Introduction
Lung cancer is one of the leading causes of cancer
associated mortality worldwide, with a 5-year survival rate of ~20%
(1). The conventional treatment of
cytotoxic chemotherapy for lung cancer appears to have reached an
effectiveness plateau and new treatment options are required
(2,3).
Angiogenesis, the growth of new blood vessels from pre-existing
ones, is a fundamental step in the transition of tumors between the
dormant and malignant stages (4,5). Vascular
endothelial growth factor (VEGF), through activation of VEGF
receptor 1 (VEGFR1) and 2 (VEGFR2), is a potent pro-angiogenic
factor and a key mediator of angiogenesis in malignant tissues
(6). Previous studies have revealed
that increased levels of VEGF and high microvessel density are
associated with poor prognosis in lung cancer patients (7,8). In
addition, the anti-VEGF monoclonal antibody bevacizumab has been
tested to improve tumor response and progression-free survival in
patients with non-small cell lung cancer (NSCLC) (3,9). Tumor
angiogenesis has been the target of major drug discovery programs
and angiogenesis inhibitors have been evaluated in clinical trials
for lung cancer treatment for the last decade (2,3,10–12).
Azithromycin is a Food and Drug
Administration-approved antibiotic and the primary drug for the
treatment of chlamydia and mycoplasma pneumonia (13). It destroys bacteria by inhibiting
protein synthesis through reversible binding to the 50S subunit of
bacterial ribosomes (14). With the
exception of its antimicrobial activity, azithromycin exerts
anti-inflammatory effects by suppressing the secretion of
pro-inflammatory cytokines, including interluekin-8 (15). Previously, azithromycin was reported
to inhibit the proliferation of cervical and gastric tumor cells
(16,17) and enhance the effects of chemotherapy
in NSCLC, however little is known about the underlying mechanisms
of its anti-cancer activities (18).
In the present study, the effect of azithromycin in
lung tumor angiogenesis was investigated using in vitro and
in vivo angiogenesis models, and a lung tumor xenograft
model. The present results demonstrated that azithromycin potently
inhibits angiogenesis in vitro and in vivo by
inducing apoptosis and inhibiting the spreading and proliferation
of human lung tumor associated-endothelial cells (HLT-ECs).
Azithromycin also effectively inhibits lung tumor growth by
suppressing angiogenesis. Finally, the inhibitory effects of
azithromycin on angiogenesis are identified to be attributed to its
inhibition of VEGFR2-mediated downstream signaling pathways.
Materials and methods
HLT-EC, reagents and drugs
Primary HLT-EC was purchased from Cell Biologics,
Inc. (Chicago, IL, USA) and grown in Complete Human Endothelial
Cell Medium (Cell Systems Corporation, Kirkland, WA, USA). HLT-EC
used for the experiments were from passages 2–4 and starved in
endothelial cell medium (ECM, served as the basal medium; Cell
Systems Corporation) for 3 h prior to being treated with drugs.
Human lung cancer PC-9, A549, NCI-H69, DMS-53, H157 and EBC-1 cell
lines, were purchased from American Type Culture Collection
(Manassas, VA, USA) and cultured in Dulbecco's modified Eagle's
medium (DMEM; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
containing 10% fetal bovine serum (HyClone; GE Healthcare Life
Sciences, Logan, UT, USA), 1 mM sodium pyruvate, 2 mM L-glutamine
and 1% penicillin-streptomycin (Thermo Fisher Scientific, Inc.).
Recombinant human VEGF165 was purchased from R&D
Systems, Inc. (Minneapolis, MN, USA). Azithromycin (Sigma-Aldrich;
EMD Millipore, Billerica, MA, USA) was dissolved in dimethyl
sulfoxide.
In vitro capillary network
formation
The HLT-ECs (2×104/well), together with
various concentrations of azithromycin, were seeded onto solidified
Matrigel matrix (BD Biosciences, Franklin Lakes, CA, USA) in a
96-well plate. Following 6 h of incubation at 37°C, capillary
network formation was analyzed under light microscopy (Zeiss GmbH,
Jena, Germany). The total tube length was quantified using NIH
Image J 1.32 software (National Institutes of Health, Bethesda, MD,
USA).
Transwell migration assay
A Transwell migration assay (Cell Biolabs, Inc., San
Diego, CA, USA) was performed using 6.5 mm diameter polycarbonate
filters pre-coated with 0.1% gelatin. The HLT-ECs
(4×104/well) were seeded into the upper chambers of the
Transwell plates, and subsequently treated with azithromycin. ECM
with or without 20 ng/ml VEGF was placed into the lower chamber.
Following 4–6 h of incubation at 37°C, cells spreading on the upper
surfaces of the filter (non-migrated cells) were wiped away with
cotton swabs, and the migrated cells on the lower surface of the
filter were fixed and stained with 0.4% Giemsa (Sigma-Aldrich; EMD
Millipore).
Cell spreading assay
HLT-ECs together with various concentrations (1, 5
and 10 µM) of azithromycin were seeded onto a 20X diluted
Matrigel-coated 96-well-plate for 2 h. The attached cells were then
photographed under light microscopy. Images were taken using
magnification ×20 under phase-contrast.
Measurement of proliferation and
apoptosis
HLT-ECs were treated with azithromycin in ECM medium
with or without 20 ng/ml VEGF. Lung cancer cells were treated with
azithromycin in DMEM medium. Following 2 days of treatment,
cellular proliferation activity was measured by the CellTiter
96® aqueous one solution cell proliferation assay kit
(Promega Corporation, Madison, MI, USA). To measure cell apoptosis,
cells were stained with Annexin V-fluorescein isothiocyanate (FITC)
and then analyzed on a Beckman Coulter FC500 flow cytometer. The
percentage of Annexin V-positive cells was determined by CXP
software analysis (Beckman Coulter, Inc., Brea, CA, USA).
Denaturing SDS-PAGE and western blot
(WB) analysis
HLT-ECs were treated with azithromycin in
endothelial cell medium with or without 20 ng/ml VEGF for 30 min
and subsequently lysed using a radioimmunoprecipitation assay
(RIPA) buffer (Thermo Fisher Scientific, Inc.) to extract total
protein. Frozen tumor tissues were homogenized using a polytron
homogenizer in ice-cold RIPA buffer for 10 min. Equal amounts (10
µg) of protein from cell extracts or tumor tissues were resolved
using denaturing 10–15% SDS-PAGE and analyzed by WB analysis.
