Introduction
Breast cancer is the second leading cause of
cancer-associated mortality in females in the USA, and it had an
estimated annual diagnosis rate of 246,660 people and annual
mortality rate of 40,450 cases in 2016 (1). Currently, the primary therapeutic
strategy for breast cancer patients is surgery, while chemotherapy,
radiotherapy and hormonal therapy are also used. Despite the
impressive advances in breast cancer treatment, relapse and
metastasis remain inevitable in breast cancer patients (2). Therefore, determining the molecular
mechanisms underlying breast cancer migration and invasion may
facilitate the identification of novel therapeutic targets and
consequently lead to the improvement of prognosis in the
future.
MicroRNAs (MiRNAs) are noncoding RNAs with 20–22
nucleotides. They post-transcriptionally modulate gene expression
by binding to the 3′-untranslated region (3′-UTR) of the target
mRNAs (3). As is well known, miRNAs
are involved in tumour cell proliferation, migration and invasion
(4). Moreover, altered miRNA
expression is involved in breast cancer pathogenesis via the
modulation of oncogenes and tumour suppressors that subsequently
affect the downstream signalling pathway (5–7). For
instance, miR-411, miR-490-3p and miR-145 were significantly
down-regulated in breast cancer cases and played a critical role in
the suppression of tumour metastasis (8–10).
Although the pathogenesis of breast cancer metastasis was already
investigated, the mechanism was not completely illustrated. Crucial
miRNAs that might play significant roles in breast cancerogenesis
must be identified. The newly discovered miRNA hsa-miR-599 plays an
important role in the development of hepatocellular carcinoma
(11). However, the mechanism
underlying the development and progression of breast cancer remains
unknown.
In this study, we firstly detected the expression
level of hsa-miR-599 in breast cancer tissues and metastatic cell
lines. Secondly, we investigated the role of hsa-miR-599 in breast
cancer proliferation, migration and invasion in vitro and
in vivo. Finally, bromodomain containing 4 (BRD4) was
identified as a direct target of hsa-miR-599. The results revealed
that hsa-miR-599 functioned as a tumour suppressor in breast
cancer.
Materials and methods
Cell culture
Breast cancer cell lines MCF-7 (TCHu74), MDA-MB-231
(TCHu104) were purchased from the Shanghai Cell Bank, Chinese
Academy of Sciences. BT474 (HTB-20™), and T-47D (HTB-133) and
normal mammary epithelial cell line MCF 10A (CRL-10317) were
obtained from the American Type Culture Collection (Manassas, VA,
USA). The cell identity was confirmed by STR analysis. The MCF-7,
BT474, MDA-MB-231 and T-47D cells were cultured in Dulbecco's
modified Eagle's medium (DMEM; HyClone; GE Healthcare Life
Sciences, Logan, UT, USA) supplemented with 10% fetal bovine serum
(FBS) (HyClone; GE Healthcare Life Sciences). The MCF-10A cells
were cultured in DMEM/F12 medium (HyClone; GE Healthcare Life
Sciences) containing 5% horse serum (Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA), 1% penicillin/streptomycin
(HyClone; GE Healthcare Life Sciences), 20 ng/ml of EGF (Gibco;
Thermo Fisher Scientific, Inc.), 0.5 µg/ml of hydrocortisone
(Sigma, St. Louis, MO, USA). All cells were incubated in a
humidified atmosphere with 5% CO2 and humidified sphere
of 95% at 37°C.
Cell transfection
An hsa-miR-599 mimic (sense
5′-CUGUCCACAGUGUGUUUGAUAAG-3′) were chemically synthesised by
Shanghai GenePharma Co., Ltd. (Shanghai, China). RNA with no
homology to any human genomic sequence was used as negative control
(NC) (sense 5′-ACUACUGAGUGACAGUAGA-3′). For convenience, the
hsa-miR-599 mimic and the NC were designated as miR-599 and NC,
respectively. A BRD4 overexpression plasmid (sc-425356-ACT) and
control vector (sc-437275) were obtained from Santa Cruz
Biotechnology, Inc. (Dallas, TX, USA). During cell transfection,
the cells were seeded in six-well plates and then cultured until 50
to 70% confluency was reached in 1 day. Transfection was performed
using a Lipofectamine® 2000 Reagent (Invitrogen; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) according to the
manufacturer's protocols. The transfection mixture was replaced in
a medium containing 10% FBS after 6 to 8 h.
Cell viability assay
Cell viability was measured using a Cell Counting
Kit-8 (CCK-8) assay (Beyotime Institute of Biotechnology, Haimen,
China). The cells were transfected with miR-599 and control and
then cultured overnight. Subsequently, the cells were trypsinised
and seeded at 3,000 cells/well in a 96-well plate. After culturing
at indicated time periods (0, 24, 48 and 72 h), 10 µl of the CCK-8
solution was added into each well. The resulting mixtures were
incubated at 37°C. After 3 h, the absorbance of each well was
measured using a Multiskan MK3 spectrophotometer set at a
wavelength of 450 nm.
Colony formation assay
The cells transfected with the miR-599 mimic and
control were seeded at 200 cells/well in the 6-well plates. After 1
week of culture, the colonies were stained with 0.5% crystal violet
(Beyotime Institute of Biotechnology), and the images of the
stained colonies were captured using a CKX41 light microscope. The
number of the colonies was counted using the images.
