Introduction
Cancer has long been recognized as a major causes of
mortality in humans. Bladder cancer, a heterogeneous neoplasm, is
either a low-grade tumor located in the superficial mucosa (80% of
cases) or a muscle-invasive carcinoma (20% of cases) (1). Urothelial carcinoma is the most common
type of bladder cancer and is a major cause of morbidity and
mortality (2). In Western
populations, bladder cancer represents the fourth most common
malignancy affecting males (3,4) and ranks
thirteenth in terms of cancer-associated mortality worldwide
(5,6).
The majority (>90%) of bladder tumors are of epithelial origin
and arise from the bladder urothelium (7). The 2014 report from the Ministry of
Health and Welfare of Taiwan indicated that bladder cancer was one
of the most common types of cancer in Taiwan in 2006, with 1,985
new cases and 681 mortalities (8).
Currently, the majority of treatments for superficial bladder
cancer are derived from natural products. The aim of this treatment
is to reduce tumor recurrence and prevent tumor progression.
Cantharidin (CTD), a natural toxin, is secreted by
beetles of the Mylabris genus (blister beetle) and has been
used as a traditional drug to treat Molluscum contagiosum viral
infections and cancer in China and Vietnam (9,10). For
treating superficial bladder cancer, the correct dosage of CTD is
important; an overdose of CTD may lead to mortality (11,12). A
number of studies have demonstrated that CTD induces cytotoxic
effects on numerous human cancer cell lines. CTD was demonstrated
to repress cancer cell growth through cell cycle arrest and the
induction of apoptosis in cancer cells, including pancreatic cancer
(13–15), leukemia U937 (16), human HepG2 (17), human colon cancer colo205 (18), human lung cancer A549 (19) and human bladder cancer (20) cells, in addition hepatocellular
carcinoma in vivo (21). It
has been reported that tamoxifen represses the phosphorylation of
protein kinase C (PKC) and amplifies the anticancer effect induced
by CTD (22). Previous studies have
revealed a CTD-induced cytotoxic effect in melanoma A375.S
(23), lung cancer NCI-H460 (24) and bladder cancer TSGH-8301 (25) cells.
Cancer arises from alterations in the structure,
expression and function of tumor suppressor genes (26). A number of genetic mutations have been
identified as biomarkers for the diagnosis and treatment of certain
types of human cancer, including glioblastoma multiforme, but
intratumoral heterogeneity presents challenges for personalized
treatment strategies (27).
Furthermore, genetic mutations in oncogenes and tumor suppressors
have been identified in numerous types of cancer cells (28,29). It
has been determined, through constructing a protein-protein
interaction network of differentially expressed genes and
co-expressed genes, that G2/M phase-specific cyclin B1
and H2A histone family member Z are significantly associated with
bladder cancer (30). A previous
study suggested that fibroblast growth factor receptor 2 gene
mutations in bladder cancer may only serve a minor role in bladder
carcinogenesis (31). Another
previous study demonstrated that CTD alters gene expression in
human lung cancer NCI-H460 cells in vitro (32). It has also been hypothesized that CTD
alters the gene expression of bladder cancer cells; however, to the
best of our knowledge this has not yet been tested. In the present
study, the effects of CTD treatment on NKG2D-associated immune
response in TSGH-8301 human bladder carcinoma cells were
investigated, including its effects on gene expression.
Materials and methods
Chemicals and reagents
CTD, propidium iodide, and dimethyl sulfoxide (DMSO)
were purchased from Sigma-Aldrich (Merck KGaA, Darmstadt, Germany).
All organic solvents used were of high performance liquid
chromatography grade. RPMI-1640 medium, fetal bovine serum (FBS)
and penicillin-streptomycin were obtained from Gibco (Thermo Fisher
Scientific, Inc., Waltham, MA, USA). Tissue culture plastic wares
were obtained from TPP Techno Plastic Products AG (Trasadingen,
Switzerland). The ApoBrdU DNA Fragmentation Assay kit [terminal
deoxynucleotidyl transferase dUTP nick end labeling (TUNEL)] was
obtained from BioVision, Inc. (Milpitas, CA, USA). CTD was
dissolved in DMSO and stored at −20°C prior to use.
Human bladder cancer cell culture
TSGH-8301 human bladder carcinoma cells were
purchased from the Food Industry Research and Development Institute
(Hsinchu, Taiwan). Cells were cultured in T75 tissue culture flasks
with RPMI-1640 medium supplemented with 10% FBS and 1%
penicillin-streptomycin (100 U/ml penicillin and 100 µg/ml
streptomycin). Cell cultures were maintained in a humidified
incubator at 37°C with 5% CO2.
Cell morphology assay
Cell morphology was examined as described previously
(25). Briefly, TSGH-8301 cells
(1×105 cells/well) were trypsinized, seeded into a
12-well plate with RPMI-1640 medium and cultured for 24 h. The
culture medium was then replaced with fresh medium containing CTD
(7.5 µM) and the plates were incubated for 12, 24 and 48 h at 37°C.