Antibodies (dilution, 1:1,000) used in WB analyses included
anti-p-VEGFR2 (no. 2478), anti-VEGFR2 (no. 2479), anti-VEGF (no.
2463), anti-hypoxia-inducible factor (HIF; no. 3716),
anti-phosphorylated (p)-focal adhesion kinase (FAK; no. 3283),
anti-FAK (no. 3285), anti-p-phosphatidylinositol 3-kinase (PI3K;
no. 4228), anti-PI3K (no. 4292), anti-p-protein kinase B (Akt; no.
4060), anti-Akt (no. 9272) and anti-actin (no. 4967; all from Cell
Signaling Technology, Inc., Danvers, MA, USA). The membranes were
incubated with primary antibodies at 4°C overnight.
Directed in vivo angiogenesis
assay
The in vivo angiogenesis in Matrigel plug was
determined using the directed in vivo angiogenesis assay kit
(Trevigen, Inc., Gaithersburg, MD, USA) (19). The angioreactors were filled with
basement membrane extracts alone, in combination with VEGF (20
ng/ml)/basic fibroblast growth factor (bFGF; 30 ng/ml) or with
VEGF/bFGF in combination with 1, 5 or 10 µM azithromycin, and
incubated at 37°C for 1 h to allow gelling. The angioreactors
containing total 20 µl volume were implanted under the skin of
right flank of severe combined immunodeficiency mice for two weeks.
All work was conducted with the formal approval of the Jianghan
University Animal Care Committee (Jianghan, China) and strictly
followed the ethical guidelines on the care and use of animals.
Male SCID mice with 20–25 g were purchased from Animal Resources
Centre Australia (Murdoch, WA, Australia) and housed in specific
pathogen-free rooms in animal holding unit of Jianghan University.
A maximum of 5 mice were maintained in a cage with water bottle and
overhead food hopper. A total of 25 mice at 6-week old were used.
The invaded endothelial cells were isolated and labeled with
FITC-lectin. The quantification of FITC-lectin labeled endothelial
cells was performed using a SPECTRAmax microplate
spectrofluorometer (excitation 485 nm and emission 510 nm;
Molecular Devices, LLC, Sunnyvale, CA, USA).
Lung xenograft mouse tumor in SCID
mice and blood vessel analysis
All procedures were conducted according to the
guidelines approved by the Institutional Animal Care and Use
Committee. SCID mice at 6 weeks old were purchased from Animal
Resources Centre Australia (Murdoch, WA, Australia). In total 20
male mice (20–25 g) were used in the present study. Mice were
housed in pathogen-free room with temperature ~25°C. A maximum of 5
were maintained in a cage with water bottle and overhead food
hopper. A 14/10 h light/dark cycle is used. Lung tumor A549 cells
at 1×106/100 µl PBS were injected into the flank of each
mouse. When the tumor volume reached ~200 mm3, the mice
were treated with vehicle control (20/80%, DMSO/Saline) or
intraperitoneal azithromycin at 20 mg/kg daily (n=10 per group).
Tumor length and width was measured every three days and the volume
was calculated as: Length × width2 × 0.5. For tumor
blood vessel analysis, tumor frozen section slides (5 µm) were
fixed with 4% paraformaldehyde (Sigma-Aldrich; EMD Millipore,
Billerica, MA, USA) and stained with 1:200-diluted anti-CD31
antibody (no MA3100) followed by 1:500-diluted Alex-fluor
488-conjugated 2nd antibody (no A-11034; Molecular Probes; Thermo
Fisher Scientific, Inc.). The nuclei were stained with 4, 6-DAPI
(Molecular Probes; Thermo Fisher Scientific, Inc.). Blood vessels
were viewed with ×40 magnification under confocal microscopy (Zeiss
GmbH). For vessel quantification, the average number of vessels per
microscopic field from 3 microscopic fields per tumor section were
analyzed.
Analysis of actin stress fibers and
focal adhesions in HLT-ECs
HLT-ECs were treated with azithromycin in ECM medium
with or without 20 ng/ml VEGF for 60 min at 37°C. Cells were fixed
with 4% formalin for 10 min and washed with 1X PBS. Cells were then
permeabilized with 0.5% Triton X-100 for 10 min and stained with
1:500 diluted anti-paxillin antibody (no sc-373880) and 1:500
diluted FITC-conjugated immunoglobulin G (no sc-20052; both from
Santa Cruz Biotechnolgy, Inc., Dallas, TX, US).
Tetramethylrhodamine-conjugated phalloidin (Sigma-Aldrich; EMD
Millipore) was used to stain the actin cytoskeleton.
Statistical analysis
All data are expressed as the mean ± standard
deviation to indicate data variability. Statistical analyses were
performed using Student's unpaired t-test. A one way analysis of
variance was used for comparison in instances of multiple
parameters. All statistical analyses were performed using the
Graphpad Prism v6.0 software (GraphPad Software, Inc., La Jolla,
CA, USA). P<0.05 was considered to indicate a statistically
significant difference.
Results
Azithromycin effectively inhibits lung
tumor angiogenesis in vitro and in vivo
To investigate the effects of azithromycin in lung
tumor angiogenesis, in vitro and in vivo angiogenesis
assays were performed using HLT-ECs isolated from human lung
tumors. ECs can rapidly align and form tubular structures within 6
h when cultured on complete Matrigel matrix, which is rich in
extracellular matrix and various growth factors, including VEGF and
bFGF (20).
