Introduction
Hepatoblastoma is the most common type of pediatric
malignant liver tumor in childhood worldwide, and demonstrates an
increasing incidence; the incidence is estimated to be
1.2–1.5/million children per year, accounting for 80% of malignant
liver tumors in childhood (1–4). The type of tumor is usually diagnosed
prior to the age of 4 years, with poor prognosis and aggressive
behavior (5). Despite certain
previous studies investigating the metastasis mechanisms underlying
the HepG2 cell line (6–8), the mechanisms remain obscure. The
approaches to suppress the migration and invasion viability in
hepatoblastoma cells remain limited.
Previous studies have suggested that EMT is a
crucial process inducing cell cancer migration and invasion
(9,10). A growing number of tumors were
demonstrated to be involved in the EMT process when migrated and
invaded, including pancreatic cancer, colorectal cancer, prostate
cancer and breast cancer (11–14).
At the molecular level, a feature of EMT is the loss
or decreased expression level of epithelial cell markers,
E-cadherin (E-ca), and the upregulation of mesenchymal properties,
Vimentin and N-cadherin (N-ca) (15,16). The
loss of E-ca is considered to be the primary event of EMT, the
consequent loss of E-ca induces enhanced migration and invasion
potential of tumor cells (17).
Furthermore, the expression level of E-ca is often inversely
associated with tumor stage and grade (18,19).
Solamargine (SM) extracted from the Chinese herb
Solanum incanum L., is a major steroidal glycoalkaloid and
belongs to the Solanaceae (or nightshade) family (20). Our previous study demonstrated that SM
strongly inhibited growth and induced apoptosis of HCC cells, and
we further investigated mechanisms underlying apoptosis induced by
SM (21). Given this effect on the
proliferation and apoptosis of HCC cells, the present study aimed
to observe and determine whether SM contributes to an effective
action on metastasis in hepatoblastoma cells and explore the
possible underlying mechanisms.
Materials and methods
Materials
SM (purity >98%) was purchased from Yilin
Biotechnology Co., Ltd. (Shanghai, China). Fetal bovine serum (FBS)
and Dulbecco's modified Eagle's medium (DMEM) were obtained from
Gibco (Thermo Fisher Scientific, Inc., Waltham, MA, USA). MTT and
crystal violet staining solution were purchased from Beyotime
Institute of Biotechnology (Shanghai, China). The Transwell was
obtained from Corning Incorporated (Corning, NY, USA). Rabbit
anti-human monoclonal epithelial-cadherin (E-ca; 1:1,000 dilution;
cat. no. 3195S), rabbit anti-human monoclonal α-N-catenin (N-ca;
1:1,000 dilution; cat. no. 2163S), rabbit anti-human monoclonal
Vimentin (1:1,000 dilution; cat. no. 5741P) and mouse anti-human
monoclonal β-actin (loading control; 1:1,000 dilution; cat. no.
3700S) primary antibodies were purchased from Cell Signaling
Technology, Inc. (Danvers, MA, USA). Horseradish
peroxidase-conjugated goat anti-mouse (1:2,000 dilution; cat. no.
7072S) and goat anti-rabbit (1:2,000 dilution; cat. no. 7071S)
immunoglobulin G secondary antibodies were purchased from Cell
Signaling Technology, Inc. Other chemicals used in the present
study were commercial products of reagent grade.
Cell lines and culture
Human hepatoblastoma HepG2 cells were purchased from
the Cell Bank of the Chinese Academy of Sciences (Shanghai, China).
The HepG2 cells were maintained in DMEM, supplemented with 10% FBS
and 100 U/ml penicillin (Gibco; Thermo Fisher Scientific, Inc.), as
well as 100 µg/ml streptomycin (Gibco; Thermo Fisher Scientific,
Inc.) in a humidified atmosphere of 5% CO2 at 37°C.
Evaluation of cell viability
Cell viability was detected by MTT assay. HepG2
cells were seeded in 96-well plates (Corning Incorporated) at a
density of 2×105 and treated with serial concentrations of 0, 5, 10
or 20 µM SM for 24 h at 37°C. Following incubation, the medium
solution was removed and 20 µl of medium supplemented with MTT
reagent (5.0 mg/ml) was added to the well. Following a 4-h
incubation at 37°C, the medium solution was removed again.
Subsequently, 100 µl DMSO was added to each well and agitated for
15 min at 37°C. The absorbance at 490 nm (A490) was determined
using an ELISA reader (Bio-Rad 680; Bio-Rad Laboratories, Inc.,
Hercules, CA, USA). The cell survival ratio was evaluated based on
the treated group A490 results vs. the untreated group results.
