Introduction
Gastric cancer is one of the most common
malignancies worldwide, with an estimated 951,600 novel stomach
cancer cases and 723,100 mortalities occurring in 2012 (1). Treatment options currently available for
gastric cancer are inadequate. Recent advances in surgical
techniques have led to an increase in the 5-year survival rate of
patients with gastric cancer from 10 to 30%. However, patients with
gastric cancer in advanced stages are untreatable (2–5).
Therefore, there is an urgent requirement to identify novel
therapeutic agents that can decrease the mortality of cancer
patients with fewer side effects.
Natural products have been identified to exhibit
anticancer effects by targeting multiple cellular signaling
pathways, mediated by complex signal transduction (5). In our previous studies, extracts of the
herbal plants Citrus aurantium L., Lonicera japonica
Thunb. and Scutellaria baicalensis Georgi were identified to
exhibit anticancer effects in human gastric, liver and lung cancer
cells, respectively (6–8).
Flavonoids are natural polyphenolic compounds
extensively present in vegetables and fruits. The use of flavonoids
as anticancer compounds has been investigated previously (6). In previous a in vitro study,
S. baicalensis Georgi extract was reported to be cytotoxic
to a broad spectrum of human cancer cell lines (7). In spite of the molecular mechanisms
underlying these effects remaining unclear, previous studies
suggest that the cytotoxicity of S. baicalensis Georgi
regulates the viability of human cancer cells by inducing
apoptosis, arresting cell cycle progression and regulating
metastasis in various cancer cell lines. S. baicalensis
Georgi has been widely used in traditional Chinese herbal medicine
to treat inflammation, hypertension and cardiovascular disease, and
has officially been listed in the Chinese Pharmacopoeia as a
medicinal plant (8–11).
Apoptosis, or type 1 programmed cell death, serves a
fundamental role in the normal development and differentiation of
multicellular organisms. Activation of apoptosis in cancer cells
may be instrumental in the process of cancer cell elimination.
There are two primary apoptotic signaling pathways: The extrinsic
or death receptor signaling pathway, and the intrinsic or
mitochondrial signaling pathway (12). Apoptosis is characterized by
cytoplasmic shrinking, extensive plasma membrane blebbing and
nuclear condensation formation (13,14).
Apoptosis progresses in either a caspase-dependent or a
caspase-independent manner (15).
Furthermore, the expression ratio of anti- and pro-apoptotic
mitochondrial proteins, including B cell lymphoma 2 (Bcl-2)
-associated X protein (Bax)/B cell lymphoma extra-large (Bcl-xL),
is crucial for the induction of apoptosis, and determines the
susceptibility of cells to apoptosis (16).
In the present study, the anticancer effect of
flavonoid extract from Korean S. baicalensis Georgi (FSB)
was investigated with the aim of elucidating the underlying
molecular mechanisms of the anticancer effect of FSB on AGS human
gastric cancer cells.
Materials and methods
Chemicals
RPMI-1640 medium, antibiotics
(penicillin/streptomycin) and fetal bovine serum were purchased
from Gibco; Thermo Fisher Scientific, Inc. (Waltham, MA, USA).
Antibodies against caspase-3 (#9662; 1:1,000), caspase-9 (#9502;
1:1,000), cleaved caspase-3 (#9664; 1:1,000), poly (ADP-ribose)
polymerase (PARP) (#9542; 1:1,000), cleaved PARP (#5625; 1:1,000),
tumor necrosis factor regulator superfamily member 6 (Fas) (#4233;
1:500), Fas ligand (FasL) (#4273; 1:500), Bcl-xL (#2764; 1:1,000)
and Bax (#2774; 1:1,000) were purchased from Cell Signaling
Technology, Inc. (Danvers, MA, USA). Anti-β-actin antibody was
obtained from EMD Millipore (#MAB-1501; 1:10,000; Billerica, MA,
USA). Anti-rabbit IgG (#ADI-SAB-300; 1:2,000) and horseradish
peroxidase-conjugated goat anti-mouse IgG (#ADI-SAB-100-J; 1:2,000)
were purchased from Enzo Life Sciences (Postfach CH-4415 Lausen,
Switzerland). MTT was obtained from Sigma-Aldrich; Merck KGaA
(Darmstadt, Germany). An Annexin V-fluorescein isothiocyanate
(FITC) apoptosis detection kit (FITC Annexin V Apoptosis Detection
kit I) was purchased from BD Pharmingen (San Diego, CA, U.S.A.).
Propidium iodide (PI) was procured from Sigma-Aldrich (Merck KGaA).
A caspase-8 activity assay kit (Caspase 8 Colorimetric Activity
Assay kit, IETD) was purchased from EMD Millipore (Billerica, MA,
USA), Hoechst 33342 trihydrochloride stain was purchased from
Invitrogen; Thermo Fisher Scientific (Eugene, OR97402).
Electrophoretic materials and chemicals were procured from Bio-Rad
Laboratories, Inc. (Hercules, CA, USA).
Isolation of flavonoids from Korean S.
baicalensis Georgi
Korean S. baicalensis Georgi radix was
obtained from the Animal Bioresources Bank (Jinju, Korea). The
flavonoids were isolated using HPLC-tandem mass spectrometry
(MS/MS) at the Department of Chemistry, Gyeongsang National
University (Jinju, Korea) by Professor Sung Chul Shin. The sample
was prepared according to a previously described method (16). Samples were stored at −20°C until
use.
Cell culture
Human gastric cancer AGS cells were obtained from
the Korea Cell Line Bank (Seoul, Korea) and cultured and maintained
in RPMI-1640 medium supplemented with 10% (v/v) heat-inactivated
fetal bovine serum and 1% penicillin/streptomycin in a humidified
atmosphere with 5% CO2 at 37°C.
Cell viability assay
To assess the effect of FSB on AGS cell viability,
cells were seeded (1×105 cells/ml) on a 12-well plate
and were treated with 50, 100, 200 and 400 µg/ml FSB or vehicle
(DMSO) alone for 24 h at 37°C. A 100 µl volume of MTT solution (0.5
mg/ml) was subsequently added to each well prior to incubation at
37°C for 3 h. The culture medium was removed and 500 µl
dimethylsulfoxide (DMSO) was added to each well to dissolve the
formazan crystals that had formed. Following mixing, absorbance was
determined at 540 nm using a plate reader (Bio-Rad Laboratories,
Inc.). Cell viability was expressed as a percentage of viability
relative to that of the vehicle-treated control, which was set as
100%.