TSGH-8301 cell morphology was subsequently examined using an
Olympus phase-contrast microscope (Olympus Corporation, Tokyo,
Japan).
TUNEL assay for cell apoptosis
The TUNEL assay was performed with the ApoBrdU DNA
Fragmentation Assay kit according to the manufacturer's protocol
(33). TSGH-8301 cells were seeded at
a density of 1×105 cells/well onto coverslips, cultured
with 7.5 µM CTD for 48 h and then fixed with 4% formaldehyde in PBS
for 15 min, permeabilized with 0.1% (v/v) Triton X-100 for 1 h and
washed with PBS. TUNEL staining solution was added to the
coverslips to bind the fluorescein-dUTP to the DNA break terminals.
TUNEL-positive cells were examined and images captured using a
Leica TCS SP2 confocal microscope (Leica Microsystems GmBH,
Wetzlar, Germany), using an excitation wavelength of 488–623 nm and
a detection wavelength of 488–520 nm.
Complementary (c)DNA microarray
assay
TSGH-8301 cells were seeded into a 10 cm dish at a
density of 1.5×106 cells/dish with RPMI-1640 medium and
cultured for 24 h. Cells were treated with 7.5 µM CTD for 48 h and
then collected and washed twice with PBS. Total RNA from the
untreated control and CTD-treated groups were extracted using a
Qiagen RNeasy Mini kit according to the manufacturer's protocol
(Qiagen, Inc., Valencia, CA, USA). RNA concentrations were
determined using a Qubit™ Fluorometer (Invitrogen;
Thermo Fisher Scientific, Inc.) as described previously (32). For cDNA synthesis, the oligo (dT)
Maxime RT premix kit according to the manufacturer's protocol
(Intron Biotechnology, Inc., Seongnam, Korea) was used. The samples
were hybridized to the Affymetrix GeneChip® Human Gene
1.0 ST array as described previously (Affymetrix, Inc., Santa
Clara, CA, USA) (34), then the
sample fluorescence was quantified by Asia BioInnovations
Corporation (Taipei, Taiwan). The data was analyzed using
Affymetrix® Expression Console™ software
(Soft version 1.1.2; Affymetrix, Inc.) with default robust
multiarray parameters. Following comparison of the control group
with the CTD treated group, a 2-fold change in gene expression was
used as the threshold to indicate a significant effect on
expression (7–10).
Statistical analysis
Data are representative of three assays. Differences
between control and CTD-treated groups were presented when they
were >2-fold.
Results
CTD induces morphological changes in
TSGH-8301 cells in vitro
Following treatment with 7.5 µM CTD for 12, 24 and
48 h, cells were examined and imaged using phase-contrast
microscopy at a magnification of ×200 (Fig. 1). Control cells without CTD treatment
