Introduction
The difference in energy production between cancer
cells and normal cells is considered to be one of the available
targets for the treatment of cancer (1). Unlike normal cells, cancer cells obtain
energy mostly via glycolysis, while mitochondrial oxidative
phosphorylation is reduced (2). As
tumor cells actively proliferate, they require a constant energy
supply and it has been postulated that they switch to aerobic
glycolysis characterized by rapid ATP production, which is known as
the Warburg effect (2). It is
hypothesized that the purpose of the Warburg effect is to avoid the
excessive generation of reactive oxygen species (ROS) during
oxidative phosphorylation in mitochondria, and the subsequent
oxidative stress that causes cell damage (1,3). The
differences in the energy metabolism between tumor and normal cells
have previously been exploited in cancer treatment, and the
efficacy of a number of currently used chemotherapeutic agents,
including methotrexate, depends on ROS production (3).
Excessive oxidative stress can be generated by
forcing cancer cells to produce energy via mitochondrial oxidation,
which is expected to cause anticancer effects without injury to
benign cells, as for the latter it is a normal metabolic process
(4). A previous study indicated that
ketone bodies and medium-chain fatty acids (MCFAs) may be used as a
tool to induce cancer cells to switch between glycolysis and
oxidative phosphorylation for energy production, indicating that
ketogenic and MCFA-enriched diets may be beneficial in cancer
therapy (5).
In mitochondria, fatty acids are metabolized to
acetyl-CoA via β-oxidation, which can be diverted to the formation
of ketone bodies under the conditions of glucose starvation
(5,6).
β-hydroxybutyric acid (β-HBA, also termed 3-HBA) is one of the
three ketone bodies generated from fatty acids; it has been
revealed that 3-HBA can be used for energy production in
mitochondria through the tricarboxylic acid cycle, where it is
converted into acetoacetic acid and then to acetyl-CoA (7,8). Ketogenic
diets rich in medium-chain triglycerides demonstrated inhibitory
effects on cancer growth, and it was observed that prostate cancer
cells had lower ability to utilize dietary fatty acids compared
with normal prostate cells, indicating possible therapeutic
potential. Lauric acid (LAA) is an MCFA with an aliphatic tail of
12 carbons, which, contrary to long-chain fatty acids (LCFAs), can
be transported to the mitochondrial intermembrane space directly
without the carnitine shuttle (9).
Therefore, LAA is utilized through β-oxidation with higher efficacy
than LCFAs (10). The present study
examined the effect of 3-HBA and LAA on cancer cell proliferation,
oxidative stress and stemness, with the aim of providing
mechanistic insights into a potentially therapeutic effect of
ketogenic diets for patients with cancer.
Materials and methods
Cell culture and treatment
The mouse CT26 colon cancer cell line was provided
by Professor I. J. Fidler (MD Anderson Cancer Center, Texas
University, Austin, TX, USA). Cells were cultured in Dulbecco's
modified Eagle's medium containing high glucose (450 mg/dl), low
glucose (100 mg/dl) or no glucose (Wako Pure Chemical Industries,
Ltd., Osaka, Japan), and supplemented with 10% fetal bovine serum
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) at 37°C in 5% CO2.
Cells were treated with 3-HBA (1 mM; Sigma-Aldrich; Merck KGaA)
and/or LAA (1.5 mM; Wako Pure Chemical Industries, Ltd.) for 24 h
at 37°C, and analyzed for viability using a tetrazolium dye (MTT;
Sigma-Aldrich; Merck KGaA) assay, as previously described (11). 3-HBA was solubilized using PBS at room
temperature. LAA was solubilized in 60°C PBS and kept at 37°C until
use. All procedures were performed in accordance with the Ethical
Guidelines for Human Genome/Gene Research enacted by the Japanese
Government, which was approved by the Ethics Committee of Nara
Medical University (approval no., 937-1).
Mitochondrial volume was assessed by staining
mitochondria with MitoGreen by adding MitoGreen-dimethyl sulfoxide
solution (200 µM) to cultured cells (50 nM at working
concentration) for 1 h at 37°C (Takara Bio, Inc., Otsu, Japan).
Cell images were captured under a fluorescence microscope equipped
with a camera (Leica Microsystems, Inc., Buffalo Grove, IL, USA),
and fluorescence was evaluated in the images using Photoshop Image
Analyzer (CS2; version 9.0.2; Adobe Systems, Inc., San Jose, CA,
USA). Alkaline phosphatase (ALP) activity was determined using a
commercial kit according to the manufacturer's protocol (LabAssay
ALP; catalog no., 291-58601; Wako Pure Chemical Industries, Ltd.).
GSSG (by High-Throughput Glutathione Assay kit) and 4-HNE (by
OxiSelect HNE Adduct ELISA Kit) were measured in cell lysates using
ELISA kits (R&D Systems, Inc., Minneapolis, MN, USA), according
to the manufacturer's protocol. Whole-cell lysates were prepared in
0.1% Nonidet 40-containing lysis buffer (Wako Pure Chemical
Industries, Ltd.) as previously described (12).