Cell cycle distribution and
measurement of cell apoptosis
AGS cells were seeded in 60-mm dishes
(1×105 cells/ml) and incubated at 37°C. When cells had
reached between 70 and 80% confluence, they were treated with 0, 50
and 100 µg/ml FSB added directly to the culture medium and
incubated at 37°C for 24 h. Following incubation, whole cells were
harvested by trypsinization and washed with ice-cold PBS and fixed
in 70% ethanol at −20°C for 1 h. Fixed cells were washed in PBS and
stained with PI (50 µg/ml) including RNase A (0.1 mg/ml) in PBS for
30 min in the dark. Flow cytometry analyses were performed using a
Cytomics FC 500 flow cytometer (Beckman Coulter, Inc., Brea, CA,
USA). In each sample, 10,000 cells were analyzed. Data were
analyzed using CXP software (version 2.2; Beckman Coulter,
Inc.).
Annexin V-PI apoptosis detection
assay
AGS cells were treated with various concentrations
of FSB (0, 50 and 100 µg/ml) for 24 h at 37°C. Cells were harvested
by trypsinization and washed with PBS, then resuspended in binding
buffer [0.1 M Hepes/NaOH (pH-7.4), 1.4 M NaCl, 25 mM
CaCl2]. Cells were stained with Annexin V-FITC and PI
for 15 min at room temperature in the dark, and 500 µl binding
buffer was added. The stained apoptotic cells were measured using a
FACSCalibur flow cytometer (BD Biosciences, Franklin Lakes, NJ,
USA). In each sample, 10,000 cells were analyzed. The data were
analyzed using Cell Quest software (version 7.5.3; BD
Biosciences).
Hoechst 33,258 staining
Cell nuclear morphology was evaluated using
fluorescence microscopy following Hoechst 33,258 DNA staining. AGS
cells were seeded (1×105 cells/ml) on coverslips and
treated with treated or not with 100 µg/ml FSB for 24 h at 37°C.
The cells were then washed in ice-cold PBS and fixed in 3.7%
paraformaldehyde solution for 15 min at room temperature. The cells
were stained with Hoechst 33258 (5 µg/ml in PBS) for 10 min at room
temperature and structural changes were observed using a
fluorescence microscope (Leica Microsystems, Ltd., Milton Keynes,
UK).
Measurement of mitochondrial membrane
potential
AGS cells were incubated with or without FSB at
various concentrations (0, 50 and 100 µg/ml) at 37°C for 24 h in
RPMI-1640 medium. After 24 h of incubation, cells were harvested by
trypsinization and washed with 1X PBS. Washed cells were stained
with 3,3′-dihexyloxacarbocyanine iodide (DiOC6) dye and the change
in fluorescent intensity was analyzed using a Cytomics FC 500 flow
cytometer. In each sample, 10,000 cells were analyzed. The data
were analyzed using CXP software.
Caspase-8 activity assay
Caspase-8 activity was determined using a
colorimetric assay with a caspase-8 activity kit, according to the
manufacturer's protocol. AGS cells were incubated with or without
FSB at various concentrations (0, 50 and 100 µg/ml) at 37°C for 24
h in complete medium. After 24 h of incubation, cells were
harvested by trypsinization and lysed using lysis buffer supplied
in the kit and incubated for 10 min in an ice bath. Subsequently,
samples were centrifuged at 10,000 × g for 5 min in a
microcentrifuge. The supernatants were collected and incubated at
37°C for 1 h with assay buffer and caspase-8 substrate
(N-acetyl-Ile-Glu-Thr-Asp-p-nitroanilide) supplied in the kit. The
optical density of the reaction mixture was quantified using a
plate reader at 405 nm.
Determination of proteins involved in
the apoptotic signaling pathway using western blot analysis
AGS cells were treated with various concentrations
of FSB (0, 50 and 100 µg/ml) at 37°C for 24 h and cell lysates were
prepared using lysis buffer [1% (w/w) NP40, 1% (w/v) sodium
deoxycholate, 0.1% (w/v) SDS, 0.15 M NaCl, 0.01 M sodium phosphate
buffer, pH 7.2, 2 mM EDTA and protease inhibitors (Thermo Fisher
Scientific, Inc.)]. Protein lysates were centrifuged at 14,500 × g
for 15 min at 4°C to remove insoluble material. The protein
concentration in the supernatants was determined using a Bradford
protein assay kit (Bio-Rad Laboratories, Inc.), according to the
manufacturer's protocol. A total of 50 µg of proteins were
separated by SDS-PAGE (12% gel) and transferred onto a
polyvinyldene fluoride membrane using a TE 77 Semi-Dry Transfer
Unit (GE Healthcare Life Sciences, Chalfont, UK). Following
blocking in 5% skimmed milk powder in Tris-buffered saline
containing Tween-20 for 1 h at room temperature, membranes were
incubated overnight with primary antibodies at 4°C followed by
incubation for 3 h with horseradish peroxidase-conjugated secondary
antibody at room temperature. Blots were developed using an
enhanced chemiluminescence detection system (GE Healthcare Life
Sciences). Bands were quantitatively analyzed using the ImageJ
1.50i program (rsb.info.nih.gov), normalized to β-actin expression.
Experiments were performed three times.
Statistical analysis
All statistical analyses were performed using SPSS
software for Windows (version 10.0; SPSS, Inc., Chicago, IL, USA).
Results are presented as the mean ± standard deviation of at least
three independent experiments. The statistical significance between
the control and test groups was determined using one-way analysis
of variance followed by Student's t-test. P<0.05 was considered
to indicate a statistically significant difference.
Results
Characterization and quantification of
FSB
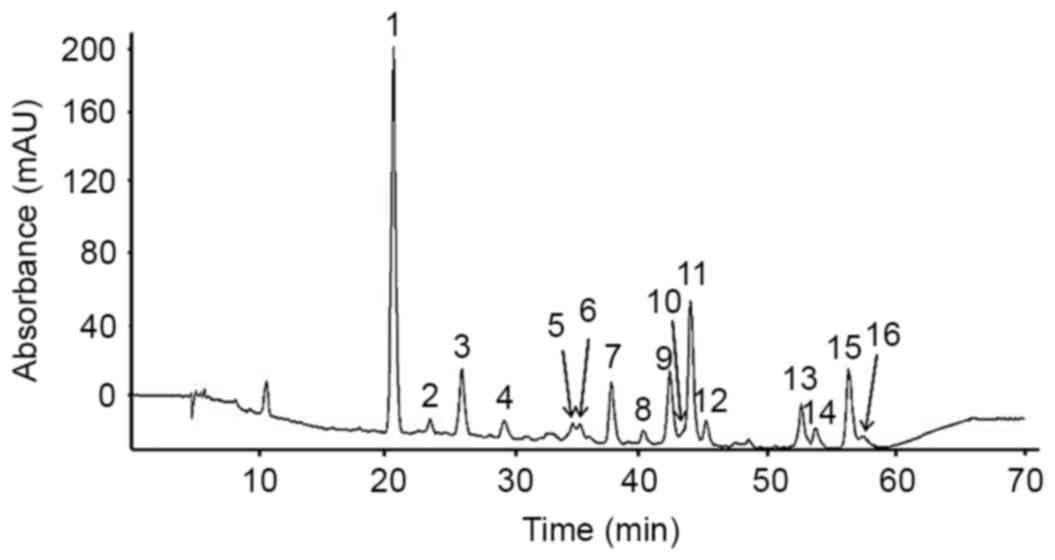
Flavonoids were isolated from Korean S.
baicalensis Georgi using HPLC-MS/MS. A total of 16 peaks at 280
nm were identified on the basis of the HPLC retention time,
molecular ion masses and the ultraviolet-visible spectra of a
library of standard compounds (Fig.