Colony formation assay
For the colony formation assays, the HepG2 cells
were seeded into 6-well plates (Corning Incorporated) at a low
density of 500 cells per well. Subsequently, the cells were treated
with 0, 5, 10 or 20 µM SM and incubated for 2 weeks in a humidified
atmosphere of 5% CO2 at 37°C. Subsequently, the cells
were fixed with 4% paraformaldehyde for 30 min at room temperature
and stained with crystal violet for 15 min at room temperature,
followed with colony counting by eye. Images were captured using a
fluorescence microscope (Eclipse TS100; magnification, ×10; Nikon
Corporation, Tokyo, Japan).
Determination of migration using
wound-healing assays
In the wound-healing migration assays, cells were
seeded in 6-well plates at a density of 1×106 cells/well, incubated
for 24 h at 37°C and scratched using a yellow pipette tip when the
cells covered the well. The cells were washed with PBS twice to
clear the floating cells and various concentrations (0, 5, 10 or 20
µM) of SM were added. Images were captured at 12 and 24 h using a
fluorescence microscope at ×40 magnification.
Evaluation of migration by Transwell
assay
Cell migration was determined using the Transwell
Boyden chamber containing 8-µm pore size membranes (Corning
Incorporated). Briefly, suspended HepG2 cells were separately
treated with 0, 5, 10 or 20 µM SM in serum-free DMEM at a density
of 2×105 cells, and then added to the upper chamber of the
Transwell. DMEM supplemented with 10% FBS was added to the lower
chamber as a chemoattractant. Following incubation for 24 h at
37°C, non-invading cells remaining on the upper surface were
removed using a wet cotton swab, whereas cells on the lower surface
were fixed with 4% paraformaldehyde for 30 min at room temperature
and stained with crystal violet for 15 min at room temperature.
Images were captured using a fluorescence microscope at ×200
magnification.
Matrigel-coated Transwell assay to
detect invasion
In the cell invasion experiment, the Transwells were
coated with Matrigel (BD Biosciences, Franklin Lakes, NJ, USA).
Subsequent steps were similar to those described in the migration
assay protocol, and detailed procedures were performed as
previously described (22).
Western blot analysis
Western blot analysis was performed as described
previously (21), with minor
modifications. Cells were treated with 0, 5, 10 or 20 µM SM for 24
h. Cell lysates were collected using a lysis buffer (Beyotime
Institute of Biotechnology) and β-actin was used as the loading
control. The cell lysate proteins were separated by electrophoresis
based on their molecular weight, size and charge, and 40 µg/lane
cell lysate proteins were loaded and separated using 10% SDS-PAGE
by electrophoresis for 2 h at room temperature. They were then
transferred onto polyvinylidene difluoride membranes (EMD
Millipore, Billerica, MA, USA) for 90 min at −20°C, then incubated
with 3% bovine serum albumin for 1 h at room temperature.
Subsequently, the primary antibodies as aforementioned were added
and incubated overnight at 4°C. Following this, cells were
incubated with secondary antibodies as aforementioned for 1 h at
room temperature. Immunoreactive bands were detected using
chemiluminescence reagent (EMD Millipore) and all the blots were
quantified using LANE 1D software version 1 (Sage Creation Science
Co., Ltd., Beijing, China). The experiments were performed ≥3
times.
Statistical analysis
Data are presented as the mean ± standard deviation,
and the differences between two groups were analyzed using
Student's t-test. Statistical analyses were performed using SPSS
16.0 (SPSS, Inc., Chicago, IL, USA). P<0.05 was considered to
indicate a statistically significant difference. All the
experiments were performed in at least in triplicate.