Reverse transcription-polymerase chain
reaction (RT-PCR)
Total RNA was extracted using an RNeasy kit from
1×107 CT26 cells according to the manufacturer's
protocol (Qiagen, Inc., Valencia, CA, USA). cDNA was synthesized
from 0.5 µg RNA using ReverTra Ace RT-qPCR kit (Toyobo Co., Ltd.,
Osaka, Japan) and used as a template for q-PCR. The thermocycler
settings were 30 cycles of 96°C for 30 sec, 64°C for 30 sec and
72°C for 30 sec, followed by 72°C for 10 min. The expression of
monocarboxylic acid transporter (Mct)1, Mct5, succinyl-CoA
transferase (Scot), NADH dehydrogenase (ubiquinone) 1 α subcomplex,
9 (Ndufa9), succinate dehydrogenase complex, subunit A,
flavoprotein (Sdha), Ubiquinol cytochrome c reductase core
protein 2 (Uqcrc2), cytochrome c oxidase II (mtCo2), ATP
synthase, H+ transporting, mitochondrial F0 complex, subunit F2
(Atp5j2), F1 ATPase inhibitor (If1), PTEN-induced putative kinase 1
(Pink1) and GAPDH were quantified. Primers were synthesized by
Sigma Genosys (Ishikari, Japan); the sequences are presented in
Table I. PCR products were separated
by electrophoresis in 2% agarose, and then stained with ethidium
bromide (Wako Pure Chemical Industries, Ltd.) for 30 min at room
temperature. The level present in untreated cells grown under 450
mg/dl glucose was set as 10%. For Pink1, semi-quantitative
analysis, the level present in untreated cells grown under glucose
free-conditions was set as 100%. Semi-quantitation was performed on
captured gel images from an EPSON image scanner (Tokyo, Japan). The
captured images were analyzed by densitometry with NIH Image
(version 1.63; National Institute of Mental Health, Bethesda, MD,
USA). Each experiment was performed in triplicate.
 | Table I.List of primer sets for reverse
transcription-polymerase chain reaction. |
Table I.
List of primer sets for reverse
transcription-polymerase chain reaction.
| Gene name | Gene symbol | Figure
abbreviation | Accession no. | Upper primer (5′ to
3′) | Lower primer (5′ to
3′) |
|---|
| ATPase inhibitor | IF1 | If1 | AF002718.1 |
tctggggtatgaaggtcctg |
tcgaatggtggtcaatctca |
| Succinyl CoA
transferase | Scot | Scot | AB085609.1 |
ctgccttctacaccagcaca |
catgctttcaccagagcaaa |
| Solute carrier family
16 (monocarboxylic acid transporters), member 1 | Slc16a1 | Mct1 | NM_009196.4 |
gctggaggtcctatcagcag |
agttgaaagcaagcccaaga |
| Solute carrier family
16 (monocarboxylic acid transporters), member 5 | Slc16a5 | Mct5 | NM_001080934.1 |
cctgtccatgtgtggagttg |
cccacacccaggataagaga |
| NADH dehydrogenase
(ubiquinone) 1 alpha subcomplex, 9 | Ndufa9 | Ndufa9, Com I | NM_009933.4 |
acacagacctggtggagacc |
ggatgggcttggagtaatca |
| Succinate
dehydrogenase complex, subunit A, flavoprotein | Sdha | Sdha, Com II | NM_025358.3 |
actgtgtttggggctacagg |
gattgatgaccacgttgctg |
| Ubiquinol
cytochrome c reductase core protein 2 | Uqcrc2 | Uqcrc2, Com
III | NM_025899.2 |
gtcagagggcttcctgagtg |
actcgtcgagaaaaggcgta |
| Cytochrome c
oxidase II | mtCo2 | mtCo2, Com IV | AB284309.1 |
ggcagaacgactcggttatc |
acgaaatcaacaaccccgta |
| ATP synthase, H+
transporting, mitochondrial F0 complex, subunit F2 | Atp5j2 | Atp5j2, Com V | NM_020582.2 |
ccttccaccgggatttttat |
aatttggcagctatgggaga |
| PTEN induced
putative kinase 1 | Pink1 | Pink1 | NM_026880.2 |
cccacaccctaacatcatcc |
actgggagtctgctcctcaa |
|
Glyceraldehyde-3-phosphate
dehydrogenase | Gapdh | Gapdh | NM_001289726.1 |
aactttggcattgtggaagg |
acacattgggggtaggaaca |
Statistical analysis
Statistical significance was calculated using
two-tailed Fisher's exact, χ2 and unpaired Student
t-tests using InStat software (version 3.0; GraphPad Software,
Inc., La Jolla, CA, USA). Data are expressed as the mean ± standard
deviation of three independent experiments. P<0.05 (two-sided)
was considered to indicate a statistically significant
difference.
Results
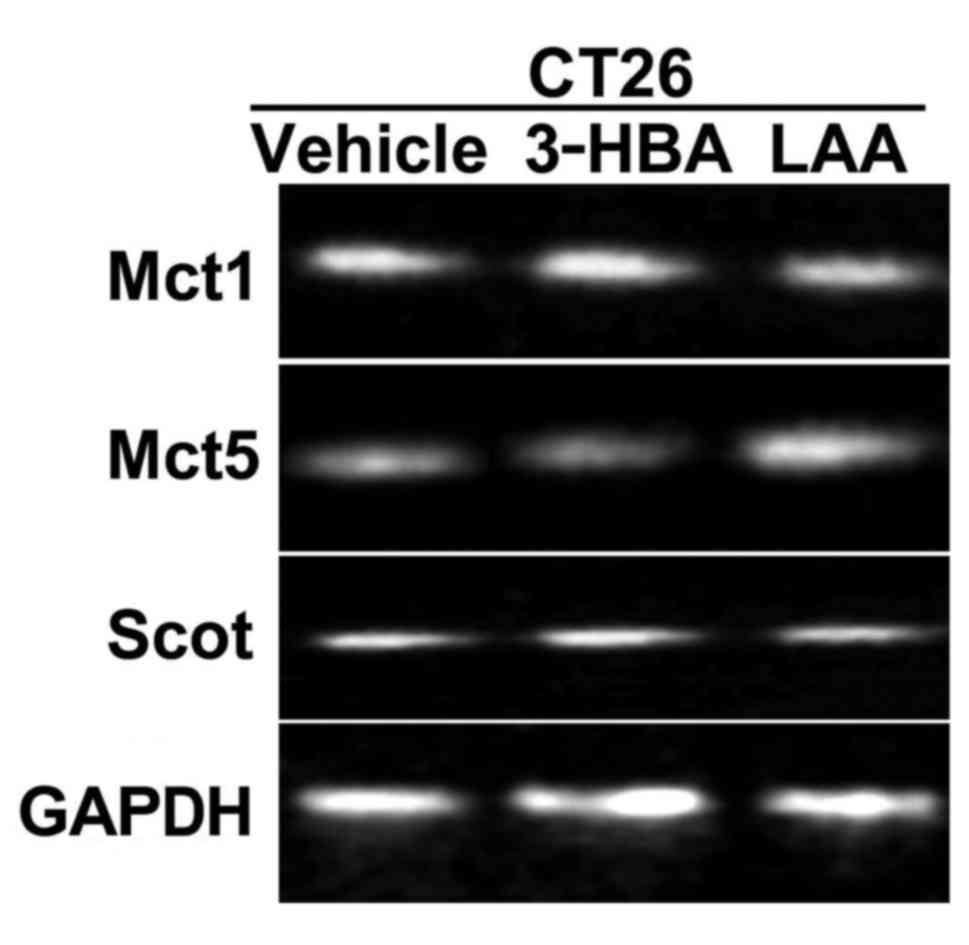
Expression of fatty acid transporters
and ketone body metabolism-associated genes in CT26 cells
First, the mRNA expression of transporters for
short-chain fatty acids (SCFAs) and MCFAs in CT26 cells treated
with 3-HBA and LAA was determined (Fig.