Cell migration and invasion assay
Cell migration and invasion were assessed by
performing a Boyden chamber assay. For the invasion assay, the
upper sides of the filters were coated with 50 µl of Matrigel (BD
Biosciences, Bedford, MA, USA). Cells were harvested at 48 h
post-transfection. Approximately 5×104 cells with 200 µl
of serum-free medium were seeded in the upper chamber. The lower
chamber was filled with medium supplemented with 5% FBS. After
incubation at 37°C with 5% CO2 for 8 h (migration) or 12
h (invasion), the cells on the lower filter were fixed with
methanol and stained with crystal violet. The stained cells were
then counted under a light microscope (CKX41; Olympus Corporation,
Tokyo, Japan).
Luciferase assay
BRD4 3′-UTR-luciferase reporter vectors were created
by ligating the BRD4 3′-UTR polymerase chain reaction (PCR)
products into the XhoI and NotI restriction sites of
the psiCHECK-2™ Vector (Promega Corporation, Madison, WI, USA). The
mutant 3′-UTR regions were chemically synthesised and ligated into
the psiCHECK-2™ vector. The cells were cultured in 24-well plates,
and each well was transfected with 250 ng of vectors with 50 nM
Hsa-miR-599 mimic or control. After 48 h of co-transfection, the
luciferase activity was measured using a dual-luciferase reporter
assay system (Promega Corporation) according to the manufacturer's
instructions.
Western blot analysis
Equal amounts of proteins (30 µg) from the lysates
of the cells were subjected to electrophoresis through a 10%
SDS-PAGE (Beyotime Institute of Biotechnology) at 80 V for 30 min
and at 100 V for 1.5 h. The proteins were then transferred onto
polyvinylidene difluoride membranes. After blocking in 5% skimmed
milk, the membranes were then incubated with the following diluted
primary antibodies: Rabbit polyclonal BRD4 (Abcam, Cambridge, MA,
USA), mouse monoclonal β-actin (Beyotime Institute of
Biotechnology) overnight at 4°C and horseradish
peroxidase-conjugated goat anti-rabbit antibody (Santa Cruz
Biotechnology, Inc.) at room temperature for 2 h. Specific bands
were visualised on an autoradiographic film using an enhanced
chemiluminescence reagent (Nanjing KeyGen Biotech Co., Ltd.,
Nanjing, China).
Quantitative PCR (qPCR)
The total RNA from cells was isolated using TRIzol
reagent (Invitrogen; Thermo Fisher Scientific, Inc.) and then
reverse-transcribed using a PrimeScript RT Reagent kit with gDNA
Eraser (Takara Biotechnology Co., Ltd., Dalian, China) according to
the manufacturer's instructions. qPCR was performed using a SYBR
Premix Ex Taq. The specificity of the amplification was verified
using a melting curve and electrophoresis in agarose gel. The
following PCR conditions were used for the detection of the mRNAs:
95°C for 30 sec, followed by 40 cycles of 95°C for 30 sec, 60°C for
30 sec and 72°C for 30 sec. The β-actin and U6 small nuclear RNA
were used as internal controls for the detection. The relative
expression levels of miR-599 and BRD4 were calculated as the
inverse log of ΔΔCq and normalised to the reference. The primers
used for amplification were the following: miR-599 forward
5′-GUUGUGUCAGUUUAUCAAAC-3′ and reverse 5′-GUUGUGUCAGUUUAUCAAAC-3′.
U6 forward 5′-TGCGGGTGCTCGCTTCGCAGC-3′ and reverse
5′-CCAGTGCAGGGTCCGAGGT-3′. BRD4 forward 5′-CATGGACATGAGCACAATCA-3′
and reverse 5′-TCATGGTCAGGAGGGTTGTA-3′. β-actin forward
5′-GATCATTGCTCCTCCTGAGC-3′ and reverse
5′-ACTCCTGCTTGCTGATCCAC-3′.
Clinical sample collection
Forty cases of surgically resected breast cancer
samples (tumour and adjacent nontumour tissue) were obtained from
Weifang People's Hospital. These samples were frozen in liquid
nitrogen immediately and stored at −80°C. None of the patients
received chemotherapy or radiotherapy before the surgery. All the
patients gave their written informed consents, and the study was
approved by the Ethics Committee of Weifang People's Hospital.
Animal studies
All animal studies were approved by the Animal Care
and Welfare Committee of WeiFang Medical University and conducted
in strict accordance with the guidelines of the National Animal
Welfare Law of China. Four-week-old female BALB⁄c nude mice were
purchased from the Laboratory Animal Center of Yangzhou University
(Yangzhou, China) and maintained in a specific pathogen-free
environment. Subcutaneous tumour xenografting was performed by
subcutaneously injecting the mice with 200 µl PBS containing
1×106 MCF-7 cells transfected with miR-599 mimic or
miR-NC. Tumour volume (mm3) was calculated every 5 days
using the formula: V=0.5 × length × width2. After 25
days, the mice were euthanised, and the tumours were isolated,
weighed, photographed and processed for immunohistochemistry.
Statistical analysis
All experiments were performed in triplicate. Unless
otherwise indicated, the experimental values were expressed as mean
± SEM. Statistical significance was determined by unpaired
Student's t-test using SPSS 13.0 (SPSS, Inc., Chicago, IL, USA).
P<0.05 was considered to indicate a statistically significant
difference.
