Introduction
The standard treatment for ovarian cancer is surgery
followed by anticancer therapy (1). A
total of ~70% of advanced ovarian cancer recurs, so advanced
ovarian cancer ultimately has a 5-year survival rate of between 30
and 40% (2). Ovarian cancer has the
highest mortality rate among gynecological malignancies (3).
Photodynamic therapy (PDT) has garnered attention as
a novel therapy to reduce the tumor burden on a patient. PDT has
been used to treat superficial esophageal cancer, early lung cancer
and early gastric cancer (4–6). In gynecology, PDT has been used to treat
early cancer of the cervix (7).
Younger patients tend to opt to receive treatment that preserves
fertility, and PDT may be such a treatment (7). Therefore, attention has focused on the
usefulness of PDT in that regard. Unlike conventional approaches,
PDT leaves the cervix intact, has a high cure rate, and does not
hamper pregnancy or delivery following surgery (7).
PDT is a therapy involving photosensitizers (or
their precursors) with an affinity for tumors. The specific
accumulation of photosensitizers in tumor tissue and blood vessels
and the excitation of light are used by PDT to produce singlet
oxygen (a type of reactive oxygen species) with potent cytocidal
action. PDT using hematoporphyrin derivatives to treat skin
metastasis of breast cancer was first studied by Dougherty et
al (8) in 1979. Subsequently, PDT
has been investigated in numerous studies. PDT does not
substantially damage normal tissue and it treats lesions with low
levels of light energy, so various lesions may be treated by
irradiation using a laser beam. Previously, we have described the
potential effectiveness of using PDT to treat ovarian cancer;
however, certain types of ovarian cancer are resistant to PDT
(9). Results suggested that
protoporphyrin IX (PpIX) in cells is crucial to satisfactory
treatment of cancer with PDT (9).
PpIX is a metabolite that is converted from aminolevulinic acid via
heme synthesis and degradation pathways (10).
In the present study, five ovarian cancer cell lines
with distinct histological types were used to produce subcutaneous
tumors in mice. Subsequently, the effectiveness of PDT in treating
those tumors was determined. In addition, PDT-sensitive cells and
therapy-resistant cells were identified, and a microarray technique
was used to compare the expression of genes that code for enzymes
involved in heme synthesis and degradation. Glutathione transferase
Omega 1 (GSTO1) is involved in the conversion of PpIX into heme.
The level of expression of GSTO1 was compared in the various cell
lines.
Materials and methods
Cell lines and cell culture
HTOA cells were obtained from RIKEN BioResource
Center (Tsukuba, Japan). MCAS and TOV21G cells were obtained from
the American Type Culture Collection (Manassas, VA, USA). HRA and
DISS cells were provided by Dr Y. Kikuchi (National Defense Medical
College, Tokorozawa, Japan) (11) and
Dr Y. Saga (Jichi Medical School, Tochigi, Japan), respectively.
HTOA, HRA and DISS cells were derived from human ovarian serous
adenocarcinoma. MCAS and TOV21G cells were derived from human
ovarian mucinous and clear cell adenocarcinoma, respectively. These
five cell lines were verified in writing as being ovarian in
origin. All cell lines were grown in RPMI-1640 medium with 10%
fetal bovine serum, 100 units/ml penicillin and 100 µg/ml
streptomycin at 37°C in a humidified atmosphere containing 5%
CO2/95% air.
Animals
Animal experiments were approved by the Animal
Research Committee of Hirosaki University (Hirosaki, Japan) and all
animals were cared for and handled in accordance with the Rules for
Animal Experimentation of Hirosaki University and animal practices
as defined by national and local bodies governing animal welfare
(Guide for the Care and Use of Laboratory Animals published by the
National Institutes of Health). A total of 10 8-week-old female
BALB/c nu/nu mice weighing 20–25 g (CLEA Japan, Inc., Tokyo, Japan)
were used in the present study. All mice were group housed in
plastic cages with stainless steel grid tops in an air-conditioned
room with a 12-h light/12-h dark cycle and fed with water and food
ad libitum in the Institute for Animal Experiments of Hirosaki
University.