were incubated under identical conditions. The results indicated
that CTD induced cell morphological changes, including cell size
reduction, and decreased cell number in a dose-dependent
manner.
CTD induces TSGH-8301 cell death in
vitro
Following treatment with 7.5 µM CTD for 48 h, cells
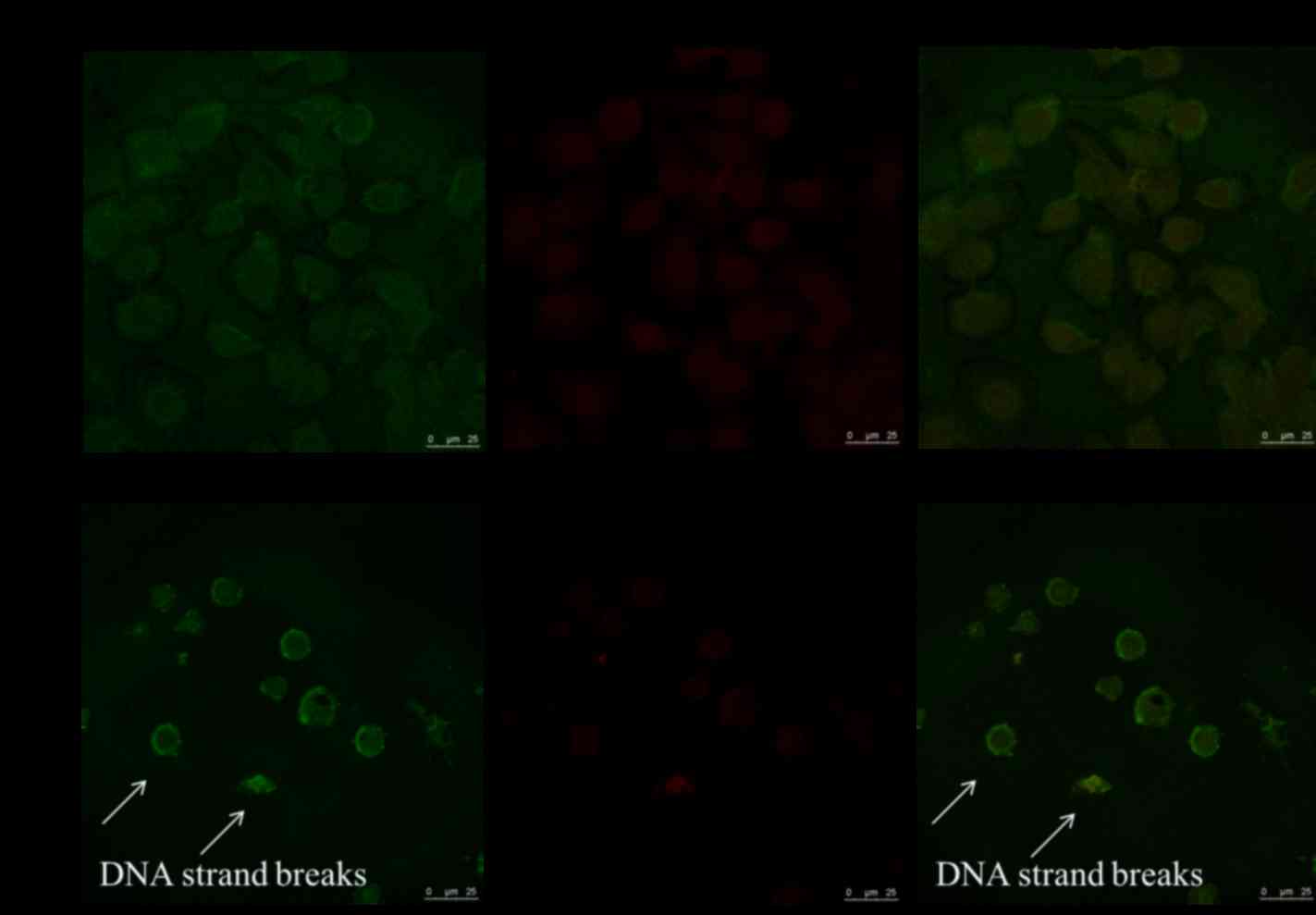
were examined using a TUNEL assay (Fig.
2). TUNEL-positive cells were observed in the CTD-treated
group, but not in the control group. The number of apoptotic nuclei
(green color) was higher in the CTD treated group compared with
that of the control group (data not shown). These observations
indicate that CTD induces apoptotic cell death in TSGH-8301
cells.
CTD alters the expression of genes
associated with the NKG2D-associated immune response in TSGH-8301
human bladder carcinoma cells
Following treatment with 7.5 µM CTD for 48 h, total
RNA was extracted from TSGH-8301 cells for cDNA microarray analysis
in order to examine changes in gene expression. The genes that were
upregulated or downregulated, calculated from the microarray data,
are displayed in in Tables I and
II. A total of 269 genes were
>2-fold upregulated and 286 genes were >2-fold downregulated
(Table I). DNA-damage-inducible
transcript 3 (DDIT3) was 4.75-fold upregulated, activating
transcription factor 3 was 5.41-fold upregulated and
dehydrogenase/reductase (SDR family) member 2 was 6.08-fold
upregulated compared with the untreated control cells.
Microfibrillar associated protein 5 was 4.79-fold downregulated,
insulin-like growth factor binding protein 5 was 5.41-fold
downregulated and matrix-remodeling associated 5 (MXRA5) was
7.98-fold downregulated compared with the untreated control cells
(Table II).
 | Table I.Number of genes that were upregulated
or downregulated in TSGH-8301 cells following treatment with 7.5 µM
CTD compared with untreated control cells. |
Table I.
Number of genes that were upregulated
or downregulated in TSGH-8301 cells following treatment with 7.5 µM
CTD compared with untreated control cells.
| A, Upregulated
genes |
|---|
|
|---|
| Fold change | Number of
genes | Total |
|---|
| ≥5 and <10 | 7 | 269 |
| ≥4 and <5 | 12 |
|
| ≥3 and <4 | 19 |
|
| ≥2 and <3 | 231 |
|
|
| B, Downregulated
genes |
|
| Fold change | Number of
genes | Total |
|
| >-3 and ≤-2 | 233 | 286 |
| >-4 and ≤-3 | 34 |
|
| >-5 and ≤-4 | 11 |
|
| >-10 and
≤-5 | 8 |
|
| Table II.Representative genes that were
upregulated or downregulated in response to CTD treatment in
TSGH-8301 cells. |
Table II.
Representative genes that were
upregulated or downregulated in response to CTD treatment in
TSGH-8301 cells.