Results
Proliferation and viability of HepG2
cells is suppressed by SM
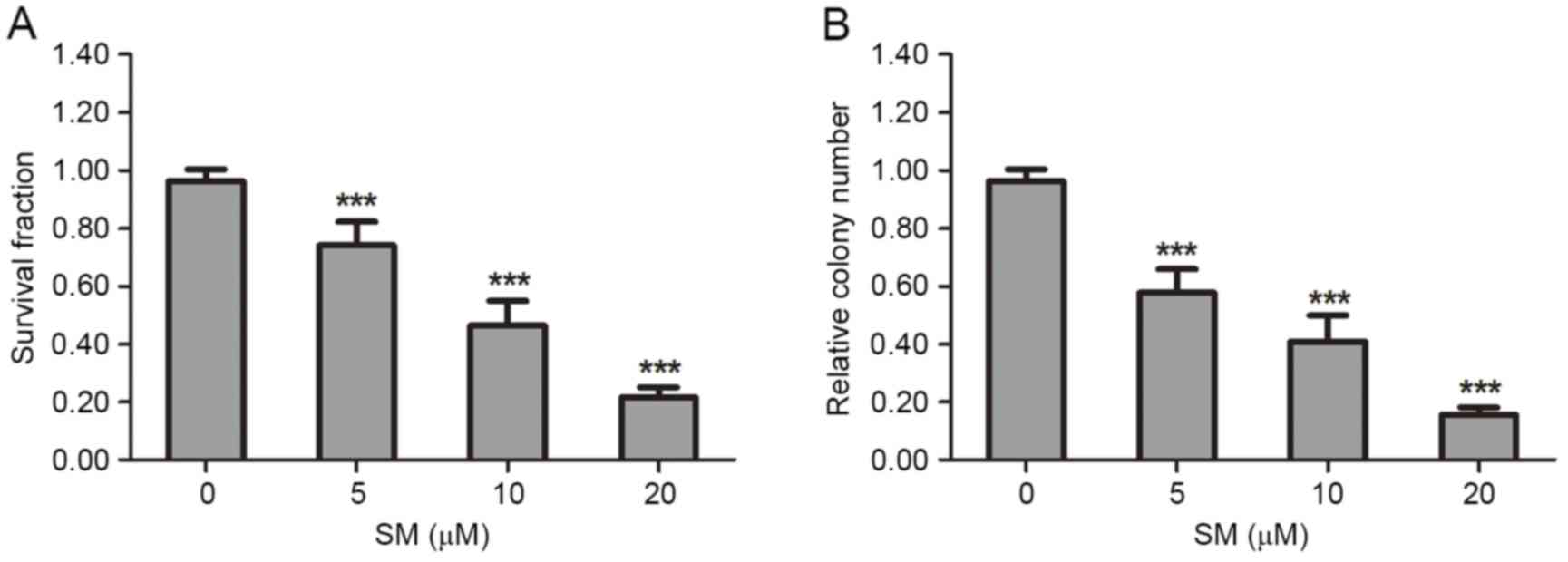
The suppressive effect of SM on HepG2 cell viability
and proliferation was determined by MTT and colony formation
assays. In the MTT analysis, HepG2 cells were treated with various
concentrations (0, 5, 10 or 20 µM) of SM for 24 h. The results
revealed that the survival fraction of HepG2 cells was affected in
a dose-dependent manner. At 20 µM SM the survival rate (21.7±3.5%)
was the lowest (Fig. 1A). In the
colony formation experiments, each well was seeded with 500 HepG2
cells, then incubated for 2 weeks and stained, and the colony
number was then counted. Following exposure to 5, 10 or 20 µM SM,
the relative colony numbers were 57.7±4.3, 38.1±4.9 and 15.7±2.5%
compared with the control group (P<0.001). The colony number
decreased in a dose-dependent manner (Fig. 1B).
Migration of HepG2 cells is attenuated
by SM treatment
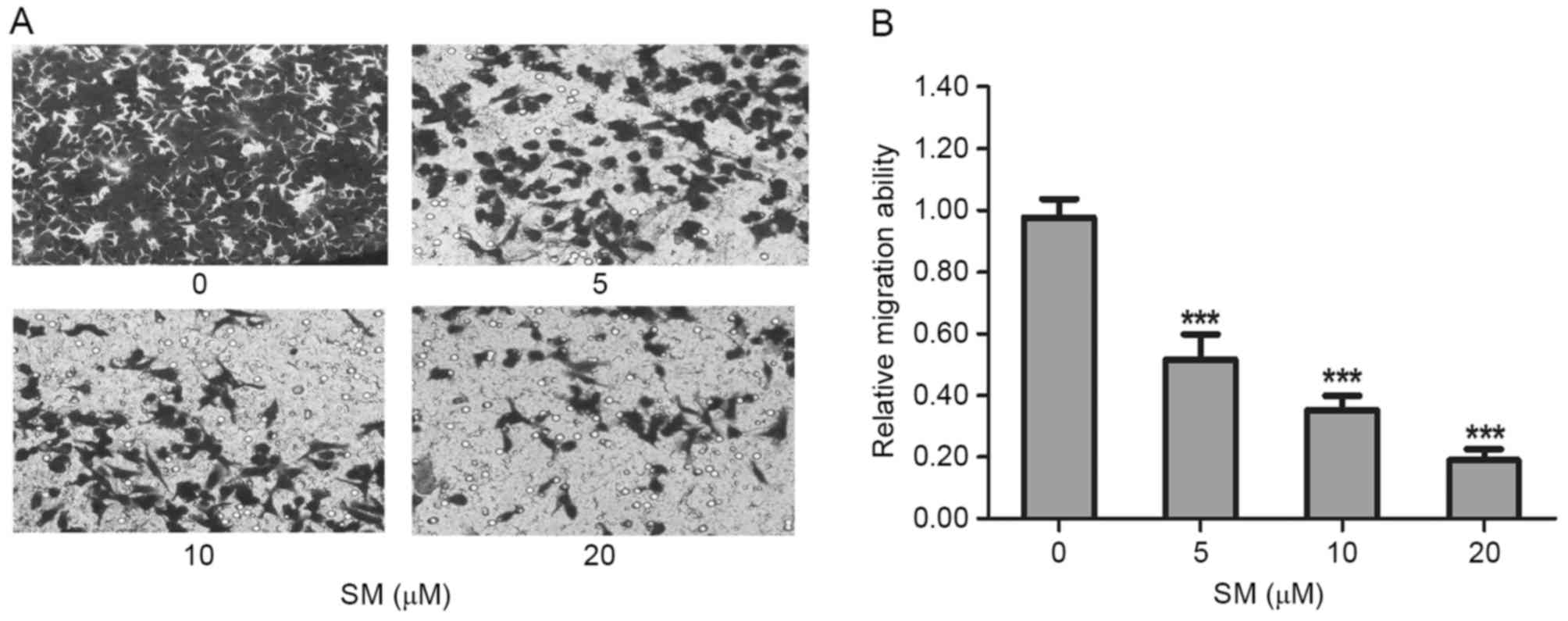
The migration was evaluated by wound-healing and
Transwell assays. In the wound-healing assay, HepG2 cells were
treated with 0, 5, 10 or 20 µM SM supplemented with 1% FBS.
Following 12 and 24 h, the width of the wound exhibited a lower
propensity for closure compared with the that of the untreated
cells. The untreated HepG2 cells filled the majority of the wounded
area after 24 h, whereas a distinct gap remained in the SM-treated
groups after 24 h, and the gaps were affected in a dose-dependent
manner (Fig. 2). Furthermore, after
24 h and treatment with 10 and 20 µM SM, respectively, the width of
the gaps were increased compared with the gap at 12 h (Fig. 2). To confirm the inhibition effects, a
Transwell assay was performed. HepG2 cells were treated with SM for
