Introduction
In the white population of European and American
countries, squamous cell carcinoma (SCC) is the second most common
skin morbidity after basal cell carcinoma (BCC) (1). In China, there are ~1.5–2.0 million new
cases a year, accounting for around 5–12% of all malignant tumors
(1). The overall effects of various
factors, such as ultraviolet (UV) exposure, autoimmune disease,
inflammation and bruising are thought to play a role in the
pathogenesis of cutaneous carcinoma (2). Tumorigenesis is closely related with
aberrant expression of transcription factors and proteins. The
forkhead box (FOX) protein family is a highly evolutionary
conserved protein family and includes a 7 same nucleotide core
sequence motif [5-(G/A)(T/C)(A/C)AA(C/T)A-3], which regulates the
expression of downstream target genes (3). FOXC2 regulates cell proliferation,
differentiation and invasion, and lymphangiogenesis and
angiogenesis in a variety of malignant tumors, including lung,
breast and cervical cancers (4,5). The
endogenous telomerase-inhibiting gene PinX1, is a potential tumor
suppressor that helps downregulate the activity of telomerase and
shorten the telomere, inducing apoptosis of cancer cells (6). Ki-67, a proliferating cell nuclear
antigen (PCNA), may indirectly reflect the proliferation of tumor
cells (7). Cyclin D1, a positive
regulator of cell cycle, also influences the proliferation activity
of tumor cells (8). We hypothesized
that the altered expression of FOXC2 in the skin plays a role in
the underlying mechanism of SCC, and determined the expression of
possible downstream factors whose expressions might also be changed
in SCC tissues.
Materials and methods
Study objects
Pathology skin specimens from patients diagnosed
with either SCC or BCC from October 2015 to October 2016 were used
for experiments (thirty samples in each group). Additionally,
thirty normal skin tissue samples were included as a control
group.
Among SCC patients, there were 16 men and 14 women,
aged from 35 to 68 years, with an average of 52.5±13.6 years. The
tumors were located at the head and face in 20 patients and in the
limbs in 10 patients. TNM staging placed 6 patients in stage I, 10
patients in stage II, 3 patients in stage III and 3 patients in
stage IV. The largest diameter of the tumors ranged from 0.3 to 3.5
cm (with an average of 1.6±0.9 cm). According to classification by
pathological grade, 3 patients were grade I, 6 were grade II, 12
grade III and 9 grade IV.
Among BCC patients, there were 14 men and 16 women,
their ages ranged from 39–75, with an average of 54.4±16.7 years. A
total of 22 patients had tumors in the head and face and 8 patients
in the limbs. According to TNM staging, there were 5 patients in
stage I, 8 patients in stage II, 13 patients in stage III and 4
patients in stage IV. The largest diameter of the tumors ranged
from 0.2 to 3.2 (1.4±0.7 cm on average). According to pathological
grading, there were 5 patients in grade I, 8 in grade II, 11 in
grade III and 6 in grade IV.
Finally, in the control group there were 15 men and
15 women, their ages ranged from 30 to 70 years (52.2±15.8 years on
average). A total of 16 normal tissues were taken from the head and
face and 14 from the limbs.
The baseline data of all groups is comparable. This
study was approved by the Ethics Committee of Affiliated Hospital
of Weifang Medical University. Signed written informed consents
were obtained from all participants before the study.
Immunohistochemistry (IHC)
IHC was used to measure the protein expression of
FOXC2, PinX, Ki-67 and Cyclin D1 in the skin specimens obtained.
After fixing and paraffin embedding, the tissues were cut at 5 µm,
then de-paraffinized and hydrated. The samples were incubated with
3% H2O2 solution for 20 min at 27°C for
blocking, then incubated with normal goat serum working solution
(Biosharp, Hefei, China) for 30 min at 27°C. Next, the slices were
incubated overnight at 4°C with mouse monoclonal FOXC2 antibody
(dilution, 1:500; cat. no. SAB1412035), mouse monoclonal PinX1
antibody (dilution, 1:500; cat. no. SAB2500795), mouse monoclonal
Ki-67 antibody (dilution, 1:500; cat. no. WH0004288M1), mouse
monoclonal Cyclin D1 antibody (dilution, 1:500; cat. no.