| Probe set ID | Gene symbol | Fold change | Gene
description |
|---|
| 7998927 | TRNAP24P | 6.7 | Transfer RNA
proline 24 (anticodon AGG) pseudogene |
| 7903530 | FNDC7 | 6.59 | Fibronectin type
III domain containing 7 |
| 8114572 | HBEGF | 6.15 | Heparin-binding
EGF-like growth factor |
| 7973433 | DHRS2 | 6.08 |
Dehydrogenase/reductase (SDR family)
member 2 |
| 7909610 | ATF3 | 5.41 | Activating
transcription factor 3 |
| 8092578 | ETV5 | 5.26 | Ets variant 5 |
| 8122724 | ULBP1 | 5.11 | UL16 binding
protein 1 |
| 7982868 | CHAC1 | 4.89 | ChaC, cation
transport regulator homolog 1 (E. coli) |
| 8160912 | C9orf131 | 4.82 | Chromosome 9 open
reading frame 131 |
| 7964460 | DDIT3 | 4.75 |
DNA-damage-inducible transcript 3 |
| 7963534 | KRT4 | −4.76 | Keratin 4 |
| 7960919 | MFAP5 | −4.79 | Microfibrillar
associated protein 5 |
| 8040430 | VSNL1 | −5.25 | Visinin-like 1 |
| 8015337 | KRT15 | −5.28 | Keratin 15 |
| 8115623 | ATP10B | −5.36 | ATPase, class V,
type 10B |
| 8058857 | IGFBP5 | −5.41 | insulin-like growth
factor binding protein 5 |
| 8136336 | AKR1B10 | −5.48 | Aldo-keto reductase
family 1, member B10 (aldose reductase) |
| 8104758 | NPR3 | −5.78 | Natriuretic peptide
receptor C/guanylate cyclase C (atrionatriuretic peptide receptor
C) |
| 8113709 | LOX | −6.35 | Lysyl oxidase |
| 8171172 | MXRA5 | −7.98 | Matrix-remodelling
associated 5 |
CTD-treatment alters signaling
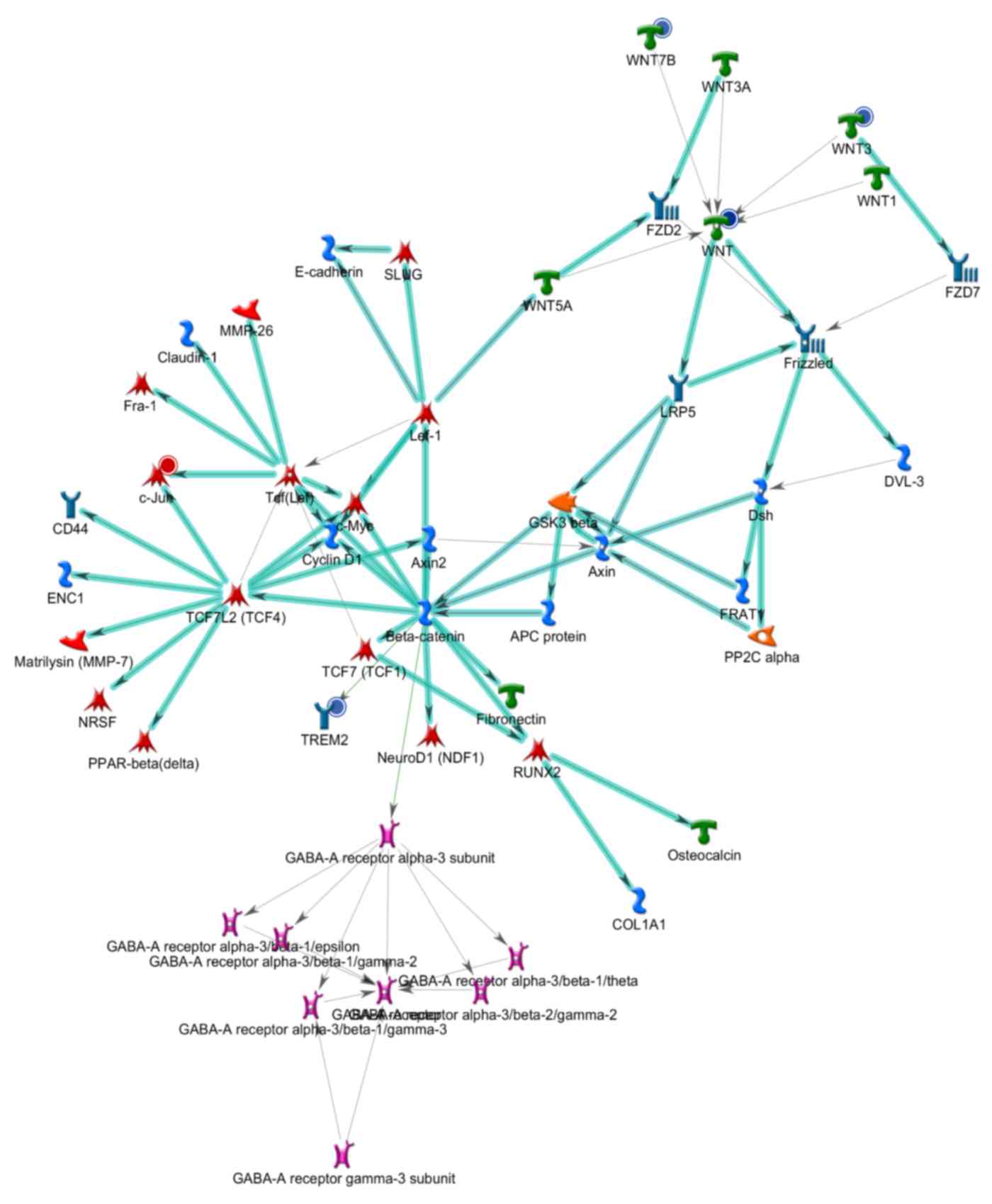
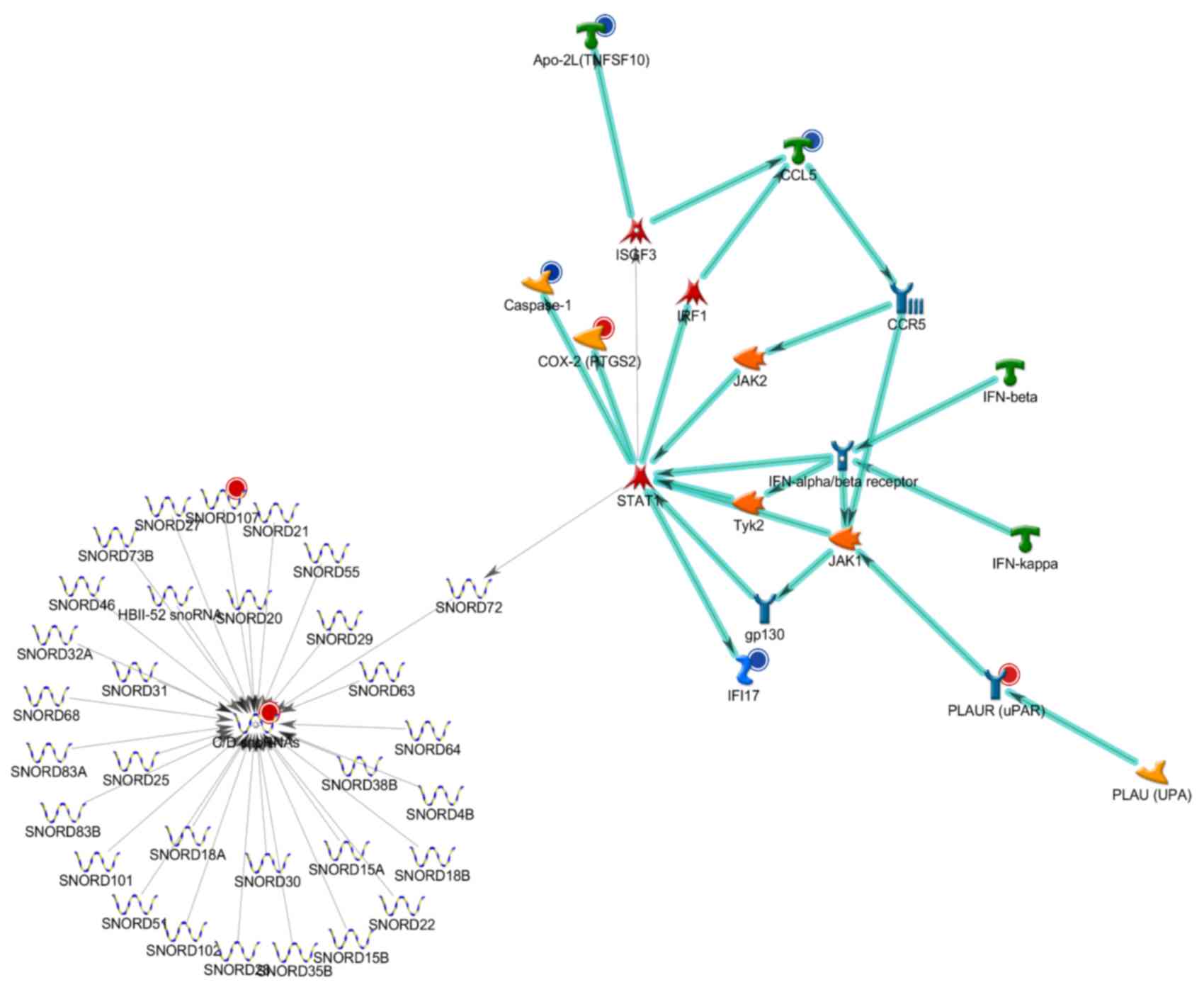
pathways in TSGH-8301 cells
In order to investigate the molecular signaling
pathways associated with the genes whose expression was altered by
CTD, a GeneGo analysis was performed. The analysis from GeneGo for
immune responses for human NKG2D type II integral membrane protein