24 h, then stained, imaged and evaluated. Compared with the
untreated group, the mobility ratio of 5, 10 and 20 µM SM-treated
cells were 51.7±8.1, 35.3±4.5 and 19.0±3.6%, respectively (Fig. 3A). The relative migration ratios of
the SM-treated HepG2 cells revealed significant differences
compared with the untreated group (P<0.001; Fig. 3B).
Invasion of HepG2 cells is suppressed
by SM
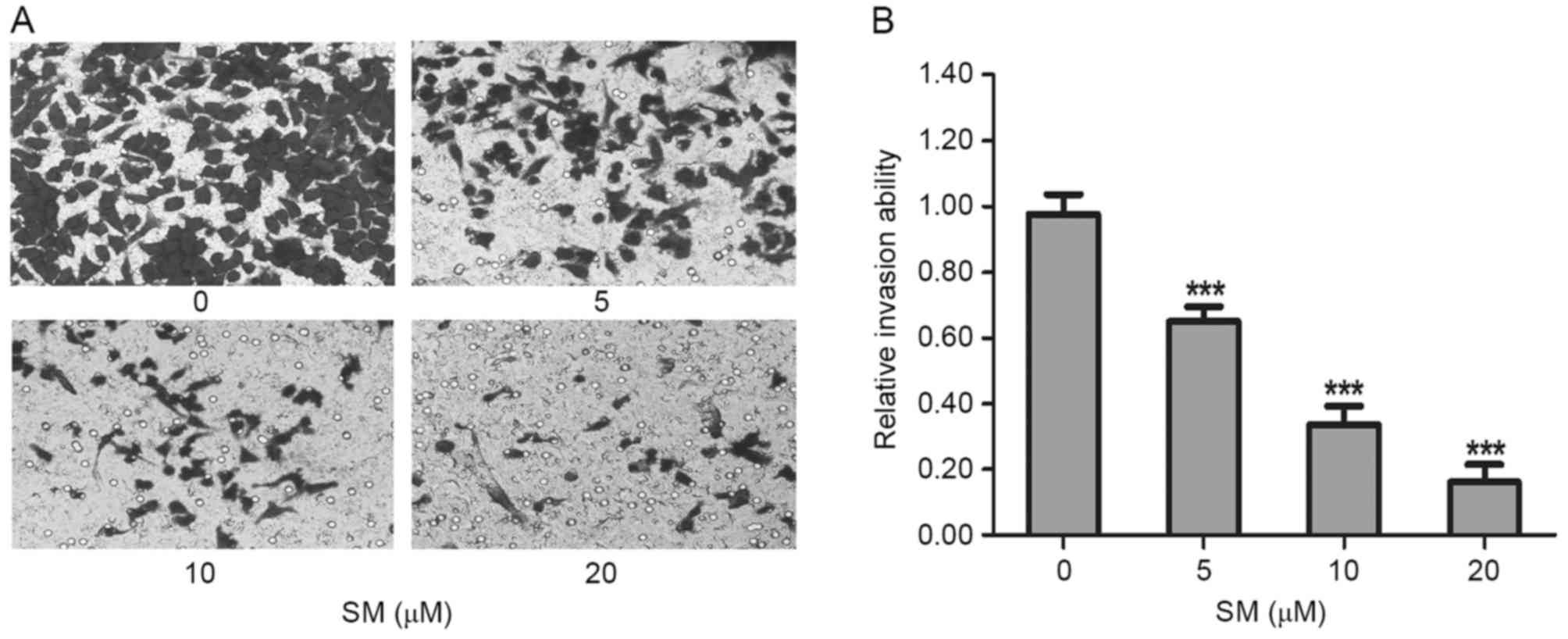
Cell invasive capacity was evaluated by
matrigel-coated Transwell assays. As presented in Fig. 4A, the invasive ability of the
SM-treated cells decreased significantly as the SM concentration
increased. Compared with the untreated cells, the relative invasion
ratios of 5, 10 and 20 µM SM-treated HepG2 cells were 65.0±4.6,
33.7±5.5 and 16.1±5.6%, respectively. There were significant
differences between the treated and untreated groups (P<0.001;
Fig. 4B). These results indicated
that SM inhibited the invasive capacity of HepG2 cells.
Changes in expression level of EMT
representative proteins in SM-treated cells
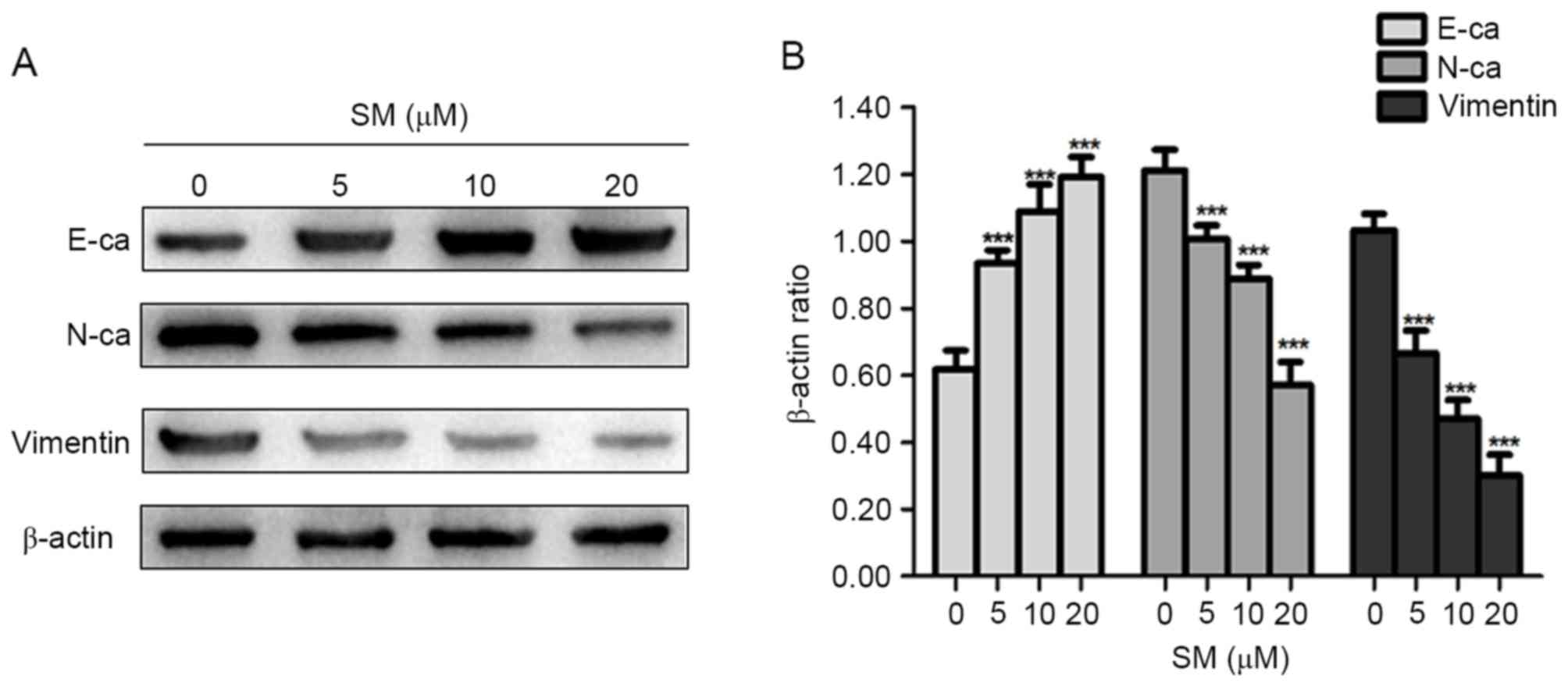
EMT is an important process in inducing cancer
migration and invasion, which is often associated with a loss or
decrease of E-ca, and an increase of Vimentin and N-ca expression
levels (15,16). Thus, the present study examined
whether EMT-associated protein expression levels changed in
SM-treated cells. As presented in Fig.
5, the epithelial marker, E-ca, was upregulated compared with
that in the control group, whereas the mesenchymal markers,
Vimentin and N-ca were consistently downregulated in the SM-treated
cells. Therefore, it was suggested that SM treatment decreased the
migration and invasion abilities in hepatoblastoma cells by