1). 3-HBA has an aliphatic chain similar to that of SCFAs and
is transported by monocarboxylate transporters (Mcts) 1–4,
while LAA is an MCFA transported by Mcts 5–7 (13,14). The
results indicated that CT26 cells expressed Mct1 and
Mct5. CT26 cells also expressed 3-oxoacid CoA transferase
(SCOT), a key enzyme involved in metabolizing 3-HBA to acetoacetate
(15,16); however, mRNA levels were low and not
significantly affected by 3-HBA or LAA treatment (Fig. 1).
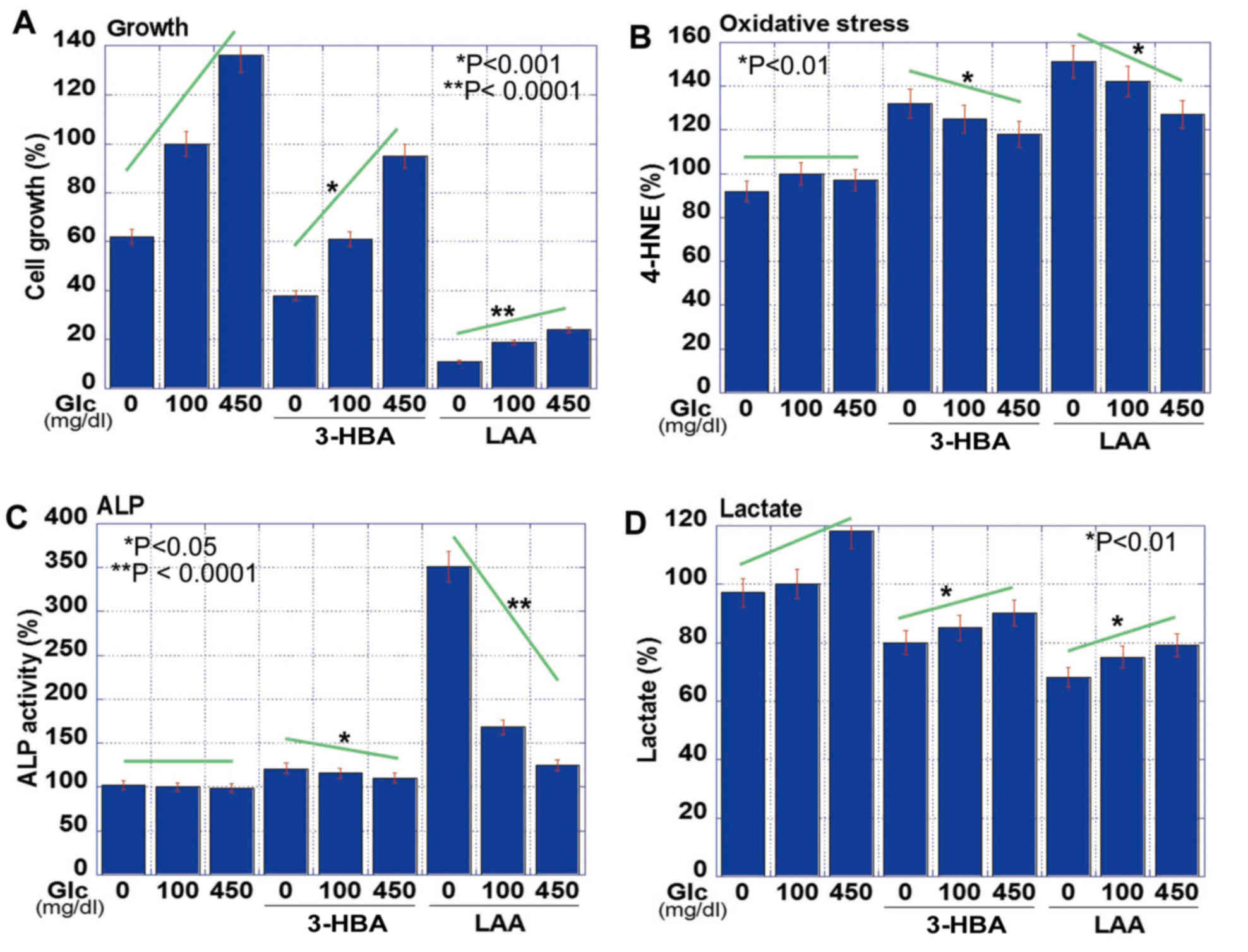
Effect of 3-HBA and LAA on CT26 cell
proliferation, stemness and oxidative stress
The effect of 3-HBA and LAA on CT26 cells grown in
the presence of various glucose concentrations was subsequently
examined (Fig. 2). The results
revealed that the proliferation of CT26 cells was correspondingly
increased with the glucose concentration, but the addition of 3-HBA
or LAA into the medium inhibited cell proliferation at each glucose
concentration (Fig. 2A). The
suppressive effect of LAA on cell growth was markedly high. Lipid
peroxidation assessed based on 4-HNE levels was not significantly
different in any glucose concentration. By contrast, 4-HNE levels
were upregulated with 3-HBA, and more so with LAA (Fig. 2B). Cancer cell stemness was evaluated
via ALP activity (17), which was
inversely associated with the glucose concentration (Fig. 2C). The 3-HBA increased ALP activity in
glucose-free, but not in glucose-supplemented, medium. By contrast,
LAA induced ALP activity under all conditions. Lactate
concentration in the medium increased with the glucose
concentration, but was decreased following the administration of
3-HBA and LAA (Fig. 2D).
