelial cells were large and PCNA expression was high (Fig. 2A). In the involuting hemangioma tissue
samples, the nuclei of endothelial cells were flat, there were few
nuclei and nuclei contained a small amount of brown-yellow
granules, indicating that PCNA expression was low (Fig. 2B).
H&E staining and the
immunohistochemistry S-P method were used to detect PCNA
Classification indicated 24 cases of proliferative
hemangioma and 14 cases of involuting hemangioma. In addition, 5
cases had healthy skin surrounding the hemangioma and were thus
used as the control group.
Detection of Bcl-2 and p53 through the
immunohistochemistry S-P method
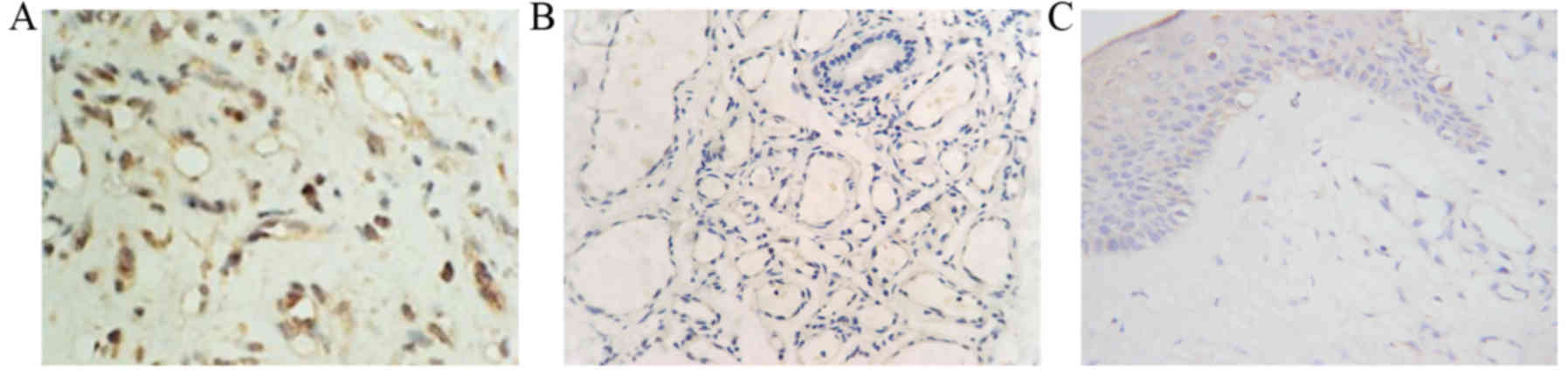
Numerous brown-yellow granules were identified in
the endochylema of endothelial cells in proliferative hemangioma
tissue, indicating that Bcl-2 expression was high (Fig. 3A). No brown-yellow granules were
identified in the endochylema of endothelial cells in involuting
hemangioma samples (Fig. 3B) or cells
in healthy skin tissue samples (Fig.
3C), suggesting Bcl-2 was not expressed.