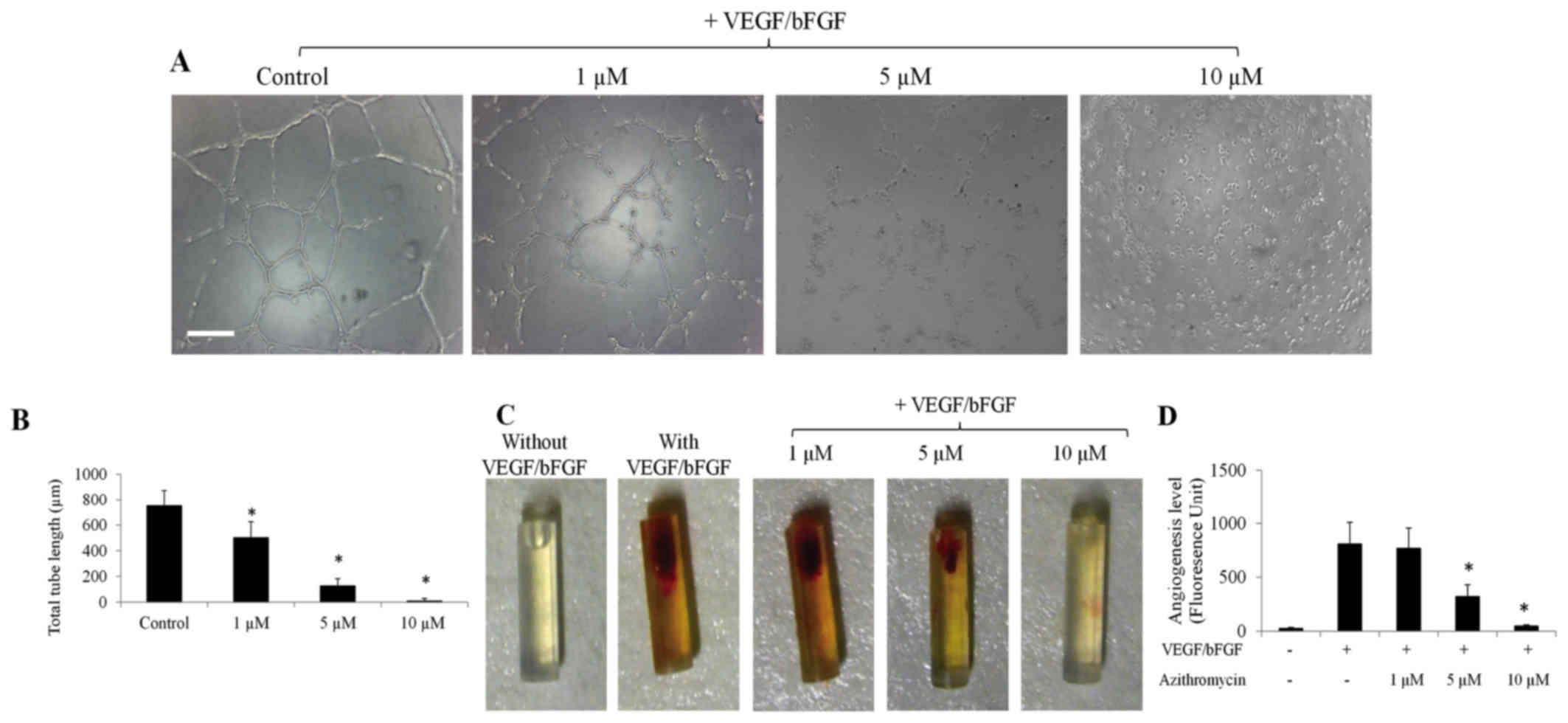
Azithromycin was revealed to effectively inhibit
capillary network formation of HLT-ECs on complete Matrigel matrix
in a dose-dependent manner (Fig. 1A and
B). The effector concentration for half maximal response of
azithromycin was as low as ~3 µm (Fig.
1B). It was then examined whether azithromycin could inhibit
angiogenesis in vivo using a modified basic Matrigel plug
angiogenesis assay (19). Basic
Matrigel together with drugs were retained in a silicon tube and
implanted subcutaneously into the mouse for two weeks. As shown in
Fig. 1C, a mixture of VEGF and bFGF
induced potent angiogenesis compared with the control (basic
Matrigel alone). When azithromycin was added together with
VEGF/bFGF, it suppressed VEGF/bFGF-induced angiogenesis in a
dose-dependent manner, as assessed by measuring the number of ECs
using fluorescently labeled EC-binding lectin (Fig. 1D). Collectively, the present results
demonstrate that azithromycin acts as an angiogenesis inhibitor
in vitro and in vivo.
Azithromycin suppresses spreading,
proliferation and induces apoptosis of HLT-ECs without affecting
HLT-EC migration
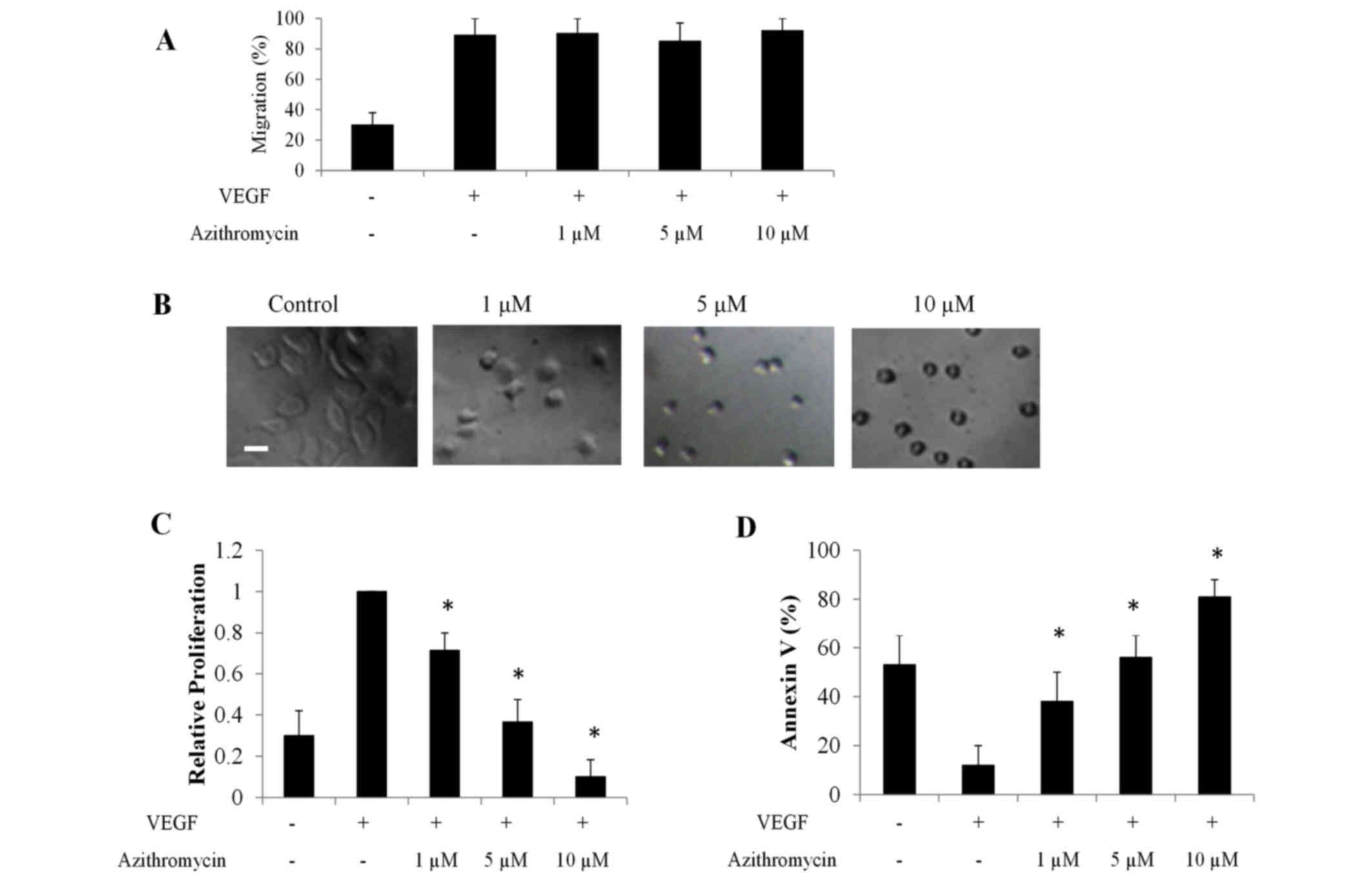
Capillary network formation is a multi-step and
dynamic process involving cell attachment to matrix, migration,
spreading, morphogenesis and apoptosis (21). Therefore, migration, spreading,
proliferation and apoptosis assays were performed using HLT-ECs to