Cancer-bearing mouse model
Cancer cells (0.5×106 cells) in 200 µl
PBS were injected subcutaneously into the dorsal region of the nude
mice under general anesthesia. Tumors were grown until the largest
diameter reached 3 mm prior to commencing treatment. Then, the mice
were divided into two groups, a treatment group and a control
group, containing 5 mice each. The tumor dimensions were measured
three times/week using a caliper and tumor volume was calculated
using the equation V (mm3)=AxB2/2, where A is
the largest diameter and B is the smallest diameter.
PDT
5-Aminolevulinic acid methyl ester hydrochloride
(methyl-ALA) was obtained from Cosmo Bio International (Tokyo,
Japan). Immediately following dissolution in PBS at a concentration
of 10 mg/ml, PBS. The methyl-ALA solution was injected
intraperitoneally into the nude mice of the treatment group. The
mice in the treatment group and in the control group received an
intraperitoneal injection of 250 mg/kg methyl-ALA and PBS alone,
respectively (9). PDT was
administered by 10 min irradiation using a 150 W halogen light, 3 h
after methyl-ALA or PBS injection (control) (9). Each mouse received PDT twice/week for 3
weeks.
Determination of intracellular PpIX
following methyl-ALA exposure
The cellular uptake of methyl-ALA, which was
converted into fluorescent PpIX inside HTOA, HRA, DISS, MCAS and
TOV21G cells, was determined using high-performance liquid
chromatography (HPLC). Each cell line was seeded in 100-mm culture
dishes and incubated with 5.5 µM freshly prepared methyl-ALA for 3
h. Methyl-ALA was removed and the cells were trypsinized and
centrifuged at 10,000 × g for 5 min at 4°C, then washed twice with
ice-cold PBS. The cell pellet was then resuspended in 1 ml
radioimmunoprecipitation assay buffer (Sigma-Aldrich; Merck KGaA;
Darmstadt, Germany). The mixture was agitated gently for 15 min on
ice and centrifuged at 10,000 × g for 5 min at 4°C to pellet the
cell debris. The supernatant was transferred to a new tube and
subjected to HPLC analysis (LC-20AT Prominence; Shimadzu, Kyoto,
Japan) equipped with a reversed-phase C18 column (CAPCELL PAK, C18;
SG300; 5 µm, 4.6×250 mm; Shiseido, Tokyo, Japan). Elution was
started with 10% solvent A (50 mM phosphate buffer) and 90% solvent
B (acetonitrile) for 7 min. The elution flow throughout was kept
constant at a rate of 2.0 ml/min. The excitation wavelength was set
at 404 nm and the fluorescence emission wavelength was set at 620
nm. Subsequent to dissolving 10 mg of PpIX disodium salt
(Sigma-Aldrich; Merck KGaA) with a few drops of 1 M HCl, a stock
PpIX standard was prepared by diluting it in 50 ml of
N,N-dimethylformamide. All results are presented for three
independent experiments.
Total RNA isolation
Cells were homogenized using a homogenizer and RNA
was extracted using TRIzol® reagent (Invitrogen; Thermo
Fisher Scientific, Inc., Waltham, MA, USA), according to the
manufacturer's protocol. The extracted total RNA was then purified
using an RNeasy MiniElute Cleanup kit column (Qiagen, Inc.,
Valencia, CA, USA) with incubation with DNase (Qiagen, Inc.). Total
RNA extracted was quantified using an Eppendorf UV
spectrophotometer, and the integrity of the RNA samples was
monitored using an Agilent 2100 Bioanalyzer (Agilent Technologies,
Inc., Santa Clara, CA, USA) and a NanoDrop ND-1000 kit (Thermo
Fisher Scientific, Inc.). Only RNAs with an
A260/A280 ratio >1.8 and an RNA integrity
number >7 were used for microarray experiments. The remaining
good-quality RNAs were retained for subsequent confirmative reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
experiments.
