Introduction
Gastric cancer (GC) is highly prevalent in Asia, and
is the one of the leading causes of mortality, following lung
cancer and liver cancer, in Korea (1). Its development has been revealed to be a
multi-step process, ranging from chronic gastritis to atrophy,
intestinal metaplasia, dysplasia and, finally, invasive cancer
(2). The major histological type of
GCs are adenocarcinomas, which are subdivided into intestinal type,
diffuse type and mixed/unclassifiable type by Lauren classification
(3). Previous studies have
demonstrated that intestinal and diffuse types of GC evolve via
distinct genetic pathways (3–6).
Telomeres, which contain TTAGGG repeat sequences in
humans, are nucleoprotein complexes that cap each end of a
eukaryotic chromosome (7).
Mitochondrial DNA (mtDNA) differs from nuclear DNA, and multiple
copies of mtDNA are present in each mitochondrion (8). Previous studies have demonstrated that
telomere length (TL) and the mtDNA copy number (mtCN) are
associated with numerous diseases, particularly specific types of
cancer (9–12). In a number of examined cancer types,
TL and mtCN changes were significantly associated with clinical and
prognostic characteristics, suggesting an early and important
effect on carcinogenesis (10–12).
Previous studies have demonstrated a sequential accumulation of
mitochondrial genetic changes during the progression from chronic
gastritis to cancer via intestinal metaplasia and dysplasia
(11–14).
Recent studies have indicated that TL and mtCN are
positively correlated in healthy individuals and in pregnant
females (15–17). Genetic changes in the telomeres and
mtDNA independently serve important roles in cellular senescence
(18). mtDNA damage caused by
cellular senescence may contribute to the production of reactive
oxygen species (ROS), resulting in telomere shortening (18). Past reviews have emphasized the
importance of the telomere-p53-mitochondrion axis for cancer,
suggesting that this may be targeted in future cancer therapy
(19,20). Our recent studies (21,22) have
also investigated these genetic changes in GC; however, their
association has yet to be studied in cancer tissue samples.
In the present study, TL and mtCN were evaluated in
GC, including 27 diffuse and 82 intestinal type tissue samples. The
results of the present study may aid improvements in the current
understanding of GC, through identifying the role of mtDNA and TL
in GC pathogenesis.
Materials and methods
Patients and DNA extraction
Tissue samples from a total of 109 patients
(57.97±11.74 years old; 82 male and 27 female patients), who
underwent gastrectomy to treat gastric adenocarcinoma between
October 1999 and December 2001, were selected from an archive of
paraffin blocks at Keimyung University Dongsan Hospital (Daegu,
Korea). Two experienced pathologists reviewed all cases, and the
tumor and adjacent normal mucosa tissue areas were defined
according to hematoxylin and eosin stained sections. Tissue
sections were deparaffinized with 500 µl 100% xylene, and left in a
65°C water bath for 15 min. To remove the residual xylene, the
samples were washed five times with ethanol. The selected areas
from the paraffin-embedded tissues then underwent DNA extraction.
DNA was isolated using an Absolute™ DNA Extraction kit
(BioSewoom, Inc., Seoul, Korea), according to the manufacturer's
protocol. DNA quantity and quality were measured using NanoDrop
1000 (Thermo Fisher Scientific, Inc., Pittsburgh, PA, USA).
Determination of TL and mtCN
TL and mtCN were analyzed by quantitative-polymerase
chain reaction (q-PCR). For the quantitative determination of TL
and mtCN (T) relative to β-globin (a control from nucleic DNA, S),
primers for the specific amplification of telomeric repeats and
cytochrome c oxidase subunit I (COX I; a gene in mtDNA) were
selected (Table I). q-PCR was then
performed using a LightCycler® 480 II system (Roche
Diagnostics GmbH, Mannheim Germany). The PCR conditions were: 95°C
denaturation for 1 min; 40 cycles of 95°C for 10 sec; and 60°C for
30 sec. Relative TL and mtCN were determined by calculating T/S
values using the following formula:
T/S=2−∆Cq, where ∆Cq=(mean Cq
telomere or COX I)-(mean Cq β-globin) (23,24). Each
measurement was repeated in triplicate; five serially diluted
control samples were also included in each experiment.
 | Table I.Primers used in this study. |
Table I.