Results
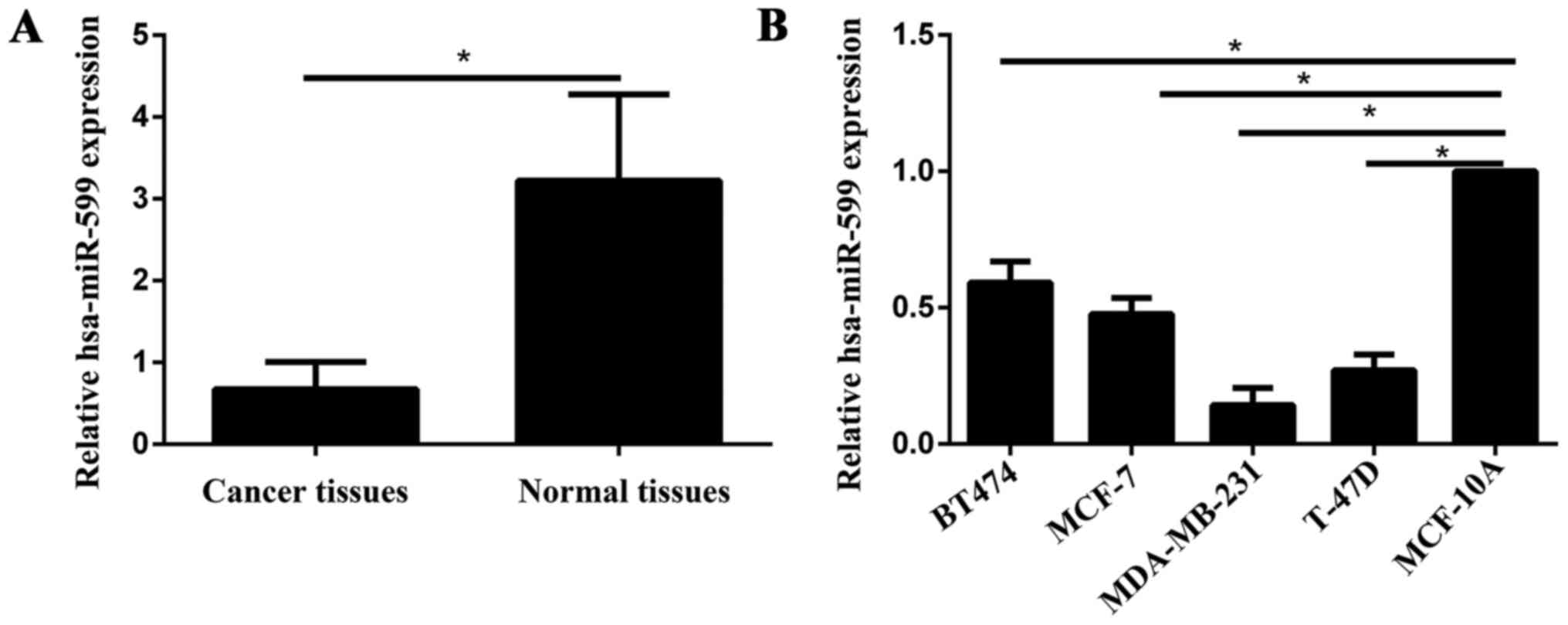
Hsa-miR-599 was downregulated in
breast cancer tissues and cell lines
The expression level of hsa-miR-599 was measured in
breast cancer specimens, and the adjacent normal tissues by qPCR.
As shown in Fig. 1A, the hsa-miR-599
expression levels in the breast cancer tissues was significantly
downregulated compared with those of the adjacent non-neoplastic
tissues (P<0.05). In the breast cancer cell lines, hsa-miR-599
expression levels were downregulated in the low-invasive cell lines
(MCF-7 and BT474) and high-invasive cell lines (MDA-MB-231 and
T-47D) compared with those in the normal mammary epithelial cell
line MCF-10A (Fig. 1B). Furthermore,
in the low-invasive and high-invasive cell lines, the expression of
hsa-miR-599 was low in the MCF-7 and MDA-MB-231, respectively.
Therefore, MCF-7 and MDA-MB-231 were selected for further
studies.
