Introduction
Flavonoids are the most abundant polyphenols in the
human diet and may be subdivided into anthocyanidins, flavanols,
flavanones, flavonols, flavones and isoflavones, according to
structural and functional features (1). In a previous review, dietary flavonoids
were highlighted for their beneficial biological activities in the
improvement of human health against osteoporosis, cancer,
cardiovascular diseases, neurodegenerative diseases and diabetes in
a variety of epidemiological studies (2). As a subfamily of flavonoids,
anthocyanins are important plant pigments with a red, blue or
purple color; they are principally identified in berries,
pomegranates, red cabbage, grapes, sweet potatoes and red wine
(3). Although >400 anthocyanins
have been identified in nature, only six anthocyanidins, including
cyanidin, pelargonidin, peonidin, delphinidin, petunidin and
malvidin, are ubiquitously distributed (4). Amongst anthocyanidins, delphinidin is
known to have effective biological functions in the prevention of
oxidative stress, inflammation, angiogenesis, metastasis and
carcinogenesis (5–7). An increasing number of studies have
identified the potential protective effects of delphinidin against
carcinogenesis of the breast (8),
prostate (9), lungs (10), liver (11) and colon (12) and against fibrosarcoma (13) by regulating cell signal transduction.
However, little is known about the functional role of delphinidin
in the female reproductive system and gynecologic types of cancer,
including of the cervix, ovary, endometrium, vagina and vulva
(14,15).
Gynecological cancer subtypes are malignant
carcinomas of the female reproductive tract, the most common of
which is epithelial ovarian cancer (EOC); EOC is the fifth leading
cause of cancer-associated mortalities in women, due to a lack of
symptoms and effective biomarkers for diagnosis in the early stages
(16). The current first-line
treatment strategy for EOC is intensive surgical cytoreduction,
followed by systemic chemotherapy with a platinum- or taxane-based
regimen (17). However, advanced EOC
[International Federation of Gynecological Oncology stages III–IV
(18)] accounts for ~75% of total
patients diagnosed with ovarian cancer with low (5-year) survival
rates and a high recurrence rate of ~80% with failure to respond to
conventional chemotherapy (19).
Thus, it is necessary to develop novel chemotherapeutic approaches
to increase the susceptibility of EOC to novel treatments, in order
to improve the median progression-free survival and overall
survival times. Therefore, the present study determined the
potential role of delphinidin in EOC through investigating the
following: Effects of delphinidin on the viability of SKOV3 ovarian
adenocarcinoma cells; cell signaling mechanisms regulated by
delphinidin against the proliferation of SKOV3 cells; comparative
chemotherapeutic effects between delphinidin and conventional
chemotherapy drugs on the proliferation of SKOV3 cells. The results
of the present study indicate that delphinidin exerts
anti-proliferative effects in SKOV3 cells via inactivation of the
phosphoinositide-3 kinase (PI3K) and extracellular-regulated kinase
(ERK)1/2 mitogen-activated protein kinase (MAPK) signaling
pathways. Therefore, this may be an effective synergistic regimen
for the enhancement of conventional chemotherapy for patients with
EOC.
Materials and methods
Chemicals
Delphinidin was purchased from Indofine Chemical
Company, Inc. (Hillsborough, NJ, USA). SB203580 and U0126 were
purchased from Enzo Life Sciences, Inc. (Farmingdale, NY, USA) and
LY294002 was obtained from Cell Signaling Technology, Inc.
(Danvers, MA, USA). The antibodies against phosphorylated-AKT
[serine (Ser)473] (catalog no. 4060), ERK1/2 [threonine
(Thr)202/Tyr204, (catalog no. 9101; Cell
Signaling Technology, Inc.)] c-Jun N-terminal kinase (JNK;
Thr183/Thr185, catalog no. 4668), p38
(Thr180/Thr182; catalog no. 4511), ribosomal
protein S6 kinase β-1 (P70S6K; Thr421/Ser424;
catalog no. 9204) and S6 (Ser235/236, catalog no. 2211),
and total AKT (catalog no. 9272), ERK1/2 (catalog no.4695), JNK
(catalog no. 9252), p38 (catalog no. 9212), P70S6K (catalog no.
9202) and S6 (catalog no. 2217) were purchased from Cell Signaling
Technology, Inc. All antibodies were diluted to 1:1,000 for western
blot analysis. Cisplatin and paclitaxel were purchased from
Sigma-Aldrich (Merck KGaA, Darmstadt, Germany).
Cell culture
SKOV3 cells were purchased from the American Type
Culture Collection (Manassas, VA, USA) and maintained in McCoy's 5A
(modified) medium (cat. no. 16600-082; Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) supplemented with 10% fetal
bovine serum (Hyclone; GE Healthcare Life Sciences, Logan, UT, USA)
at 37°C in an atmosphere containing CO2.
Proliferation assay
Proliferation assays were conducted using a Cell
Proliferation ELISA, 5-bromo-2′-deoxyuridine (BrdU) kit (catalog
no. 11647229001; Roche Diagnostics, Indianapolis, IN, USA),
according to the manufacturer's protocol. Briefly, SKOV3 cells
(3×103 cells/100 µl) were seeded onto a 96-well plate
and incubated at 37°C for 24 h in serum-free McCoy's 5A (modified)
medium. Cells were then treated with 10 µM delphinidin alone or
with various inhibitors of cell signaling proteins including 20 µM
LY294002, 20 µM SB203580 and 10 µM U0126 or conventional
chemotherapy including 20 µM paclitaxel and 50 µM cisplatin in a
final volume of 100 µl/well. Following 48 h of incubation at 37°C,
10 µM BrdU was added to the cell culture and the cells were
incubated for an additional 2 h at 37°C. After labeling cells with
BrdU, the fixed cells were incubated with anti-BrdU-peroxidase
(POD) working solution for 90 min at room temperature. The
anti-BrdU-POD binds to BrdU incorporated in newly synthesized
cellular DNA, and these immune complexes were detected by the
reaction to 3,3′,5,5′-tetramethylbenzidine substrate. The
absorbance values of the reaction product were quantified by
measuring the absorbance at 370 and 492 nm using an ELISA reader.
These experiments were performed in triplicate.
TUNEL assay
The SKOV3 cells (3×104 cells/300 µl) were
seeded onto confocal dishes (catalog no. 100350; SPL Life Sciences
Co., Ltd., Pocheon, Korea) and then incubated for 24 h at 37°C in
serum-free McCoy's 5A. The cells were subsequently treated with
delphinidin at a final concentration of 10 µM in 10 ml for 48 h at
37°C in a CO2 incubator. Following incubation, the cells were
subjected to TUNEL staining using an in situ Cell Death
Detection kit, TMR red (Roche Diagnostics) according to the
manufacturer's protocol. Fluorescence was detected using an LSM 710
(Carl Zeiss AG, Oberkochen, Germany) confocal microscope at
magnifications 40x and 80x fitted with a digital microscope AxioCam
camera and using Zen 2009 software (Carl Zeiss AG). These
experiments were performed in triplicate.
Western blot analysis
The concentration of proteins from whole-cell
extracts was determined using a Bradford protein assay (Bio-Rad
Laboratories, Inc., Hercules, CA, USA) with bovine serum albumin
(Sigma-Aldrich; Merck KGaA) as the standard. Proteins were
denatured, separated using 10% SDS-PAGE and transferred to
nitrocellulose membranes. The membranes were blocked using 5%
bovine serum albumin in TBST buffer for 1 h at room temperature.