understand how azithromycin interfered with capillary network
formation of lung tumor ECs. VEGF is the most important EC specific
angiogenic growth factor, stimulating various aspects of
angiogenesis (22); therefore, the
effects of azithromycin on VEGF-stimulated angiogenesis were
investigated.
Azithromycin was revealed to exhibit no effect on
VEGF-induced HLT-ECs migration (Fig.
2A). By contrast, azithromycin significantly inhibited HLT-EC
spreading to diluted complete Matrigel [composed of laminin,
collage type IV, heparan sulfate proteoglycans and entactin
(23); Fig.
2B]. VEGF is known to stimulate proliferation of ECs and
protect ECs against apoptosis (22).
Results consistently demonstrated that there were more
proliferating and less apoptotic HLT-ECs in the presence of VEGF
compared with the control (basic medium without VEGF; Fig. 2C and D). Notably, azithromycin
inhibited VEGF-induced proliferation of HLT-ECs and induced
apoptosis in a dose-dependent manner, even in the presence of VEGF
(Fig. 2C and D). Compared with
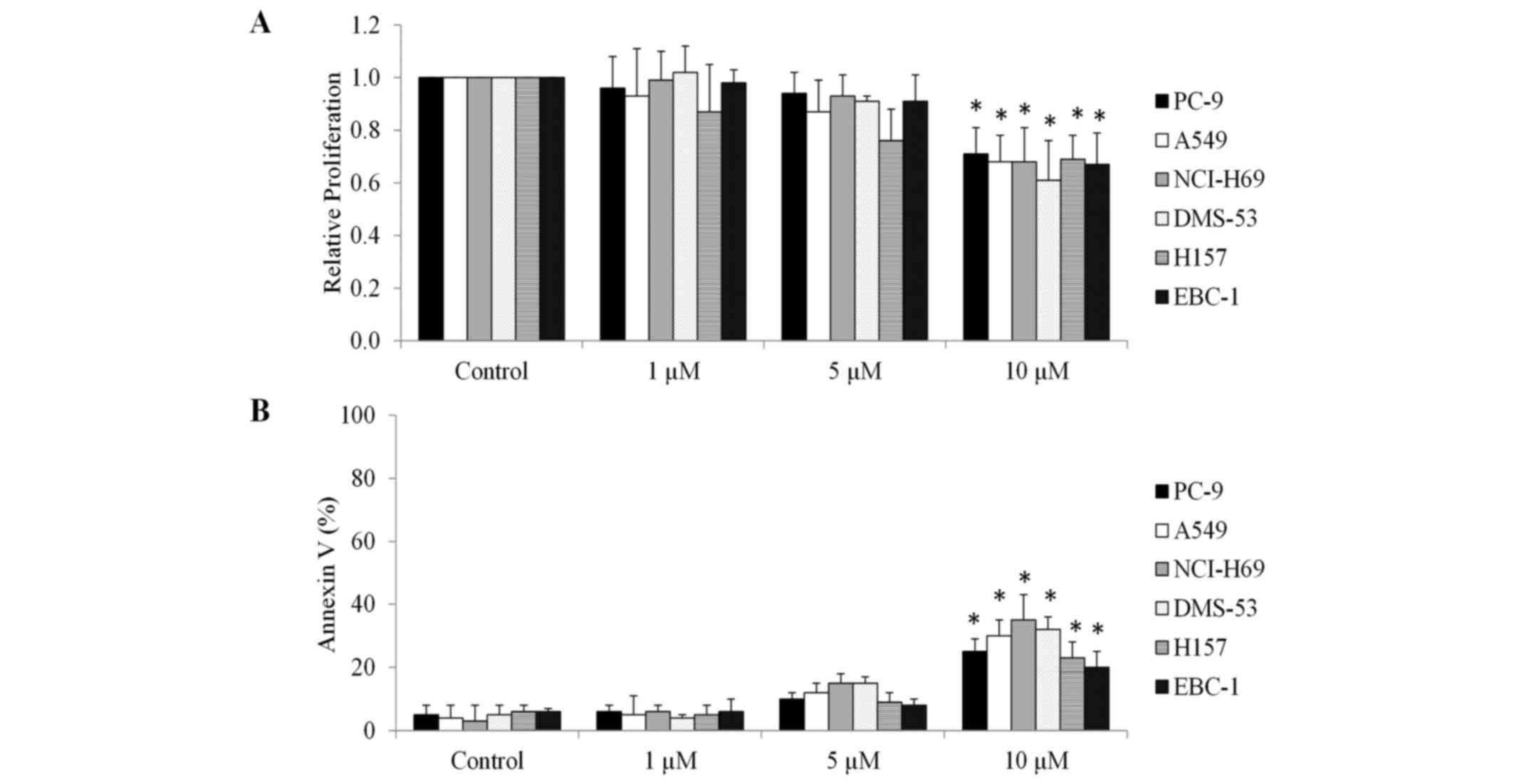
HLT-ECs, azithromycin at the same concentration (10 µm) inhibited
proliferation and induced apoptosis of multiple lung cancer cell
lines, including PC-9, A549, NCI-H69, DMS-53, H157 and EBC-1, to a
significantly reduced extent (Fig. 3A and
B), suggesting that HLT-ECs were more susceptible compared with
lung cancer cells to azithromycin treatment.