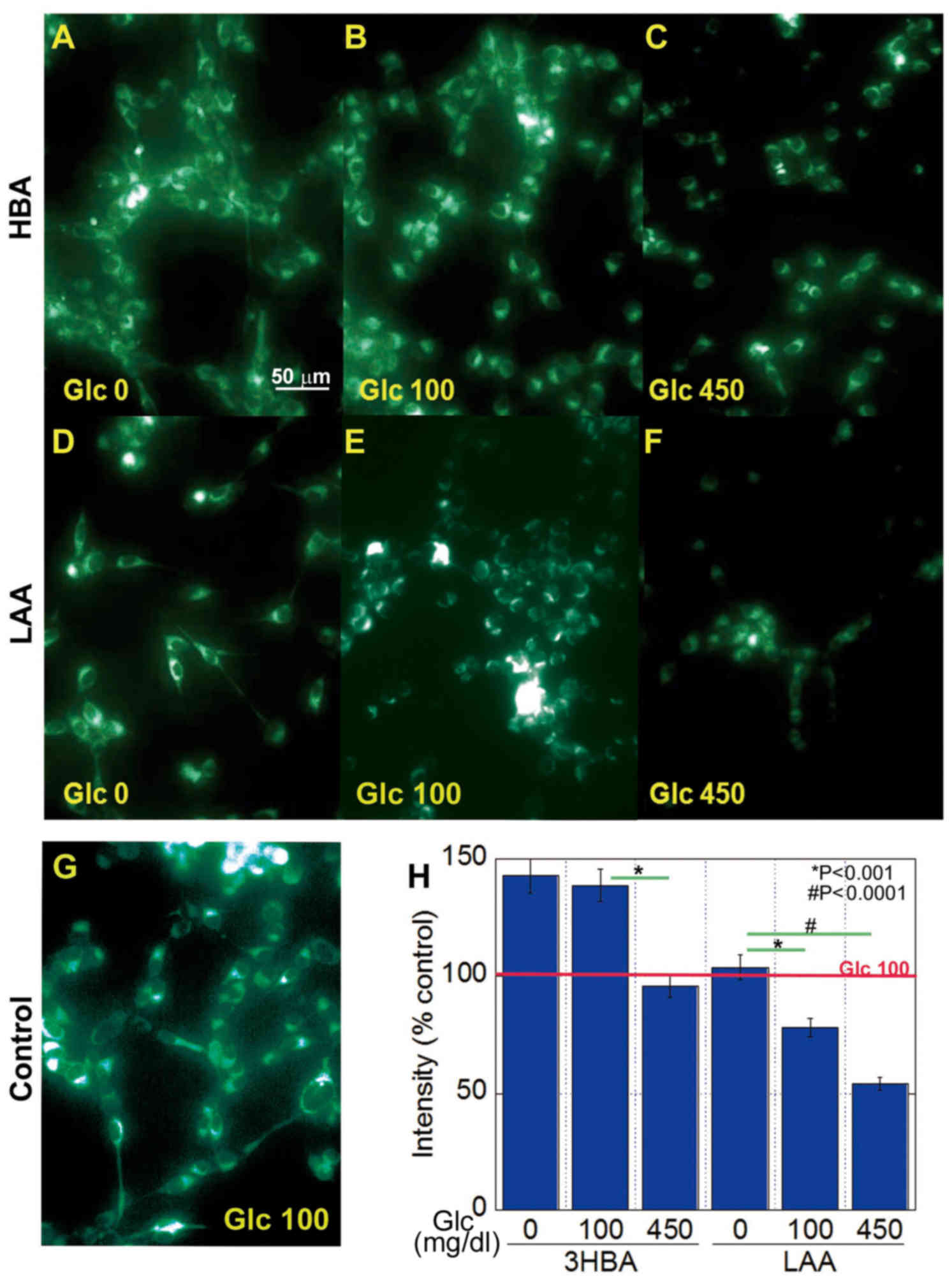
Effect of 3-HBA and LAA on
mitochondrial volume in CT26 cells
The effect of 3-HBA and LAA on mitochondrial volume
in CT26 cells was then examined, and it was revealed that it
inversely associated with the glucose concentration in the cells
treated with 3-HBA or LAA (Fig. 3).
The 3-HBA increased mitochondrial volume at glucose concentrations
of 0 and 100 mg/dl (Fig. 3A and B).
By contrast, LAA decreased mitochondrial volume at glucose
concentrations of 100 and 450 mg/dl (Fig.
3D-H).