Immunoreactive proteins were detected using goat anti-rabbit
polyclonal antibodies against phosphorylated proteins and total
proteins at a dilution of 1:1,000 (as mentioned in the previous
section). The blots were developed using enhanced chemiluminescence
detection (SuperSignal™ West Pico; Pierce; Thermo Fisher
Scientific, Inc.) and quantified by measuring the intensity of
light emitted from correctly sized bands under ultraviolet light,
using a ChemiDoc™ EQ system and Quantity One® software
(version 4.5; Bio-Rad Laboratories, Inc.). Total protein
normalization was used to quantify the abundance of the
phosopho-proteins. As a loading control, total proteins were used
to normalize the results from the detection of proteins by western
blotting. Multiple exposures of each western blot were performed to
ensure linearity of the chemiluminescent signals. These experiments
were performed in triplicate.
Determination of apoptosis by Annexin
V and propidium iodide (PI) staining
Apoptosis of SKOV3 cells induced by delphinidin was
analyzed using a fluorescein isothiocyanate (FITC)-Annexin V
apoptosis detection kit I (BD Biosciences, Franklin Lakes, NJ,
USA). The cells (4×105 cells) were seeded onto 6-well
plates and incubated for 24 h at 37°C in a CO2 incubator in
serum-free medium until the cells reached 70–80% confluence. The
cells were then treated separately with 10 µM delphinidin, 20 µM
LY294002, 10 µM U0126 or a combination of 10 µM delphinidin with
each inhibitor for 48 h at 37°C in a CO2 incubator. Supernatants
were removed from the culture dishes using a 10 ml serological
pipette, and adherent cells were detached using 0.25% trypsin-EDTA.
The cells were collected by centrifugation (300 × g) at room
temperature for 5 min, washed with PBS and resuspended in 1X
binding buffer (BD Biosciences). The cell suspension (100 µl) was
then transferred into a 5 ml culture tube and incubated with 5 µl
FITC-Annexin V and 5 µl PI for 15 min at room temperature in the
dark. Subsequently, 400 µl 1X binding buffer was added to the 5 ml
culture tube. Fluorescence intensity was analyzed using
FACSCalibur™ at 488 nm (BD Biosciences).
Cell cycle assay
SKOV3 cells (3×105 cells) were seeded
onto 6-well plates and incubated for 24 h in serum-free medium at
37°C until the cells reached 70–80% confluence. The cells were then
treated separately with 10 µM delphinidin, 20 µM LY294002, 10 µM
U0126 or a combination of 10 µM delphinidin with each inhibitor for
48 h at 37°C in an incubator containing CO2. Supernatants were
transferred from the culture dishes into collecting tubes and
adherent cells were detached using 0.25% trypsin-EDTA. The cells
were collected via centrifugation (300 × g) at room temperature for
5 min, washed twice with cold PBS and fixed in chilled-70% ethanol
at 4°C overnight. Cells were subsequently washed twice with PBS,
incubated with 10 µg/ml RNase A (Sigma-Aldrich; Merck KGaA) and 50
µg/ml PI (BD Biosciences) in PBS at room temperature for 30 min in
the dark and subjected to flow cytometry (FACSCalibur™; BD
Biosciences).
Statistical analysis
Data obtained from the proliferation and Annexin V
and PI assays were subjected to two-way analysis of variance
according to the general linear model (PROC-GLM) of the SAS program
(SAS Institute, Cary, NC, USA) in order to detect the effects of
treatments on SKOV3 cells proliferation. Data are presented as
least square means with standard error of the mean, unless
otherwise stated. P<0.05 was considered to indicate a
statistically significant difference.
Results
Inhibitory effects of delphinidin on
the viability of SKOV3 cells
The current study aimed to determine the effects of
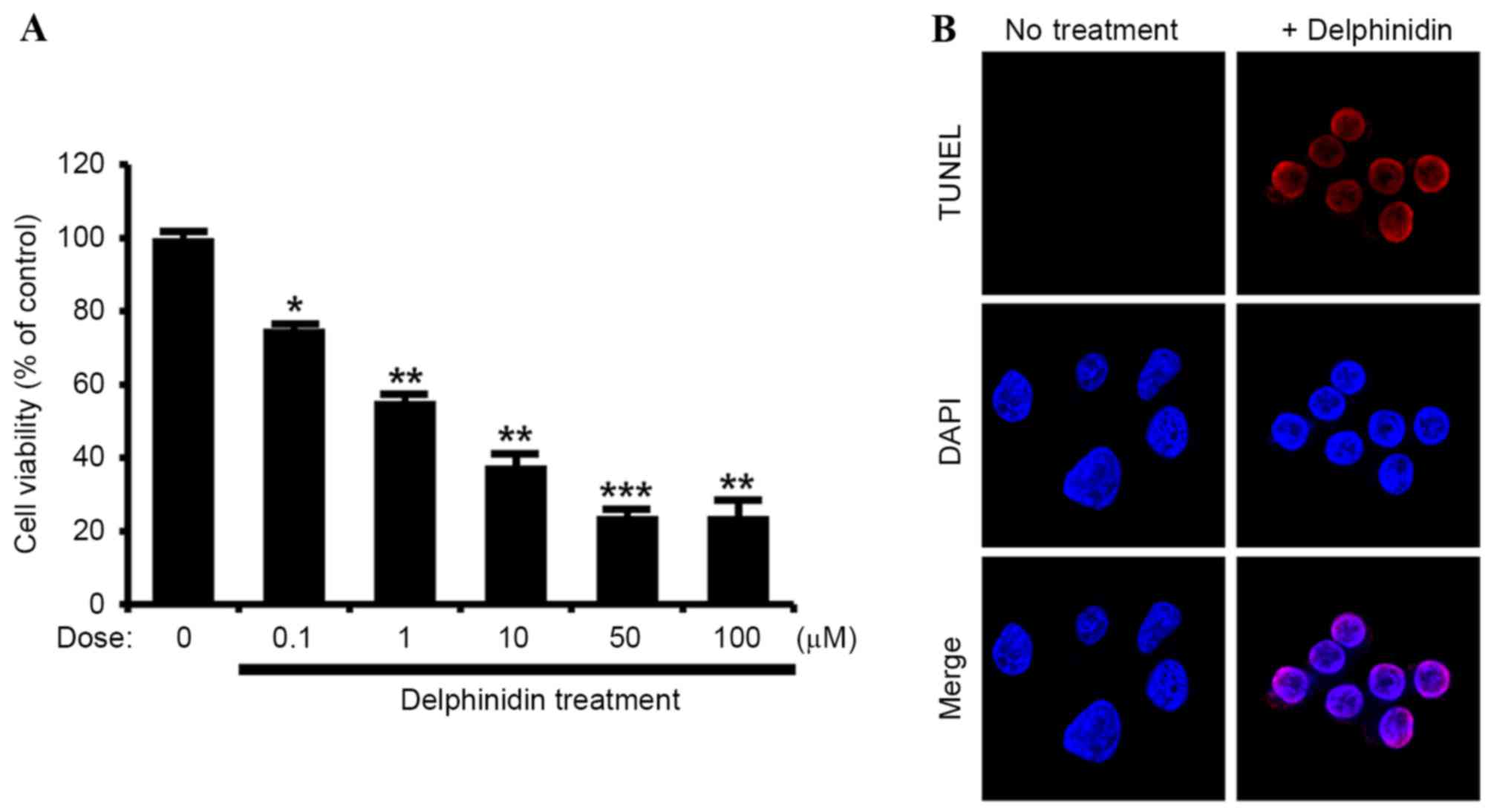
delphinidin on SKOV3 cells derived from ovarian adenocarcinoma. The
results demonstrated a dose-dependent effect of delphinidin to
inhibit the proliferation of SKOV3 cells between concentrations of
0, 0.1, 1, 10, 50 and 100 µM (Fig.