Primers used in this study.
| Primer | Sequence |
|---|
| Telomere |
|
|
Forward |
5′-CGGTTTGTTTGGGTTTGGGTTTGGGTTTGGGTTTGGGTT-3′ |
|
Reverse |
5′-GGCTTGCCTTACCCTTACCCTTACCC-TTACCCTTACCCT-3′ |
| COX I |
|
|
Forward |
5′-TTCGCCGACCGTTGACTATTCTCT-3′ |
|
Reverse |
5′-AAGATTATTACAAATGCATGGGC-3′ |
| β-globin |
|
|
Forward |
5′-TGTGCTGGCCCATCACTTTG-3′ |
|
Reverse |
5′-ACCAGCCA-CCACTTTCTGATAGG-3′ |
Statistical analysis
The SPSS statistical package version 19.0 for
Windows (IBM SPSS, Armonk, NY, USA) was used for all statistical
analyses. TL and mtCN are presented as the mean ± standard
deviation (SD). Pearson correlation coefficients were calculated to
evaluate the association between TL and mtCN. To further explore
the correlation between these markers and the prognosis of GC, a
fold change in the TL or mtCN in tumors (T), compared with that in
paired normal tissues (N), was calculated (T/N). Patients were
categorized into two subgroups, ‘longer’ and ‘shorter’ for TL, and
‘low’ and ‘high’ for mtCN, according to their median TL and mtCN
T/N values. Survival curves, estimated using the Kaplan-Meier
method (univariate analysis), were compared with a log-rank test.
Overall survival (OS) time was defined as the time from diagnosis
to mortality from cancer or other causes. A two-tailed P<0.05
was considered to indicate a statistically significant
difference.
Results
TL and mtCN do not significantly
differ between intestinal and diffuse GC
TL and mtCN were analyzed in samples obtained from
109 patients with GC (60.0±11.8 years of age) using q-PCR. TL and
mtCN T/N values were 11.48±1.14 and 14.86±1.35, respectively (fold
change; mean ± SD; Table I). When
stratified by Lauren classification, the mtCN was similar in
intestinal (14.39±1.61) and diffuse (16.34±2.44) types of GC
(P=0.540). TL was shorter in intestinal type samples (10.36±1.27),
as compared with in diffuse type samples (15.02±2.45); however,
this was not to a statistically significant extent (P=0.074).
TL and mtCN are not associated with
clinicopathological characteristics in patients with GC
For the consideration of the association between
clinicopathological characteristics and the TL and mtCN in GC, two
groups, ‘longer’ and ‘shorter’ for TL, and ‘low’ and ‘high’ for
mtCN, were assigned based on the median TL and mtCN T/N fold change
of the tissue samples. The clinicopathological characteristics of
each group are presented in Table
II. A shorter TL was more frequent in intestinal GC (64.6%)
compared with in diffuse GC (46.2%), although there was no
statistically significant difference (P=0.093). TL was identified
to be shorter in females (70.4%) than in males (57.3%); this was
also not statistically significant (P=0.229). Additionally, age and
other clinicopathological parameters were not determined to have
any significant associations with TL and mtCN.
| Table II.Telomere length and mitochondrial DNA
copy number in gastric cancer. |
Table II.
Telomere length and mitochondrial DNA
copy number in gastric cancer.