1). The mass spectral and quantification data of the 16
flavonoids are presented in Table
I.
 | Table I.List of identified flavonoids from
Scutellaria baicalensis Georgi and quantification data. |
Table I.
List of identified flavonoids from
Scutellaria baicalensis Georgi and quantification data.
| Peak no. | Compound | Rt, min | MS
[M-H]- | MS/MS | Quantity
(mg/kg) |
|---|
| 1 |
Pentahydroxyflavanone derivative | 20.64 | 629 | 303, 285, 275, 217,
201, 177, 149, 125 | 174.844±4.017 |
| 2 |
Pentahydroxyflavanone | 23.49 | 303 | 303, 285, 275, 259,
217, 193, 177, 149, 125, 109 | 14.833±0.167 |
| 3 | Viscidulin
I-O-diglucoside | 25.98 | 625 | 301, 283, 273, 258,
229, 185, 151, 125 | 23.500±0.388 |
| 4 |
Pentahydroxyflavone | 29.31 | 301 | 301, 283, 269, 259,
240, 191, 179, 161, 139, 124, 121, 109 | 14.448±0.059 |
| 5 | Unidentified | 34.65 | 647(+) | 647, 501, 467, 347,
321, 303, 285 | – |
| 6 | Viscidulin
III-O-glucoside | 35.22 | 507 | 345, 330, 315 | – |
| 7 |
Tetrahydroxyflavone | 37.69 | 285 | 285, 268, 241, 217,
199, 177, 151, 133, 107 | 50.577±0.158 |
| 8 | Iridin | 40.20 | 521 | 383, 359, 344, 329,
313, 300, 285, 212 | 16.639±0.075 |
| 9 | Eriodictyol
(4′-hydroxynaringenin) | 42.25 | 289(+) | 289, 271, 247, 179,
163, 153, 147 | 23.509±0.244 |
| 10 | Puerarin | 43.24 | 415 | 415, 295, 267, 253,
223 | 6.665±0.045 |
| 11 | Viscidulin III | 43.88 | 347(+) | 347, 332, 317, 314,
289, 286, 183, 169, 150, 142 | – |
| 12 |
Pentahydroxyflavone | 45.11 | 301 | 301, 283, 269, 241,
225, 197, 179, 165, 161, 139, 133, 124, 107 | 16.108±0.541 |
| 13 | Unidentified | 52.55 | 675 | 675, 529, 481, 361,
335, 317, 285 | – |
| 14 | Baicalin | 53.67 | 447(+) | 447, 343, 271, 253,
225, 169, 149, 123, 105 | 1.998±0.004 |
| 15 | Scutellarein | 56.25 | 285 | 285, 267, 257, 239,
213, 195, 185, 167, 165, 137, 119, 117 | 25.741±0.119 |
| 16 |
Isoscutellarein | 57.35 | 285 | 285, 267, 257, 241,
239, 229, 213, 185, 167, 165, 137, 119, 117 | 7.143±0.025 |
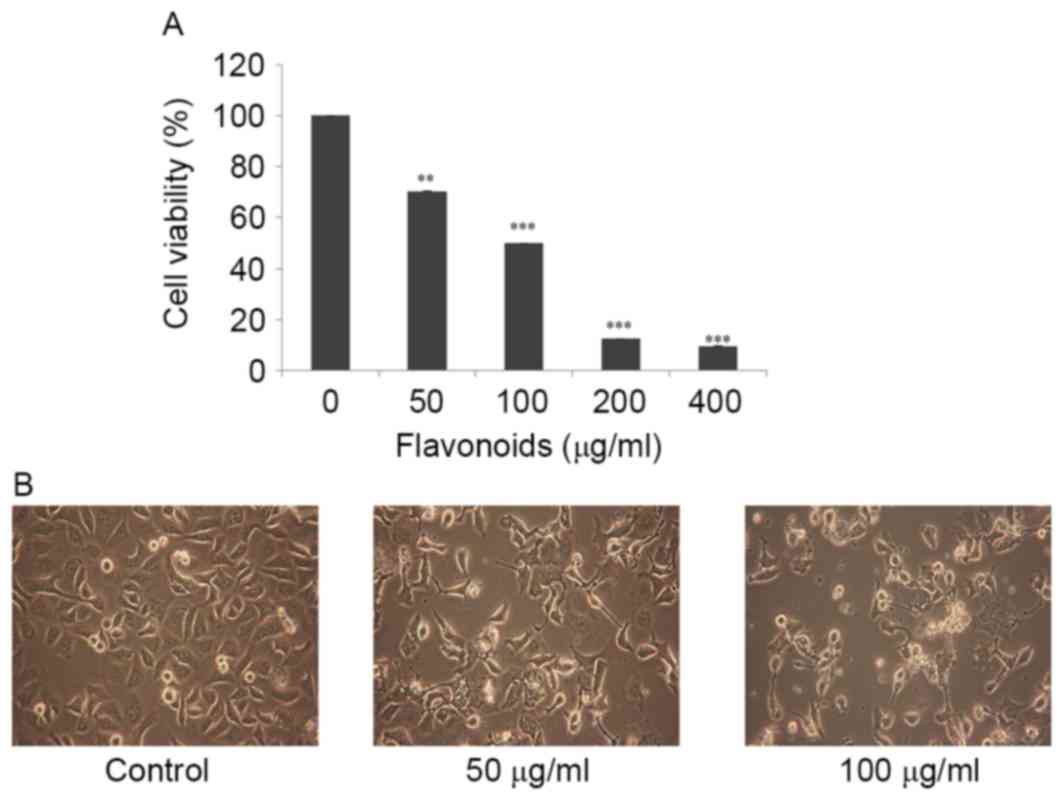
FSB inhibits the viability of AGS
cells
To determine appropriate inhibitory concentrations
of FSB on AGS cells, cells were treated with various concentrations
(0, 50, 100, 200 and 400 µg/ml) FSB for 24 h and an MTT assay was
performed to evaluate cell viability. As presented in Fig. 2A, FSB exhibited a
concentration-dependent inhibitory effect on AGS cells compared
with the control (DMSO only) at 24 h and the half-maximal
inhibitory concentrations (IC50) value was identified to be ~100
µg/ml (P<0.01 for the FSB-treated groups compared with the
control). Previously, FSB demonstrated a cytotoxic effect in AGS
human gastric cancer cells, but not in normal cells (9), therefore the cytotoxicity of FSB is
cancer cell-specific. In the present study, concentrations of FSB
of 0, 50 and 100 µg/ml were used for further experiments. Further
to the results of the MTT assay, microscopic examination revealed
morphological changes, including cell shrinkage, floating of dead
cells and a decrease in cell numbers were observed in FSB-treated
cells (Fig. 2B), as primary
indications of apoptosis.