suppressing the process of EMT.
Discussion
Previously, Chinese herbs have been demonstrated to
contribute an effective function in the treatment of numerous
diseases and have attracted worldwide attention (23,24).
Certain previous studies have reported that they Chinese herbs
served important roles in regulating tumor cell behavior, including
proliferation, apoptosis and metastasis (25–27). SM,
extracted from Solanum incanum, is a traditional Chinese
herb that was demonstrated to possess an effective function in
inducing apoptosis of HepG2 and SMMC7721 cells in our previous
study (21). In the present study, as
conclusive results were not previously observed when investigating
metastasis in SMMC7721 cells, the effects of SM on the migration
and invasion on HepG2 cells were evaluated, and the concentration
and treated durations were used as referenced in our previous study
(21).
Previous studies have indicated that SM may suppress
cell proliferation in numerous cancer cell lines (28–30). In
the present study, the results of the MTT and colony formation
assays further confirmed this hypothesis showing that SM
effectively reduced cell proliferation in a concentration-dependent
manner in HepG2 cells.
The results of wound-healing and Transwell
experiments demonstrated that the migration ability of the HepG2
cells was decreased following treatment with various concentrations
of SM. Furthermore, as the doses of SM increased, the suppression
effect was enhanced. In the wound-healing assay, it was
hypothesized that the width of the gap would decrease in a
time-dependent manner, whereas after 24 h of treatment with 10 and
20 µM concentrations of SM, the width of the gap was increased