|
|
| Lauren
classification |
|
|---|
|
|
|
|
|
|---|
| Variables | Total | Intestinal | Diffuse | P-value |
|---|
| TL (mean ± SD) | 11.48±1.14 | 10.36±1.27 | 15.02±2.45 | 0.074 |
| MtCN (mean ± SD) | 14.86±1.35 | 14.39±1.61 | 16.34±2.44 | 0.540 |
TL and mtCN are correlated in
intestinal, but not diffuse, GC
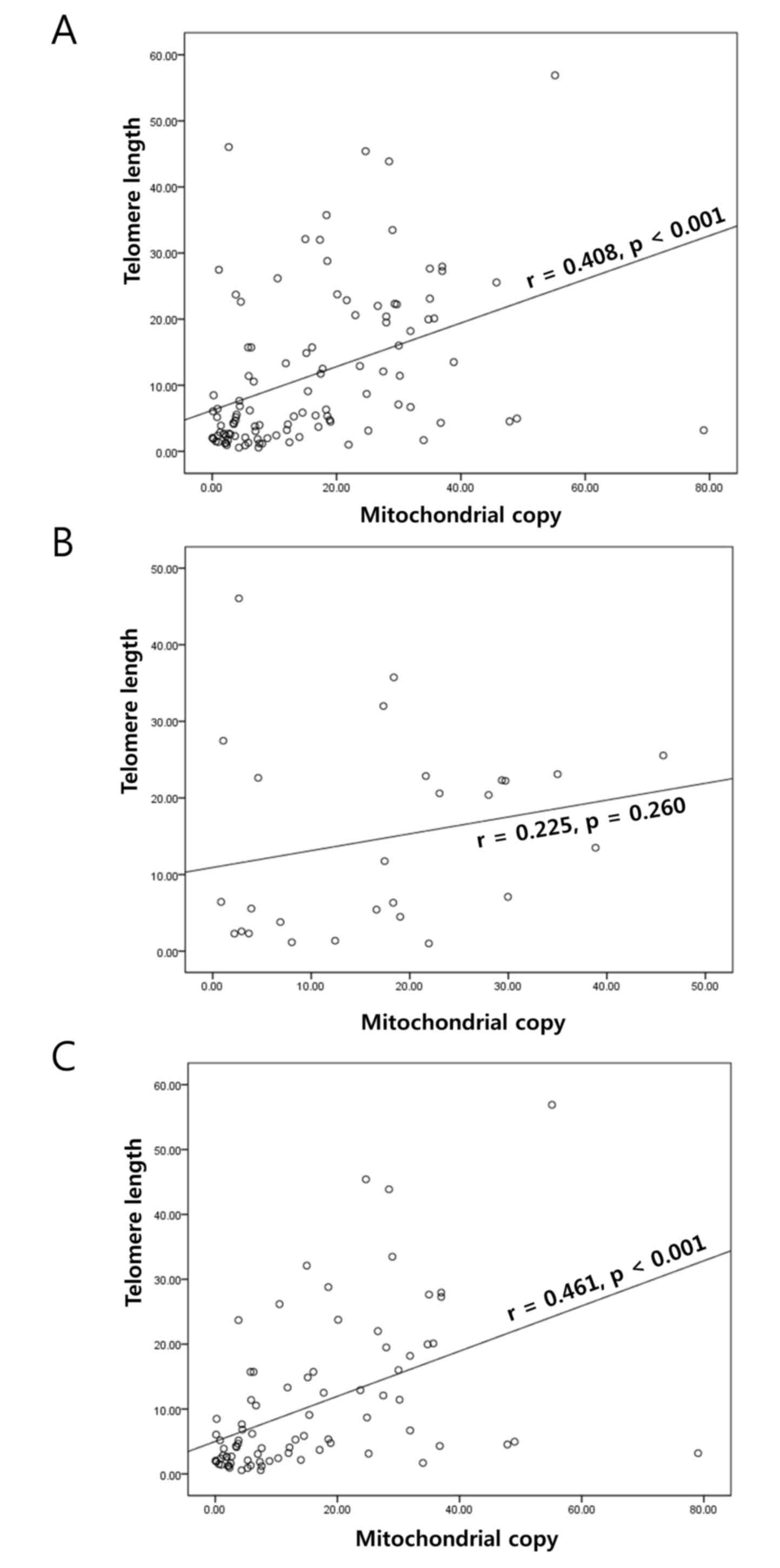
A significant positive correlation in the T/N fold
change of TL and mtCN was identified in GC (r=0.408; P<0.001;
Fig. 1A). This correlation was also
apparent in the adjacent normal mucosa (r=0.363; P<0.001)
obtained from the patients with GC. When stratified by Lauren
classification, a significant correlation was not identified in
diffuse GC (r=0.225; P=0.260; Fig.
1B); however, there was a correlation in the intestinal type
samples (r=0.461; P<0.001; Fig.
1C). The stratification did not reveal any significant
associations with other parameters (Table III).
| Table III.Clinicopathological characteristics of
TL and mtCN in gastric cancer. |
Table III.