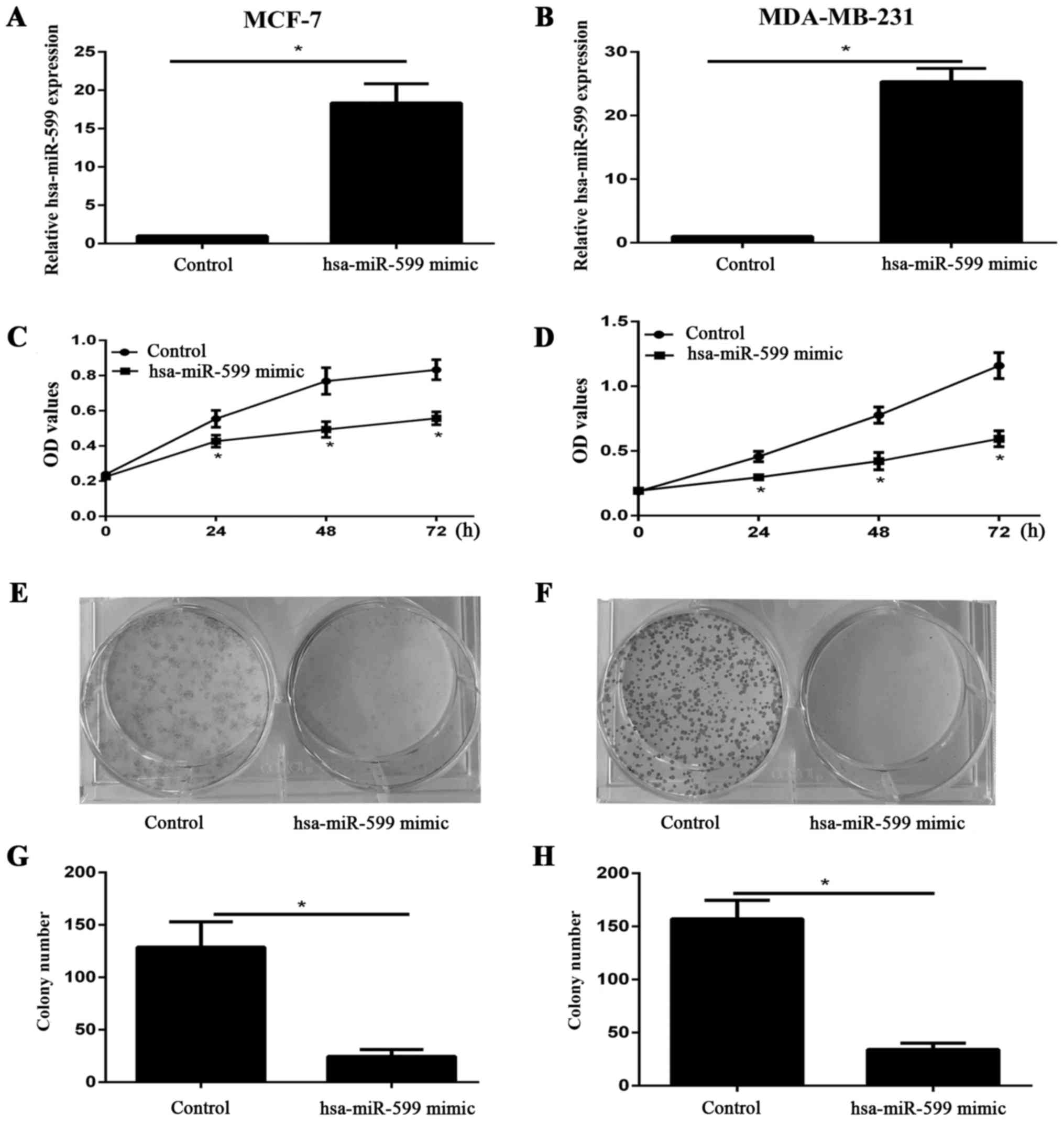
Hsa-miR-599 suppressed the viability
and proliferation of breast cancer cells in vitro
We examined the expression level of hsa-miR-599 in
cells after the transfection of hsa-miR-599 mimics or control into
MCF-7 and MDA-MB-231 through qPCR. The expression of hsa-miR-599 in
the mimic group was higher than that in the control group (Fig. 2A and≈B). The viability of MCF-7 and
MDA-MB-231 cells was investigated using CCK-8 assay after 24, 48,
and 72 h of incubation. Hsa-miR-599 mimic evidently inhibited the
viability of both cell lines (Fig. 2C and
D). Furthermore, the results of the colony formation assay
showed that the proliferation of hsa-miR-599 mimic cells was
severely inhibited (Fig. 2E-H). These
results suggested that hsa-miR-599 inhibited the growth of breast
cancer cells.
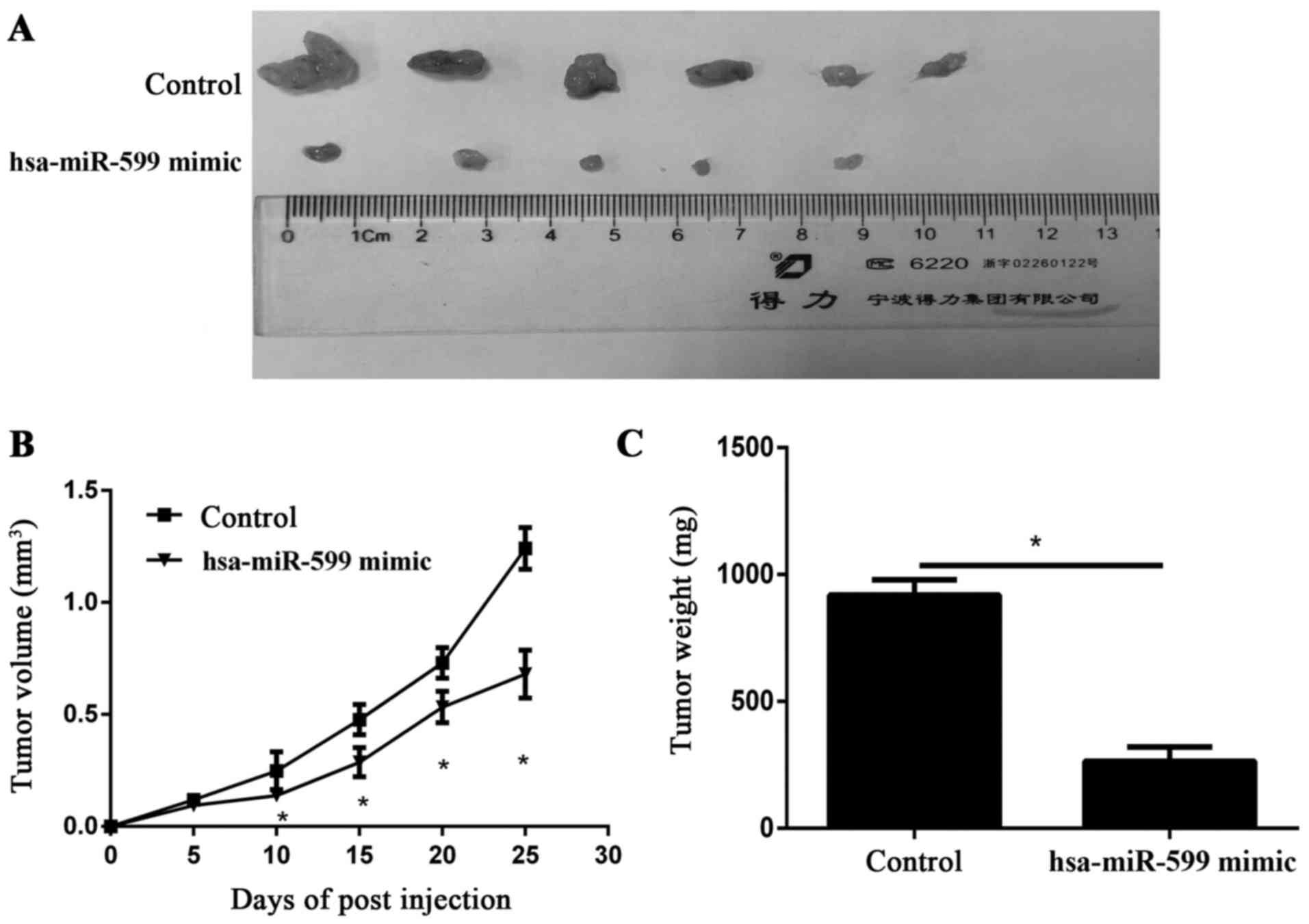
Hsa-miR-59 inhibited the breast cancer
progression in vivo
To study the relevance of hsa-miR-599 to breast