compared with the size of the gap following 12 h. It was considered
that this phenomenon was associated with the SM function in
triggering HepG2 cells death by apoptosis (21). Use of a Matrigel-coated Transwell
assay is a classical method to detect cell invasion, whereby the
invasive HepG2 cells would digest the gel and penetrate the upper
chamber to the lower wells. Following exposure to various
concentrations of SM, the number of HepG2 cells in the lower
chamber decreased greatly as the SM concentrations increased. The
Matrigel-coated Transwell experiment indicated that SM inhibited
the invasive viability of HepG2 cells efficiently.
Given the suppression effects of migration and
invasion revealed in HepG2 cells, the present study subsequently
investigated the underlying mechanism. EMT is a crucial event for
metastasis in numerous types of cancer (10,31–33), and
was first recognized as a feature of embryogenesis in the 1980s
(34). During the transformation from
the epithelial to the mesenchymal phenotype, epithelial cells lose
their polygonal shape and acquire a spindle-shaped morphology,
which provides the tumor cells enhanced motility and invasive
abilities (35–37). At the molecular level, numerous
previous studies have described the molecular pathways involved in
EMT, and revealed that it is characterized an increase in N-ca and
Vimentin, but particularly by the downregulation of E-ca (38,39). The
western blot analysis of the present study demonstrated that the
expression of E-ca was upregulated compared with the control group,
whereas N-ca and Vimentin were consistently downregulated in
SM-treated cells. Thus, it was suggested that SM inhibited the
migration and invasion of HepG2 cells by blocking EMT.