Microarray analysis
Each good-quality sample was hybridized to the
Affymetrix HG-U133 plus 2.0 GeneChip (Affymetrix, Inc., Santa
Clara, CA, USA). This gene chip analyzes the expression level of
38,500 well-characterized human genes. First, 10 µg of the total
RNA was reverse-transcribed with the SuperScript Choice System
(Invitrogen; Thermo Fisher Scientific, Inc.) with oligo dT primers
containing a T7 RNA polymerase promoter site. cDNA was then in
vitro-transcribed and labeled with biotin using the IVT labeling
kit (Affymetrix, Inc.), followed by the fragmentation of the
biotinylated cRNA. The quality of this cRNA was then assessed with
the Agilent 2100 Bioanalyzer (Agilent Technologies, Inc.). The
fragmented cRNA was hybridized overnight to Affymetrix Human Genome
U133APlus 2.0 Arrays (Affymetrix, Inc.) and scanned according to
the manufacturer's guidelines. The chips were washed and stained
using the GeneChip Fluidics Station 400 (Affymetrix, Inc.) and then
scanned with the GeneChip Scanner 3000 (Affymetrix, Inc.).
Labeling, hybridization, image scanning and data analysis were
performed by Kurabo Industries Ltd. (Osaka, Japan).
RT-qPCR
qPCR was used to evaluate the expression of the
GSTO1 gene. TaqMan® Gene Expression assays (Thermo
Fisher Scientific, Inc.) was used with TaqMan® probes
for GSTO1 and β-actin (ACTB). Primers used for PCR were as follows:
GSTO1, 5′-AGGATCCACGATGTCCGGGGAGTCAG-3′ (forward) and
5′-CGAATTCAGAGCCCATAGTCACAG-3′ (reverse), and ACTB,
5′-AGTCCCTTGCCATCCTAAAAGC-3′ (forward) and
5′-GGGAGAGGACTGGGCCATT-3′ (reverse). Prior to qPCR, reverse
transcription of 500 ng total RNA was performed using a
SuperScript® VILO™ cDNA Synthesis kit (Thermo
Fisher Scientific, Inc.), according to the manufacturer's protocol.