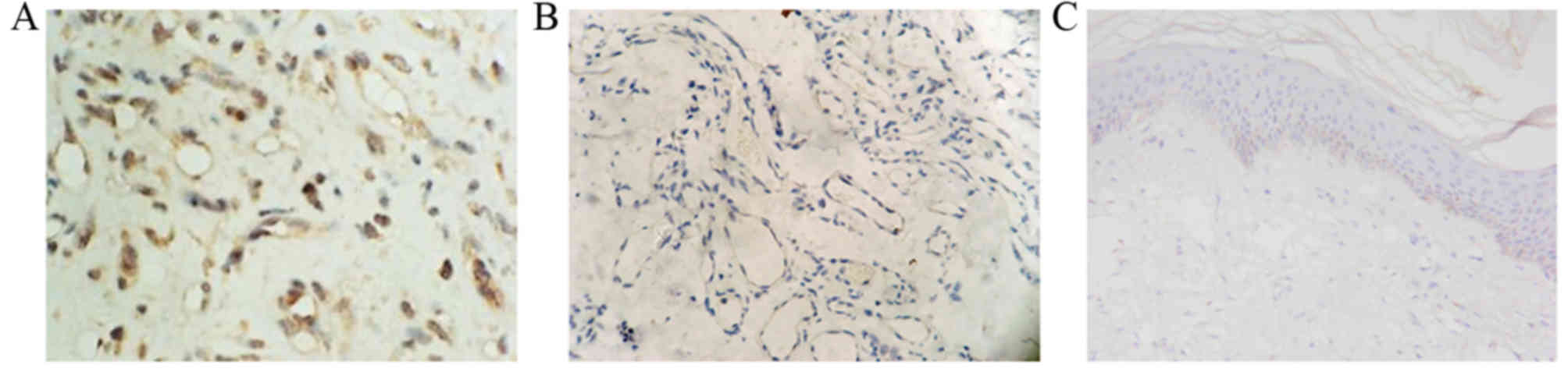
Brown-yellow granules were observed in the
endochylema of endothelial cells in proliferative hemangioma
tissue, indicating that p53 expression was high (Fig. 4A). No brown-yellow granules were
identified in the endochylema of endothelial cells in involuting
hemangioma tissue samples (Fig. 4B)
or healthy skin tissue samples, revealing weak or no p53 expression
(Fig. 4C), suggesting little or no
p53 expression.
Detection of Bcl-2, p53 and PCNA
through the quantum dot double staining method
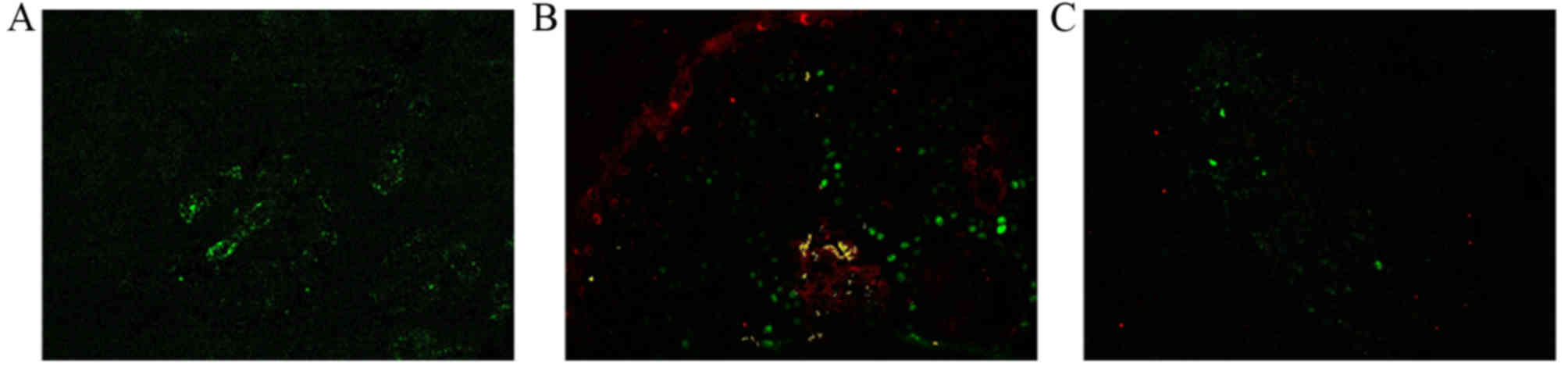
The presence of PCNA was visible as green
fluorescence, and the presence of Bcl-2 was visible as red
fluorescence (Fig. 5). No Bcl-2
expression was identified in the vascular tissue cells of healthy
skin specimens (Fig. 5A). Bcl-2 was
highly expressed in the endochylema and cytomembrane of cells in
proliferative hemangioma tissue samples (Fig. 5B). Conversely, Bcl-2 and p53
expression was low in the endochylema and cytomembrane of cells in
involuting hemangioma tissue samples (Fig. 5C).