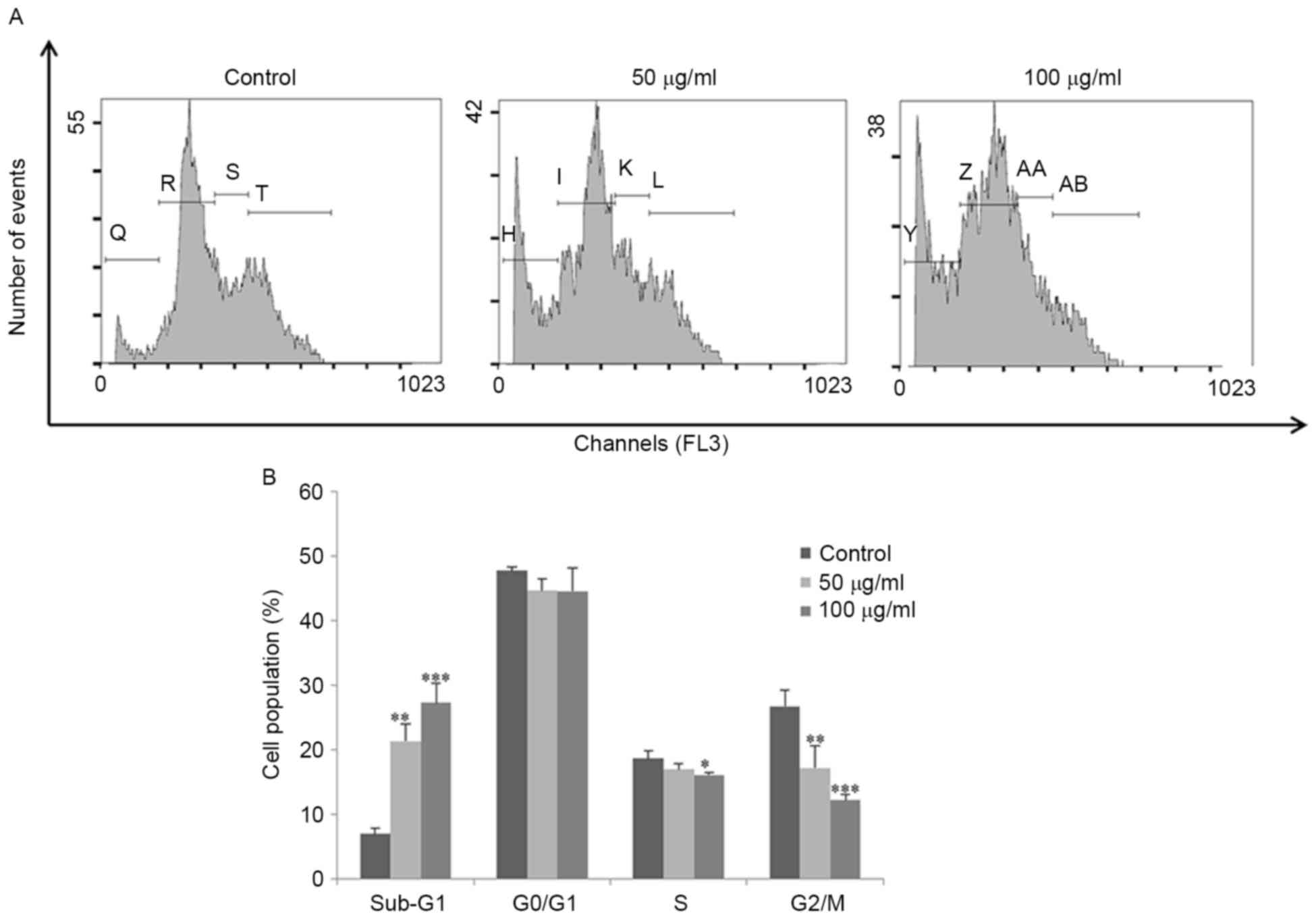
FSB induces sub-G1 phase
accumulation and apoptosis
Flow cytometric analysis was performed to determine
the cell cycle distribution and the population of cell death in
FSB-treated AGS cells. Treatment with FSB increased the proportion
of sub-G1 phase content (representing the apoptotic cell
population) from 7 to 21% (P<0.05) and 27% (P<0.001) for 50
and 100 µg/ml, respectively; FSB substantially decreased the G0/G1,
S and G2/M phase populations (Fig. 3A and
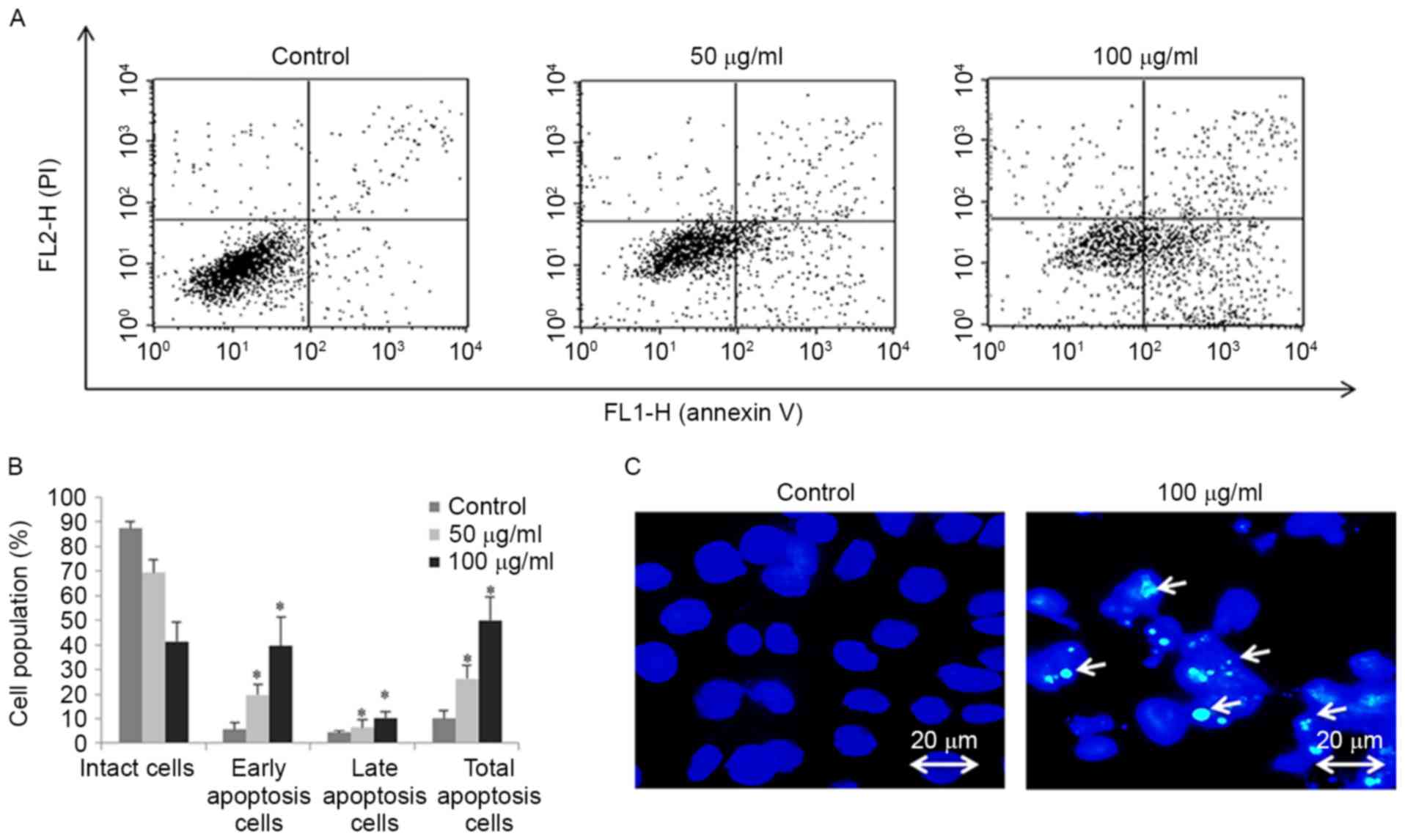
B). The effect of FSB on the induction of apoptosis in AGS
cells was also assessed using Annexin V-FITC/PI double-labeled flow
cytometry (Fig. 4A). FSB
significantly increased early apoptotic cell proportions and late
apoptotic cell proportions of AGS cells in a
concentration-dependent manner (P<0.05; Fig. 4B). Furthermore, apoptosis was