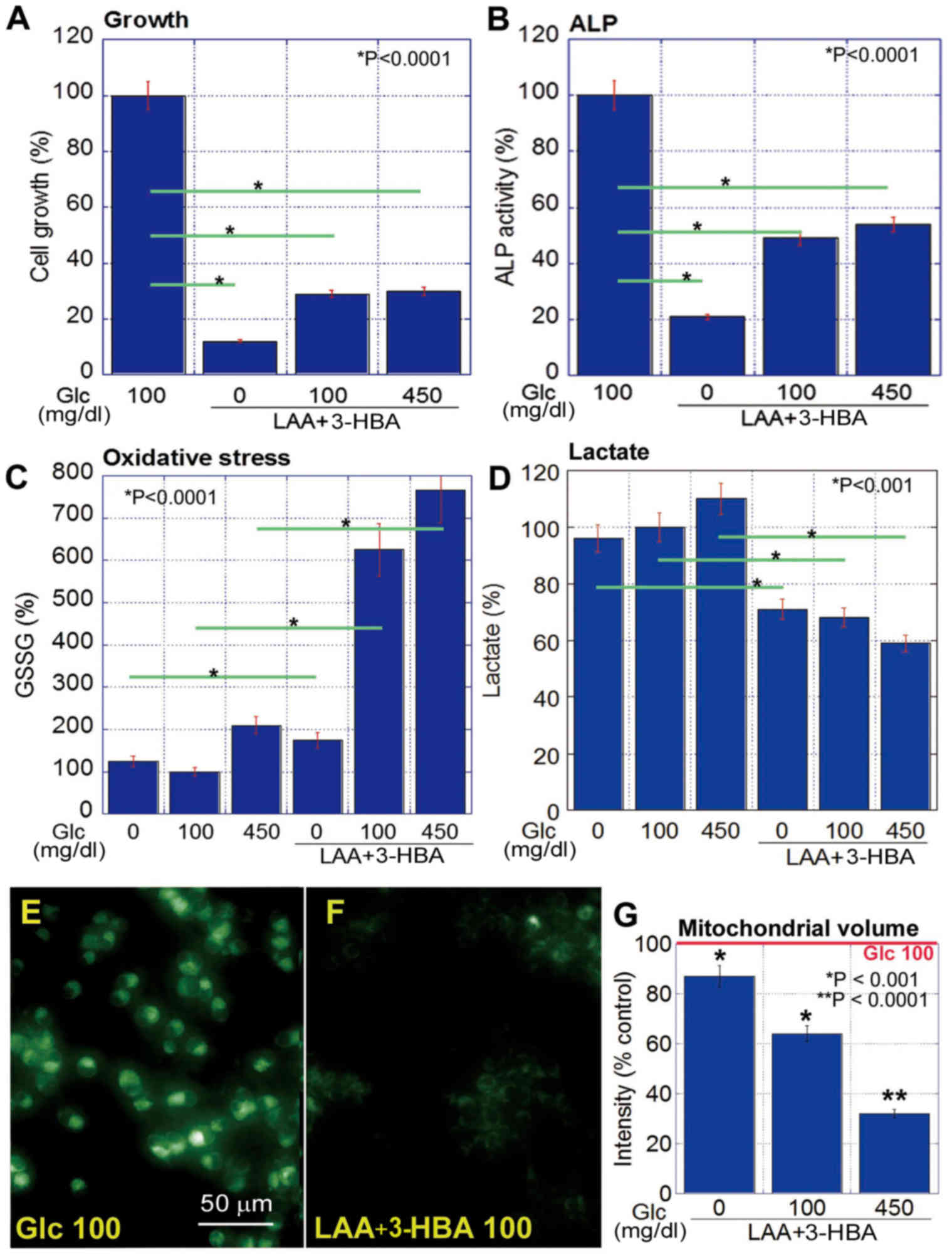
Effect of concurrent LAA and 3-HBA
treatment on CT26 cell proliferation, stemness and oxidative
stress
The concurrent treatment with LAA and 3-HBA
inhibited cell proliferation and ALP activity in CT26 cells
(Fig. 4A and B) and significantly
increased GSSG (Fig. 4C), whilst also
decreasing lactate (Fig. 4D) under
all glucose concentrations (P<0.05). LAA and 3-HBA decreased
mitochondrial volume at all glucose concentrations, compared with
that at 100 mg/ml glucose alone; however, higher glucose
concentrations induced a more pronounced decrease in mitochondrial
volume (Fig. 4E-G).
Effect of 3-HBA and LAA on the
expression of enzyme complexes involved in oxidative
phosphorylation
The increase in oxidative stress and alterations to
mitochondrial volume indicated that 3-HBA and LAA may affect
oxidative phosphorylation in mitochondria. Therefore, the mRNA
expression of enzymatic complexes in the oxidative phosphorylation
system was examined by RT-PCR (Fig.
5). Semi-quantitative analysis of the results revealed that the
expression of electron transfer system complexes in the
mitochondria was altered by glucose concentration, 3-HBA and/or LAA
treatments in comparison with those in glucose-free medium
(Fig. 5B). In detail, treatment with
3-HBA decreased the levels of complex V under glucose-free and
normal (100 mg/dl) glucose concentrations, whereas LAA administered
alone downregulated the expression of all complexes in glucose-free
conditions, and when in combination with 3-HBA reduced the
expression of complexes under glucose-free as well as high-glucose
conditions (Fig. 5B).