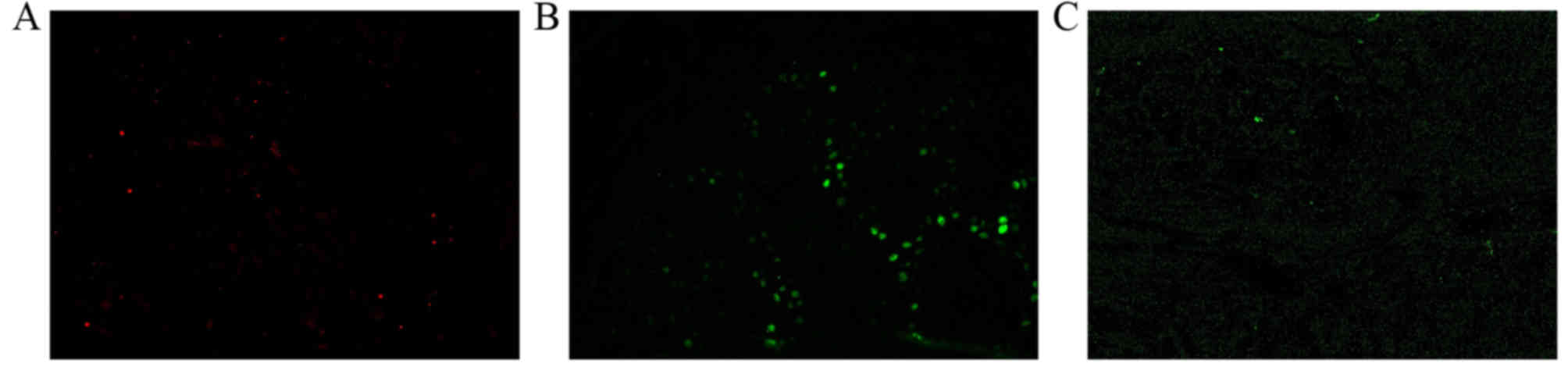
The presence of p53 was visible as green
fluorescence in a separate experiment. No p53 expression was
observed in the vascular tissue cells of healthy skin tissue
(Fig. 6A). However, p53 was highly
expressed in the nuclei of proliferative hemangioma cells (Fig. 6B). Conversely, p53 expression was low
in the nuclei of involuting hemangioma cells (Fig. 6C).
Statistical analysis of the detection
of, Bcl-2, p53 and PCNA through the quantum dot double staining
method
As illustrated in Tables
I and II, significant
differences in the average optical density and positive area rate
of Bcl-2 and p53, respectively, were identified in proliferative
stage hemangioma tissue samples compared with healthy tissue
samples (P<0.05). By contrast, no significant differences in the
average optical density and positive area rate of Bcl-2 and p53
were identified between involuting stage hemangioma tissue samples
and healthy surrounding skin tissue samples (all P>0.05).
 | Table I.Average optical density and positive
area rate of B-cell lymphoma 2 in different hemangioma stages
determined through quantum dot double staining. |
Table I.
Average optical density and positive
area rate of B-cell lymphoma 2 in different hemangioma stages
determined through quantum dot double staining.
| Sample | No. of cases | No. of
sections | Average optical
density | Positive area
rate |
|---|
| Proliferative
hemangioma | 24 | 145 |
0.3941±0.0210a |
0.4012±0.0227a |
| Involuting
hemangioma | 14 | 115 |
0.1749±0.0195b |
0.1573±0.0074b |
| Healthy surrounding
skin | 5 | 35 |
0.1704±0.0184c |
0.1486±0.0057c |
| Table II.Average optical density and positive
area rate of tumor protein p53 in different hemangioma stages
determined through quantum dot double staining. |
Table II.
Average optical density and positive
area rate of tumor protein p53 in different hemangioma stages
determined through quantum dot double staining.
| Sample | No. of cases | No. of
sections | Average optical
density | Positive area
rate |
|---|
| Proliferative
hemangioma | 24 | 145 |
0.4735±0.0307a |
0.3987±0.0304a |
| Involuting
hemangioma | 14 | 115 |
0.1781±0.0214b |
0.0954±0.0059b |
| Healthy surrounding
skin | 5 | 35 |
0.1760±0.0197c |
0.0902±0.0051c |
Discussion
Infantile cutaneous hemangioma is a common type of
benign tumor, and is characterized by the rapid growth of
endothelial cells at birth, followed by a slow involuting phase
that lasts for a number of years, and this may lead to the tumor
completely disappearing (16).