qPCRs were set up in 20 µl volumes with 50-fold cDNA dilutions, 20X
TaqMan® gene expression assay mixture, 2X
TaqMan® Universal PCR master mix II (Thermo Fisher
Scientific, Inc.) and distilled water. PCRs were performed in
quadruplicate following the manufacturer's protocols on a model ABI
Prism 7000 Real-Time PCR system (Thermo Fisher Scientific, Inc.)
using the following protocol. Initial denaturation and polymerase
activation at 95°C for 10 min, followed by 40 cycles of
denaturation at 95°C for 15 sec and annealing and extension at 60°C
for 1 min. ACTB was used as a reference gene for normalization of
samples.
Statistical analysis
The results are presented as the mean ± standard
deviation. Differences in tumor volume between the groups were
analyzed using a non-parametric Mann-Whitney U test. Other
statistical analyses were carried out using Student's t-test,
χ2 test or Fisher's exact probability test. P<0.05
was considered to indicate a statistically significant
difference.
Results
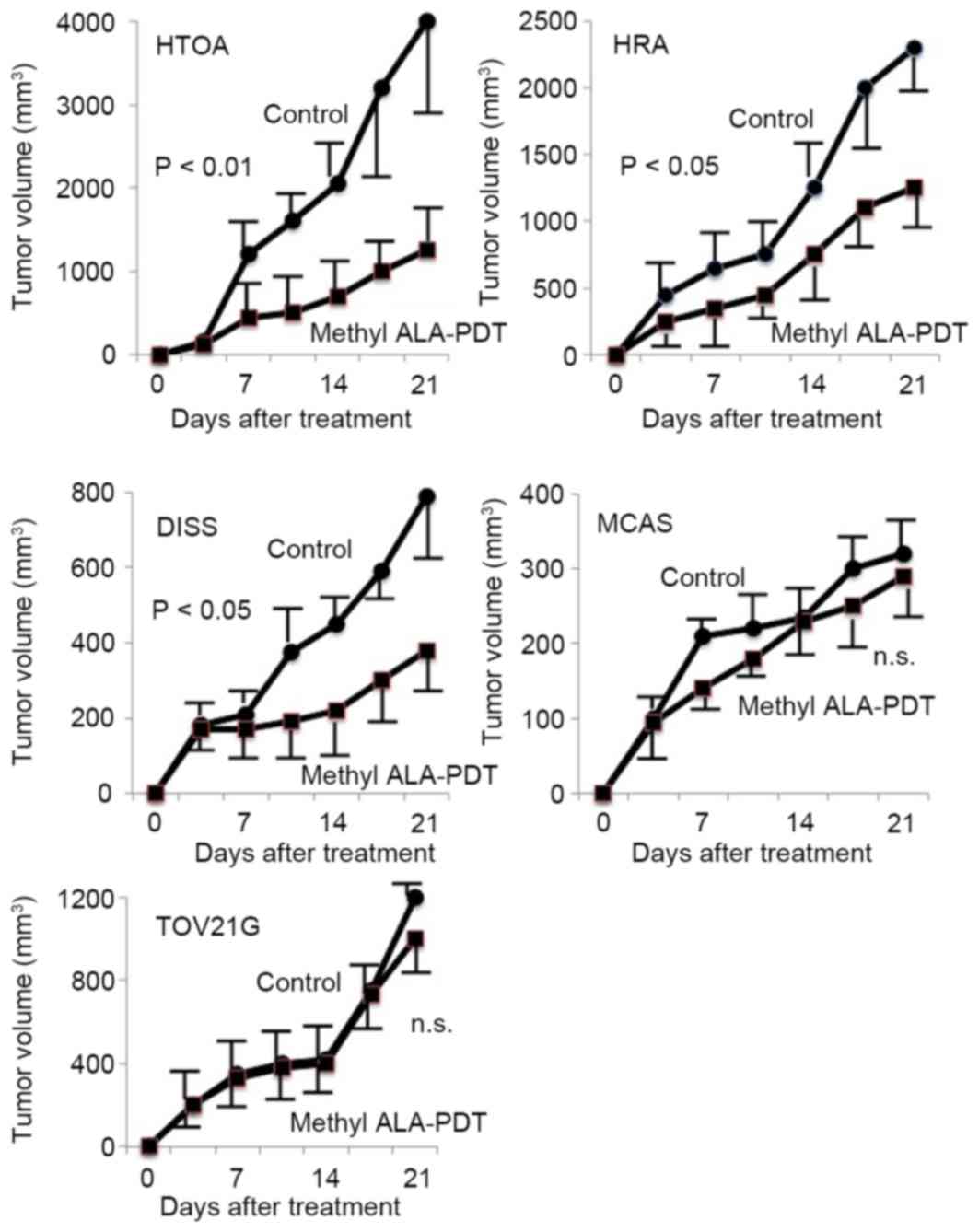
Antitumor action of PDT in ovarian
cancer cell lines
PDT resulted in significant tumor shrinkage in HTOA,
HRA and DISS tumor cells in comparison with the controls
(P<0.05; Fig. 1). No significant
differences in tumor volume were identified in MCAS and TOV21G
cells between the PDT group and the control group (Fig. 1).
PpIX levels in ovarian cancer
cells
Methyl-ALA is converted into PpIX. PpIX levels
increased significantly in HTOA, DISS and HRA cells in comparison
with levels in MCAS and TOV21G cells (Table I).
 | Table I.Determination of protoporphyrin IX in
five ovarian cancer cell lines. |
Table I.
Determination of protoporphyrin IX in
five ovarian cancer cell lines.