Clinicopathological characteristics of
TL and mtCN in gastric cancer.
|
| TL (%, n) | mtCN (%, n) |
|---|
|
|
|
|
|---|
| Group | Longer | Shorter | Low | High |
|---|
| All patients | 39.5 (43) | 60.5 (66) | 69.7 (76) | 30.3 (33) |
| Age |
|
|
|
|
|
<60 | 37.0 (20) | 63.0 (34) | 72.2 (39) | 27.8 (15) |
| ≥60 | 41.8 (23) | 58.2 (32) | 67.3 (37) | 32.7 (18) |
| Gender |
|
|
|
|
| Male | 42.7 (35) | 57.3 (47) | 68.3 (56) | 31.7 (26) |
|
Female | 29.6 (8) | 70.4 (19) | 74.1 (20) | 25.9 (7) |
| pT stage |
|
|
|
|
|
1/2 | 42.3 (30) | 57.7 (41) | 68.1 (49) | 31.9 (23) |
|
3/4 | 34.2 (13) | 65.8 (25) | 73.0 (27) | 27.0 (10) |
| pN stage |
|
|
|
|
|
0/1 | 40.3 (27) | 59.7 (40) | 68.2 (60) | 31.8 (28) |
|
2/3 | 38.1 (16) | 61.9 (26) | 76.2 (16) | 83.8 (5) |
| Lauren
classification |
|
|
|
|
|
Diffuse | 53.8 (14) | 46.2 (12) | 65.4 (17) | 34.6 (9) |
|
Intestinal | 35.4 (29) | 64.6 (53) | 70.7 (58) | 29.3 (24) |
| Depth of
invasion |
|
|
|
|
|
Early | 39.1 (18) | 60.9 (28) | 71.7 (33) | 28.3 (13) |
|
Advanced | 39.7 (25) | 60.3 (38) | 68.3 (43) | 31.7 (20) |
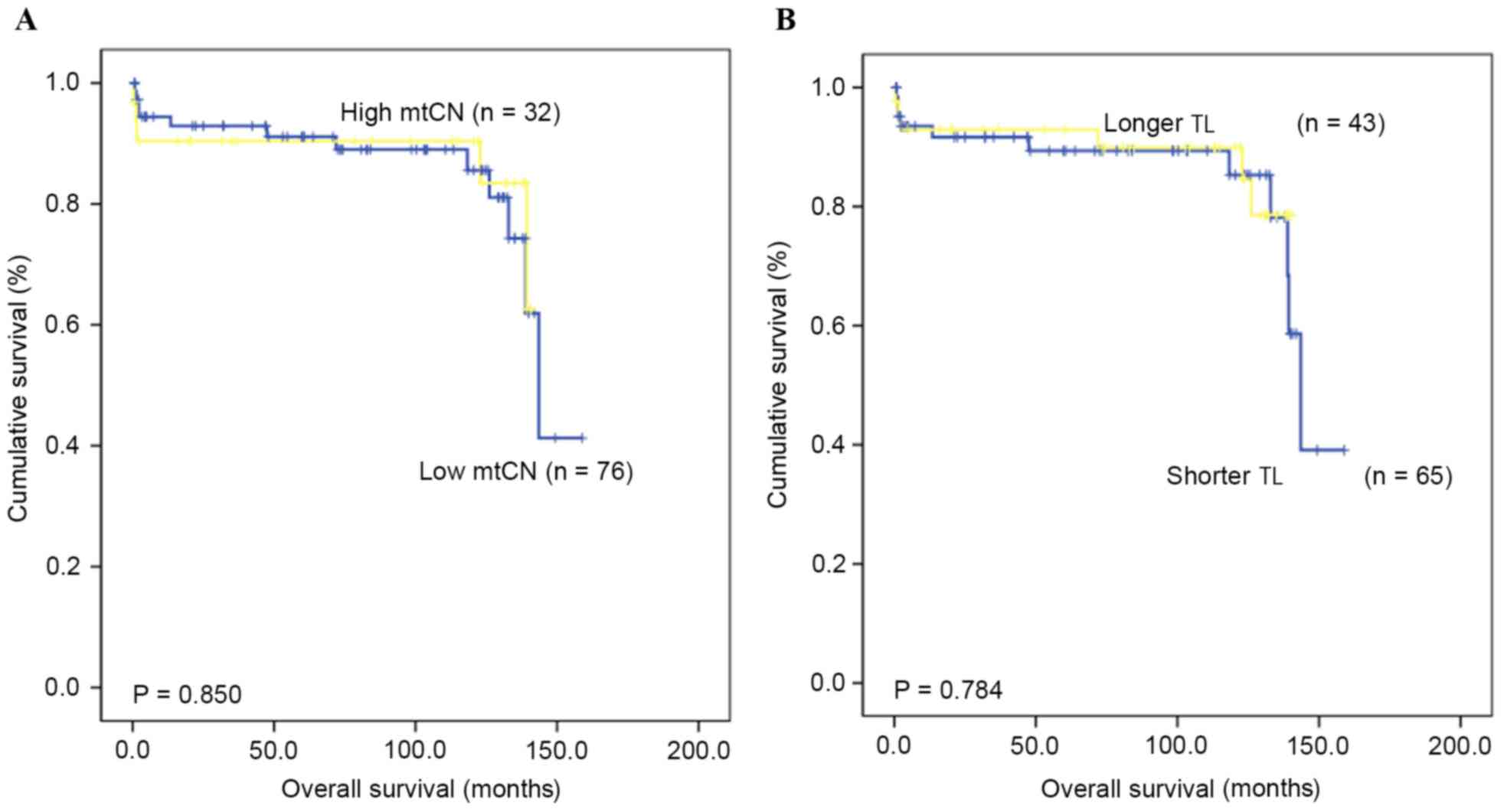
TL and mtCN are not associated with
the rate of OS
The OS time and rate for patients with GC were
assessed in order to identify if there was a prognostic value for
TL and mtCN. The median follow-up duration was 82.2 months (range,
3.7–158.8 months), and the 5-year OS rate was 79.3%. The results
indicated no prognostic value of the mtCN (P=0.850) and TL
(P=0.784) in GC, based on comparing those patients with ‘high’ and
‘low’ mtCN and TL values (Fig.
2).
Discussion
The present study identified that TL is positively
correlated with mtCN in GC tissues and paired normal tissues; this
was consistent with the results of previous studies conducted on
healthy volunteers (15–17). A correlation between TL and mtCN was
initially identified in a study of 129 healthy elderly females
(15); in a further study, it was
also demonstrated in healthy adults and pregnant females (16,17).
Furthermore, certain neuropsychiatric conditions were also
associated with TL shortening and mtCN increase, although detailed
data concerning their association were not presented (25). Taken together, there is a positive