SAB4503500), all purchased from Sigma (St. Louis, MO, USA). Goat
anti-mouse IgG secondary polyclonal antibodies (dilution, 1:1,000;
cat. no. ab6789; Abcam, Cambridge, MA, USA) were used next for
incubation at 27°C for 20 min. Finally, the tissues were incubated
with horse-radish peroxidase labelled streptavidin working solution
at 27°C for 20 min. The slices were washed with PBS 3 times, each
for 5 min, and DAB solution was added. The following procedures
included hematoxylin staining, dehydrating, and mounting of the
slices with neutral balsam. A microscope (BX-42; Olympus, Tokyo,
Japan) was used to observe the staining of the target proteins on
the tissues.
Slices were evaluated according to the staining
intensity and the percentage of stained cells using
semi-quantitative method. The staining intensity in both cytoplasm
and nucleus was scored and stratified as follows: grade 0 (no
staining or negative), grade 1, (light yellow or weak positive),
grade 2 (yellow or moderate positive) and grade 3 (yellow-brown or
strong positive). A staining score was defined as follows: score 0
(the number of positive cells ≤5%), score 1 (6–25% of cells were
stained), score 2 (26–50% of cells were stained), score 3 (51–75%
of cells were stained) and score 4 (>76% of cells were stained).
A final immunoreactivity score (IRS) was obtained for each case by
multiplying the grade times the score. Protein expression levels
were defined as negative if the IRS equaled 0–3, or positive if the
IRS value equaled 4–12.
Reverse transcription polymerase chain
reaction (RT-PCR)
Semi-quantitative RT-PCR was used to measure the
gene expression of FOXC2, PinX, Ki-67 and Cyclin D1.
Total RNA samples were extracted from each specimen
with TRIzol solution (ZSGB-Bio, Beijing, China). The purity and
concentration of total RNA samples were verified with an
ultraviolet spectrophotometer (Europe B.V., Venlo, The
Netherlands). cDNA was synthesized with a reverse transcription kit
(Takara Bio, Inc., Otsu, Japan) according to the manufacturers
instructions. Primers were designed according to the Gene Bank
sequences and they were purchased from Sangon Biotech (Shanghai,
China). RT-PCR primer sequence information is shown in Table I.
 | Table I.RT-PCR primer sequences. |
Table I.
RT-PCR primer sequences.
| Genes | Primer sequences |
|---|
| FOXC2 | F:
5-GCCGACGGATTCCTGCGCTC-3 |
|
| R:
5-CGCTCCTCGCTGGCTCCA-3 |
| PinX1 | F:
5-GGTGGTCTAAAGGAAAGGGTTT-3 |
|
| R:
5-ATGGGCAATCCAGTTGTCTT-3 |
| Ki-67 | F:
5-GAGAATCTGTGAATCTGGGTAA-3 |
|
| R:
5-CAGGCTTGCTGAGGGAAT-3 |
| Cyclin D1 | F: 5-
GGTTTCATCCAGGATCGAGCAGG-3 |
|
| R:
5-ACAAAGATGGTCACGGTCTGCC-3 |
| GAPDH | F:
5-CGCGAGAAGATGACCCAGAT-3 |
|
| R:
5-GCACTGTGTTGGCGTACAGG-3 |
A total of 25 µl reactions were prepared for each
RT-PCR containing 2 µl cDNA, 3 µl of F-primer, 3 µl of R-primer,
0.5 µl of Taq DNA polymerase, 1 µl of dNTPs and 3 µl of
MgCl2. An initial amplification using primers was done
with a denaturation step at 95°C for 5 min, followed by 30 repeated
cycles of 95°C for 30 sec, 58°C for 30 sec and 72°C for 60 sec. A
final elongation step at 72°C for 10 min followed. Next, 2% agarose
gel electrophoresis was used to identify the PCR products by their
predicted size. Analyze the grey level using ultraviolet
spectrometry imaging with gel imaging analysis system (Applied
Biosystems Life Technologies, Foster City, CA, USA). The result was
measured by the method of 2−∆∆Cq.