1A). Based on these results, delphinidin was used at a
concentration of 10 µM in further experiments, in order to
determine its effects on the apoptosis of SKOV3 cells. As indicated
by the TUNEL assay, apoptosis was induced in SKOV3 cells by nuclear
DNA fragmentation in response to delphinidin treatment (Fig. 1B). In addition, the dose-dependent
effects of delphinidin on the PI3K/AKT and MAPK signaling pathways
were investigated (Fig. 2). Compared
with basal values, delphinidin decreased the phosphorylation of
PI3K downstream proteins, including AKT, P70S6K and S6, in a
dose-dependent manner. In addition, the phosphorylated MAPK
signaling proteins, including ERK1/2 MAPK and P38 MAPK, were
downregulated with increasing concentrations of delphinidin,
whereas JNK phosphorylation was not affected in SKOV3 cells treated
with delphinidin. These results indicate that delphinidin exhibits
anti-proliferative effects on SKOV3 cells and inhibits
phosphorylation of the PI3K/AKT, ERK1/2 MAPK and P38 MAPK cell
signaling proteins.
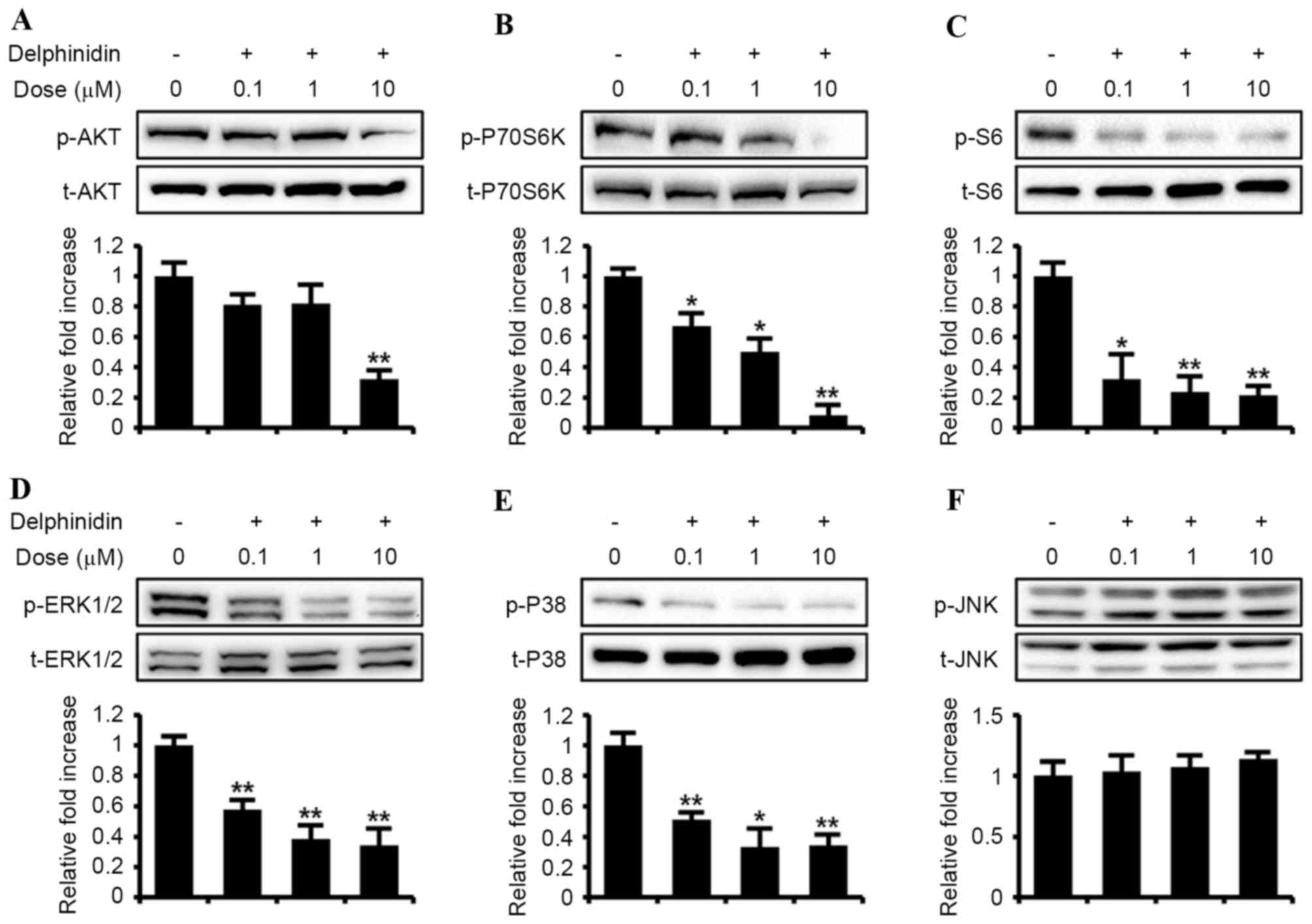 | Figure 2.Dose-response effects of delphinidin
on the phosphorylation of target signaling proteins in SKOV3 cells.
Serum-starved SKOV3 cells were treated with delphinidin (0, 0.1, 1
and 10 µM) for 30 min. Phosphorylation of (A) AKT, (B) P70S6K, (C)
S6, (D) ERK1/2, (E) p38 and (F) JNK was estimated in SKOV3 cells
incubated with delphinidin using western blot analysis. Blotting
membranes were imaged to analyze the normalized values by
calculating the abundance of phosphorylated protein relative to the
total protein. **P<0.01; *P<0.05. ERK,
extracellular-regulated kinase; JNK, c-Jun N-terminal kinase; p,
phosphorylated; t, total protein; AKT, protein kinase B. |
Effects of the inhibition of PI3K/AKT
and MAPK signal transduction on SKOV3 cell proliferation
It was examined whether the delphinidin-suppressed
proliferation of SKOV3 cells is associated with the PI3K/AKT,
ERK1/2 MAPK and P38 MAPK signal transduction pathways. Cell
proliferation assays were conducted in the absence or presence of
delphinidin (10 µM), LY294002 (20 µM; PI3K inhibitor), SB203580 (20
µM; P38 MAPK inhibitor) and U0126 (10 µM; ERK1/2 MAPK inhibitor;
Fig. 3A). Compared with non-treated
SKOV3 cells, delphinidin inhibited the proliferation of SKOV3 cells
by ~40% (P<0.01). Treatment of SKOV3 cells with LY294002 or
U0126 alone decreased cell proliferation by ~30% (P<0.05) and
70% (P<0.001), respectively. In addition, the co-incubation of
LY294002 (P<0.05) and U0126 (P<0.001) with delphinidin
exhibited a synergistic effect to inhibit the proliferation of
SKOV3 cells, as compared with delphinidin or pharmacological
inhibitor alone. However, no significant effect of SB203580 on the
anti-proliferative effects of delphinidin on SKOV3 cells was
observed, whereas SB203580 alone slightly reduced the viability of
SKOV3 cells. Thus, the expression of target signaling proteins in
response to the blockage of PI3K/AKT and ERK1/2 MAPK transduction
pathways was investigated using LY294002 or U0126 in order to
identify those cell signaling pathways mediated by delphinidin. As
illustrated in Fig. 3B, the
suppression of AKT in SKOV3 cells was markedly inhibited by
combined treatment with LY294002 and delphinidin, and with U0126
and delphinidin, respectively. The levels of phosphorylated ERK1/2
proteins in SKOV3 cells were significantly reduced by administering
delphinidin alone (P<0.05; Fig.