correlation between TL alteration and mtCN not only in healthy
individuals, but also in cancer tissues.
The clinicopathological significance of TL and
mitochondrial genetic change has been previously studied in GC by a
number of groups (11,12,14,21,22).
TL was previously demonstrated to differ in GC according to
Helicobacter pylori infection status, microsatellite
instability (MSI), and non-steroidal anti-inflammatory drug use
(26–29). Although TL change was identified in
gastric carcinogenesis, it was not associated with
clinicopathological features or prognosis in the present study.
However, Pascua et al (27)
reported that a combination of TL with MSI status has prognostic
value. Mitochondrial genetic change has been frequently studied in
GC, and our previous study (30)
identified mtMSI to be a novel genetic marker for GC
susceptibility. It was demonstrated that mtMSI was present in 10.2%
of GC samples and 12.5% of gastric dysplasia samples, and that
mtMSI was associated with a poor prognosis and an increased
potential for progression.
In the present study, TL was marginally shorter in
intestinal type GC, as compared with in diffuse type GC, whereas TL
and mtCN did not have any significant association with clinical
characteristics. However, a notable association was identified in
GC tissues; in accordance with previous studies on healthy
individuals (15–17,25), mtCN
and TL were identified to be correlated with each other in normal
and cancerous tissues obtained from patients with GC. A positive
association was also identified in intestinal type GC, but not in
the diffuse type. These results suggest that the normal positive
correlation between telomeres and mitochondrial function is
disrupted during the carcinogenesis of diffuse type GC, inducing TL
change. Therefore, loss of the co-regulation of telomere length and
mitochondrial copy number may serve a pivotal initiating role in
gastric carcinogenesis. However, the survival analysis from the
present study did not observe that this affected the clinical
characteristics and further progression of GC.
Mitochondria and telomeres are considered to be key
instigators of natural ageing (18).