Statistics analysis
SPSS 20.0 software (SPSS Inc., Chicago, IL, USA) was
used to conduct the statistical analyses. The numeric data were
expressed as mean ± standard deviation. One-way analysis of
variance (ANOVA) was used to assess the difference between groups.
LSD t-test was used to compare the mean between two groups. Case
number or % was used to indicate categorical data. χ2
test was used for comparison between groups. It was considered as
statistically significant at p<0.05.
Results
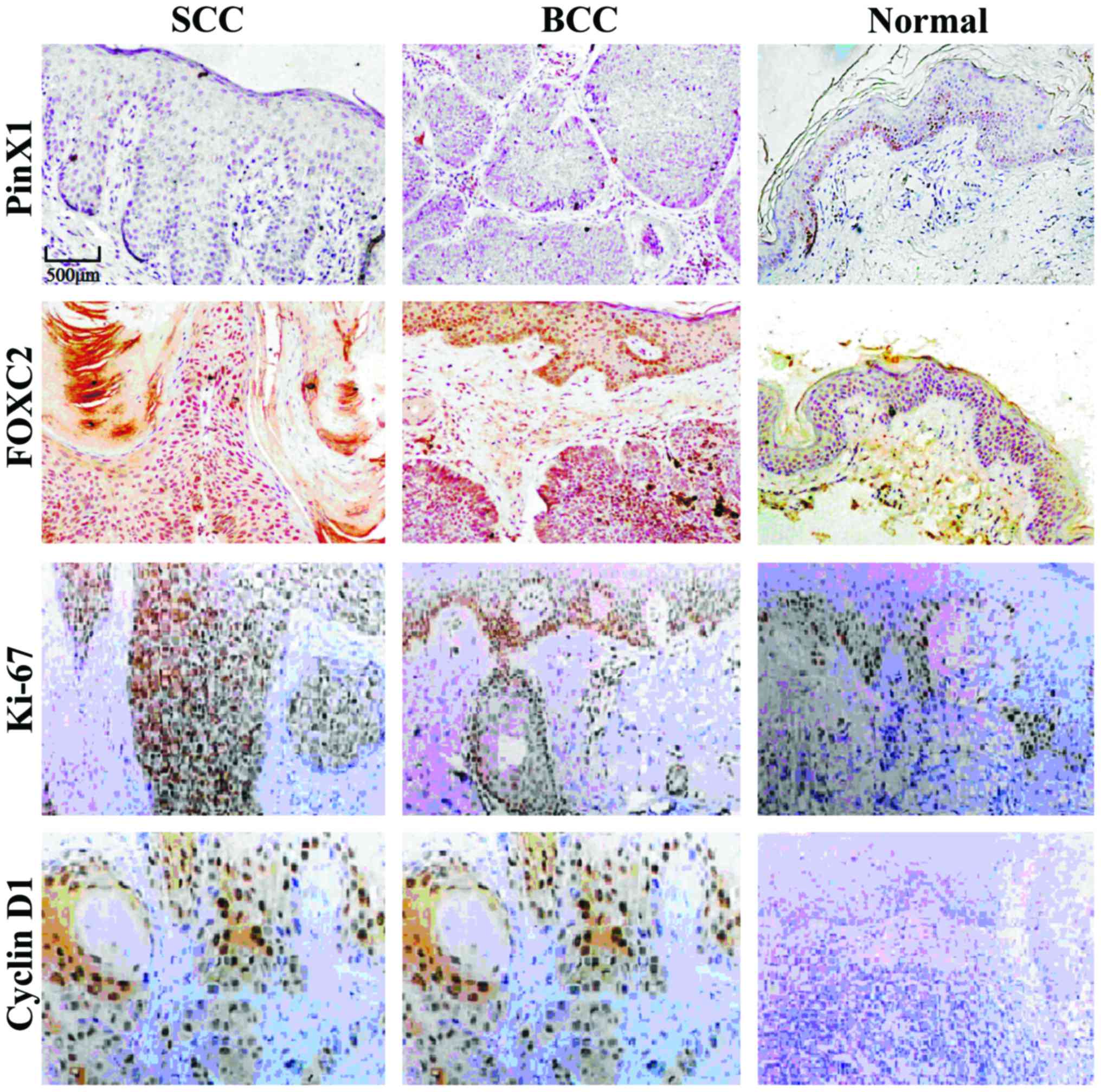
Protein expression of FOXC2, PinX,
Ki-67 and Cyclin D1
The expression levels of PinX1 in SCC and BCC were
significantly lower than that in normal skin tissues. However, the
protein expression levels of FOXC2, Ki-67 and Cyclin D1 were
significantly higher than those of normal tissues (p<0.05).