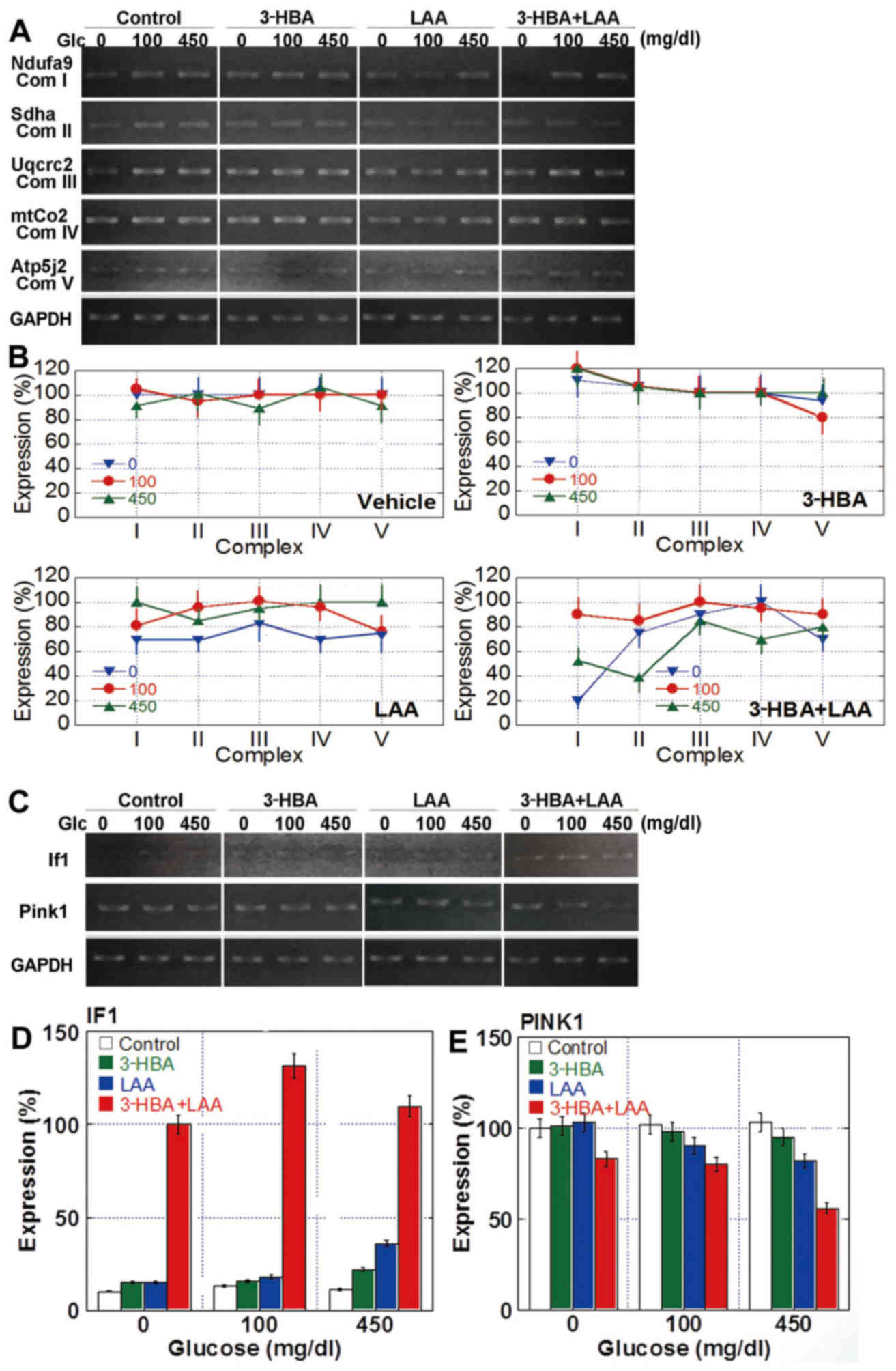 | Figure 5.Expression of mitochondrial genes in
CT26 cells treated with 3-HBA and/or LAA. (A) Expression of the
Ndufa9 (complex I), Sdha (complex II), Uqcrc2
(complex III), mtCo2 (complex IV) and Atp5j2 (complex
V) genes were examined by RT-PCR. GAPDH, loading control. (B)
Semi-quantitative analysis of the expression of mitochondrial
complex enzymes. (C) Expression of the If1- and
Pink1-encoding genes was examined by RT-PCR. GAPDH, loading
control. (D) Semi-quantitative analysis of IF1 expression. The
level present in untreated cells grown under 450 mg/dl glucose was
set as 10%. (E) Semi-quantitative analysis of Pink1 expression. The
level present in untreated cells grown under glucose
free-conditions was set as 100%. The data are expressed as the mean
± standard deviation of three independent experiments. RT-PCR,
reverse transcription-polymerase chain reaction; 3-HBA,
3-hydroxybutyric acid; LAA, lauric acid; If1, F1 ATPase inhibitor;
Pink1, protein phosphatase and tensin homolog-induced kinase 1;
Sdha, succinate dehydrogenase complex, subunit; Uqcrc2, ubiquinol
cytochrome c reductase core protein 2; mtCo2, cytochrome
c oxidase II; Ndufa9, NADH dehydrogenase (ubiquinone) 1 α
subcomplex, 9; Atp5j2, ATP synthase, H+ transporting, mitochondrial
F0 complex, subunit F2. |
Furthermore, gene expression of the mitochondrial
volume-associated factors inhibitor factor 1 (If1) and
protein phosphatase and tensin homolog-induced kinase 1
(Pink1) was examined (Fig. 5C and
D). In the untreated control cells, IF1 expression was not
detected, irrespective of the glucose concentration; however, 3-HBA
and LAA upregulated If1 mRNA to detectable levels in each
condition, and the concurrent treatment of 3-HBA with LAA markedly
increased IF1 expression. Pink1 expression was constitutive
in control cells, irrespective of the glucose concentration. The
3-HBA treatment did not significantly affect Pink1 levels;
however, they were decreased by treatment with LAA and with 3-HBA +
LAA, particularly in cells cultured at high glucose
concentrations.
Discussion
In the present study, it was identified that the
addition of 3-HBA or LAA inhibited the growth of CT26 colon cancer
cells. In normal tissues, 3-HBA and LAA are utilized for metabolic
processes, including energy production and lipogenesis,
particularly in hypoglycemic conditions (13–16). The
MCT isoforms 1–4 are transporters of SCFAs and monocarboxylates,
including 3-HBA (13). It has been
demonstrated that, in a hypermetabolic state, MCTs 1–4 are
upregulated in the brain in order to transport ketone bodies and
lactate (14). MCFAs, however, are
transported by MCTs 5–7 and, in contrast to LCFAs, do not require
the carnitine shuttle to be imported into mitochondria (6,15).