Mitochondrial dysfunction or genetic change during aging may be
associated with the aging process through increased ROS production
and decreased adenosine triphosphate generation (18). A previous study demonstrated that
mitochondrial biogenesis and energy production were decreased in
telomerase-deficient mice with severe telomere dysfunction
(17). It has been hypothesized that
telomere change influences not only oxidative defense mechanisms
but also mitochondrial functions, including biogenesis and
metabolism, in transcriptomic, molecular, genetic and functional
analyses of various cells and organs, including proliferative and
post-mitotic tissues (16,17). Therefore, this telomere-mitochondria
axis may explain how shortened telomeres can cause mitochondrial
change. The results of the present study support this hypothesis,
suggesting that deregulation of the telomere-mitochondria axis, as
caused by aging or other physiological factors, triggers the
carcinogenesis of diffuse type GC.
In summary, TL and mtCN are correlated in normal and
cancerous stomach tissues. Additionally, abnormal regulation of the
telomere-mitochondria axis was identified in diffuse type GC,
although the mechanism underlying this process remains unclear.
Therefore, a change in the regulation of mitochondria and telomeres
may be essential for diffuse GC carcinogenesis, suggesting that
this signaling pathway could be targeted for cancer prevention.
Furthermore, this result improves current understanding of how
telomere change may contribute not only to age-associated
disorders, but also to tumorigenesis.
Acknowledgements
The present study was supported by grants from the
Basic Science Research Program through the National Research
Foundation of Korea funded by the Ministry of Education
(NRF-2014R1A6A3A04058057) and by the Korean Government (grant no.
2014R1A5A2010008).
References
|
1
|
Jung KW, Won YJ, Oh CM, Kong HJ, Cho H,
Lee JK, Lee DH and Lee KH: Prediction of cancer incidence and
mortality in Korea, 2016. Cancer Res Treat. 48:451–457. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Correa P and Shiao YH: Phenotypic and
genotypic events in gastric carcinogenesis. Cancer Res. 54 7
Suppl:1941s–1943s. 1994.PubMed/NCBI
|
|
3
|
Grabsch HI and Tan P: Gastric cancer
pathology and underlying molecular mechanisms. Dig Surg.
30:150–158. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yakirevich E and Resnick MB: Pathology of
gastric cancer and its precursor lesions. Gastroenterol Clin North
Am. 42:261–284. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Akhavan-Niaki H and Samadani AA: Molecular
insight in gastric cancer induction: An overview of cancer stemness
genes. Cell Biochem Biophy. 68:463–473. 2014. View Article : Google Scholar
|
|
6
|
Carcas LP: Gastric cancer review. J
Carcinog. 13:142014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Harley CB, Futcher AB and Greider CW:
Telomeres shorten during ageing of human fibroblasts. Nature.
345:458–460. 1990. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Anderson S, Bankier AT, Barrell BG, De
Bruijn MH, Coulson AR, Drouin J, Eperon IC, Nierlich DP, Roe BA,
Sanger F, et al: Sequence and organization of the human
mitochondrial genome. Nature. 290:457–465. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Blackburn EH, Greider CW and Szostak JW:
Telomeres and telomerase: The path from maize, Tetrahymena and
yeast to human cancer and aging. Nat Med. 12:1133–1138. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Blasco MA: Telomeres and human disease:
Ageing, cancer and beyond. Nat Rev Genet. 6:611–622. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cui H, Huang P, Wang Z, Zhang Y, Zhang Z,
Xu W, Wang X, Han Y and Guo X: Association of decreased
mitochondrial DNA content with the progression of colorectal
cancer. BMC Cancer. 13:1102013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lee HC, Li SH, Lin JC, Wu CC, Yeh DC and
Wei YH: Somatic mutations in the D-loop and decrease in the copy
number of mitochondrial DNA in human hepatocellular carcinoma. Mut
Res. 547:71–78. 2004. View Article : Google Scholar
|
|
13
|
Liao LM, Baccarelli A, Shu XO, Gao YT, Ji
BT, Yang G, Li HL, Hoxha M, Dioni L, Rothman N, et al:
Mitochondrial DNA copy number and risk of gastric cancer: A report
from the Shanghai Women's Health Study. Cancer Epidemiol Biomarkers
Prev. 20:1944–1949. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wu X, Amos CI, Zhu Y, Zhao H, Grossman BH,
Shay JW, Luo S, Hong WK and Spitz MR: Telomere dysfunction: A
potential cancer predisposition factor. J Natl Cancer Inst.