cancer progression in vivo, we subcutaneously transplanted
hsa-miR-599 mimics or miR-NC MCF-7 cells into nude mice
(n=6/group). Twenty five days after cell implantation, the tumours
were removed and photographed (Fig.
3A). The tumour volumes in the hsa-miR-599 mimic of the tumour
xenograft mice were smaller than those of the mock tumour xenograft
mice. Furthermore, the periodic measurements of the tumour volume
and the final weight of the excised tumours demonstrated that the
hsa-miR-599 mimics disrupted tumour growth at termination compared
with the control group (Fig. 3B and
C).
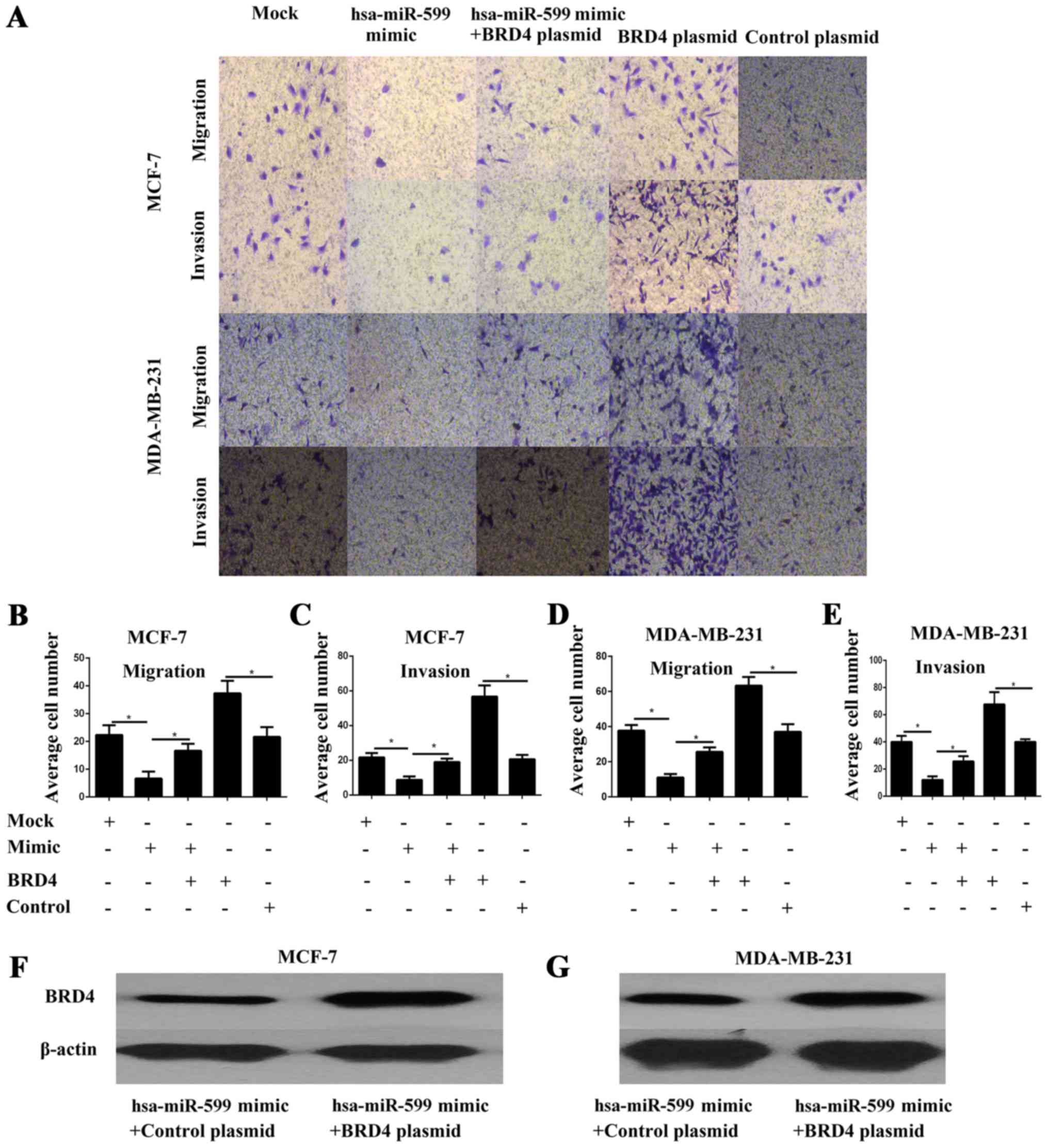
Hsa-miR-599 suppressed the migration
and invasion potential of breast cancer cells
Cell migration and invasion are critical events in
tumour metastasis. Thus, the effects of hsa-miR-599 on the
migration and invasion of breast cancer cells were explored in
vitro. The cell migration and invasion of MCF-7 and MDA-MB-231
were investigated through a transwell assay. The results showed