confirmed by Hoechest 33342 staining in AGS cells treated with FSB
at a concentration of 100 µg/ml, with nuclear fragmentation and
apoptotic bodies observed (Fig. 4C).
These results suggest that FSB may induce apoptotic cell death in
AGS cells.
FSB induces the mitochondrial
apoptotic signaling pathway in AGS cells
In order to assess the underlying molecular
mechanism by which FSB induced apoptotic cell death in AGS cells,
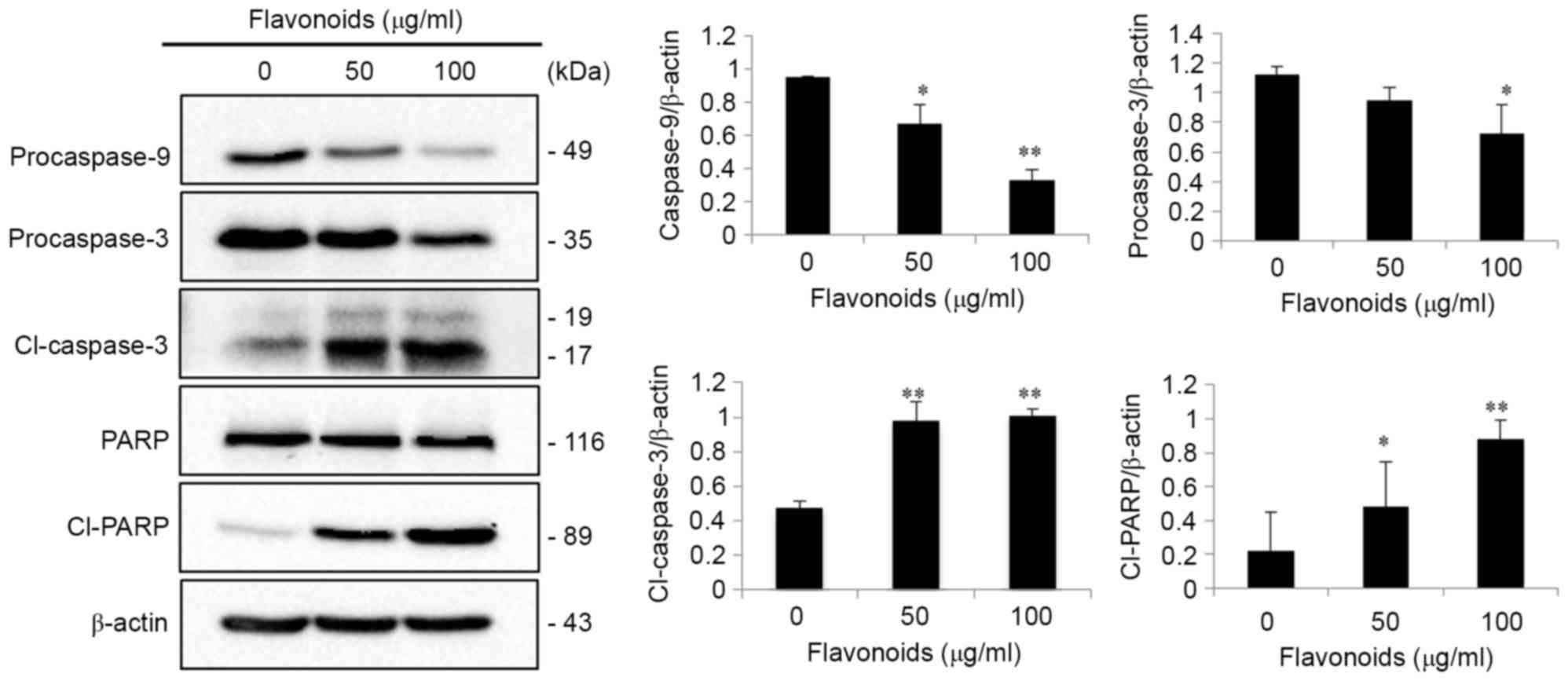
western blot analysis of the caspase cascade was performed in AGS
cells. The western blot results indicated that the expression of
procaspase-3 and procaspase-9 was significantly decreased, and,
following caspase activation, the expression of cleaved caspase-3
and cleaved PARP, a substrate of activated caspase-3, was
significantly increased in a concentration-dependent manner
(P<0.05 or <0.01; Fig. 5).