Azithromycin inhibits tumor
angiogenesis by suppressing vascular endothelial growth factor
receptor 2 (VEGFR2) -mediated signaling pathways
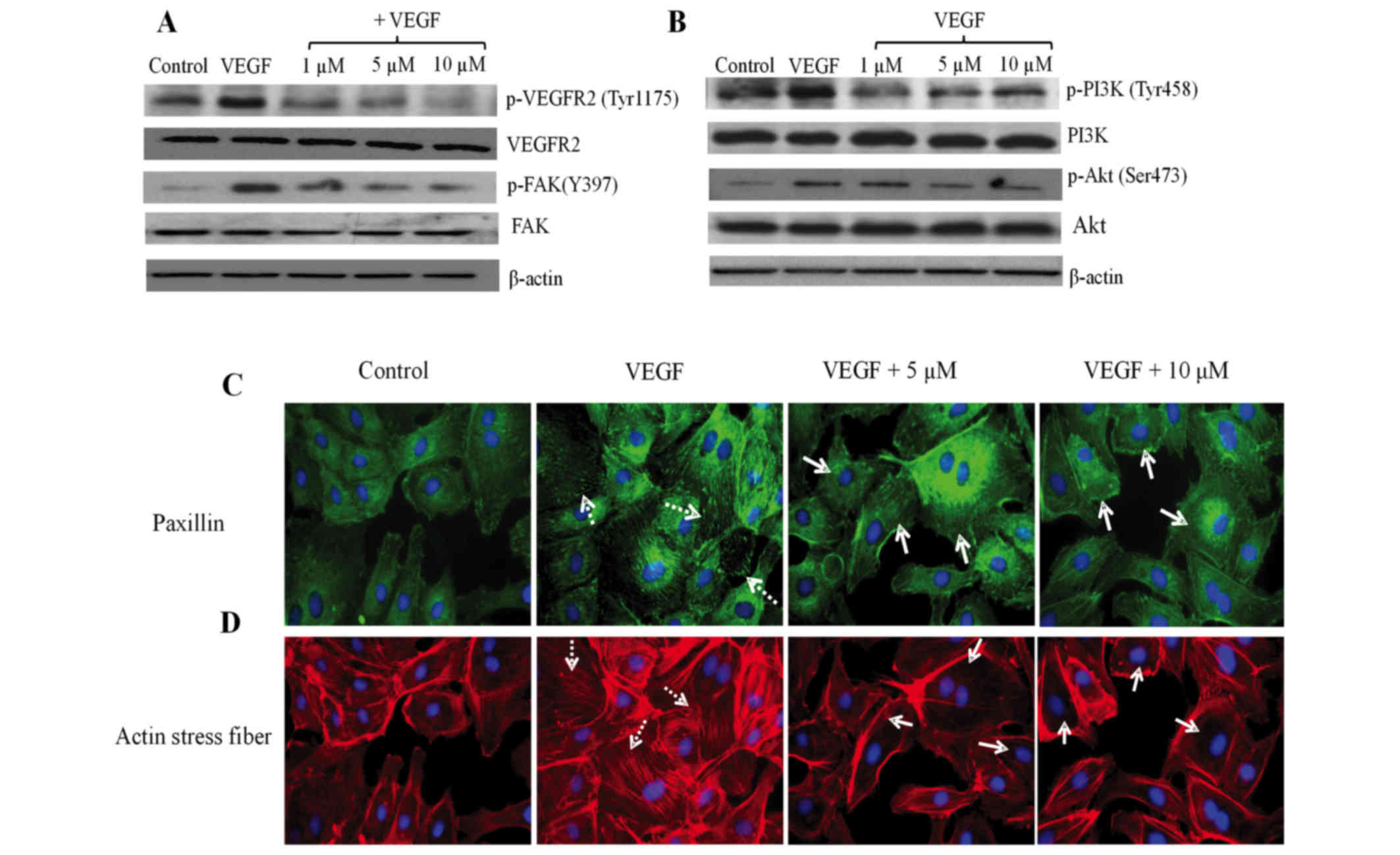
To understand the molecular basis of
azithromycin-mediated anti-angiogenic effects, the action of
azithromycin on the phosphorylation of VEGFR2 was firstly examined,
since VEGFR2 is the crucial and main receptor mediating
VEGF-stimulated angiogenic activities (24). Azithromycin inhibited VEGFR2
activation by suppressing phosphorylation of VEGFR2 at Tyr1175 at
1, 5 and 10 µm in HLT-ECs (Fig. 4A),
which was consistent with its in vitro and in vivo
functions. As a consequence of the deactivation of VEGFR2,
suppressed phosphorylation of PI3K, Akt and FAK was observed in
azithromycin-treated HLT-ECs (Fig. 4A and
B).