that the hsa-miR-599 mimics significantly inhibited cell migration
and invasion (P<0.05; Fig.
4A-E).
BRD4 is a direct target gene of
hsa-miR-599 in vitro
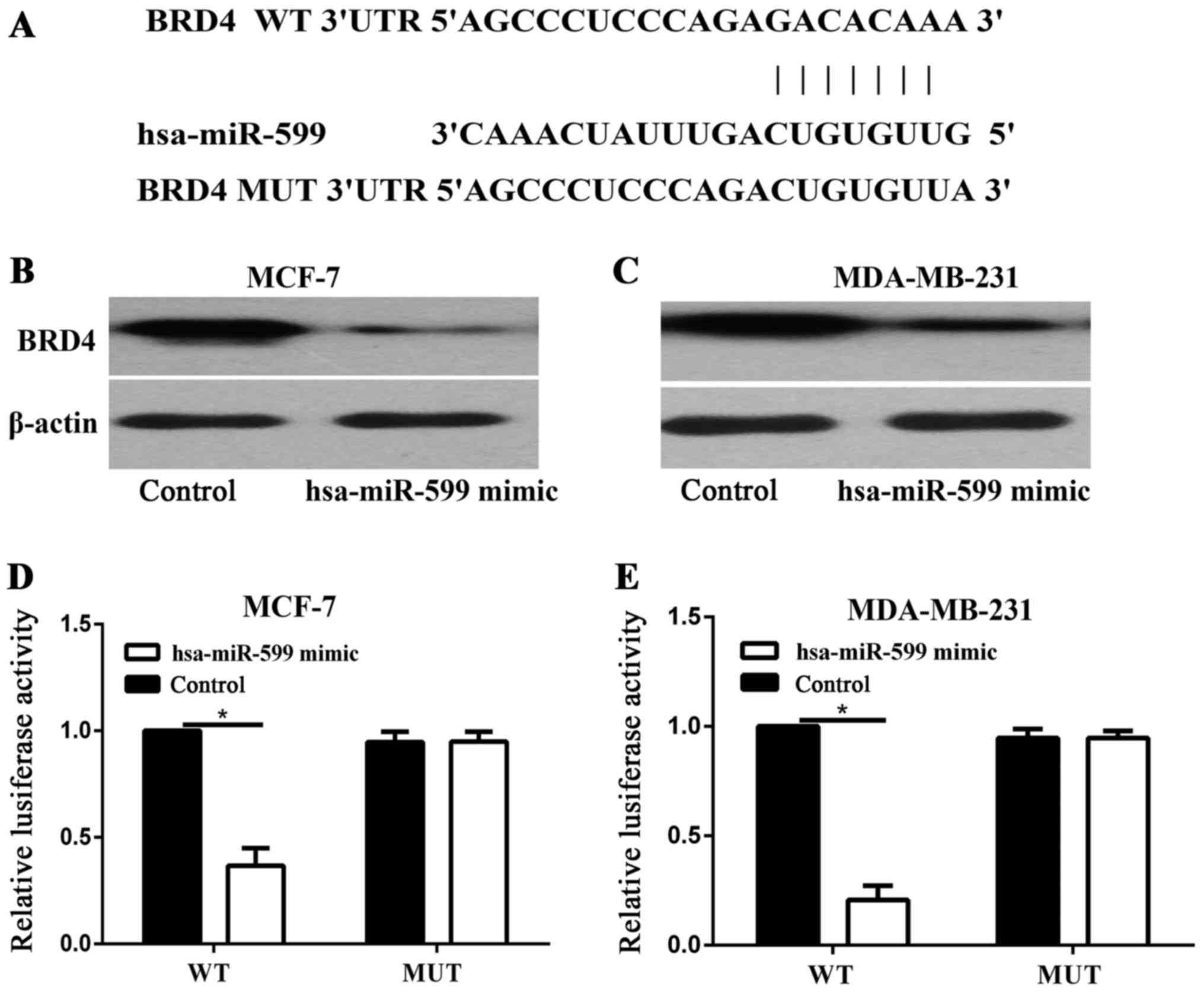
TargetScan was used to identify the target of
hsa-miR-599. As BRD4 was predicted to be a target of hsa-miR-599
(Fig. 5A), Western blot analysis was
performed to determine whether BRD4 was downregulated at the
protein level in breast cancer cells after it was transfected with
hsa-miR-599. As shown in Fig. 5B and
C, the expression levels of BRD4 was significantly reduced in
the MCF-7 and MDA-MB-231 cells after BRD4 was transfected with
hsa-miR-599 (P<0.05). Luciferase assays were then performed to
determine whether hsa-miR-599 directly target BRD4. As shown in
Fig. 5D and E, hsa-miR-599
significantly inhibited the luciferase activity of BRD4 WT, but not
of BRD4 MUT, in the MCF-7 and MDA-MB-231 cells. Overall, these
results indicated that BRD4 was a direct target gene of hsa-miR-599
in vitro.