3C). In addition, treating cells with a combination of
delphinidin with U0126 highly suppressed the abundance of
phosphorylated ERK1/2 protein in SKOV3 cells compared with cells
treated with delphinidin alone. By contrast, treatment with
delphinidin and LY294002 did not result in a significant decrease
in the levels of phosphorylated ERK1/2 compared with treatment with
dephinidin alone (Fig. 3C). The
phosphorylation of P70S6K was also inhibited (P<0.05) in SKOV3
cells when treated with a combination of LY294002 and delphinidin,
but not in cells treated with a combination of U0126 and
delphinidin (Fig. 3D). Additionally,
pattern of changes in the level of phospho-proteins in response to
each treatment in SKOV3 cells were similar for phospho-S6 and
phospho-AKT (Fig. 3E). These results
indicate that the inactivation of PI3K/AKT and EKR1/2 MAPK signal
transduction serves a crucial role in delphinidin-induced
suppression of SKOV3 cell proliferation.
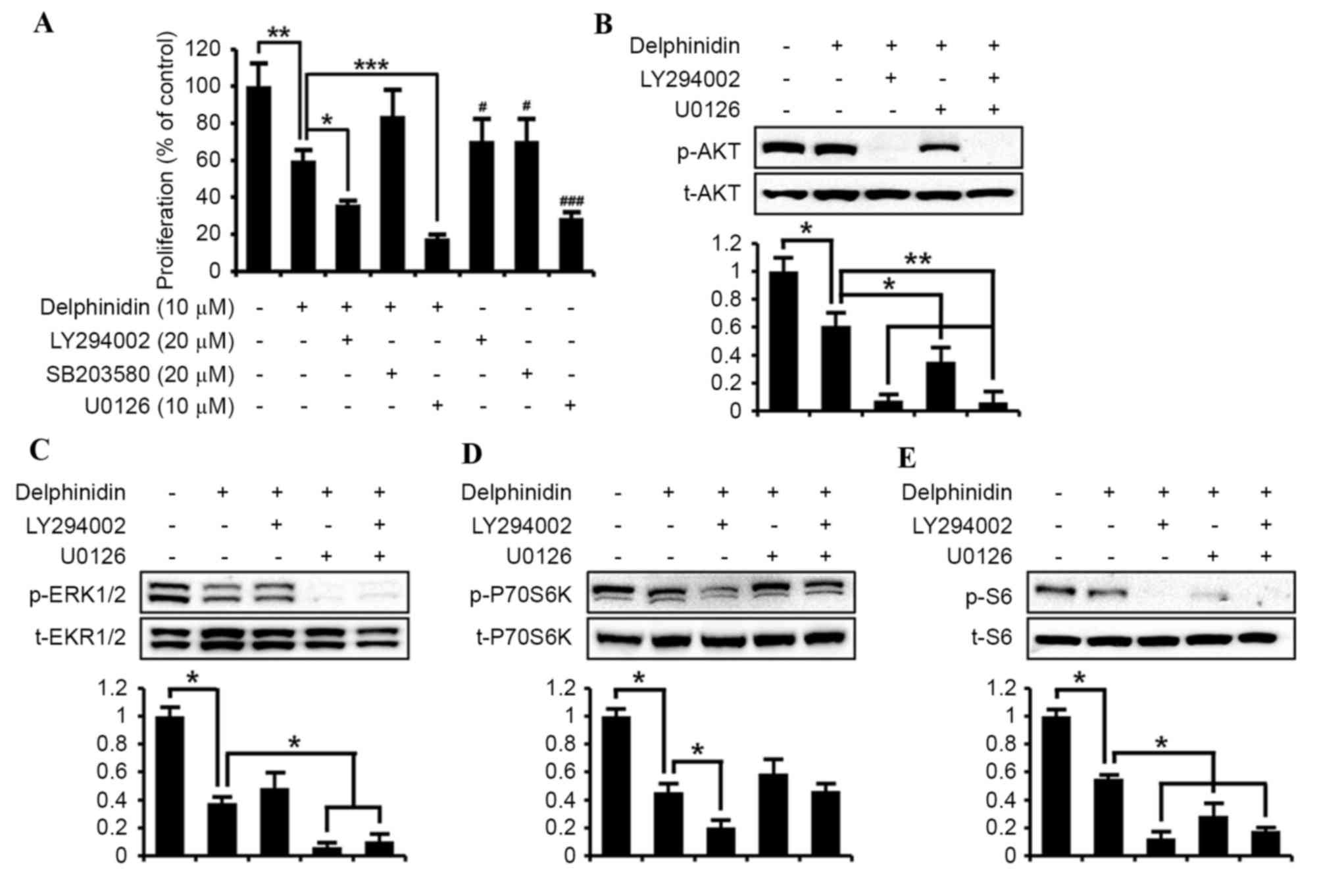 | Figure 3.Delphinidin suppresses the
proliferation of SKOV3 cells through the PI3K/AKT and ERK1/2 MAPK
signaling pathways. (A) Effects of pharmacological inhibitors,
including 20 µM LY294002 (a PI3K inhibitor), 20 µM SB203580 (a P38
MAPK inhibitor) or 10 µM U0126 (an ERK1/2 MAPK inhibitor), of cell
signaling pathways on the proliferation of SKOV3 cells treated with
delphinidin. The results exhibit relative differences in cell
proliferation, as compared with non-treated SKOV3 cells set at
100%. Starved SKOV3 cells were pretreated with 20 µM LY294002 or 10
µM U0126, or with a combination of both inhibitors for 1 h and then
incubated with 10 µM delphinidin for 30 min. Phosphorylation of (B)
AKT, (C) ERK1/2, (D) P70S6K and (E) S6 was determined in SKOV3
cells by western blot analysis. Blotting membranes were imaged to
analyze the normalized values by calculation of the abundance of
phosphorylated protein relative to total protein. All quantitative
data are presented as least square means with overall standard
error of the mean. ***P<0.001; **P<0.01; *P<0.05, compared
with delphinidin treatment.. PI3K, phosphoinositide-3 kinase; ERK,
extracellular-regulated kinase; MAPK, mitogen-activated protein
kinase; p, phosphorylated; t, total protein; AKT, protein kinase
B. |
Effects of PI3K/AKT and MAPK signal
transduction inhibition on the necrosis and apoptosis of SKOV3
cells
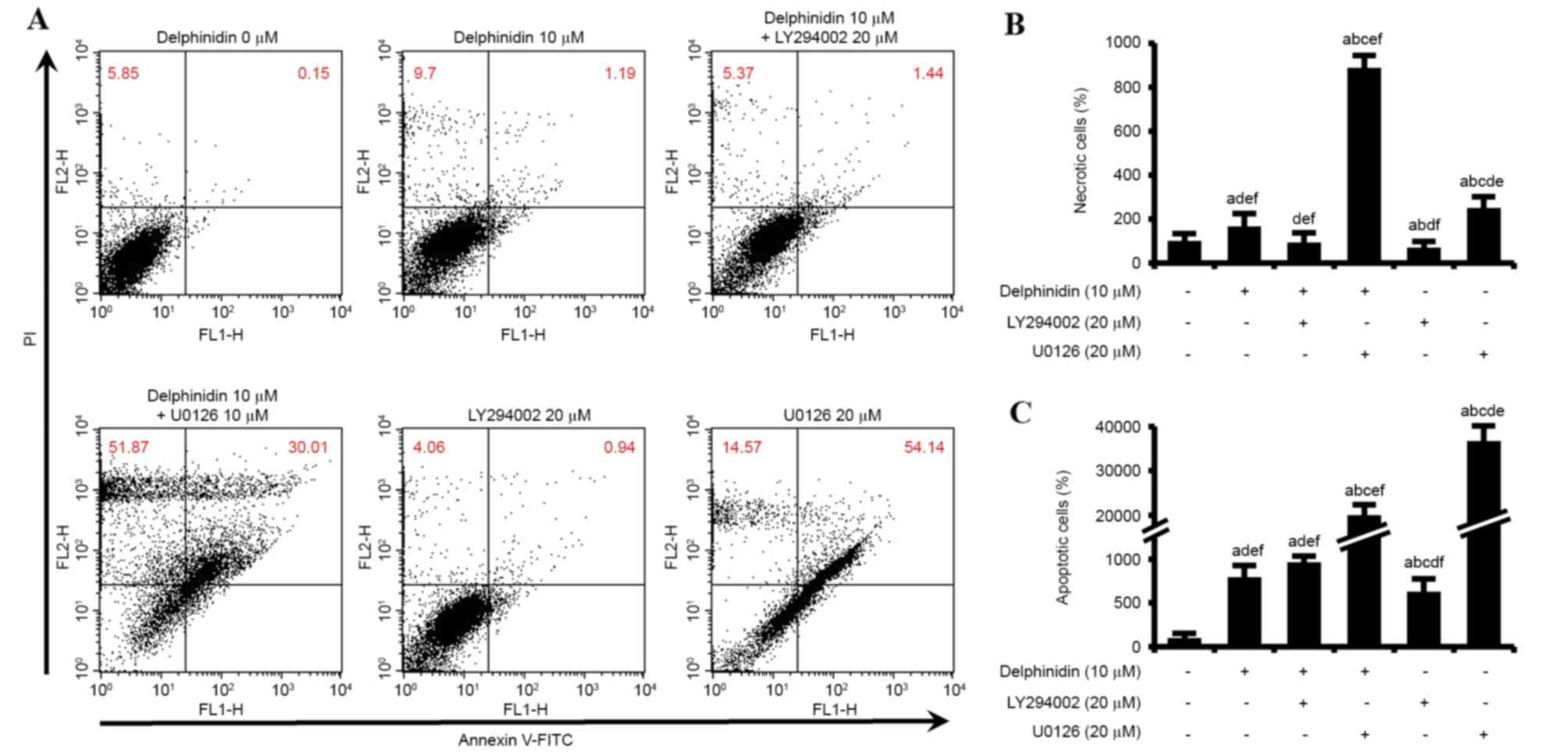
In order to compare the apoptotic effects of various
drug treatments (10 µM delphinidin, 20 µM LY294002, 10 µM U0126 and
a combination of 10 µM delphinidin with each inhibitor) in SKOV3
cells, Annexin V/PI staining and cell cycle assays were performed.
As illustrated in Fig. 4A, the number