There was no significant correlation between SCC and BCC
(p>0.05, Fig. 1 and Table II). The expression level of PinX1 in
SCC and BCC tissues was significantly lower than that of normal
tissues, while the protein expression levels of FOXC2, Ki-67 and
Cyclin D1 were significantly higher than those in normal tissues.
No significant correlation was observed between SCC and BCC.
| Table II.Protein expression of PinX1, FOXC2,
Ki-67 and Cyclin D1 [n (%)] |
Table II.
Protein expression of PinX1, FOXC2,
Ki-67 and Cyclin D1 [n (%)]
| Group | n | PinX1 | FOXC2 | Ki-67 | Cyclin D1 |
|---|
| SCC | 30 | 6 (20.0) | 22 (73.3) | 20 (66.7) | 18 (60.0) |
| BCC | 30 | 8 (26.7) | 21 (70.0) | 18 (60.0) | 16 (53.3) |
| Normal | 30 | 24 (80.0) | 10 (33.3) | 6 (20.0) | 8 (26.7) |
| χ2 |
| 26.599 | 12.208 | 15.296 | 7.500 |
| P-value |
| <0.001 | 0.002 | <0.001 | 0.024 |
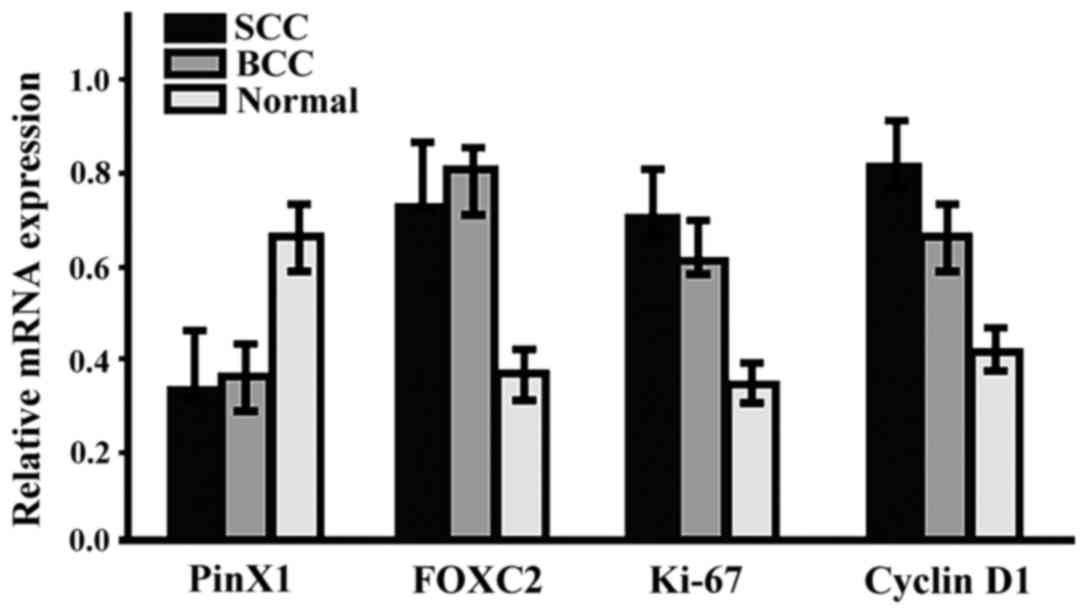
The gene expression of PinX1, FOXC2,
Ki-67 and Cyclin D1
The mRNA expression level of PinX1 in SCC and BCC
was significantly lower than that in normal tissues, while the mRNA
expression levels of FOXC2, Ki-67 and Cyclin D1 were significantly
higher than those in normal tissues (p<0.05). No significant
correlation between SCC and BCC was observed (p>0.05, Fig. 2).