95:1211–1218. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kim JH, Kim HK, Ko JH, Bang H and Lee DC:
The relationship between leukocyte mitochondrial DNA copy number
and telomere length in community-dwelling elderly women. PloS one.
8:e672272013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Qiu C, Enquobahrie DA, Gelaye B, Hevner K
and Williams MA: The association between leukocyte telomere length
and mitochondrial DNA copy number in pregnant women: A pilot study.
Clin Lab. 61:363–369. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tyrka AR, Carpenter LL, Kao HT, Porton B,
Philip NS, Ridout SJ, Ridout KK and Price LH: Association of
telomere length and mitochondrial DNA copy number in a community
sample of healthy adults. Exp Gerontol. 66:17–20. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Passos JF, Saretzki G and von Zglinicki T:
DNA damage in telomeres and mitochondria during cellular
senescence: Is there a connection? Nucleic Acids Res. 35:7505–7513.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hu J, Hwang SS, Liesa M, Gan B, Sahin E,
Jaskelioff M, Ding Z, Ying H, Boutin AT, Zhang H, et al:
Antitelomerase therapy provokes ALT and mitochondrial adaptive
mechanisms in cancer. Cell. 148:651–663. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sahin E, Colla S, Liesa M, Moslehi J,
Müller FL, Guo M, Cooper M, Kotton D, Fabian AJ, Walkey C, et al:
Telomere dysfunction induces metabolic and mitochondrial
compromise. Nature. 470:359–365. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lee H, Lee JH, Kim DC, Hwang I, Kang YN,
Gwon GJ, Choi IJ and Kim S: Is mitochondrial DNA copy number
associated with clinical characteristics and prognosis in gastric
cancer? Asian Pac J Cancer Prev. 16:87–90. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lee H, Jin JD, La BM, Park WJ, Choi IJ and
Lee JH: TERT promoter mutation, telomere length and TERT expression
in gastric cancer. Int J Clin Exp Pathol. 9:1758–1763. 2016.
|
|
23
|
Gil ME and Coetzer TL: Real-time
quantitative PCR of telomere length. Mol Biotechnol. 27:169–172.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lee JH, Hwang I, Kang YN, Choi IJ and Kim
DK: Genetic characteristics of mitochondrial DNA was associated
with colorectal carcinogenesis and its prognosis. PloS One.
10:e01186122015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tyrka AR, Parade SH, Price LH, Kao HT,
Porton B, Philip NS, Welch ES and Carpenter LL: Alterations of
mitochondrial DNA copy number and telomere length with early
adversity and psychopathology. Biol Psychiatry. 79:78–86. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Aslan R, Bektas A, Bedir A, Alacam H,
Aslan MS, Nar R, Yildirim B, Goren I, Ecemis O, Ustaoglu M, et al:
Helicobacter pylori eradication increases telomere length in
gastric mucosa. Hepatogastroenterology. 60:601–604. 2013.PubMed/NCBI
|
|
27
|
Pascua I, Fernández-Marcelo T,
Sánchez-Pernaute A, De Juan C, Head J, Torres-García AJ and Iniesta
P: Prognostic value of telomere function in gastric cancers with
and without microsatellite instability. Eur J Gastroenterol
Hepatol. 27:162–169. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tahara T, Shibata T, Kawamura T, Ishizuka
T, Okubo M, Nagasaka M, Nakagawa Y, Arisawa T, Ohmiya N and Hirata
I: Telomere length in non-neoplastic gastric mucosa and its
relationship to H. pylori infection, degree of gastritis and NSAID
use. Clin Exp Med. 16:65–71. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tahara T, Shibata T, Okubo M, Kawamura T,
Horiguchi N, Ishizuka T, Nakani N, Nagasaka M, Nakagawa Y and
Ohmiya N: Demonstration of potential link between helicobacter
pylori related promoter CpG island methylation and telomere
shortening in human gastric mucosa. Oncotarget. 7:43989–43996.
2016.PubMed/NCBI
|
|
30
|
Jeong CW, Lee JH, Sohn SS, Ryu SW and Kim
DK: Mitochondrial microsatellite instability in gastric cancer and
gastric epithelial dysplasia as a precancerous lesion. Cancer
Epidemiol. 34:323–327. 2010. View Article : Google Scholar : PubMed/NCBI
|