of necrotic (upper left quadrant) and apoptotic (upper right
quadrant) SKOV3 cells in response to various drug treatments were
investigated by Annexin V-FITC and PI flow cytometry. The number of
necrotic SKOV3 cells increased by 165%, as compared with the number
of cells in the control cell group (Fig.
4B). In addition, a combination of delphinidin and U0126
increased the abundance of necrotic SKOV3 cells by ~886%, compared
with non-treated SKOV3 cells. This necrotic effect on SKOV3 cells
was greater than that observed for delphinidin or U0126 treatment
alone. However, delphinidin in combination with LY294002 only
slightly enhanced the necrosis of SKOV3 cells, as compared with
LY294002 administered alone. Delphinidin significantly increased
the rate of apoptosis in SKOV3 cells (793%, P<0.05), compared
with non-treated SKOV3 cells (Fig.
4C). In addition, the combination of delphinidin and LY294002
increased the number of apoptotic cells to a greater extent than
that observed with delphinidin or LY294002 alone. Furthermore, the
apoptosis of SKOV3 cells was significantly increased by treatment
with U0126. These results indicate that the inhibition of PI3K/AKT
and ERK1/2 MAPK signaling pathways in SKOV3 cells treated with
delphinidin induces necrosis and apoptosis.
Comparison of cell cycle regulation in
SKOV3 cells by delphinidin, inhibitors of PI3K and ERK1/2 MAPK and
a combination
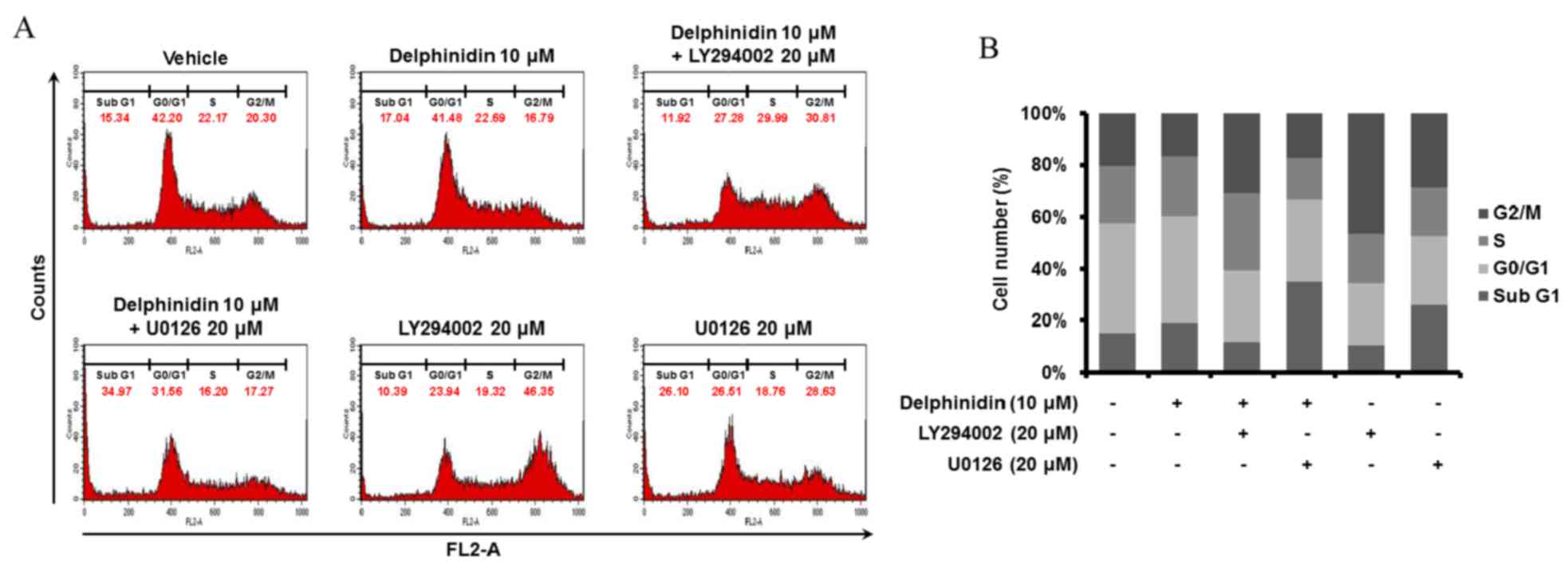
To determine the regulatory effects of delphinidin
and pharmacological inhibitors on the cell cycle distribution of
SKOV3 cells, cell cycle assays were conducted via quantitation of
DNA content using PI and flow cytometry (Fig. 5). Delphinidin increased the number of
cells in the sub-G1 phase, whereas the proportions of SKOV3 cells
in the G0/G1 and G2/M phases were decreased following treatment
with delphinidin due to cell cycle arrest, compared with
non-treated cells. In addition, the treatment of SKOV3 cells with
an ERK1/2 MAPK inhibitor (U0126) increased the proportion of cells
in the sub-G1 phase compared with non-treated cells. There was a
greater increase in the number of SKOV3 cells in the sub-G1 phase
when treated with a combination of delphinidin and U0126 compared
with treating with delphinidin or U0126 alone. In addition,
LY294002 administered alone and in combination with delphinidin
increased the proportion of SKOV3 cells in the S and G2/M phases,
as compared with SKOV3 cells treated with delphinidin alone. These
results indicate that delphinidin stimulates the apoptosis of SKOV3
cells by inducing cell cycle arrest and that a combination of
delphinidin and U0126 primarily induces sub-G1 phase arrest in
SKOV3 cells.