Forced BRD4 expression partially
rescued the migration and invasion capability of
hsa-miR-599-transfected breast cancer cells
To further investigate whether the effects of
hsa-miR-599 on cell migration and invasion are mediated by BRD4, we
performed transwell assays in MCF-7 and MDA-MB-231 cells
co-transfected with hsa-miR-599 and BRD4 overexpressed plasmid.
BRD4 protein expression was confirmed by Western blot analysis
(Fig. 4F-G). The results showed that
forced BRD4 expression increased the migration and invasion of
breast cancer cells (Fig. 4A-E). More
importantly, the partial but significant restoration of BRD4
rescued the migration and invasion of the hsa-miR-599 cells
(Fig. 4A-E). Overall, these results
illustrated that the effects of hsa-miR-599 on cell migration and
invasion were partly mediated by BRD4.
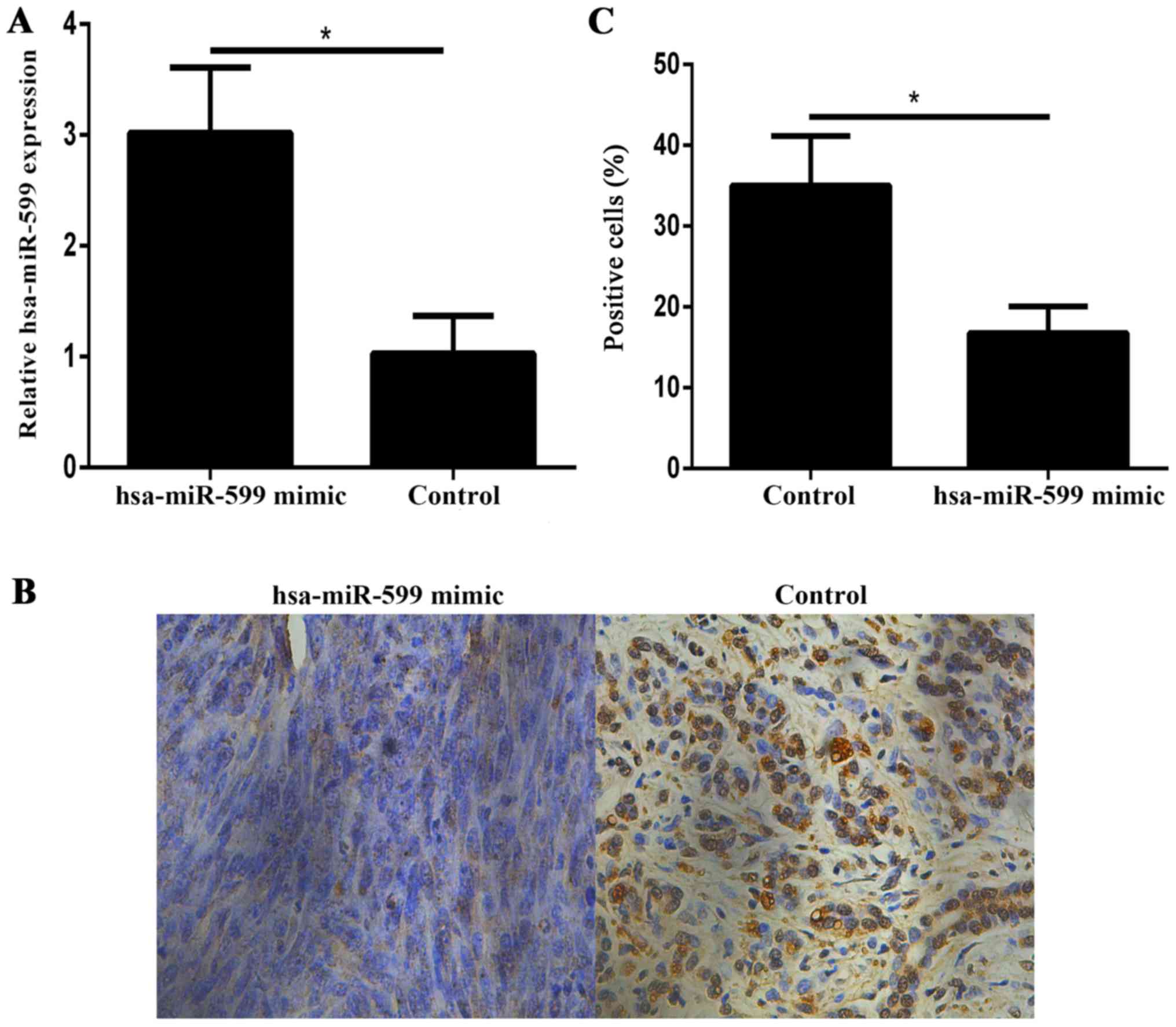
Hsa-miR-599 downregulates BRD4
expression in vivo
Immunohistochemistry showed that the BRD4 expression
levels in the hsa-miR-599 mimic tumour xenografts were lower than
those in the control xenografts (Fig.