The present study aimed to reveal the function and
effects of SM in hepatoblastoma cells. However, as a traditional
Chinese herb, SM has various functions and further studies are
required to detect them. Taken together, the present findings
suggested that SM may inhibit the migration and invasion viability
of HepG2 cells by blocking EMT.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant no. 81502663), the
Social Development Foundation of Jiangsu Province (grant no.
BE2015668) and the Natural Science Foundation of Colleges and
Universities in Jiangsu Province (grant no. 14KJD310001).
References
|
1
|
Linabery AM and Ross JA: Trends in
childhood cancer incidence in the U.S. (1992–2004). Cancer.
112:416–432. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Fracp KJGA: Pathology of pediatric
gastrointestinal and liver disease. Second edition. Anticancer
Research. 35:234. 2015.
|
|
3
|
Mclaughlin CC, Baptiste MS, Schymura MJ,
Nasca PC and Zdeb MS: Maternal and infant birth characteristics and
hepatoblastoma. Am J Epidemiol. 163:818–828. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Finegold MJ, Lopez-terrada DH, Bowen J,
Washington MK and Qualman SJ; College of American Pathologists, :
Protocol for the examination of specimens from pediatric patients
with hepatoblastoma. Arch Pathol Lab Med. 131:520–529.
2007.PubMed/NCBI
|
|
5
|
Litten JB and Tomlinson GE: Liver tumors
in children. Oncologist. 13:812–820. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Liu Z, Wang J, Mao Y, Zou B and Fan X:
MicroRNA-101 suppresses migration and invasion via targeting
vascular endothelial growth factor-C in hepatocellular carcinoma
cells. Oncol Lett. 11:433–438. 2016.PubMed/NCBI
|
|
7
|
Dai W, Wang F, He L, Lin C, Wu S, Chen P,
Zhang Y, Shen M, Wu D, Wang C, et al: Genistein inhibits
hepatocellular carcinoma cell migration by reversing the
epithelial-mesenchymal transition: Partial mediation by the
transcription factor NFAT1. Mol Carcinog. 54:301–311. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bi Z, Liu W, Ding R, Wu Y, Dou R, Zhang W,
Yuan X, Liu X, Xiong L, Guo Z and Mao C: A novel peptide, 9R-P201,
strongly inhibits the viability, proliferation and migration of
liver cancer HepG2 cells and induces apoptosis by down-regulation
of FoxM1 expression. Eur J Pharmacol. 796:175–189. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Iwatsuki M, Mimori K, Yokobori T, Ishi H,
Beppu T, Nakamori S, Baba H and Mori M: Epithelial-mesenchymal
transition in cancer development and its clinical significance.
Cancer Sci. 101:293–299. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelial-mesenchymal transitions in development and disease.
Cell. 139:871–890. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Nagathihalli NS and Merchant NB:
Src-mediated regulation of E-cadherin and EMT in pancreatic cancer.
Front Biosci (Landmark Ed). 17:2059–2069. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bates RC and Mercurio AM: The
epithelial-mesenchymal tansition (EMT) and colorectal cancer
progression. Cancer Biol Ther. 4:365–370. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhu C, Li J, Cheng G, Zhou H, Tao L, Cai
H, Li P, Cao Q, Ju X, Meng X, et al: MiR-154 inhibits EMT by
targeting HMGA2 in prostate cancer cells. Mol Cell Biochem.
379:69–75. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Creighton CJ, Chang JC and Rosen JM:
Epithelial-mesenchymal transition (EMT) in tumor-initiating cells
and its clinical implications in breast cancer. J Mammary Gland
Biol Neoplasia. 15:253–260. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Thiery JP: Epithelial-mesenchymal
transitions in tumour progression. Nat Rev Cancer. 2:442–454. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Grünert S, Jechlinger M and Beug H:
Diverse cellular and molecular mechanisms contribute to epithelial
plasticity and metastasis. Nat Rev Mol Cell Biol. 4:657–665. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yang J, Mani SA, Donaher JL, Ramaswamy S,
Itzykson RA, Come C, Savagner P, Gitelman I, Richardson A and
Weinberg RA: Twist, a master regulator of morphogenesis, plays an
essential role in tumor metastasis. Cell. 117:927–939. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cowin P, Rowlands TM and Hatsell SJ:
Cadherins and catenins in breast cancer. Curr Opin Cell Biol.