FAK is an essential component of the focal adhesion
complex that attaches cells to the extracellular matrix, and is
important for cell spreading and survival (25). Activated FAK then activates and
recruits paxillin into the focal adhesion complex, followed by an
accumulation of actin stress fibers to form proper focal adhesion
complexes. Consistent with inhibition of HLT-EC spreading and FAK
activation (Figs. 2B and 4A), azithromycin effectively blocked
VEGF-induced paxillin recruitment into focal adhesions, as shown by
the reduced paxillin staining (Fig.
4C) and VEGF-induced stress fiber formation (Fig. 4D).
The above experiments associated with each other to
reveal that inhibition of HLT-EC spreading, proliferation and
survival by azithromycin is linked with its ability to suppress the
VEGF-VEGFR2-mediated PI3K/Akt signaling pathway, FAK
phosphorylation, paxillin relocation into focal adhesions and actin
stress fiber formation.
Azithromycin inhibits lung tumor
growth and tumor angiogenesis in xenograft mice
It was then investigated whether azithromycin exerts
inhibitory effects on tumor angiogenesis in vivo and tumor
growth using a A549 lung tumor xenograft mouse model. The mice
tolerated 20 mg/kg azithromycin well, as no significant body weight
loss was observed (data not shown). Azithromycin was revealed to
significantly suppressed lung tumor growth (Fig. 5A).
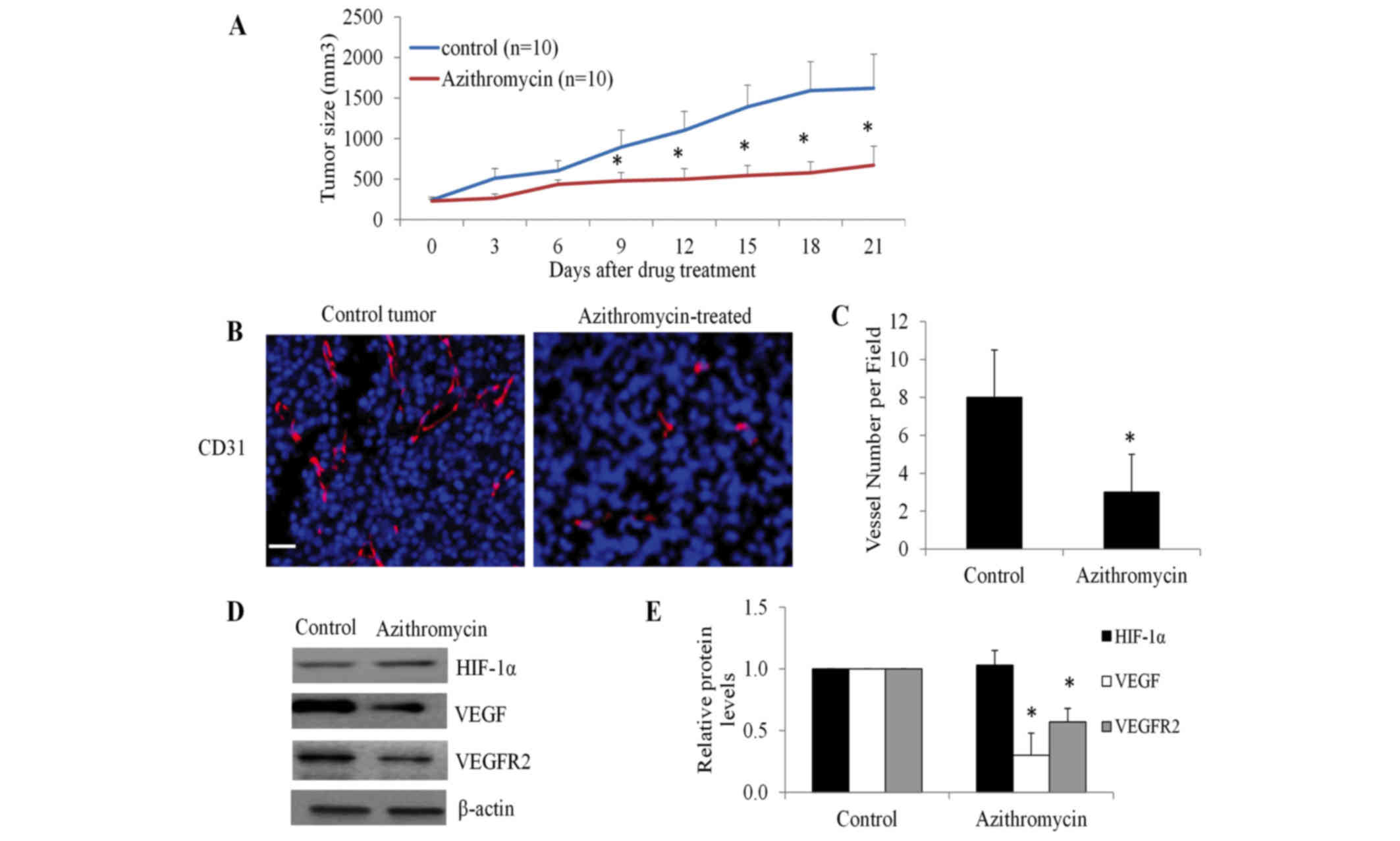 | Figure 5.Azithromycin inhibits lung tumor
growth and tumor angiogenesis in xenograft mice. (A) Azithromycin
significantly inhibits A549 lung tumor growth. SCID mice bearing
A549 lung tumor xenografts at the flanks were treated with equal
volume of vehicles, azithromycin at 20 mg/kg daily by
intraperitoneal injection (n=10). (B) Azithromycin significantly
inhibits tumor angiogenesis. Tumor blood vessels were visualized by
CD31 staining. Nucleus was stained blue by
4,6-diamidino-2-phenylindole. Scale bar represents 20 µm. (C) The
average number of vessels were analyzed from three microscopic
fields per tumor section, three tumor sections per tumor and ten
tumors for each experiment group. (D and E) Decreased protein
levels of VEGF and VEGFR2, but not HIF-1α, were observed in
azithromycin-treated tumors. Representative western blot analysis
images of tumor tissues were captured and presented. The protein
levels of HIF-1α, VEGF and VEGFR were quantified using Image-J
software with β-actin levels for normalization. *P<0.05,
compared with the control. CD31, cluster of differentiation 31;
VEGF, vascular endothelial growth factor; VEGFR2, VEGF receptor 2;
HIF-1α, hypoxia-inducible factor-1α. |
To examine whether the inhibitory effect of
azithromycin on lung tumor growth is attributable to decreased
tumor angiogenesis, blood vessel density and morphology of tumor
tissue sections was examined using anti-CD31 antibody, which
specifically stains tumor ECs (26).