Discussion
Researchers have found FOXC2 expressed in the
vascular endothelium of human and murine melanomas (9). After hypodermic injection of malignant
melanoma tumor cells into wild-type and FOXC2 heterozygous mutant
(FOXC2+/−) B16 mice, it was clear that the tumor growth
and angiogenesis in the mutant mice was significantly slower than
the same in the wild-type mice, suggesting that FOXC2 plays
critical roles in tumor angiogenesis. FOXC2 inhibits apoptosis of
vascular endothelium by increasing the expression of vascular
endothelium growth factor A (VEGF-A) (10). In addition, FOXC2 sets up a local
tumor microenvironment by enhancing the secretion and expression of
matrix metalloproteinase-2 (MMP-2), stromal-derived factor-1
(SDF-1) and some other active molecules (11). Moreover, FOXC2 has been shown to be
involved in tumor invasion and metastasis, as well as in the
development of the epithelial-mesenchymal transition (EMT) observed
in many tumors (12).
In this study, both the protein and mRNA levels of
PinX1 in SCC and BCC were significantly lower than those in normal
skin tissues, while the expression rates of FOXC2, Ki-67 and Cyclin
D1 were higher than those of normal tissues (p<0.05). However,
no significant differences were found when comparing levels between
SCC and BCC (p>0.05). This evidence indicates that FOXC2 may be
involved in the development of SCC and BCC and (based partly on
prior literature) it may do so by changing the levels of PinX1,
Ki-67 and Cyclin D1 in turn. However, FOXC2 cannot be used as a
specific biomarker to diagnose SCC. Nevertheless, the levels of
FOXC2 combined with clinical features of various malignant tumors,
may serve as a valuable biomarker for early diagnosis and prognosis
(13).
PinX1 can directly interact with human telomerase
reverse transcriptase (hTERT), which is a key speed-limiting enzyme
for the activity of telomerase (6).
PinX1 is widely expressed in human tissue and mostly locates in the
cell nucleolus and telomere proximity. However, the expression of
PinX1 in tumor tissues is limited at best (14). Ki-67 correlates with the
aggressiveness of a tumor (15), and
can thus be used as an objective indicator of the proliferation
activity of an SCC. Cyclin D1, on the other hand, is a key factor
regulating cell cycle of both normal and tumorigenic cells and its
expression correlates with tumor proliferation and differentiation
(16).
We hypothesized that the altered expression of FOXC2
in the skin plays a role in the underlying mechanism of SCC, and
determined the expression of possible downstream factors whose
expression might also be changed in SCC tissues. The aim of this
investigation was to show the role of FOXC2 in the development of
SCC and BCC. Additionally FOXC2 may promote the tumor development
by affecting the expression level of PinX1, Ki-67 and Cyclin D1,
which could be an important mechanism of oncogenesis and be used as
an early intervention in SCC. We intend to further analyze the
correlation between FOXC2 and the clinical features of SCC and BCC.
The reason why there is no significant difference of FOXC2
expression between SCC and BCC remains to be explored. The small
sample size of this study may influence the results. We plan to
adopt methods of in vitro gene expression silencing or the
overexpression of genetic vectors to further explore the specific
mechanism of FOXC2-induced tumorigenesis.
References
|
1
|
Farasat S, Yu SS, Neel VA, Nehal KS,
Lardaro T, Mihm MC, Byrd DR, Balch CM, Califano JA, Chuang AY, et
al: A new American Joint Committee on Cancer staging system for
cutaneous squamous cell carcinoma: Creation and rationale for
inclusion of tumor (T) characteristics. J Am Acad Dermatol.