| Cell line | Amount of
protoporphyrin IX, mg/dl |
|---|
| HTOA | 62.6±9.4a |
| HRT | 67.7±2.7a |
| DISS | 59.3±8.9a |
| MCAS | 22.4±4.7 |
| TOV21G | 24.1±3.9 |
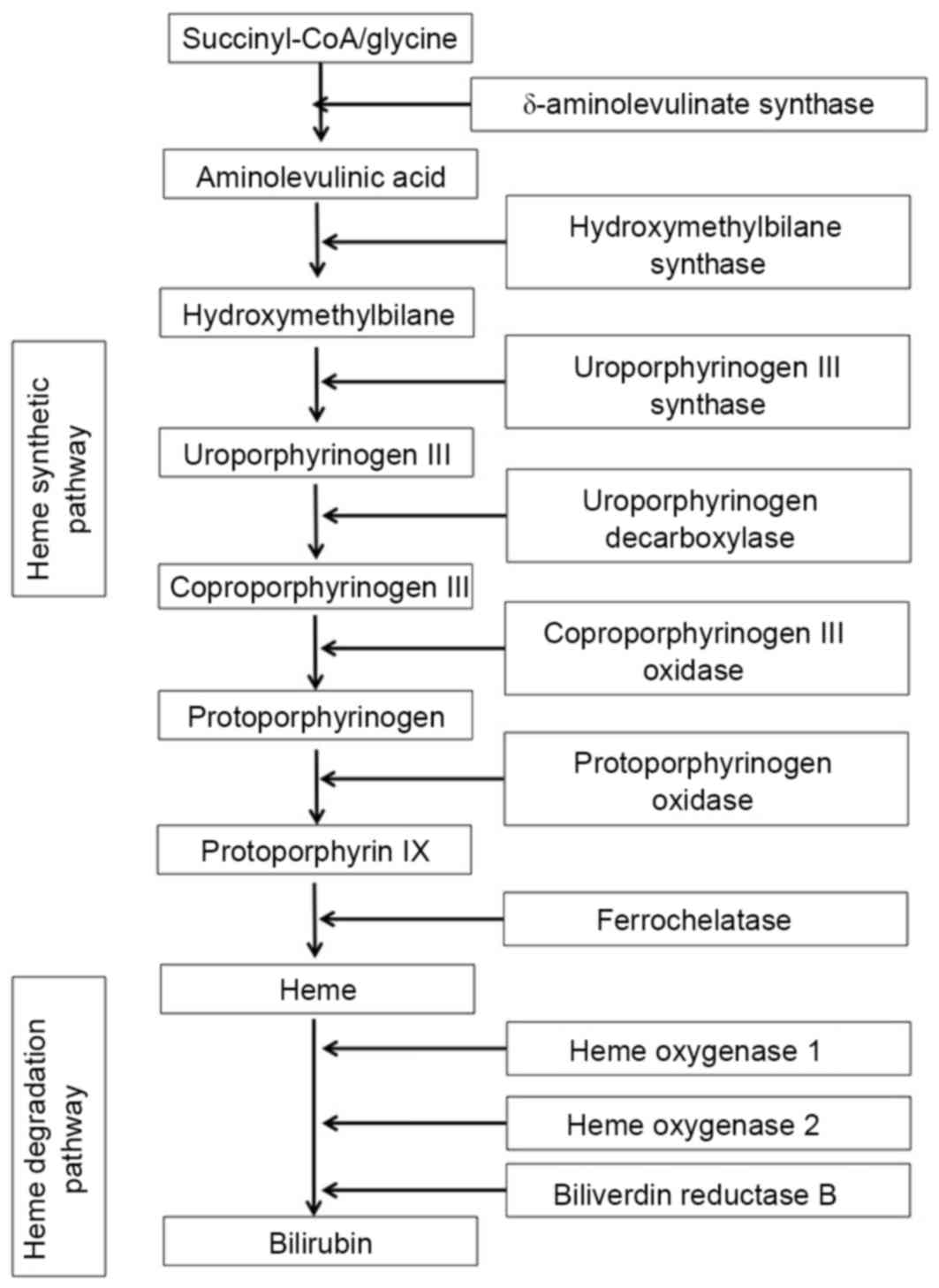
Genetic profile of enzymes involved in
heme synthesis and degradation in DISS and MCAS cells
The genetic profile of enzymes involved in heme
synthesis and degradation (Fig. 2)
was determined using microarray assay. A 3-fold increase in the
level of expression was defined as marked upregulation (Table II). δ-aminolevulinate (δ-ALA)
synthase is a rate-limiting enzyme in heme synthesis (Fig. 2), and expression of δ-ALA synthase was
increased markedly in MCAS cells in comparison with that in DISS
cells (Table II). Heme oxygenase 2
is an enzyme that degrades heme into biliverdin and biliverdin
reductase B is an enzyme that reduces biliverdin into bilirubin
(Fig. 2). The expression levels of
heme oxygenase 2 and biliverdin reductase B increased 11.5-fold and
6.8-fold, respectively, in MCAS cells in comparison with levels of
expression in DISS cells (Table
II).
| Table II.Results of microarray analysis on the
expression of enzymes involved in the heme synthetic and
degradation pathway in DISS and MCAS cells. |
Table II.