signaling were the top, second and third highest scored Analyze
Network (AN) networks (data not shown), by the number of pathways
(Figs. 3–5). Therefore, NKG2D was selected as there
were significant effects following CTD treatment compared with that
of control.
Discussion
The results from the present study demonstrated that
CTD induces cell morphological changes and apoptosis, as revealed
using phase-contrast microscopy and a TUNEL assay, respectively, in
TSGH-8301 human bladder carcinoma cells. Tumor cell lines are
invaluable research tools that are able to be experimentally
manipulated (35). The TSGH-8301
cells are derived from a well-differentiated human transitional
cell carcinoma of the urinary bladder (36). In addition, CTD treatment resulted in
the upregulation and downregulation of a number of immune
response-associated genes. Numerous studies have demonstrated that
CTD induces a cytotoxic effect in a number of human cancer cell
lines. For example, CTD affects gene expression in human lung
cancer NCI-H460 cells (32)
Additionally, CTD induces cell apoptosis through
mitochondrial-dependent signaling pathways (25), and inhibits cell migration and
invasion through the inhibition of the matrix
metalloproteinase-2/-9 signaling pathway in human bladder cancer
TSGH-8301 cells (37). Other studies
in human bladder cancer T24 cells have also demonstrated that CTD
induces apoptosis through the calcium/PKC-regulated endoplasmic
reticulum stress signaling pathway (38). However, to the best of our knowledge
no studies have demonstrated the effect of CTD on gene expression
in human bladder cancer cells. Thus, in the present study, the
effect of CTD on gene expression in human bladder cancer TSGH-8301
cells was investigated in vitro.