Cancer cells can utilize ketone bodies or LAA
(16,18). It was previously revealed that gastric
cancer cells could continue growing in glucose-free medium
supplemented with ketone bodies (16). Utilization of ketone bodies in gastric
cancer cells is regulated by the expression of SCOT, a key enzyme
involved in ketone body metabolism (16), and it was reported that neuroblastoma
cells were unable to use ketone bodies as an energy source, due to
decreased SCOT expression (18).
Therefore, the present study examined the expression
of MCT1, MCT5 and SCOT, which enable the utilization of 3-HBA and
LAA in colon cancer cells. However, the levels of SCOT expression
were low, which is consistent with the lower SCOT levels in cancer
cells compared with in cancer-associated fibroblasts observed in
colorectal cancer (19). By contrast,
the carnitine shuttle enzyme carnitine palmitoyltransferase 1A
(CPT1), which is important for fatty acid transport into
mitochondria, was expressed at high levels in CT26 cells (20). The experimental concentration of 3-HBA
(1 mM) in the present study was equivalent to serum levels of
ketone bodies present in individuals fed a carbohydrate-limited
diet (21). The experimental
concentration of LAA was also equivalent to the serum fatty acid
concentration (22). Thus, LAA may
provide higher mitochondria-activating effect on energy metabolism
compared with 3-HBA.
In normal tissues, SCFAs and MCFAs inhibit
glycolysis and stimulate lipogenesis or gluconeogenesis (15). SCFAs and MCFAs exhibit no or weak
protonophoric and lytic activities in mitochondria, and decrease
the efficacy of oxidative ATP synthesis (15). However, SCFAs and MCFAs also increase
the mitochondrial respiratory capacity in normal physiological and
inflammatory conditions (23). MCFAs
also stimulate fatty acid oxidization and energy production in
mitochondria in type II skeletal muscle (24). In cancer, SCOT, the rate-limiting
enzyme for the production of ketone bodies, and CPT1, the carnitine
shuttle protein, is associated with the cell metastatic potential,
as evidenced by their increased expression in the highly metastatic
SW620 cells, as compared with in the low metastatic parental SW480
cells (25). In the present study,
3-HBA and LAA decreased lactate fermentation, particularly under
glucose starvation, indicating that 3-HBA and LAA switch the energy
production pathway from glycolysis and lactate fermentation to
oxidative phosphorylation.
Oxidative phosphorylation is considered a major
intracellular source of oxidative stress (26). Activation of mitochondria is followed
by oxidative stress, which may induce cell and tissue damage
(26). Hydrogen peroxide release is
increased in mitochondria during the oxidation of MCFAs supplied in
the form of carnitine esters (27).
Consistent with these findings, the present results indicated that
3-HBA and LAA increased oxidative stress in parallel with the
enhancement of energy production in mitochondria.
The imbalance in the complexes of the electron
transport chain (ETC), characteristic of aged mitochondria, induces
the generation of ROS (28). Notably,
3-HBA and LAA caused the imbalance of ETC complexes in addition to
the downregulation of PINK1, which is a major factor in
PINK1-Parkin-mediated mitophagy (29). The decrease in PINK1 suggests
retardation of the mitochondrial turnover, which may promote the
imbalance of ETC complexes and result in ROS production, which in
turn may damage the mitochondria. The present data suggested that
3-HBA- and LAA-associated oxidative stress is associated with the
decrease in mitochondrial volume. The depletion of proteins
involved in autophagy leads to mitochondrial dysfunction, decreased
ATP production and increased ROS release, ultimately resulting in
the apoptosis of mesenchymal stem cells (30).
Thus, the present data suggested that ketone bodies
and MCFAs may exert anticancer effects via the deviation of energy
metabolism from glycolysis-lactate fermentation to oxidative
phosphorylation. Additional studies are required to compare the
responses to ketone bodies and MCFAs in normal and cancer tissues,
in order to verify the therapeutic effect of LAA and 3-HBA dietary
supplementation for patients with cancer.
Acknowledgements
The authors thank Ms. Tomomi Masutani for expert
assistance with the preparation of this manuscript. The present
study was supported by Mext Kakenhi (grant nos. 14478268 and
16675788).
References
|
1
|
Chen X, Qian Y and Wu S: The Warburg
effect: Evolving interpretations of an established concept. Free
Radic Biol Med. 79:253–263. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Warburg O: On respiratory impairment in
cancer cells. Science. 124:269–270. 1956.PubMed/NCBI
|
|
3
|
Hess JA and Khasawneh MK: Cancer
metabolism and oxidative stress: Insights into carcinogenesis and
chemotherapy via the non-dihydrofolate reductase effects of
methotrexate. BBA Clin. 3:152–161. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Boland ML, Chourasia AH and Macleod KF:
Mitochondrial dysfunction in cancer. Front Oncol. 3:2922013.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Dueregger A, Schöpf B, Eder T, Höfer J,
Gnaiger E, Aufinger A, Kenner L, Perktold B, Ramoner R, Klocker H
and Eder IE: Differential utilization of dietary fatty acids in
benign and malignant cells of the prostate. PLoS One.