Results of microarray analysis on the
expression of enzymes involved in the heme synthetic and
degradation pathway in DISS and MCAS cells.
|
| Signal |
|
|---|
|
|
|
|
|---|
| Gene title | DISS | MCAS | Fold difference
(MCAS/DISS) |
|---|
| Heme synthetic
pathway |
|
δ-aminolevulinic synthase | 1,040 | 3,380 | 3.25 |
|
Hydroxymethylbilane
synthase | 830 | 770 | 0.93 |
|
Uroporphyrinogen III
synthase | 1,010 | 1,530 | 1.51 |
|
Uroporphyrinogen
decarboxylase | 4,710 | 2,320 | 0.49 |
|
Coproporphyrinogen III
oxidase | 2,220 | 1,620 | 0.73 |
|
Protoporphyrinogen
oxidase | 690 | 510 | 0.74 |
|
Ferrochelatase | 500 | 250 | 0.50 |
| Heme degradation
pathway |
| Heme
oxygenase 1 | 150 | 310 | 2.07 |
| Heme
oxygenase 2 | 90 | 1,040 | 11.56 |
|
Biliverdin reductase B | 540 | 3,680 | 6.81 |
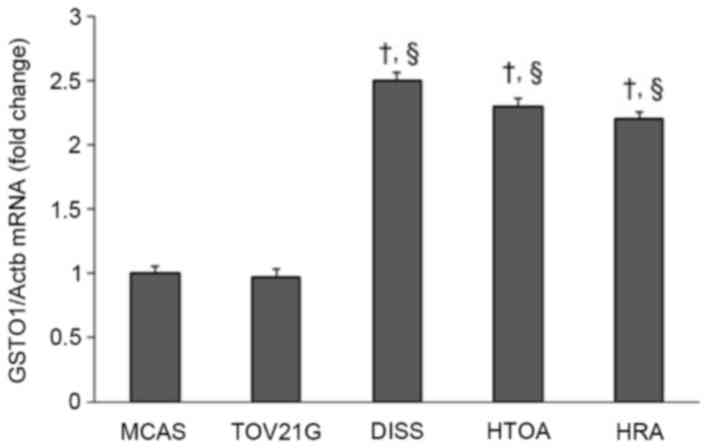
Expression of GSTO1 in ovarian cancer
cells
The expression level of GSTO1 in HTOA, HRA and DISS
cells was ~2.5-fold increased compared with that in either MCAS or
TOV21G cells, a difference which was determined to be significant
(P<0.001; Fig. 3).
Discussion
During PDT, methyl-ALA is taken up by cells and
converted into PpIX. PpIX levels are a crucial aspect of the
antitumor action of PDT. In the present study, it was identified
that antitumor action various depending on PpIX levels in cells. In
other words, PDT is markedly more effective at treating tumors
developing from ovarian cancer cells with high PpIX levels.
PpIX is an intermediate in heme synthesis and
degradation (10). When 5.5 µM
methyl-ALA was added to ovarian cancer cells, PDT-sensitive HTOA,
HRA and DISS cells exhibited significantly higher PpIX levels in
comparison with PDT-resistant MCAS and TOV21G cells. A microarray
assay was used to analyze the genetic profile of converting enzymes
involved in heme synthesis and degradation in DISS and MCAS cells.
This analysis indicated that MCAS cells exhibited increased
expression (3-fold) of the δ-ALA synthase gene (which codes for a
rate-limiting enzyme in heme synthesis), increased expression
(10-fold) of the heme oxygenase 2 gene (which codes for an enzyme
that degrades heme into biliverdin), and increased expression
(7-fold) of the biliverdin reductase B gene (which codes for an
enzyme that reduces biliverdin into bilirubin) in comparison with
DISS cells. These results suggest that PpIX tends not to accumulate
in PDT-resistant cells despite active heme synthesis and
degradation, and despite the uptake of a derivative of an exogenous
amino acid.