However, no significant alterations in total PARP expression in
FSB-treated cells were identified. To investigate the effect of FSB
on mitochondrial membrane potential, AGS cells untreated or treated
with FSG for 24 h were stained with DiOC6 dye and the alteration in
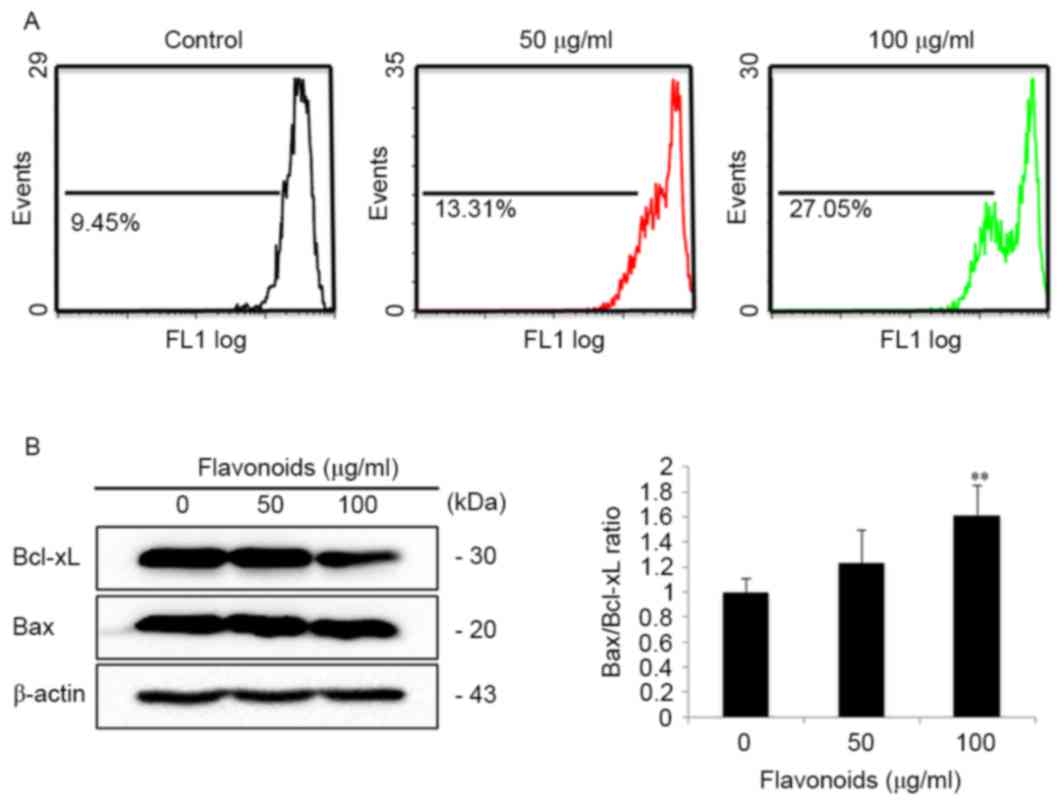
fluorescence intensity was assessed using flow cytometry. It was
observed that FSB treatment markedly decreased the mitochondrial
membrane potential of AGS cells (Fig.
6A). In addition, the expression of the
mitochondrion-associated apoptotic proteins Bax and Bcl-xL was also
analyzed in FSB-treated AGS cells. FSB increased the expression of
the Bax/Bcl-xL ratio of AGS cells in a concentration-dependent
manner (Fig. 6B). These results
suggest that FSB induced caspase-dependent mitochondrion-mediated
apoptosis in AGS cells.
FSB induces apoptosis in AGS cells are
independent of the extrinsic apoptotic signaling pathway
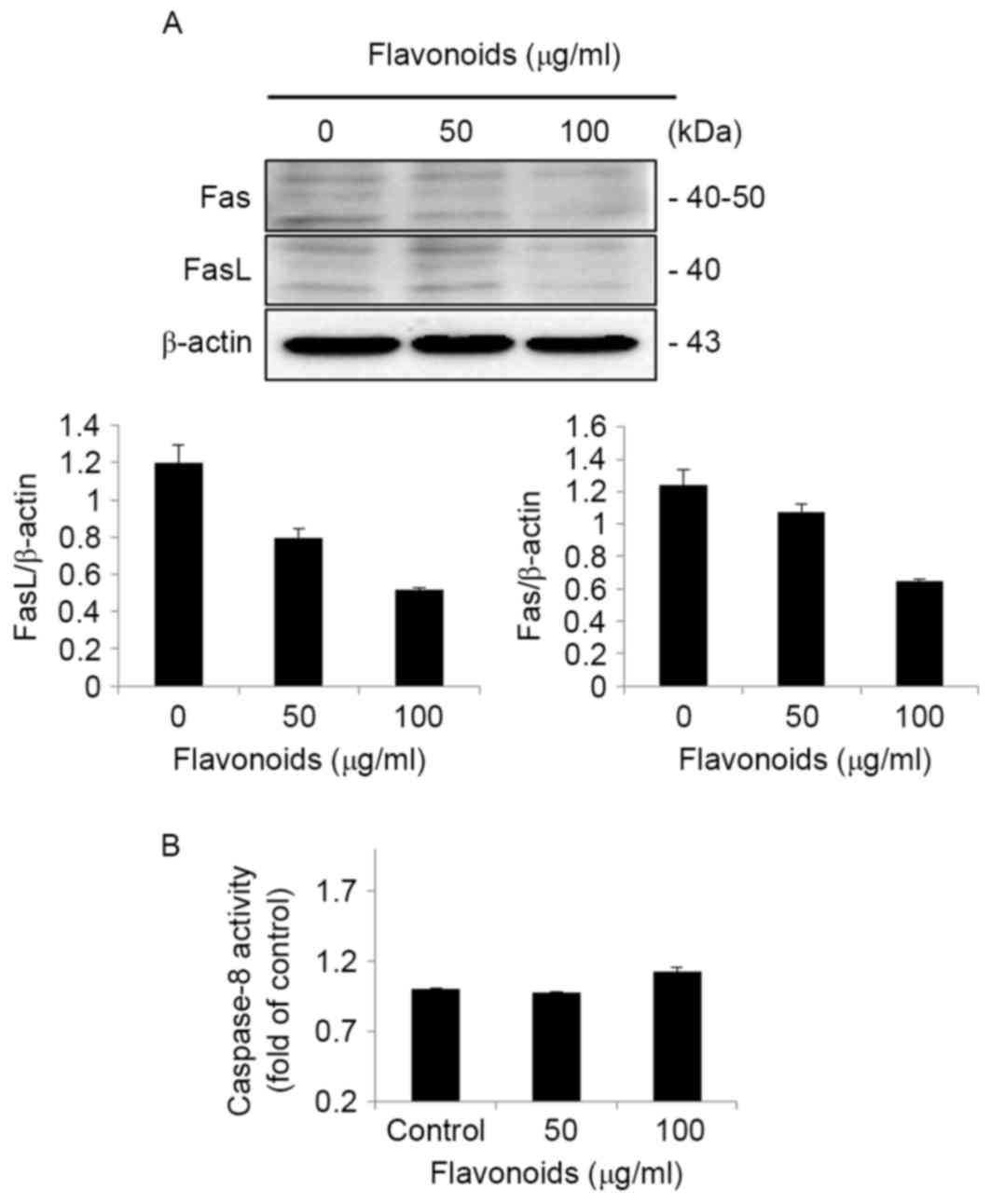
In order to determine the involvement of the
extrinsic apoptotic signaling pathway in FSB-induced apoptotic cell
death in AGS cells, western blot analysis of extrinsic apoptotic
signaling pathway proteins was performed in AGS cells. The western
blot results identified that the expression of Fas and FasL were
decreased by FSB treatment (Fig. 7A).