10:e01357042015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Papamandjaris AA, MacDougall DE and Jones
PJ: Medium chain fatty acid metabolism and energy expenditure:
Obesity treatment implications. Life Sci. 62:1203–1215. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Newman JC and Verdin E: β-hydroxybutyrate:
Much more than a metabolite. Diabetes Res Clin Pract. 106:173–181.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fukao T, Mitchell G, Sass JO, Hori T, Orii
K and Aoyama Y: Ketone body metabolism and its defects. J Inherit
Metab Dis. 37:541–551. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
McCarty MF and DiNicolantonio JJ: Lauric
acid-rich medium-chain triglycerides can substitute for other oils
in cooking applications and may have limited pathogenicity. Open
Heart. 3:e0004672016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fulgencio JP, Kohl C, Girard J and
Pégorier JP: Troglitazone inhibits fatty acid oxidation and
esterification, and gluconeogenesis in isolated hepatocytes from
starved rats. Diabetes. 45:1556–1562. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kuniyasu H, Yano S, Sasaki T, Sasahira T,
Sone S and Ohmori H: Colon cancer cell-derived high mobility group
1/amphoterin induces growth inhibition and apoptosis in
macrophages. Am J Pathol. 166:751–760. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kuniyasu H, Oue N, Wakikawa A, Shigeishi
H, Matsutani N, Kuraoka K, Ito R, Yokozaki H and Yasui W:
Expression of receptors for advanced glycation end-products (RAGE)
is closely associated with the invasive and metastatic activity of
gastric cancer. J Pathol. 196:163–170. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Schutkowski A, Wege N, Stangl GI and König
B: Tissue-specific expression of monocarboxylate transporters
during fasting in mice. PLoS One. 9:e1121182014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Takimoto M and Hamada T: Acute exercise
increases brain region-specific expression of MCT1, MCT2, MCT4,
GLUT1, and COX IV proteins. J Appl Physiol (1985). 116:1238–1250.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Schönfeld P and Wojtczak L: Short- and
medium-chain fatty acids in energy metabolism: The cellular
perspective. J Lipid Res. 57:943–954. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sawai M, Yashiro M, Nishiguchi Y, Ohira M
and Hirakawa K: Growth-inhibitory effects of the ketone body,
monoacetoacetin, on human gastric cancer cells with succinyl-CoA:
3-oxoacid CoA-transferase (SCOT) deficiency. Anticancer Res.
24:2213–2217. 2004.PubMed/NCBI
|
|
17
|
Ṧtefková K, Procházková J and Pacherník J:
Alkaline Phosphatase in Stem Cells. Stem Cell Int.
2015:6283682015.
|
|
18
|
Skinner R, Trujillo A, Ma X and Beierle
EA: Ketone bodies inhibit the viability of human neuroblastoma
cells. J Pediatr Surg. 44:212–216. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chao C, Carmical JR, Ives KL, Wood TG,
Aronson JF, Gomez GA, Djukom CD and Hellmich MR: CD133+ colon
cancer cells are more interactive with the tumor microenvironment
than CD133- cells. Lab Invest. 92:420–436. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Park JH, Vithayathil S, Kumar S, Sung PL,
Dobrolecki LE, Putluri V, Bhat VB, Bhowmik SK, Gupta V, Arora K, et
al: Fatty acid oxidation-driven src links mitochondrial energy
reprogramming and oncogenic properties in triple-negative breast
cancer. Cell Rep. 14:2154–2165. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Carneiro L, Geller S, Fioramonti X, Hébert
A, Repond C, Leloup C and Pellerin L: Evidence for hypothalamic
ketone body sensing: Impact on food intake and peripheral metabolic
responses in mice. Am J Physiol Endocrinol Metab. 310:E103–E115.
2016.PubMed/NCBI
|
|
22
|
Allen BG, Bhatia SK, Anderson CM,
Eichenberger-Gilmore JM, Sibenaller ZA, Mapuskar KA, Schoenfeld JD,
Buatti JM, Spitz DR and Fath MA: Ketogenic diets as an adjuvant
cancer therapy: History and potential mechanism. Redox Biol.
2:963–970. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hecker M, Sommer N, Voigtmann H, Pak O,
Mohr A, Wolf M, Vadász I, Herold S, Weissmann N, Morty RE, et al:
Impact of short- and medium-chain fatty acids on mitochondrial
function in severe inflammation. JPEN J Parenter Enteral Nutr.
38:587–594. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ishizawa R, Masuda K, Sakata S and
Nakatani A: Effects of different fatty acid chain lengths on fatty
acid oxidation-related protein expression levels in rat skeletal
muscles. J Oleo Sci. 64:415–421. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Provenzani A, Fronza R, Loreni F, Pascale
A, Amadio M and Quattrone A: Global alterations in mRNA polysomal
recruitment in a cell model of colorectal cancer progression to
metastasis. Carcinogenesis. 27:1323–1333. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chen Y, Zhang H, Zhou HJ, Ji W and Min W:
Mitochondrial redox signaling and tumor progression. Cancers
(Basel). 8:pii: E402016. View Article : Google Scholar
|
|
27
|
Schonfeld P and Wojtczak L: Brown adipose
tissue mitochondria oxidizing fatty acids generate high levels of
reactive oxygen species irrespective of the uncoupling protein-1
activity state. Biochim Biophys Acta. 1817:410–418. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lee YK, Jee BA, Kwon SM, Yoon YS, Xu WG,
Wang HJ, Wang XW, Thorgeirsson SS, Lee JS, Woo HG and Yoon G:
Identification of a mitochondrial defect gene signature reveals
NUPR1 as a key regulator of liver cancer progression. Hepatology.
62:1174–1189. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ploumi C, Daskalaki I and Tavernarakis N:
Mitochondrial biogenesis and clearance: A balancing act. FEBS J.
284:183–195. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ghanta S, Tsoyi K, Liu X, Nakahira K, Ith
B, Coronata AA, Fredenburgh LE, Englert JA, Piantadosi CA, Choi AM
and Perrella MA: Mesenchymal stromal cells deficient in autophagy
proteins are susceptible to oxidative injury and mitochondrial
dysfunction. Am J Respir Cell Mol Biol. 56:300–309. 2016.
View Article : Google Scholar
|