Vascular endothelial cells serve an important role in the
development of hemangioma through the modulation of the expression
of a series of positive and negative regulators (9–11). It is
typically understood that the proliferation and regression of
vascular endothelial cells is the primary reason for the growth and
involution of hemangioma (31).
Apoptosis is the process of spontaneous cell death through gene
regulation in order to regulate the development of organisms and
maintain a stable environment (32).
Under certain conditions, the activation of enzymes, such as
endogenous DNA incision enzymes, can induce cell death through a
series of signaling cascade reactions via exogenous or endogenous
processes. The Bcl-2 protein, encoded by the Bcl-2 gene, serves an
essential role in the formation of a variety of malignant tumors.
Bcl-2 does not promote cell proliferation, however it inhibits
apoptosis, which is the opposite of other members of the Bcl-2
family such as Bad, Bid and Bax that promote cellular apoptosis
(33,34). Bcl-2 was first isolated by Tsujimoto
et al (35) from follicular
non-Hodgkin's lymphoma B cells and is one of the most well
investigated apoptosis-regulating genes. The results of a study
performed by McDonnell et al (36) on Bcl-2 transgenic mice demonstrated
that high expression of Bcl-2 can inhibit the apoptosis of
cells.
From the results of previous studies and the present
study, it can be hypothesized that the important role of Bcl-2
serves in inhibiting apoptosis accounts for its association with
hemangioma formation. The results of a study by Turner et al
(37) demonstrated that the
expression of Bcl-2 was positively correlated with tumor
microvessel density in pituitary tumors. In addition, the results
of a study by Gazzaniga et al (38) revealed that the expression of Bcl-2
was positively correlated with VEGF and bFGF expression, indicating
that Bcl-2 is associated with tumor angiogenesis. Furthermore, a
previous study identified higher VEGF and bFGF expression in the
proliferative stage of hemangioma compared with the involuting
stage (39).
The present study demonstrated that the expression
of Bcl-2 in the endothelial cells of proliferative stage hemangioma
samples was significantly increased compared with the expression in
involuting stage hemangioma samples and healthy skin tissue
samples. It has been suggested that Bcl-2 inhibits the apoptosis of
endothelial cells, resulting in an imbalance between cell
proliferation and apoptosis. An increase in cell proliferation and
inhibition of cell apoptosis are involved in the formation and
development of hemangioma.
p53 is tumor suppressor protein that is associated
with tumor development and serves an essential role in the
regulation of cell cycle progression, cell differentiation and
apoptosis. Previous studies have revealed that >50% of tumors
are associated with p53 gene mutation (40–43). p53
is ~53 kDa and is encoded by the TP53 gene. p53 is important for
the maintenance of normal cell growth and inhibition of malignant
proliferation. The primary mechanisms of action of p53 are the
regulation of the cell cycle and induction of cell death. In the
case of DNA damage, increased p53 expression can induce cell cycle
arrest at the G1 stage to allow for DNA damage repair
and if the damage cannot be repaired, then cell apoptosis is
induced. p53 has wild-type (wt-TP53) and mutant (mt-TP53) forms.
mt-TP53 cannot inhibit cell proliferation and induce apoptosis,
enabling malignant cells to proliferate. In a previous study, the
expression of wt-p53 in healthy tissue surrounding a hemangioma was
low and only mt-p53 was detected through immunohistochemistry and
quantum dot analyses (44).
In the present study, the presence of mt-p53 in
human hemangioma tissue was detected through immunohistochemical
and quantum dot analysis in 38 cases, and relatively accurate
staging was conducted through measuring PCNA expression. The
results of the current study demonstrated that compared with
healthy skin tissue samples there was no significant difference in
endothelial cell p53 expression in involuting stage hemangioma.
Proliferative stage hemangioma endothelial cell p53 expression was
significantly increased compared with involuting stage hemangioma.