Chemotherapeutic effects of
delphinidin compared with conventional chemotherapy regimens
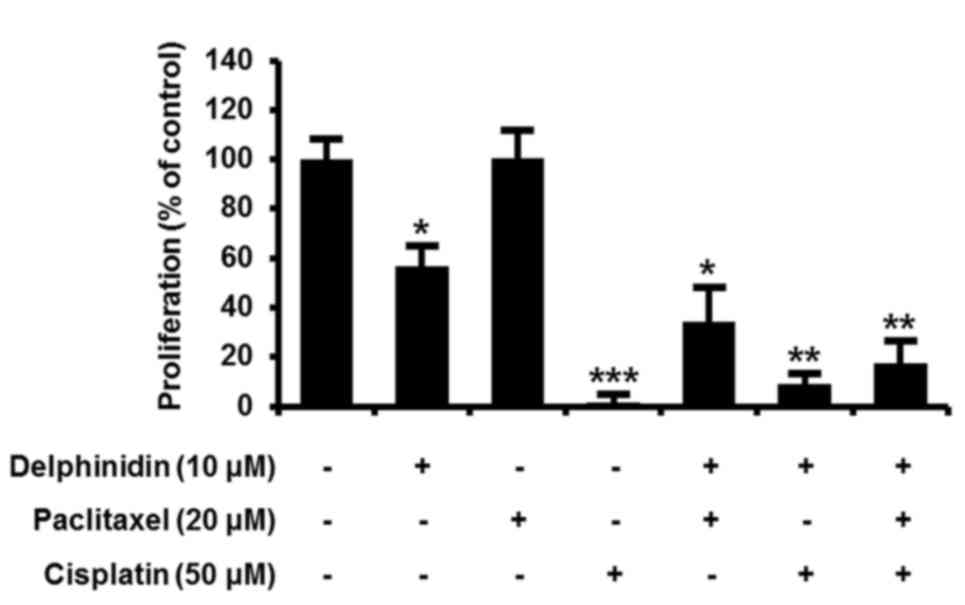
To investigate chemotherapeutic effects of
delphinidin on the proliferation of SKOV3 cells compared with
conventional chemotherapy, the therapeutic efficacy of 10 µM
delphinidin, 20 µM paclitaxel (a taxane-based drug) and 50 µM
cisplatin (a platinum-based drug) alone and in combination were
assessed (Fig. 6). Monotherapy using
delphinidin or cisplatin inhibited the proliferation of SKOV3 cells
by 44% (P<0.05) and 95% (P<0.001), respectively. By contrast,
the SKOV3 cells were not affected by paclitaxel treatment alone. In
comparison with paclitaxel monotherapy, the proliferation of SKOV3
cells was markedly decreased by delphinidin plus paclitaxel. These
results indicate that co-treatment with delphinidin and paclitaxel
effectively increases the chemotherapeutic activity of paclitaxel
monotherapy in SKOV3 cells.
Discussion
The results of the present study indicate a
functional role for delphinidin, a member of the anthocyanidin
family, in suppressing the progression of EOC using SKOV3 cells as
a model system. The anti-proliferative effects of delphinidin in
SKOV3 cells were regulated by its inhibition of PI3K/AKT and ERK1/2
MAPK signal transduction protein phosphorylation. In addition, a
combination of delphinidin and the pharmacological inhibitors of
PI3K and ERK1/2 MAPK induced apoptosis and cell cycle arrest in
SKOV3 cells. Furthermore, comparative analyses between delphinidin
and conventional chemotherapy of EOC revealed that delphinidin
exhibits a synergistic effect with paclitaxel in promoting
anti-proliferative effects in SKOV3 cells. These findings support
the hypothesis that delphinidin inhibits cell proliferation and
induces apoptosis in EOC, and that appropriate therapeutic
approaches using delphinidin may be useful for overcoming
chemoresistance in ovarian cancer cells.
Delphinidin, a natural bioactive product belonging
to the anthocyanidins, has a diphenylpropane-based polyphenolic
ring structure, including a positive charge in its core ring
(21). The clinical interest in
delphinidin is increasing due to its potential in preventing aging,
cardiovascular diseases, neurodegenerative diseases, diabetes and
cancer through various antioxidative (7), anti-inflammatory (22), anti-angiogenic (23) and anti-mutagenic (24) properties. In particular, the
antiproliferative effects of delphinidin have been identified in
various types of cancer cells to involve the regulation of
intracellular signaling pathways. In hepatocellular carcinoma,
delphinidin induces apoptotic cell death through the induction of
B-cell lymphoma-2 (Bcl-2) associated X protein and the reduction of
Bcl-2 (25). Delphinidin also
sensitizes TNF-related apoptosis-inducing ligand-resistant prostate
cancer cells to undergo apoptosis by inducing death receptor 5
mediating histone deacetylase 3 cleavage (9). In addition, it suppresses the
progression of lung cancer by inhibiting the aberrant expression of
endothelial growth factor receptor and vascular endothelial growth
factor receptor 2, leading to the decreased phosphorylation of PI3K
and MAPK (10). In addition,
delphinidin blocks the phosphorylation of ERK1/2 MAPK in breast
cancer cells (8). However, little is
known about the function and underlying mechanisms of delphinidin
in EOC. The present study demonstrated that the antiproliferative
effects of delphinidin on SKOV3 cells are dose-dependent (Fig. 1A). In addition, delphinidin increased
the percentage of apoptotic SKOV3 cells. These results indicate
that delphinidin inhibits the proliferation and induces the
apoptosis of SKOV3 cells.
EOC initiation and progression are complex events
regulated by various intracellular signaling proteins (26). For a novel therapeutic strategy to
incorporate delphinidin as an anticancer agent, understanding of
the molecular mechanisms underlying the process of carcinogenesis
from ovarian surface epithelia is essential. Activation of the
PI3K/AKT pathway is considered a key signal transduction pathway
for preventing tumorigenesis through promoting cell survival and
proliferation, and blocking apoptosis via the activation of
pro-apoptotic proteins and the control of cell cycle regulatory
proteins (27,28). Activated AKT may stimulate
phosphorylation of mechanistic target of rapamycin (mTOR) or
directly phosphorylate p70 S6 kinase (P70S6K), regulating
translation through ribosomal protein S6 (S6) for cell
proliferation and survival (29). The
activation of P70S6K in cell proliferation, invasiveness and
angiogenesis in malignant ovarian tumors has been reported and is
recognized as a hallmark of ovarian cancer (30,31). The
aberrant alteration of the PI3K-AKT-mTOR signaling pathways
indicates that the control of target proteins in the PI3K-AKT-mTOR
signal transduction pathway may be regarded as an effective
therapeutic approach for the treatment of EOC (32). Consistent with the results of previous
studies, the data obtained in the current study indicate that
increasing concentrations of delphinidin were able to decrease the
activity of AKT in SKOV3 cells. In addition, the phosphorylation of
P70S6K and S6 was inactivated following the treatment of SKOV3
cells with delphinidin. Delphinidin administered in combination
with LY294002 suppressed the activity of phosphorylated AKT and S6
proteins, as compared with delphinidin monotherapy in SKOV3 cells,
and enhanced the antiproliferative effects of delphinidin in SKOV3
cells. These results indicate that delphinidin exerts
chemopreventive effects on EOC cells through the inhibition of
PI3K/AKT signaling pathways.