In addition, the caspase-8 activity assay results indicated that
there was no activation of caspase-8 (Fig. 7B), which is one of the primary
extrinsic apoptosis-associated proteins. These results indicate
that there was no involvement of the extrinsic apoptotic signaling
pathway in FSB-induced apoptotic cell death in AGS cells.
Discussion
Screening for natural compounds that are able to
induce apoptosis in cancer cells is a promising emerging strategy
for the treatment of cancer (17).
Furthermore, natural products, such as traditional herbal
medicines, which have fewer side effects compared with modern
chemotherapeutics, have long been used clinically to treat various
diseases, including cancer (18–20)
Therefore, the identification of naturally occurring agents with
pro-apoptotic activity in cancer cells is a promising approach for
developing novel cancer chemotherapies. Although it has been
demonstrated previously that the components of Korean S.
baicalensis Georgi extract have distinct pharmacological
properties, including anti-inflammatory and anticancer effects
(7,9),
the underlying molecular mechanism of action of its antitumor
property in gastric cancer remains largely unknown. To investigate
the anticancer effects of FSB and its underlying molecular
mechanism on AGS human gastric cancer cells, FSB, consisting of a
mixture of 16 flavonoids, was isolated and characterized by
HPLC-MS/MS, and used to assess its effect on the viability of AGS
cells. FSB significantly inhibited AGS cell viability in a
concentration-dependent manner and the IC50 was
determined to be ~100 µg/ml. Morphological examination identified
that a decrease in cell number and alterations in cell morphology,
including cell shrinkage, were observed in FSB-treated cells,
indicating apoptotic cell death.
Previous studies have indicated that various
anticancer and chemo preventive agents, including natural
compounds, exert anticancer effects via inducing apoptosis (type I
programmed cell death) (21–23). We previously demonstrated that cell
cycle aberrations frequently lead to apoptosis in various cell
lines (24–27) In the present study, the accumulation
of sub-G1 phase cells (indicating apoptosis) was
identified in FSB-treated AGS cells in a concentration-dependent
manner. Furthermore, Annexin V/FITC and PI staining confirmed
apoptosis. These results are consistent with those of a number of
previous studies on apoptotic cell death induced in various cancer
cell lines (26,28). Cleaved nuclei and apoptotic bodies,
which are hallmarks of apoptosis, were identified in FSB-treated
cells in the present study using Hoechest 33342 staining. These
results revealed that FSB effectively suppressed AGS cells and
induced apoptosis.
In apoptosis, caspases are key executioners of
apoptosis that serve a critical role in drug-induced apoptosis in a
variety of cancer cells. Members of this group of enzymes are known
as ‘initiator’ and ‘effector’ caspases. Upon activation, extrinsic
and intrinsic caspases, including caspase-8 and caspase-9,
respectively trigger the proteolysis activation of executioner
caspases including caspase-3 and caspase-6. In general, caspase-3
is the common effector for the majority of apoptotic signaling
pathways, and its active form is responsible for the cleavage of
PARP and breakdown of a number of cellular components associated
with DNA repair and regulation (13,15,29,30).
The results of the present study demonstrated that FSB
significantly decreased the expression of procaspase-3 and
procaspase-9, and increased the expression of cleaved caspase-3 and
cleaved PARP. PARP is a nuclear DNA-binding zinc-finger protein
which catalyzes the conversion of the dinucleotide nicotine-adenine
dinucleotide into nicotinamide and protein-linked chains of
ADP-ribose which leads to DNA fragmentation (31). The results of the present study
demonstrated that there was no involvement of proteins associated
with the extrinsic apoptotic signaling pathway, including Fas, FasL
and caspase-8, and activation of caspase-8 in FSB-treated AGS
cells.
Mitochondria serve an essential role in the
propagation of apoptosis; evidence suggests that decreasing
mitochondrial membrane potential is associated with mitochondrial
dysfunction. Therefore, a decrease in mitochondrial membrane
potential serves an important role during mitochondrially mediated
apoptosis. The Bcl-2 family proteins are apoptotic regulatory
proteins, which mediate the mitochondrial apoptotic process
(32). The pro-apoptotic and
anti-apoptotic proteins of Bcl-2 family in the cell determine
whether a cell lives or dies. Bcl-xL interacts with the
mitochondrial plasma membrane and protects it from other apoptotic
factors, including Bax and Bcl-2 homologous antagonist killer, thus
preventing the release of cytochrome c from the plasma
membrane. Previous studies have demonstrated that flavonoids induce
apoptosis in various tumor cell lines by inhibiting the
overexpression of Bcl-2 family proteins (26,33,34). The
Bax/Bcl-xL ratio appears to be a factor that determines apoptosis.
In the present study, it was observed that FSB treatment
significantly decreased the mitochondrial membrane potential of AGS
cells. In addition, Bcl-xL was significantly downregulated, whereas
Bax protein levels were unchanged; however, the Bax/Bcl-xL ratio
was upregulated in FSB-treated AGS cells. Cytochrome c is
able to bind to apoptotic protease-activating factor 1 when it is
released from the mitochondria into the cytosol by increasing the
Bax/Bcl-xL ratio, leading to the activation of caspase-3 and
ultimately resulting in apoptosis. The results of the present study
suggest that FSB-induced apoptosis in AGS human gastric cancer
cells is via the mitochondrially mediated caspase-dependent
intrinsic apoptotic signaling pathway.
The results of the present study indicate that FSB
significantly inhibits cell viability and induces apoptosis in AGS
cells via the mitochondrially mediated intrinsic apoptotic
signaling pathway. FSB-induced apoptosis was identified to be
mediated by caspase activation and triggered by the modulation of
Bcl-2 family proteins. To the best of our knowledge, the present
study is the first to elucidate the underlying molecular mechanism
for the anticancer activity of FSB in human gastric cancer AGS
cells. Therefore, the present study provides novel insights into
the biological effects of FSB, which may possess therapeutic
potential for the treatment of human gastric cancer.
Acknowledgements
The present study was supported by a grant from the
National Research Foundation of Korea (NRF) funded by the Ministry
of Science, ICT & Future Planning (grant nos. 2012M3A9B8019303
and 2017R1A2B4003974).