The expression of p53 was relatively low in the involuting stage,
but relatively high in the proliferative stage. These results were
consistent with results on the expression of VEGF in different
stages of hemangioma (45,46). A number of studies have indicated that
p53 can promote the expression of VEGF, increasing tumor
angiogenesis; in addition, coexpression and a positive correlation
between p53 and VEGF expression have been identified in a number of
tumor types (47–49). Therefore, p53 may promote the
formation of hemangioma through upregulation of VEGF expression.
p53 can influence apoptosis by regulating Bcl-2 and BCL2 associated
X apoptosis regulator (BAX) gene expression. p53 can specifically
inhibit the expression of Bcl-2 and promote the expression of BAX.
p53 is a direct transcription-activating factor of BAX. Therefore,
if p53 mutation occurs, it will reduce the apoptosis of cells.
In conclusion, Bcl-2 can cause an imbalance between
cell proliferation and apoptosis in endothelial cells through the
inhibition of endothelial cell apoptosis. In addition, p53 may
promote the proliferation of endothelial cells in proliferating
cutaneous hemangioma and contribute to the generation of a large
number of vascular endothelial cells. The results of the present
study have provided future directions for the study of tumor
angiogenesis. Understanding the role of these regulatory factors in
tumor angiogenesis may aid in the development of novel treatments
to inhibit tumor angiogenesis.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant nos. 30872688 and
81402242.
References
|
1
|
Kilcline C and Frieden IJ: Infantile
hemangiomas: How common are they? A systematic review of the
medical literature. Pediatr Dermatol. 25:168–173. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Takahashi K, Mulliken JB, Kozakewich HP,
Rogers RA, Folkman J and Ezekowitz RA: Cellular markers that
distinguish the phases of hemangioma during infancy and childhood.
J Clin Invest. 93:2357–2364. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mulliken JB and Young AE: Vascular
birthmarks: Hemangiomas and malformations. W. B. Saunders Co.;
Philadelphia: 1988
|
|
4
|
Risau W and Flamme I: Vasculogenesis. Annu
Rev Cell Dev Biol. 11:73–91. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Risau W: Mechanisms of angiogenesis.
Nature. 386:671–674. 1997. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Flamme I, Frölich T and Risau W: Molecular
mechanisms of vasculogenesis and embryonic angiogenesis. J Cell
Physiol. 173:206–210. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Carmeliet P: Angiogenesis in health and
disease. Nat Med. 9:653–660. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Folkman J: Angiogenesis in cancer,
vascular, rheumatoid and other disease. Nat Med. 1:27–31. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Brandling-Bennett HA, Metry DW, Baselga E,
Lucky AW, Adams DM, Cordisco MR and Frieden IJ: Infantile
hemangiomas with unusually prolonged growth phase: A case series.
Arch Dermatol. 144:1632–1637. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Stiles J, Amaya C, Pham R, Rowntree RK,
Lacaze M, Mulne A, Bischoff J, Kokta V, Boucheron LE, Mitchell DC
and Bryan BA: Propranolol treatment of infantile hemangioma
endothelial cells: A molecular analysis. Exp Ther Med. 4:594–604.
2012.PubMed/NCBI
|
|
11
|
Bourdelat D, Melki E, Mazzola C and
Marreel A: Congenital prenatal hemangioma: Diagnosis and treatment.
Arch Pediatr. 17:383–386. 2010.(In French). View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yuan WL, Qin XJ and Wang XK: Expression
and correlation of mast cell, Clusterin/apoJ and transforming
growth factor-beta in the different stages of human dermal
hemangioma. Hua Xi Kou Qiang Yi Xue Za Zhi. 27:361–365. 2009.(In
Chinese). PubMed/NCBI
|
|
13
|
Chang J, Most D, Bresnick S, Mehrara B,
Steinbrech DS, Reinisch J, Longaker MT and Turk AE: Proliferative
hemangiomas: Analysis of cytokine gene expression and angiogenesis.