Constitutive activation of the ERK1/2 MAPK pathway
promotes the growth, proliferation and differentiation of ovarian
cancer cells via the activation of downstream protein kinase or
transcription factors that may enhance carcinogenesis (33,34).
ERK1/2 MAPK signaling is activated by mitogenic factors and
inhibition of this pathway is crucial for the treatment of EOC
(34,35). In the present study, delphinidin
downregulated the phosphorylation of ERK1/2 MAPK in SKOV3 cells in
a dose-dependent manner. UO126, a pharmacological inhibitor of
ERK1/2, completely blocked the phosphorylation of ERK1/2 in SKOV3
cells, whereas LY294002 did not inhibit the phosphorylation of
ERK1/2. In addition, the combination of U0126 and delphinidin
enhanced the antiproliferative effects on SKOV3 cells, as compared
with delphinidin or U0126 alone. This effective blockade of signal
transduction using an ERK1/2 MAPK inhibitor and delphinidin may
provide insight into potential strategies for preventing
carcinogenesis from the ovarian surface epithelium.
For identifying interactions between delphinidin and
the pharmacological inhibitors of PI3K and ERK1/2 MAPK signaling,
apoptosis and cell cycle regulation in response to a combination of
delphinidin with LY294002 or U0126 was investigated. As
demonstrated in previous studies, delphinidin induces cell cycle
arrest via the downregulation of certain cell cycle regulatory
proteins, including cyclin-dependent kinase (Cdk)1, Cdk2, cyclin A
and cyclin D1, and via the upregulation of p21WAF1 in a
dose-dependent manner in prostate and colon cancer cells (12,36). In
addition, PI3K/AKT and ERK1/2-mediated suppression of cell
proliferation is due to cell cycle arrest in various types of
cancer cells (37–41). Concordantly, the results of the
present study revealed that the combination of delphinidin and
U0126 increased the percentage of necrotic cells, as compared with
the effects of delphinidin or U0126 alone. In addition, a
combination of delphinidin with LY294002 or U0126 synergistically
increased the rate of apoptosis in SKOV3 cells, as compared with
the effects of delphinidin alone. According to the cell cycle
analysis, the apoptotic effects were regulated through cell cycle
arrest at the sub-G1 phase. The cooperative effects of
delphinidin and the inhibitors of PI3K/AKT and ERK1/2 MAPK induced
the apoptosis of SKOV3 cells via regulation of the cell cycle.
Despite initial positive responses to platinum- and
taxane-based chemotherapies, persistent chemoresistance of EOC is
consistently a problem in ≤80% of patients (17). Specifically, the continued exposure of
SKOV3 cells to paclitaxel results in chemoresistance with marked
morphological changes (42,43). Therefore, novel therapeutic strategies
for the treatment of EOC are required for improvement of
progression-free survival and overall survival in patients. The
results of the present study indicated that the treatment of SKOV3
cells with paclitaxel for 48 h did not affect the viability of
SKOV3 cells, whereas cisplatin significantly decreased the
proliferation of SKOV3 cells. However, delphinidin exhibited
synergistic anticancer properties by reducing the proliferation of
paclitaxel-resistant SKOV3 cells. Therefore, combination therapy
with delphinidin and paclitaxel represents a novel treatment
strategy for additional clinical studies into advanced stages of
EOC.
Collectively, the present study presents evidence,
using the SKOV3 human ovarian cancer cell line, that delphinidin is
a crucial physiological modulator in EOC, and is also an inhibitor
of EOC cell proliferation. It was identified that delphinidin
increased the rate of apoptosis via DNA fragmentation in SKOV3
cells. In addition, delphinidin suppressed the proliferation of
SKOV3 cells through inactivating the phosphorylation of AKT,
P70S6K, S6 and ERK1/2 proteins in the PI3K/AKT and ERK1/2 MAPK
signaling pathways. Furthermore, blocking the ERK1/2 MAPK signaling
pathways reduced the levels of S6 protein in SKOV3 cells, which
regulates protein synthesis for cell death. Furthermore,
interactions between delphinidin and various pharmacological
inhibitors of PI3K/AKT and ERK1/2 MAPK, synergistically induced
apoptosis by increasing the proportion of SKOV3 cells arrested in
the sub-G1 phase of the cell cycle. Delphinidin-reduced
proliferation of SKOV3 cells was also required for paclitaxel to
decrease the proliferation of SKOV3 cells. In conclusion, these
findings provide evidence for the synergistic anticancer effects of
delphinidin combined with paclitaxel in human ovarian cancer SKOV3
cells, involving the inactivation of the PI3K/AKT and ERK1/2 MAPK
signaling pathways. Thus, delphinidin may be developed as a novel
therapeutic agent for the enhancement of survival in patients with
EOC.
Acknowledgements
The authors thank Dr Fuller W. Bazer (Texas A&M
University, College Station, USA) for their thoughtful editing and
comments on this manuscript. The present study was supported by
grants from the Basic Science Research Program (grant no.
2015R1D1A1A01059331) of the Republic of Korea and also supported by
School of Life Sciences and Biotechnology for BK21 PLUS, Korea
University.
References
|
1
|
Ververidis F, Trantas E, Douglas C,
Vollmer G, Kretzschmar G and Panopoulos N: Biotechnology of
flavonoids and other phenylpropanoid-derived natural products. Part
I: Chemical diversity, impacts on plant biology and human health.
Biotechnol J. 2:1214–1234. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zamora-Ros R, Touillaud M, Rothwell JA,
Romieu I and Scalbert A: Measuring exposure to the polyphenol
metabolome in observational epidemiologic studies: Current tools
and applications and their limits. Am J Clin Nutr. 100:11–26. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Prior RL and Wu X: Anthocyanins:
Structural characteristics that result in unique metabolic patterns
and biological activities. Free Radic Res. 40:1014–1028. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mazza G and Miniati E: Anthocyanins in
fruits, vegetables, and grains. CRC Press; Boca Raton: 1993
|
|
5
|
Favot L, Martin S, Keravis T,
Andriantsitohaina R and Lugnier C: Involvement of cyclin-dependent
pathway in the inhibitory effect of delphinidin on angiogenesis.
Cardiovasc Res. 59:479–487. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lamy S, Lafleur R, Bédard V, Moghrabi A,
Barrette S, Gingras D and Béliveau R: Anthocyanidins inhibit
migration of glioblastoma cells: Structure-activity relationship
and involvement of the plasminolytic system. J Cell Biochem.
100:100–111. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Noda Y, Kaneyuki T, Mori A and Packer L:
Antioxidant activities of pomegranate fruit extract and its
anthocyanidins: Delphinidin, cyanidin, and pelargonidin. J Agric
Food Chem. 50:166–171. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ozbay T and Nahta R: Delphinidin inhibits
HER2 and Erk1/2 signaling and suppresses growth of
HER2-overexpressing and triple negative breast cancer cell lines.
Breast cancer (Auckl). 5:143–154. 2011.PubMed/NCBI
|
|
9
|
Ko H, Jeong MH, Jeon H, Sung GJ, So Y, Kim
I, Son J, Lee SW, Yoon HG and Choi KC: Delphinidin sensitizes
prostate cancer cells to TRAIL-induced apoptosis, by inducing DR5
and causing caspase-mediated HDAC3 cleavage. Oncotarget.
6:9970–9984. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pal HC, Sharma S, Strickland LR, Agarwal
J, Athar M, Elmets CA and Afaq F: Delphinidin reduces cell
proliferation and induces apoptosis of non-small-cell lung cancer
cells by targeting EGFR/VEGFR2 signaling pathways. PLoS One.