Sustaining PpIX levels in cells is an aspect that
requires consideration in order to increase the effectiveness with
which PDT is able to treat ovarian cancer. Ferrochelatase is an
enzyme that converts PpIX into heme. Inhibiting ferrochelatase may
assist in sustaining PpIX levels in cells. Lead is a typical
ferrochelatase inhibitor; however, the use of such an inhibitor
clinically would be precluded since lead poisoning causes porphyria
(12). The inhibition of heme
oxygenase increases the accumulation of heme, i.e. it increases the
levels of iron, a potent source of free radicals (13).
GSTO1 inhibits the conversion of PpIX into heme
(14) and GSTO1 catalyzes the
detoxification of heavy metals including iron (through the
conjugation of glutathione to those substances) (15). Heme induces expression of the
erythropoietin gene through activation of hypoxia-inducible factor
1 (16). Erythropoietin is a
hematopoietic hormone, and erythropoietin is known to inhibit
sensitivity to PDT (17). Recent
studies have revealed that tyrosine protein kinase Met
(c-Met)/phosphoinositide 3-kinase (PI3K) signaling is related to
resistance to PDT (18).
Erythropoietin may induce resistance to PDT through the activation
of c-Met/PI3K signaling. GSTO1 inhibits the conversion of PpIX into
heme, so GSTO1 may act to sustain PpIX levels in cells and inhibit
expression of the erythropoietin gene, thus assisting with the
maintenance of sensitivity to PDT. In the present study, the
expression of GSTO1 in DSS cells increased significantly in
comparison with that in MCAS cells. This result is consistent with
the result that DISS cells exhibited significantly higher PpIX
levels compared with MCAS cells.
PDT is accompanied by oxidative stress (19). Oxidative stress is a biological
phenomenon whereby reactive oxygen species are produced and
subsequently cause cellular damage. In cancer tissue, inflammation
has been triggered and angiogenesis is extensive, therefore further
reactive oxygen species are produced. A recent study identified
that GSTO1 provides protection from oxidative stress (20). This result is consistent with the
results of the present study that DISS cells that expressed
increased levels of GSTO1 responded well to PDT.
The photosensitizer precursor used in the present
study was methyl-ALA. Methyl-ALA has been demonstrated to be
selectively taken up by tumor tissue (9). Other approaches are being investigated,
including delivery of a photosensitizer to a tumor via a
nanoparticle drug delivery system (21) and increasing tumor selectivity by
linking a photosensitizer to the gene encoding β-galactosidase
(22). Studies by the present authors
are underway to examine transfer of the GSTO1 gene to tumor
cells and direct induction of PpIX in tumor cells. Further research
is warranted to ensure that PDT is universally effective at
treating a range of cancer cells.
Sensitivity to PDT is associated with PpIX levels in
cells. The results of the present study suggested that PpIX tends
not to accumulate in PDT-resistant cells despite active heme
synthesis and degradation and despite the uptake of a derivative of
an exogenous amino acid. The results of the present study suggested
that high levels of GSTO1 expression are associated with increased
sensitivity to PDT.
Acknowledgements
The present study was supported by a Grant-in Aid
for Cancer Research from the Ministry of Education, Science and
Culture of Japan (grant no. 20591935 to Y.Y.).
References
|
1
|
Hofstetter G, Concin N, Braicu I, Chekerov
R, Sehouli J, Cadron I, van Gorp T, Trillsch F, Mahner S, Ulmer H,
et al: The time interval from surgery to start of chemotherapy
significantly impacts prognosis in patients with advanced serous
ovarian carcinoma-analysis of patient data in the prospective OVCAD
study. Gynecol Oncol. 131:15–20. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Akeson M, Zetterqvist BM, Dahllöf K,
Brännström M and Horvath G: Effect of adjuvant paclitaxel and
carboplatin for advanced stage epithelial ovarian cancer: A
population-based cohort study of all patients in western Sweden
with long-term follow-up. Acta Obstet Gynecol Scand. 87:1343–1352.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2013. CA Cancer J Clin. 63:11–30. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Casson AG: Photofrin PDT for early stage
esophageal cancer: A new standard of care? Photodiagnosis Photodyn
Ther. 6:155–156. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ikeda N, Usuda J, Kato H, Ishizumi T,
Ichinose S, Otani K, Honda H, Furukawa K, Okunaka T and Tsutsui H:
New aspects of photodynamic therapy for central type early stage
lung cancer. Lasers Surg Med. 43:749–754. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Spinelli P, Dal Fante M and Mancini A:
Current role of laser and photodynamic therapy in gastrointestinal
tumors and analysis of a 10-year experience. Semin Surg Oncol.