Plast Reconstr Surg. 103:1–10. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tan ST, Wallis RA, He Y and Davis PF: Mast
cells and hemangioma. Plast Reconstr Surg. 113:999–1011. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Shan S, Shan G and Zhang D: Treatment of
hemangioma by transfection of antisense VEGF gene. J Huazhong Univ
Sci Technolog Med Sci. 29:335–339. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shan G, Tang T and Zhang D: Expression of
HLA-G in hemangioma and its clinical significance. J Huazhong Univ
Sci Technolog Med Sci. 32:713–718. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Razon MJ, Kräling BM, Mulliken JB and
Bischoff J: Increased apoptosis coincides with onset of involution
in infantile hemangioma. Microcirculation. 5:189–195. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cook BD, Ferrari G, Pintucci G and
Mignatti P: TGF-beta1 induces rearrangement of
FLK-1-VE-cadherin-beta-catenin complex at the adherens junction
through VEGF-mediated signaling. J Cell Biochem. 105:1367–1373.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sharma MR, Tuszynski GP and Sharma MC:
Angiostatin-induced inhibition of endothelial cell
proliferation/apoptosis is associated with the down-regulation of
cell cycle regulatory protein cdk5. J Cell Biochem. 91:398–409.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ola MS, Nawaz M and Ahsan H: Role of Bcl-2
family proteins and caspases in the regulation of apoptosis. Mol
Cell Biochem. 351:41–58. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tobiume K: Involvement of Bcl-2 family
proteins in p53-induced apoptosis. J Nippon Med Sch. 72:192–193.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Vaseva AV and Moll UM: The mitochondrial
p53 pathway. Biochim Biophys Acta. 1787:414–420. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Crowe DL and Sinha UK: P53 apoptotic
response to DNA damage dependent on bcl2 but not bax in head and
neck squamous cell carcinoma lines. Head Neck. 28:15–23. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hainaut P: The tumor suppressor protein
p53: A receptor to genotoxic stress that controls cell growth and
survival. Curr Opin Oncol. 7:76–82. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Han Z, Chatterjee D, He DM, Early J,
Pantazis P, Wyche JH and Hendrickson EA: Evidence for a G2
checkpoint in p53-independent apoptosis induction by X-irradiation.
Mol Cell Biol. 15:5849–5857. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Marinaş MC, Mogoş DG, Simionescu CE,
Stepan A and Tănase F: The study of p53 and p16 immunoexpression in
serous borderline and malignant ovarian tumors. Rom J Morphol
Embryol. 53:1021–1025. 2012.PubMed/NCBI
|
|
27
|
Cheok CF and Lane DP: Seeking synergy in
p53 transcriptional activation for cancer therapy. Discov Med.
14:263–271. 2012.PubMed/NCBI
|
|
28
|
Jayaraman B, Valiathan GM, Jayakumar K,
Palaniyandi A, Thenumgal SJ and Ramanathan A: Lack of mutation in
p53 and H-ras genes in phenytoin induced gingival overgrowth
suggests its non cancerous nature. Asian Pac J Cancer Prev.
13:5535–5538. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Martina JD, Simmons C and Jukic DM:
High-definition hematoxylin and eosin staining in a transition to
digital pathology. J Pathol Inform. 2:452011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mulliken JB and Glowacki J: Hemangiomas
and vascular malformations in infants and children: A
classification based on endothelial characteristics. Plast Reconstr
Surg. 69:412–422. 1982. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tomek M, Akiyama T and Dass CR: Role of
Bcl-2 in tumour cell survival and implications for pharmacotherapy.
J Pharm Pharmacol. 64:1695–1702. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Xiao Z, Shan J, Li C, Luo L, Lu J, Li S,
Long D and Li Y: Mechanisms of cyclosporine-induced renal cell
apoptosis: A systematic review. Am J Nephrol. 37:30–40. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sermeus A, Genin M, Maincent A, Fransolet
M, Notte A, Leclere L, Riquier H, Arnould T and Michiels C:
Hypoxia-induced modulation of apoptosis and BCL-2 family proteins
in different cancer cell types. PLoS One. 7:e475192012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Shu R, Liu DL and Wang Q: Effect of
remifentanil on expression of Bcl-2 and caspase-3 in ratscerebral
cortex following Ischemia-reperfusion injury. Acta Medicinae
Universitatis Scientiae et Technologiae Huazhong. 41:72–75.