In the present study, human bladder cancer TSGH-8301
cells were treated with 7.5 µM CTD, followed by analysis of cell
morphology and apoptotic cell death using phase-contrast microscopy
and the TUNEL assay. The results demonstrated that CTD induces
apoptotic cell death and morphological changes in human bladder
cancer TSGH-8301 cells. Compared with the control group, cells
treated with CTD exhibited increased DNA strand breaks, as
visualized using an ApoBrdU DNA Fragmentation Assay kit. This
result indicates that CTD induces apoptotic cell death via DNA
fragmentation in TSGH-8301 cells.
The changes in cell morphology observed in the
present study as a result of CTD treatment were dose-dependent.
Additionally, CTD induced apoptotic cell death in a dose-dependent
manner, as indicated by TUNEL staining.
A previous study demonstrated that CTD affects gene
expression in the human lung cancer NCI-H460 cell line (32). Additionally, a recent study
identified, using next generation sequencing, that human bladder
cancer has numerous gene alterations compared with normal tissue,
with most predicted to be loss-of-function mutations; however,
evaluating the functional impact of each genetic alteration is
impractical (39). The present study,
to the best of our knowledge, is the first to demonstrate the
effects of CTD on gene expression in human bladder TSGH-8301 cancer
cells. Thus, the results from the present study offer an insight
into CTD-induced gene expression changes in bladder cancer cells,
which will aid in the identification of potential biomarkers for
patients with bladder cancer that require targeted therapy.
A number of studies have shown that the matrix
proteins, stromal cells and associated secreted molecules that
comprise the tumor microenvironment serve a role in mediating
responses to cancer drugs (40–42).
Therefore, in the present study, changes in gene expression in
TSGH-8301 cells following exposure to CTD for 24 h were analyzed
using a cDNA microarray. CTD treatment resulted in an >2-fold
upregulation of 269 genes and an >2-fold downregulation of 286
genes in TSGH-8301 cells. DDIT3, which is associated with DNA
damage, was upregulated 4.75-fold and MXRA5, which is associated
with cell migration and invasion, was downregulated 7.98-fold. In
order to further investigate the genes whose expression was altered
in response to CTD, and their associated molecular signaling
pathways, a GeneGo process network analysis was performed. This
suggested that CTD treatment affects a number of associated
signaling pathways, including the NKG2D signaling pathway, in
TSGH-8301 cells.
In conclusion, the data from the present study
indicates that CTD induces cytotoxic effects in TSGH-8301 cells,
based on the observed decrease in the number of cells and increase
in apoptotic cell death in CTD-treated cells compared with control
untreated cells. Additionally, CTD treatment resulted in the
upregulation or downregulation of numerous genes that are
associated with a number of immune response-associated molecular
signaling pathways. These findings may aid in future research into
the molecular targets of CTD.
Acknowledgements
The present study was supported by the China Medical
University (Taichung, Taiwan; grant no. CMU103-ASIA-01).
Experiments and data analysis were performed in part at the Medical
Research Core Facilities Center of the Office of Research &
Development at China Medical University.
References
|
1
|
Knowles MA: The genetics of transitional
cell carcinoma: Progress and potential clinical application. BJU
Int. 84:412–427. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Pudasaini S, Subedi N, Prasad KB, Rauniyar
SK, Joshi BR and Bhomi KK: Cystoscopic bladder biopsies: A
histopathological study. Nepal Med Coll J. 16:9–12. 2014.PubMed/NCBI
|
|
3
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Parkin DM: The global burden of urinary
bladder cancer. Scand J Urol Nephrol Suppl. 12–20. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Boffetta P: Tobacco smoking and risk of
bladder cancer. Scand J Urol Nephrol Suppl. 45–54. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Prasad SM, Decastro GJ and Steinberg GD:
Medscape: Urothelial carcinoma of the bladder: Definition,
treatment and future efforts. Nat Rev Urol. 8:631–642. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ministry of Health and Welfare (Taiwan), .
Taiwan Statistics of causes of death 2006. Ministry of Health and
Welfare (Taiwan); Taipei City: 2006
|
|
9
|
Moed L, Shwayder TA and Chang MW:
Cantharidin revisited: A blistering defense of an ancient medicine.
Arch Dermatol. 137:1357–1360. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang GS: Medical uses of mylabris in
ancient China and recent studies. J Ethnopharmacol. 26:147–162.