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Green D, de Leon S Ponce, Leon-Rodriguez E
and Sosa-Sanchez R: Adenocarcinoma of the stomach: Univariate and
multivariate analysis of factors associated with survival. Am J
Clin Oncol. 25:84–89. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Crew KD and Neugut AI: Epidemiology of
gastric cancer. World J Gastroenterol. 12:354–362. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Harrison LE, Karpeh MS and Brennan MF:
Extended lymphadenectomy is associated with a survival benefit for
node-negative gastric cancer. J Gastrointest Surg. 2:126–131. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Liu HL, Jiang WB and Xie MX: Flavonoids:
Recent advances as anticancer drugs. Recent Pat Anticancer Drug
Discov. 5:152–164. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Park KI, Park HS, Kang SR, Nagappan A, Lee
DH, Kim JA, Han DY and Kim GS: Korean Scutellaria baicalensis water
extract inhibits cell cycle G1/S transition by suppressing cyclin
D1 expression and matrix-metalloproteinase-2 activity in human lung
cancer cells. J Ethnopharmacol. 133:634–641. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yan H, Xin S, Wang H, Ma J, Zhang H and
Wei H: Baicalein inhibits MMP-2 expression in human ovarian cancer
cells by suppressing the p38 MAPK-dependent NF-κB signaling
pathway. Anticancer drugs. 26:649–656. 2015.PubMed/NCBI
|
|
9
|
Hong GE, Kim JA, Nagappan A, Yumnam S, Lee
HJ, Kim EH, Lee WS, Shin SC, Park HS and Kim GS: Flavonoids
identified from Korean Scutellaria baicalensis Georgi inhibit
inflammatory signaling by suppressing activation of NF-κB and MAPK
in RAW 264.7 cells. Evid Based Complement Alternat Med.
2013:9120312013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kim JA, Nagappan A, Park HS, Saralamma VV,
Hong GE, Yumnam S, Lee HJ, Raha S, Kim EH, Young PS and Kim GS:
Proteome profiling of lipopolysaccharide induced L6 rat skeletal
muscle cells response to flavonoids from Scutellaria baicalensis
Georgi. BMC Complement Altern Med. 14:3792014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kavandi L, Lee LR, Bokhari AA, Pirog JE,
Jiang Y, Ahmad KA and Syed V: The Chinese herbs Scutellaria
baicalensis and Fritillaria cirrhosa target NFκB to inhibit
proliferation of ovarian and endometrial cancer cells. Mol
Carcinog. 54:368–378. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Taylor RC, Cullen SP and Martin SJ:
Apoptosis: Controlled demolition at the cellular level. Nat Rev Mol
Cell Biol. 9:231–241. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Saraste A and Pulkki K: Morphologic and
biochemical hallmarks of apoptosis. Cardiovasc Res. 45:528–537.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hacker G: The morphology of apoptosis.
Cell Tissue Res. 301:5–17. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Logue SE and Martin SJ: Caspase activation
cascades in apoptosis. Biochem Soc Trans. 36:1–9. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Cory S and Adams JM: The Bcl2 family:
Regulators of the cellular life-or-death switch. Nat Rev Cancer.
2:647–656. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ademosun AO, Oboh G, Passamonti S, Tramer
F, Ziberna L, Boligon AA and Athayde ML: Inhibition of
metalloproteinase and proteasome activities in colon cancer cells
by citrus peel extracts. J Basic Clin Physiol Pharmacol.
26:471–477. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ye F, Jiang S, Volshonok H, Wu J and Zhang
DY: Molecular mechanism of anti-prostate cancer activity of
Scutellaria baicalensis extract. Nutr Cancer. 57:100–110. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Newman DJ and Cragg GM: Natural products
as sources of new drugs over the 30 years from 1981 to 2010. J Nat
Prod. 75:311–335. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ubah OC and Wallace HM: Cancer therapy:
Targeting mitochondria and other sub-cellular organelles. Curr
Pharm Des. 20:201–222. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Danial NN and Korsmeyer SJ: Cell death:
Critical control points. Cell. 116:205–219. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Fesik SW: Promoting apoptosis as a
strategy for cancer drug discovery. Nat Rev Cancer. 5:876–885.
2005. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang L, Cheng X, Gao Y, Zheng J, Xu Q,
Sun Y, Guan H, Yu H and Sun Z: Apigenin induces autophagic cell
death in human papillary thyroid carcinoma BCPAP cells. Food Funct.
6:3464–3472. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Nagappan A, Lee HJ, Saralamma VV, Park HS,
Hong GE, Yumnam S, Raha S, Charles SN, Shin SC, Kim EH, et al:
Flavonoids isolated from Citrus platymamma induced G2/M cell cycle
arrest and apoptosis in A549 human lung cancer cells. Oncol Lett.
12:1394–1402. 2016.PubMed/NCBI
|
|
26
|
Lee DH, Park KI, Park HS, Kang SR,
Nagappan A, Kim JA, Kim EH, Lee WS, Hah YS, Chung HJ, et al:
Flavonoids isolated from Korea Citrus aurantium L. induce G2/M
phase arrest and apoptosis in human gastric cancer AGS cells. Evid
Based Complement Alternat Med. 2012:5159012012.PubMed/NCBI
|
|
27
|
Park KI, Park HS, Nagappan A, Hong GE, Lee
DH, Kang SR, Kim JA, Zhang J, Kim EH, Lee WS, et al: Induction of
the cell cycle arrest and apoptosis by flavonoids isolated from
Korean Citrus aurantium L. in non-small-cell lung cancer cells.
Food Chem. 135:2728–2735. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Saralamma VV, Nagappan A, Hong GE, Lee HJ,
Yumnam S, Raha S, Heo JD, Lee SJ, Lee WS, Kim EH and Kim GS:
Poncirin induces apoptosis in AGS human gastric cancer cells
through extrinsic apoptotic pathway by up-regulation of fas ligand.
Int J Mol Sci. 16:22676–22691. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Stennicke HR and Salvesen GS: Properties
of the caspases. Biochim Biophys Acta. 1387:17–31. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kumar S: Regulation of caspase activation
in apoptosis: Implications in pathogenesis and treatment of
disease. Clin Exp Pharmacol Physiol. 26:295–303. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tait SW, Ichim G and Green DR: Die another
way-non-apoptotic mechanisms of cell death. J Cell Sci.
127:2135–2144. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Tait SW and Green DR: Mitochondria and
cell death: Outer membrane permeabilization and beyond. Nat Rev Mol
Cell Biol. 11:621–632. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hyun HB, Lee WS, Go SI, Nagappan A, Park
C, Han MH, Hong SH, Kim G, Kim GY, Cheong J, et al: The flavonoid
morin from Moraceae induces apoptosis by modulation of Bcl-2 family
members and Fas receptor in HCT 116 cells. Int J Oncol.
46:2670–2678. 2015.PubMed/NCBI
|
|
34
|
Zhang SD, Shan L, Li W, Li HL and Zhang
WD: Isochamaejasmin induces apoptosis in leukemia cells through
inhibiting Bcl-2 family proteins. Chin J Nat Med. 13:660–666.
2015.PubMed/NCBI
|