17:499–508. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Junghans D, Haas IG and Kemler R:
Mammalian cadherins and protocadherins: About cell death, synapses
and processing. Curr Opin Cell Biol. 17:446–452. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lorey S, Porzel A and Ripperger H: Steroid
alkaloid glycosides from Solanum coccineum. Phytochemistry.
41:1633–1635. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xie X, Zhu H, Yang H, Huang W, Wu Y, Wang
Y, Luo Y, Wang D and Shao G: Solamargine triggers hepatoma cell
death through apoptosis. Oncol Lett. 10:168–174. 2015.PubMed/NCBI
|
|
22
|
Albini A, Iwamoto Y, Kleinman HK, Martin
GR, Aaronson SA, Kozlowski JM and McEwan RN: A rapid in vitro assay
for quantitating the invasive potential of tumor cells. Cancer Res.
47:3239–3245. 1987.PubMed/NCBI
|
|
23
|
Chen W, Lim CE, Kang HJ and Liu J: Chinese
herbal medicines for the treatment of type A H1N1 influenza: A
systematic review of randomized controlled trials. PLoS One.
6:e280932011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lam W, Bussom S, Guan F, Jiang Z, Zhang W,
Gullen EA, Liu SH and Cheng YC: The four-herb Chinese medicine
PHY906 reduces chemotherapy-induced gastrointestinal toxicity. Sci
Transl Med. 2:45ra592010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liu J, Shu Y, Zhang Q, Liu B, Xia J, Qiu
M, Miao H, Li M and Zhu R: Dihydromyricetin induces apoptosis and
inhibits proliferation in hepatocellular carcinoma cells. Oncol
Lett. 8:1645–1651. 2014.PubMed/NCBI
|
|
26
|
Jiang J and Hu C: Evodiamine: A novel
anti-cancer alkaloid from Evodia rutaecarpa. Molecules.
14:1852–1859. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jang SY, Lee JK, Jang EH, Jeong SY and Kim
JH: Shikonin blocks migration and invasion of human breast cancer
cells through inhibition of matrix metalloproteinase-9 activation.
Oncol Rep. 31:2827–2833. 2014.PubMed/NCBI
|
|
28
|
Sun L, Zhao Y, Li X, Yuan H, Cheng A and
Lou H: A lysosomal-mitochondrial death pathway is induced by
solamargine in human K562 leukemia cells. Toxicol In Vitro.
24:1504–1511. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li X, Zhao Y, Wu WK, Liu S, Cui M and Lou
H: Solamargine induces apoptosis associated with p53
transcription-dependent and transcription-independent pathways in
human osteo-sarcoma U2OS cells. Life Sci. 88:314–321. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ding X, Zhu FS, Li M and Gao SG: Induction
of apoptosis in human hepatoma SMMC-7721 cells by solamargine from
Solanum nigrum L. J Ethnopharmacol. 139:599–604. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Thompson EW, Newgreen DF and Tarin D:
Carcinoma invasion and metastasis: A role for
epithelial-mesenchymal transition? Cancer Res. 65:5991–5995. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kalluri R and Weinberg RA: The basics of
epithelial-mesenchymal transition. J Clin Invest. 119:1420–1428.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
33
|
Cardiff RD: Epithelial to mesenchymal
transition tumors: Fallacious or Snail's pace? Clin Cancer Res.
11:8534–8537. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Thiery JP: Epithelial-mesenchymal
transitions in development and pathologies. Curr Opin Cell Biol.
15:740–746. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Thiery JP and Sleeman JP: Complex networks
orchestrate epithelial-mesenchymal transitions. Nat Rev Mol Cell
Biol. 7:131–142. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
36
|
Klymkowsky MW and Savagner P:
Epithelial-mesenchymal transition: A cancer researcher's conceptual
friend and foe. Am J Pathol. 174:1588–1593. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Christiansen JJ and Rajasekaran AK:
Reassessing epithelial to mesenchymal transition as a prerequisite
for carcinoma invasion and metastasis. Cancer Res. 66:8319–8326.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
De Craene B and Berx G: Regulatory
networks defining EMT during cancer initiation and progression. Nat
Rev Cancer. 13:97–110. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lamouille S, Xu J and Derynck R: Molecular
mechanisms of epithelial-mesenchymal transition. Nat Rev Mol Cell
Biol. 15:178–196. 2014. View
Article : Google Scholar : PubMed/NCBI
|