1989. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Karras DJ, Farrell SE, Harrigan RA,
Henretig FM and Gealt L: Poisoning from ‘Spanish fly’
(cantharidin). Am J Emerg Med. 14:478–483. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Swingle M, Ni L and Honkanen RE:
Small-molecule inhibitors of ser/thr protein phosphatases:
Specificity, use and common forms of abuse. Methods Mol Biol.
365:23–38. 2007.PubMed/NCBI
|
|
13
|
Li W, Xie L, Chen Z, Zhu Y, Sun Y, Miao Y,
Xu Z and Han X: Cantharidin, a potent and selective PP2A inhibitor,
induces an oxidative stress-independent growth inhibition of
pancreatic cancer cells through G2/M cell-cycle arrest and
apoptosis. Cancer Sci. 101:1226–1233. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li W, Chen Z, Gong FR, Zong Y, Chen K, Li
DM, Yin H, Duan WM, Miao Y, Tao M, et al: Growth of the pancreatic
cancer cell line PANC-1 is inhibited by protein phosphatase 2A
inhibitors through overactivation of the c-Jun N-terminal kinase
pathway. Eur J Cancer. 47:2654–2664. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li W, Chen Z, Zong Y, Gong F, Zhu Y, Zhu
Y, Lv J, Zhang J, Xie L, Sun Y, et al: PP2A inhibitors induce
apoptosis in pancreatic cancer cell line PANC-1 through persistent
phosphorylation of IKKα and sustained activation of the NF-κB
pathway. Cancer Lett. 304:117–127. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Huh JE, Kang KS, Chae C, Kim HM, Ahn KS
and Kim SH: Roles of p38 and JNK mitogen-activated protein kinase
pathways during cantharidin-induced apoptosis in U937 cells.
Biochem Pharmacol. 67:1811–1818. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chang C, Zhu YQ, Mei JJ, Liu SQ and Luo J:
Involvement of mitochondrial pathway in NCTD-induced cytotoxicity
in human hepG2 cells. J Exp Clin Cancer Res. 29:1452010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Huang WW, Ko SW, Tsai HY, Chung JG, Chiang
JH, Chen KT, Chen YC, Chen HY, Chen YF and Yang JS: Cantharidin
induces G2/M phase arrest and apoptosis in human colorectal cancer
colo 205 cells through inhibition of CDK1 activity and
caspase-dependent signaling pathways. Int J Oncol. 38:1067–1073.
2011.PubMed/NCBI
|
|
19
|
Zhang WD, Zhao HR, Yan Y, Wang XH, Zong ZH
and Liu Y: Apoptosis induced by cantharidin in human pulmonary
carcinoma cells A549 and its molecular mechanisms. Zhonghua Zhong
Liu Za Zhi. 27:330–334. 2005.(In Chinese). PubMed/NCBI
|
|
20
|
Huan SK, Lee HH, Liu DZ, Wu CC and Wang
CC: Cantharidin-induced cytotoxicity and cyclooxygenase 2
expression in human bladder carcinoma cell line. Toxicology.
223:136–143. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li W, Li DM, Chen K, Chen Z, Zong Y, Yin
H, Xu ZK, Zhu Y, Gong FR and Tao M: Development of a gene therapy
strategy to target hepatocellular carcinoma based inhibition of
protein phosphatase 2A using the α-fetoprotein promoter enhancer
and pgk promoter: An in vitro and in vivo study. BMC Cancer.
12:5472012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Xie X, Wu MY, Shou LM, Chen LP, Gong FR,
Chen K, Li DM, Duan WM, Xie YF, Mao YX, et al: Tamoxifen enhances
the anticancer effect of cantharidin and norcantharidin in
pancreatic cancer cell lines through inhibition of the protein
kinase C signaling pathway. Oncol Lett. 9:837–844. 2015.PubMed/NCBI
|
|
23
|
Hsiao YP, Tsai CH, Wu PP, Hsu SC, Liu HC,
Huang YP, Yang JH and Chung JG: Cantharidin induces G2/M phase
arrest by inhibition of Cdc25c and Cyclin A and triggers apoptosis
through reactive oxygen species and the mitochondriadependent
pathways of A375.S2 human melanoma cells. Int J Oncol.