8:e772702013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Feng R, Wang SY, Shi YH, Fan J and Yin XM:
Delphinidin induces necrosis in hepatocellular carcinoma cells in
the presence of 3-methyladenine, an autophagy inhibitor. J Agric
Food Chem. 58:3957–3964. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yun JM, Afaq F, Khan N and Mukhtar H:
Delphinidin, an anthocyanidin in pigmented fruits and vegetables,
induces apoptosis and cell cycle arrest in human colon cancer
HCT116 cells. Mol Carcinog. 48:260–270. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Filipiak K, Hidalgo M, Silvan JM, Fabre B,
Carbajo RJ, Pineda-Lucena A, Ramos A, de Pascual-Teresa B and de
Pascual-Teresa S: Dietary gallic acid and anthocyanin cytotoxicity
on human fibrosarcoma HT1080 cells. A study on the mode of action.
Food Funct. 5:381–389. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
de Hullu JA, van der Avoort IA, Oonk MH
and van der Zee AG: Management of vulvar cancers. Eur J Surg Oncol.
32:825–831. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kochli OR, Sevin BU, Averette HE and
Haller U: Overview of currently used chemosensitivity test systems
in gynecologic malignancies and breast cancer. Contrib Gynecol
Obstet. 19:12–23. 1994.PubMed/NCBI
|
|
16
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Raja FA, Chopra N and Ledermann JA:
Optimal first-line treatment in ovarian cancer. Ann Oncol. 23 Suppl
10:x118–x127. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Javadi S, Ganeshan DM, Qayyum A, Iyer RB
and Bhosale P: Ovarian cancer, the revised FIGO staging system and
the role of imaging. AJR Am J Roentgenol. 206:1351–1360. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Coleman RL, Monk BJ, Sood AK and Herzog
TJ: Latest research and treatment of advanced-stage epithelial
ovarian cancer. Nat Rev Clin Oncol. 10:211–224. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tripathi V, Kumar R, Dinda AK, Kaur J and
Luthra K: CXCL12-CXCR7 signaling activates ERK and Akt pathways in
human choriocarcinoma cells. Cell Commun Adhes. 21:221–228. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hou DX, Fujii M, Terahara N and Yoshimoto
M: Molecular mechanisms behind the chemopreventive effects of
anthocyanidins. J Biomed Biotechnol. 2004:321–325. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hou DX, Yanagita T, Uto T, Masuzaki S and
Fujii M: Anthocyanidins inhibit cyclooxygenase-2 expression in
LPS-evoked macrophages: Structure-activity relationship and
molecular mechanisms involved. Biochemical pharmacology.
70:417–425. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lamy S, Blanchette M, Michaud-Levesque J,
Lafleur R, Durocher Y, Moghrabi A, Barrette S, Gingras D and
Béliveau R: Delphinidin, a dietary anthocyanidin, inhibits vascular
endothelial growth factor receptor-2 phosphorylation.
Carcinogenesis. 27:989–996. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Azevedo L, de Lima PL Alves, Gomes JC,
Stringheta PC, Ribeiro DA and Salvadori DMF: Differential response
related to genotoxicity between eggplant (Solanum melanogena) skin
aqueous extract and its main purified anthocyanin (delphinidin) in
vivo. Food Chem Toxicol. 45:852–858. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yeh CT and Yen GC: Induction of apoptosis
by the Anthocyanidins through regulation of Bcl-2 gene and
activation of c-Jun N-terminal kinase cascade in hepatoma cells. J
Agric Food Chem. 53:1740–1749. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Smolle E, Taucher V, Pichler M, Petru E,
Lax S and Haybaeck J: Targeting signaling pathways in epithelial
ovarian cancer. Int J Mol Sci. 14:9536–9555. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Luo J, Manning BD and Cantley LC:
Targeting the PI3K-Akt pathway in human cancer: Rationale and
promise. Cancer Cell. 4:257–262. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Osaki M, Oshimura M and Ito H: PI3K-Akt
pathway: Its functions and alterations in human cancer. Apoptosis.
9:667–676. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ip CKM and Wong AST: Exploiting p70 S6
kinase as a target for ovarian cancer. Expert Opin Ther Tar.
16:619–630. 2012. View Article : Google Scholar
|
|
30
|
Meng Q, Xia C, Fang J, Rojanasakul Y and
Jiang BH: Role of PI3K and AKT specific isoforms in ovarian cancer
cell migration, invasion and proliferation through the p70S6K1
pathway. Cell Signal. 18:2262–2271. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Pon YL, Zhou HY, Cheung AN, Ngan HY and
Wong AS: p70 S6 kinase promotes epithelial to mesenchymal
transition through snail induction in ovarian cancer cells. Cancer
Res. 68:6524–6532. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Cheaib B, Auguste A and Leary A: The
PI3K/Akt/mTOR pathway in ovarian cancer: Therapeutic opportunities
and challenges. Chin J Cancer. 34:4–16. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Choi KC, Auersperg N and Leung PC:
Mitogen-activated protein kinases in normal and (pre)neoplastic
ovarian surface epithelium. Reprod Biol Endocrinol. 1:712003.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Roberts PJ and Der CJ: Targeting the
Raf-MEK-ERK mitogen-activated protein kinase cascade for the
treatment of cancer. Oncogene. 26:3291–3310. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Nicosia SV, Bai W, Cheng JQ, Coppola D and
Kruk PA: Oncogenic pathways implicated in ovarian epithelial
cancer. Hematol Oncol Clin North Am. 17:927–943. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Bin Hafeez B, Asim M, Siddiqui IA, Adhami
VM, Murtaza I and Mukhtar H: Delphinidin, a dietary anthocyanidin
in pigmented fruits and vegetables: A new weapon to blunt prostate
cancer growth. Cell Cycle. 7:3320–3326. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Fan W, Sun L, Zhou JQ, Zhang C, Qin S,
Tang Y, Liu Y, Lin SS and Yuan ST: Marsdenia tenacissima extract
induces G0/G1 cell cycle arrest in human esophageal carcinoma cells
by inhibiting mitogen-activated protein kinase (MAPK) signaling
pathway. Chin J Nat Med. 13:428–437. 2015.PubMed/NCBI
|
|
38
|
Li A, Wang J, Wu M, Zhang X and Zhang H:
The inhibition of activated hepatic stellate cells proliferation by
arctigenin through G0/G1 phase cell cycle arrest: Persistent
p27(Kip1) induction by interfering with PI3K/Akt/FOXO3a signaling
pathway. Eur J Pharmacol. 747:71–87. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lin CJ, Chen TL, Tseng YY, Wu GJ, Hsieh
MH, Lin YW and Chen RM: Honokiol induces autophagic cell death in
malignant glioma through reactive oxygen species-mediated
regulation of the p53/PI3K/Akt/mTOR signaling pathway. Toxicol Appl
Pharmacol. 304:59–69. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Liu E, Li J, Shi S, Wang X, Liang T, Wu B
and Li Q: Sustained ERK activation-mediated proliferation
inhibition of farrerol on human gastric carcinoma cell line by
G0/G1-phase cell-cycle arrest. Eur J Cancer Prev. 25:490–499. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Oh I, Cho H and Lee Y, Cheon M, Park D and
Lee Y: Blockage of autophagy rescues the dual PI3K/mTOR inhibitor
BEZ235-induced growth inhibition of colorectal cancer cells. Dev
Reprod. 20:1–10. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Jia L, Zhang S, Ye Y, Li X, Mercado-Uribe
I, Bast RC Jr and Liu J: Paclitaxel inhibits ovarian tumor growth
by inducing epithelial cancer cells to benign fibroblast-like
cells. Cancer Lett. 326:176–182. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Lamendola DE, Duan Z, Yusuf RZ and Seiden
MV: Molecular description of evolving paclitaxel resistance in the
SKOV-3 human ovarian carcinoma cell line. Cancer Res. 63:2200–2205.
2003.PubMed/NCBI
|