A significant decrease in density of blood vessels was observed in
azithromycin-treated tumors compared with the control (Fig. 5B and C). Notably, the morphology of
the majority of blood vessels was non-lumen structures (cell
clusters and spots) in azithromycin-treated tumors, whereas blood
vessels in control tumors were all luminal structures (Fig. 5B). Consistently, decreased protein
levels of VEGF and VEGFR2, but not HIFα, were observed in
azithromycin-treated tumors (Fig.
5D). These data demonstrated that azithromycin inhibited lung
tumor growth through suppressing tumor angiogenesis.
Discussion
Combination therapies using angiogenesis inhibitors
and cytotoxic chemotherapeutic agents may achieve additional
anti-tumor efficacy as it targets two aspects of tumor development:
Tumor cell growth and vascularization. Angiogenesis has been the
target of major drug discovery programs, and angiogenesis
inhibitors with different mechanisms of actions have been evaluated
in clinical trials for cancer treatment over the last decade
(2,3).
To the best of our knowledge, the present study demonstrated for
the first time that azithromycin acts as a potential angiogenesis
inhibitor and may be repositioned from its traditional use in
infection disease to the therapy of lung cancer.
The primary endothelial cells, HLT-ECs (isolated
from human lung tumor), were selected to demonstrate the biological
effects of azithromycin on a lung tumor angiogenesis model.
Azithromycin was revealed to inhibit capillary network formation of
HLT-ECs in a dose-dependent manner (Fig.
1A and B), suggesting that azithromycin inhibits lung tumor
angiogenesis in vitro. VEGF is the most important angiogenic
factor that stimulates endothelial cell proliferation, migration,
tube formation and survival- all necessary components of an
angiogenesis response (6). With the
exception of migration, azithromycin inhibited all aspects of
VEGF-induced angiogenesis, as shown by decreased proliferation and
spreading, and increased apoptosis in HLT-ECs (Fig. 2). In accordance with in vitro
data, azithromycin potently inhibited VEGF/bFGF induced
angiogenesis in vivo (Fig. 1C and
D). These results demonstrate that azithromycin is an
angiogenesis inhibitor, and blocks multiple steps of
angiogenesis.
In addition, azithromycin significantly suppressed
VEGF-induced activation of VEGFR2, followed by the decreased
phosphorylation of PI3K/Akt and FAK, and disruption of focal
adhesion assembly and actin stress fiber formation in HLT-ECs
(Fig. 4). Inhibitors targeting the
PI3K/Akt were shown to decrease VEGF secretion and angiogenesis
(27,28). The present study supports previous
studies (27,28) and additionally demonstrates that
PI3K/Akt may perform essential roles for anti-angiogenic activities
of azithromycin. Bloomstein et al (2) reported that VEGF and its main receptor
VEGFR2 are two validated molecular targets for patients with NSCLC.
The ability of azithromycin to target VEGF and VEGFR2 makes it an
attractive addition to the armamentarium in the treatment of
NSCLC.
Notably, azithromycin has a potent ability to
inhibit human lung tumor growth, an effect achieved to a great
extent by angiogenesis suppression. In xenograft lung tumor mice,
azithromycin significantly inhibited tumor growth throughout the
treatment (P=0.023; Fig. 5A).
Vascularization analysis revealed that the generation of new blood
vessels and the expression of VEGF and VEGFR2 in
azithromycin-treated mice tumors were significantly decreased
compared with the control (Fig.
5B-E), demonstrating that azithromycin inhibits angiogenesis in
lung tumors. This finding supports the previous studies that
angiogenesis inhibitors alone can inhibit tumor growth (29). Notably, azithromycin has been reported
to exhibit inhibitory effects on tumor cell proliferation (16). It is possible that the inhibition of
lung tumor growth occurs through the effects of azithromycin on the
biological function of endothelial cells and/or tumor cells. The
present data demonstrated that HLT-ECs were more susceptible
compared with lung cancer cells to azithromycin treatment (Fig. 3), suggesting that endothelial cells
were possibly the primary target of azithromycin in lung tumor
growth inhibition.
In conclusion, to the best of our knowledge, the
present study reports for the first time that azithromycin is a
novel angiogenesis inhibitor. It potently inhibits angiogenesis
in vitro and in vivo through suppressing multiple
aspects of VEGF-induced angiogenic response. The molecular
mechanisms of the action of azithromycin on HLT-ECs are attributed
to its inhibition of VEGFR2-mediated downstream signaling pathways.
Notably, azithromycin effectively inhibits lung tumor growth via
suppressing angiogenesis. These findings suggest that azithromycin
may be translated into clinical trials for lung cancer treatment
and also emphasize the therapeutic value of angiogenesis inhibition
in lung cancer.
Acknowledgements
The present study was supported by a research grant
provided by Wuhan No. 6 Hospital, Affiliated Hospital to Jianghan
University (grant no. WH6201206006) and the National Foundation
Research Grant of China (grant no. 81300413).
References
|
1
|
Dancey J and Le Chevalier T: Non-small
cell lung cancer: An overview of current management. Eur J Cancer.
33 Suppl 1:S2–S7. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Blumenschein GR Jr, Reckamp K, Stephenson
GJ, O'Rourke T, Gladish G, McGreivy J, Sun YN, Ye Y, Parson M and
Sandler A: Phase 1b study of motesanib, an oral angiogenesis
inhibitor, in combination with carboplatin/paclitaxel and/or
panitumumab for the treatment of advanced non-small cell lung
cancer. Clin Cancer Res. 16:279–290. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lind JS and Smit EF: Angiogenesis
inhibitors in the treatment of non-small cell lung cancer. Ther Adv
Med Oncol. 1:95–107. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hanahan D and Folkman J: Patterns and
emerging mechanisms of the angiogenic switch during tumorigenesis.