45:2393–2402. 2014.PubMed/NCBI
|
|
24
|
Hsia TC, Yu CC, Hsu SC, Tang NY, Lu HF,
Huang YP, Wu SH, Lin JG and Chung JG: Cantharidin induces apoptosis
of H460 human lung cancer cells through mitochondria-dependent
pathways. Int J Oncol. 45:245–254. 2014.PubMed/NCBI
|
|
25
|
Kuo JH, Chu YL, Yang JS, Lin JP, Lai KC,
Kuo HM, Hsia TC and Chung JG: Cantharidin induces apoptosis in
human bladder cancer TSGH 8301 cells through mitochondria-dependent
signal pathways. Int J Oncol. 37:1243–1250. 2010.PubMed/NCBI
|
|
26
|
Evans HJ and Prosser J: Tumor-suppressor
genes: Cardinal factors in inherited predisposition to human
cancers. Environ Health Perspect. 98:25–37. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tang C, Guo J, Chen H, Yao CJ, Zhuang DX,
Wang Y, Tang WJ, Ren G, Yao Y, Wu JS, et al: Gene mutation
profiling of primary glioblastoma through multiple tumor biopsy
guided by 1H-magnetic resonance spectroscopy. Int J Clin Exp
Pathol. 8:5327–5335. 2015.PubMed/NCBI
|
|
28
|
Heneghan HM, Miller N and Kerin MJ: MiRNAs
as biomarkers and therapeutic targets in cancer. Curr Opin
Pharmacol. 10:543–550. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Osborne C, Wilson P and Tripathy D:
Oncogenes and tumor suppressor genes in breast cancer: Potential
diagnostic and therapeutic applications. Oncologist. 9:361–377.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ai X, Jia ZM, Wang J, Di GP, Zhang XU, Sun
F, Zang T and Liao X: Bioinformatics analysis of the target gene of
fibroblast growth factor receptor 3 in bladder cancer and
associated molecular mechanisms. Oncol Lett. 10:543–549.
2015.PubMed/NCBI
|
|
31
|
Spiegelberg C, Giedl J, Gaisa NT, Rogler
A, Riener MO, Filbeck T, Burger M, Ruemmele P, Hartmann A and
Stoehr R: Frequency of activating mutations in FGFR2 exon 7 in
bladder tumors from patients with early-onset and regular-onset
disease. Int J Clin Exp Pathol. 7:1708–1713. 2014.PubMed/NCBI
|
|
32
|
Hsia TC, Yu CC, Hsu SC, Tang NY, Lu HF, Yu
CS, Wu SH, Lin JG and Chung JG: cDNA microarray analysis of the
effect of cantharidin on DNA damage, cell cycle and
apoptosis-associated gene expression in NCI-H460 human lung cancer
cells in vitro. Mol Med Rep. 12:1030–1042. 2015.PubMed/NCBI
|
|
33
|
Lu CC, Yang JS, Chiang JH, Hour MJ, Lin
KL, Lee TH and Chung JG: Cell death caused by quinazolinone HMJ-38
challenge in oral carcinoma CAL 27 cells: Dissections of
endoplasmic reticulum stress, mitochondrial dysfunction and tumor
xenografts. Biochim Biophys Acta. 1840:2310–2320. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chou YC, Chang MY, Wang MJ, Liu HC, Chang
SJ, Harnod T, Hung CH, Lee HT, Shen CC and Chung JG: Phenethyl
isothiocyanate alters the gene expression and the levels of protein
associated with cell cycle regulation in human glioblastoma GBM
8401 cells. Environ Toxicol. 32:176–187. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Barretina J, Caponigro G, Stransky N,
Venkatesan K, Margolin AA, Kim S, Wilson CJ, Lehár J, Kryukov GV,
Sonkin D, et al: The Cancer Cell Line Encyclopedia enables
predictive modelling of anticancer drug sensitivity. Nature.
483:603–607. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yeh MY, Yu DS, Chen SC, Lin MS, Chang SY,
Ma CP and Han SH: Establishment and characterization of a human
urinary bladder carcinoma cell line (TSGH-8301). J Surg Oncol.
37:177–184. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Huang YP, Ni CH, Lu CC, Chiang JH, Yang
JS, Ko YC, Lin JP, Kuo JH, Chang SJ and Chung JG: Suppressions of
Migration and Invasion by Cantharidin in TSGH-8301 human bladder
carcinoma cells through the inhibitions of matrix
metalloproteinase-2/−9 signaling. Evid Based Complement Alternat
Med. 2013:1902812013.PubMed/NCBI
|
|
38
|
Su CC, Liu SH, Lee KI, Huang KT, Lu TH,
Fang KM, Wu CC, Yen CC, Lai CH, Su YC and Huang CF: Cantharidin
induces apoptosis through the calcium/PKC-regulated endoplasmic
reticulum stress pathway in human bladder cancer cells. Am J Chin
Med. 43:581–600. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hensel J, Duex JE, Owens C, Dancik GM,
Edwards MG, Frierson HF and Theodorescu D: Patient mutation
directed shRNA screen uncovers novel bladder tumor growth
suppressors. Mol Cancer Res. 13:1306–1315. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ayala F, Dewar R, Kieran M and Kalluri R:
Contribution of bone microenvironment to leukemogenesis and
leukemia progression. Leukemia. 23:2233–2241. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Straussman R, Morikawa T, Shee K,
Barzily-Rokni M, Qian ZR, Du J, Davis A, Mongare MM, Gould J,
Frederick DT, et al: Tumour micro-environment elicits innate
resistance to RAF inhibitors through HGF secretion. Nature.
487:500–504. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wilson TR, Fridlyand J, Yan Y, Penuel E,
Burton L, Chan E, Peng J, Lin E, Wang Y, Sosman J, et al:
Widespread potential for growth-factor-driven resistance to
anticancer kinase inhibitors. Nature. 487:505–509. 2012. View Article : Google Scholar : PubMed/NCBI
|