64:1051–1059. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kwon S, Dong ZM and Wu PC: Sentinel lymph
node biopsy for high-risk cutaneous squamous cell carcinoma:
Clinical experience and review of literature. World J Surg Oncol.
9:802011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Shimeld SM, Degnan B and Luke GN:
Evolutionary genomics of the Fox genes: Origin of gene families and
the ancestry of gene clusters. Genomics. 95:256–260. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jiang W, Fan H, Qian C, Ding J, Wang Q and
Pang X: Prognostic value of high FoxC2 expression in resectable
non-small cell lung cancer, alone or in combination with E-cadherin
expression. BMC Cancer. 16:162016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Werden SJ, Sphyris N, Sarkar TR, Paranjape
AN, LaBaff AM, Taube JH, Hollier BG, Ramirez-Peña EQ, Soundararajan
R, den Hollander P, et al: Phosphorylation of serine 367 of FOXC2
by p38 regulates ZEB1 and breast cancermetastasis, without
impacting primary tumor growth. Oncogene. 35:5977–5988. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lai XF, Shen CX, Wen Z, Qian YH, Yu CS,
Wang JQ, Zhong PN and Wang HL: PinX1 regulation of telomerase
activity and apoptosis in nasopharyngeal carcinoma cells. J Exp
Clin Cancer Res. 31:122012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sobecki M, Mrouj K, Camasses A, Parisis N,
Nicolas E, Llères D, Gerbe F, Prieto S, Krasinska L, David A, et
al: The cell proliferation antigen Ki-67 organises heterochromatin.
eLife. 5:e137222016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Dong M, Wei H, Hou JM, Gao S, Yang DZ, Lin
ZH, Jia Y, Ren XP and Gao MH: Possible prognostic significance of
p53, cyclin D1 and Ki-67 in the second primary malignancy of
patients with double primary malignancies. Int J Clin Exp Pathol.
7:3975–3983. 2014.PubMed/NCBI
|
|
9
|
Sano H, Leboeuf JP, Novitskiy SV, Seo S,
Zaja-Milatovic S, Dikov MM and Kume T: The Foxc2 transcription
factor regulates tumor angiogenesis. Biochem Biophys Res Commun.
392:201–206. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sasahira T, Ueda N, Yamamoto K, Kurihara
M, Matsushima S, Bhawal UK, Kirita T and Kuniyasu H: Prox1 and
FOXC2 act as regulators of lymphangiogenesis and angiogenesis in
oral squamous cell carcinoma. PLoS One. 9:e925342014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li D, Yan D, Liu W, Li M, Yu J, Li Y, Qu Z
and Ruan Q: Foxc2 overexpression enhances benefit of endothelial
progenitor cells for inhibiting neointimal formation by promoting
CXCR4-dependent homing. J Vasc Surg. 53:1668–1678. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hollier BG, Tinnirello AA, Werden SJ,
Evans KW, Taube JH, Sarkar TR, Sphyris N, Shariati M, Kumar SV,
Battula VL, et al: FOXC2 expression links epithelial-mesenchymal
transition and stem cell properties in breast cancer. Cancer Res.
73:1981–1992. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Nishida N, Mimori K, Yokobori T, Sudo T,
Tanaka F, Shibata K, Ishii H, Doki Y and Mori M: FOXC2 is a novel
prognostic factor in human esophageal squamous cell carcinoma. Ann
Surg Oncol. 18:535–542. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhou XZ and Lu KP: The
Pin2/TRF1-interacting protein PinX1 is a potent telomerase
inhibitor. Cell. 107:347–359. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bedir R, Güçer H, Şehitoğlu I, Yurdakul C,
Bağcı P and Üstüner P: The role of p16, p21, p27, p53 and Ki-67
expression in the differential diagnosis of cutaneous squamous cell
carcinomas and keratoacanthomas: An immunohistochemical study.
Balkan Med J. 33:121–127. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shen Y, Xu J, Jin J, Tang H and Liang J:
Cyclin D1 expression in Bowens disease and cutaneous squamous cell
carcinoma. Mol Clin Oncol. 2:545–548. 2014.PubMed/NCBI
|