2012.(In Chinese).
|
|
35
|
Tsujimoto Y, Finger LR, Yunis J, Nowell PC
and Croce CM: Cloning of the chromosome breakpoint of neoplastic B
cells with the t(14;18) chromosome translocation. Science.
226:1097–1099. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
McDonnell TJ, Deane N, Platt FM, Nunez G,
Jaeger U, McKearn JP and Korsmeyer SJ: bcl-2-immunoglobulin
transgenic mice demonstrate extended B cell survival and follicular
lymphoproliferation. Cell. 57:79–88. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Turner HE, Nagy Z, Gatter KC, Esiri MM,
Wass JA and Harris AL: Proliferation, bcl-2 expression and
angiogenesis in pituitary adenomas: Relationship to tumour
behaviour. Br J Cancer. 82:1441–1445. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Gazzaniga P, Gandini O, Gradilone A,
Silvestri I, Giuliani L, Magnanti M, Gallucci M, Saccani G, Frati L
and Agliano AM: Detection of basic fibroblast growth factor mRNA in
urinary bladder cancer: Correlation with local relapses. Int J
Oncol. 14:1123–1127. 1999.PubMed/NCBI
|
|
39
|
Bielenberg DR, Bucana CD, Sanchez R,
Mulliken JB, Folkman J and Fidler IJ: Progressive growth of
infantile cutaneous hemangiomas is directly correlated with
hyperplasia and angiogenesis of adjacent epidermis and inversely
correlated with expression of the endogenous angiogenesis
inhibitor, IFN-beta. Int J Oncol. 14:401–408. 1999.PubMed/NCBI
|
|
40
|
Halasi M, Pandit B and Gartel AL:
Proteasome inhibitors suppress the protein expression of mutant
p53. Cell Cycle. 13:3202–3206. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Giurgea LN, Ungureanu C and Mihailovici
MS: The immunohistochemical expression of p53 and Ki67 in ovarian
epithelial borderline tumors. Correlation with clinicopathological
factors. Rom J Morphol Embryol. 53:967–973. 2012.PubMed/NCBI
|
|
42
|
Rao Z and Ding Y: Ubiquitin pathway and
ovarian cancer. Curr Oncol. 19:324–328. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ji Y, Li K, Xiao X, Zheng S, Xu T and Chen
S: Effects of propranolol on the proliferation and apoptosis of
hemangioma-derived endothelial cells. J Pediatr Surg. 47:2216–2223.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Porter PL, Gown AM, Kramp SG and Coltrera
MD: Widespread p53 overexpression in human malignant tumors. An
immunohistochemical study using methacarn-fixed, embedded tissue.
Am J Pathol. 140:145–153. 1992.PubMed/NCBI
|
|
45
|
Itahana Y and Itahana K: Emerging roles of
mitochondrial p53 and ARF. Curr Drug Targets. 13:1633–1640. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Crasta JA, Mishra S and Vallikad E:
Ovarian serous carcinoma: Relationship of p53 and bcl-2 with tumor
angiogenesis and VEGF expression. Int J Gynecol Pathol. 30:521–526.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Kang SM, Maeda K, Onoda N, Chung YS,
Nakata B, Nishiguchi Y and Sowa M: Combined analysis of p53 and
vascular endothelial growth factor expression in colorectal
carcinoma for determination of tumor vascularity and liver
metastasis. Int J Cancer. 74:502–507. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Koide N, Nishio A, Hiraguri M, Hanazaki K,
Adachi W and Amano J: Coexpression of vascular endothelial growth
factor and p53 protein in squamous cell carcinoma of the esophagus.
Am J Gastroenterol. 96:1733–1740. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Tian Y, Ding RY, Zhi YH, Guo RX and Wu SD:
Analysis of p53 and vascular endothelial growth factor expression
in human gallbladder carcinoma for the determination of tumor
vascularity. World J Gastroenterol. 12:415–419. 2006. View Article : Google Scholar : PubMed/NCBI
|