Cell. 86:353–364. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Folkman J: Tumor angiogenesis: Therapeutic
implications. N Engl J Med. 285:1182–1186. 1971. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ferrara N: The role of vascular
endothelial growth factor in pathological angiogenesis. Breast
Cancer Res Treat. 36:127–137. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Han H, Silverman JF, Santucci TS, Macherey
RS, d'Amato TA, Tung MY, Weyant RJ and Landreneau RJ: Vascular
endothelial growth factor expression in stage I non-small cell lung
cancer correlates with neoangiogenesis and a poor prognosis. Ann
Surg Oncol. 8:72–79. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fontanini G, Bigini D, Vignati S, Basolo
F, Mussi A, Lucchi M, Chine S, Angeletti CA, Harris AL and
Bevilacqua G: Microvessel count predicts metastatic disease and
survival in non-small cell lung cancer. J Pathol. 177:57–63. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Horn L and Sandler AB: Angiogenesis in the
treatment of non-small cell lung cancer. Proc Am Thorac Soc. 6:pp.
206–217. 2009; View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhao L, Li W, Zhang H, Hou N, Guo L and
Gao Q: Angiogenesis inhibitors rechallenge in patients with
advanced non-small-cell lung cancer: A pooled analysis of
randomized controlled trials. Onco Targets Ther. 8:2775–2781.
2015.PubMed/NCBI
|
|
11
|
Zhang TT, Wang RM, Yang Z and Chen GB:
Dual inhibiting EGFR and VEGF pathways versus EGFR-TKIs alone in
the treatment of advanced non-small-cell lung cancer: A
meta-analysis of randomized controlled trials. Clin Transl Oncol.
18:576–581. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang J, Liu J, Chen H, Wu W, Li X, Wu Y,
Zhang K and Gu L: The impact of histological types on the efficacy
of angiogenesis inhibitors in the treatment of advanced NSCLC: A
meta-analysis of randomized controlled trials. Onco Targets Ther.
8:2375–2382. 2015.PubMed/NCBI
|
|
13
|
Esposito S, Bosis S, Faelli N, Begliatti
E, Droghetti R, Tremolati E, Porta A, Blasi F and Principi N: Role
of atypical bacteria and azithromycin therapy for children with
recurrent respiratory tract infections. Pediatr Infect Dis J.
24:438–444. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Champney WS and Burdine R: Macrolide
antibiotics inhibit 50S ribosomal subunit assembly in Bacillus
subtilis and Staphylococcus aureus. Antimicrob Agents Chemother.
39:2141–2144. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kitsiouli E, Antoniou G, Gotzou H,
Karagiannopoulos M, Basagiannis D, Christoforidis S, Nakos G and
Lekka ME: Effect of azithromycin on the LPS-induced production and
secretion of phospholipase A2 in lung cells. Biochim Biophys Acta.
1852:1288–1297. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lamb R, Ozsvari B, Lisanti CL, Tanowitz
HB, Howell A, Martinez-Outschoorn UE, Sotgia F and Lisanti MP:
Antibiotics that target mitochondria effectively eradicate cancer
stem cells, across multiple tumor types: Treating cancer like an
infectious disease. Oncotarget. 6:4569–4584. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhou X, Zhang Y, Li Y, Hao X, Liu X and
Wang Y: Azithromycin synergistically enhances anti-proliferative
activity of vincristine in cervical and gastric cancer cells.
Cancers (Basel). 4:1318–1332. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chu DJ, Yao DE, Zhuang YF, Hong Y, Zhu XC,
Fang ZR, Yu J and Yu ZY: Azithromycin enhances the favorable
results of paclitaxel and cisplatin in patients with advanced
non-small cell lung cancer. Genet Mol Res. 13:2796–2805. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Guedez L, Rivera AM, Salloum R, Miller ML,
Diegmueller JJ, Bungay PM and Stetler-Stevenson WG: Quantitative
assessment of angiogenic responses by the directed in vivo
angiogenesis assay. Am J Pathol. 162:1431–1439. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Madri JA and Pratt BM: Endothelial
cell-matrix interactions: In vitro models of angiogenesis. J
Histochem Cytochem. 34:85–91. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Davis GE and Senger DR: Endothelial
extracellular matrix: Biosynthesis, remodeling, and functions
during vascular morphogenesis and neovessel stabilization. Circ
Res. 97:1093–1107. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ferrara N: The role of VEGF in the
regulation of physiological and pathological angiogenesis. EXS.
209–231. 2005.PubMed/NCBI
|
|
23
|
Kleinman HK, McGarvey ML, Hassell JR, Star
VL, Cannon FB, Laurie GW and Martin GR: Basement membrane complexes
with biological activity. Biochemistry. 25:312–318. 1986.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shibuya M: Vascular endothelial growth
factor-dependent and -independent regulation of angiogenesis. BMB
Rep. 41:278–286. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lechertier T and Hodivala-Dilke K: Focal
adhesion kinase and tumour angiogenesis. J Pathol. 226:404–412.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Parums DV, Cordell JL, Micklem K, Heryet
AR, Gatter KC and Mason DY: JC70: A new monoclonal antibody that
detects vascular endothelium associated antigen on routinely
processed tissue sections. J Clin Pathol. 43:752–757. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Karar J and Maity A: PI3K/AKT/mTOR pathway
in angiogenesis. Front Mol Neurosci. 4:512011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kim GD, Oh J, Park HJ, Bae K and Lee SK:
Magnolol inhibits angiogenesis by regulating ROS-mediated apoptosis
and the PI3K/AKT/mTOR signaling pathway in mES/EB-derived
endothelial-like cells. Int J Oncol. 43:600–610. 2013.PubMed/NCBI
|
|
29
|
He L, Wu Y, Lin L, Wang J, Wu Y, Chen Y,
Yi Z, Liu M and Pang X: Hispidulin, a small flavonoid molecule,
suppresses the angiogenesis and growth of human pancreatic cancer
by targeting vascular endothelial growth factor receptor
2-mediatedPI3K/Akt/mTOR signaling pathway. Cancer Sci. 102:219–225.
2011. View Article : Google Scholar : PubMed/NCBI
|