8:204–213. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Choi MC, Lee C and Kim SJ: Efficacy and
safety of photodynamic therapy for cervical intraepithelial
neoplasia: A systemic review. Photodiagnosis Photodyn Ther.
11:479–480. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Dougherty TJ, Lawrence G, Kaufman JH,
Boyle D, Weishaupt KR and Goldfarb A: Photoradiation in the
treatment of recurrent breast carcinoma. J Natl Cancer Inst.
62:231–237. 1979.PubMed/NCBI
|
|
9
|
Wakui M, Yokoyama Y, Wang H, Shigeto T,
Futagami M and Mizunuma H: Efficacy of a methyl ester of
5-aminolevulinic acid in photodynamic therapy for ovarian cancers.
J Cancer Res Clin Oncol. 136:1143–1150. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Furuyama K, Harigae H, Kinoshita C,
Shimada T, Miyaoka K, Kanda C, Maruyama Y, Shibahara S and Sassa S:
Late-onset X-linked sideroblastic anemia following hemodialysis.
Blood. 101:4623–4624. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kikuchi Y, Kizawa I, Oomori K, Miyauchi M,
Kita T, Sugita M, Tenjin Y and Kato K: Establishment of a human
ovarian cancer cell line capable of forming ascites in nude mice
and effects of tranexamic acid on cell proliferation and ascites
formation. Cancer Res. 47:592–596. 1987.PubMed/NCBI
|
|
12
|
Wetterberg L: Acute porphyria and lead
poisoning. Lancet. 1:4981966. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ho PS, Hoffman BM, Kang CH and Margoliash
E: Control of the transfer of oxidizing equivalents between heme
iron and free radical site in yeast cytochrome c peroxidase. J Biol
Chem. 1258:4356–4363. 1983.
|
|
14
|
Kodym R, Calkins PR and Story MD:
Anthracycline-induced erythroid differentiation of K562 cells is
inhibited by p28, a novel mammalian glutathione-binding stress
protein. Leuk Res. 25:151–156. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Paiva L, Hernández A, Martínez V, Creus A,
Quinteros D and Marcos R: Association between GSTO2 polymorphism
and the urinary arsenic profile in copper industry workers. Environ
Res. 110:463–468. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hofer T, Wenger RH, Kramer MF, Ferreira GC
and Gassmann M: Hypoxic up-regulation of erythroid
5-aminolevulinate synthase. Blood. 101:348–350. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Solár P, Koval J, Mikes J, Kleban J,
Solárová Z, Lazúr J, Hodorová I, Fedorocko P and Sytkowski AJ:
Erythropoietin inhibits apoptosis induced by photodynamic therapy
in ovarian cancer cells. Mol Cancer Ther. 7:2263–2271. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jung KA, Choi BH and Kwak MK: The
c-MET/PI3K signaling is associated with cancer resistance to
doxorubicin and photodynamic therapy by elevating BCRP/ABCG2
expression. Mol Pharmacol. 87:465–476. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Luna MC, Wong S and Gomer CJ: Photodynamic
therapy mediated induction of early response genes. Cancer Res.
54:1374–1380. 1994.PubMed/NCBI
|
|
20
|
Meng F, Zhang Y, Liu F, Guo X and Xu B:
Characterization and mutational analysis of omega-class GST (GSTO1)
from Apis cerana cerana, a gene involved in response to oxidative
stress. PLoS One. 9:e931002014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li Z, Sun L, Lu Z, Su X, Yang Q, Qu X, Li
L, Song K and Kong B: Enhanced effect of photodynamic therapy in
ovarian cancer using a nanoparticle drug delivery system. Int J
Oncol. 47:1070–1076. 2015.PubMed/NCBI
|
|
22
|
Koide Y, Urano Y, Yatsushige A, Hanaoka K,
Terai T and Nagano T: Design and development of enzymatically
activatable photosensitizer based on unique characteristics of
thiazole orange. J Am Chem Soc. 131:6058–6059. 2009. View Article : Google Scholar : PubMed/NCBI
|