6A-C).
Discussion
Increasing evidence has shown that miRNAs are
abnormally expressed in breast cancers, and changes in miRNA
expression affect the onset, development and metastasis of breast
cancers (6,12). However, miRNAs affecting breast cancer
progression require further exploration. Recent study has
identified that hsa-miR-599 served as tumour suppressor in
hepatocellular carcinoma. However, the function of hsa-miR-599 in
breast cancer has yet to be elucidated, and the mechanism through
which hsa-miR-599 inhibits tumour migration and invasion remains
unclear.
In the present study, we found that hsa-miR-599 is
downregulated in breast cancer tissues and breast cancer cell
lines. The colony formation assay showed that cells transfected
with hsa-miR-599 grew markedly slower, and the results of the
transwell assays revealed that hsa-miR-599 transfection inhibited
cell migration and invasion. Tumour xenografting revealed that
hsa-miR-599 transfection inhibited tumour volume and growth speed.
These results illustrated that hsa-miR-599 functioned as a tumour
suppressor during the proliferation and metastasis of breast
cancer.
Identifying hsa-miR-599 target genes is essential
for understanding their functions in the carcinogenesis and
progression of breast cancer. It is also important for the
investigation of novel targeted therapies for breast cancer. In the
present study, a molecular link between hsa-miR-599 and BRD4 was
identified. BRD4 is an epigenome reader and a member of the
bromodomain and extra-terminal (BET) family of proteins, which
consist of two bromodomains in tandem and an extra terminal domain.
Recent studies showed that BRD4 promotes cell cycle progression and
regulates cell growth and transcription (13). Subsequent studies demonstrated that
BRD4 played an important role in tumour proliferation and growth in
pancreatic cancer (14), melanoma
(15), neuroblastoma (16), glioblastoma (17), hepatocellular carcinoma (18), lung adenocarcinoma (19), malignant peripheral nerve sheath
tumours (20) and breast cancer
(21). However, the relation between
miRNA and BRD4 were not illustrated in these studies. In the
present study, BRD4 was confirmed as a direct target gene of
hsa-miR-599, as indicated by the results of luciferase assay and
Western blot analysis. Moreover, rescue experiments indicated that
the most important effect exerted by hsa-miR-599 on breast cancer
cells invasion and migration, which was partially reversed when
co-transfected with BRD4. These results demonstrated that BRD4 was
a functional target gene of hsa-miR-599 in breast cancer. Notably,
each miRNA can regulate numerous genes, and multiple miRNAs may
regulate the same gene, and these characteristics affect the
activities of pathways (22). In
fact, hsa-miR-599 regulates biological functions of different
diseases by targeting various genes, such as TGFB2 (23) and MYC (11). In our study, although increased
expression of hsa-miR-599 inhibited the migration and invasion of
breast cancer cells, these capabilities were partially rescued by
BRD4 overexpression. Thus, other target genes were possibly
involved in the suppressive effects of hsa-miR-599.
In conclusion, hsa-miR-599 inhibited breast cancer
cell progression and metastasis by targeting BRD4. Hsa-miR-599
restoration may provide new therapeutic methods for breast cancer
treatment.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Pivot X: Adjuvant chemotherapy for local
relapse breast cancer. Lancet Oncol. 15:125–126. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Vasudevan S, Tong Y and Steitz JA:
Switching from repression to activation: microRNAs can up-regulate
translation. Science. 318:1931–1934. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Shen J, Stass SA and Jiang F: MicroRNAs as
potential biomarkers in human solid tumors. Cancer Lett.
329:125–136. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Asghari F, Haghnavaz N, Baradaran B,
Hemmatzadeh M and Kazemi T: Tumor suppressor microRNAs: Targeted
molecules and signaling pathways in breast cancer. Biomed
Pharmacother. 81:305–317. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Peng F, Xiong L, Tang H, Peng C and Chen
J: Regulation of epithelial-mesenchymal transition through
microRNAs: Clinical and biological significance of microRNAs in
breast cancer. Tumour Biol. 37:14463–14477. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Guo L, Yuan J, Xie N, Wu H, Chen W, Song S
and Wang X: miRNA-411 acts as a potential tumor suppressor miRNA
via the downregulation of specificity protein 1 in breast cancer.
Mol Med Rep. 14:2975–2982. 2016.PubMed/NCBI
|
|
9
|
Zhao L and Zheng XY: MicroRNA-490 inhibits
tumorigenesis and progression in breast cancer. Onco Targets Ther.
9:4505–4516. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhao H, Kang X, Xia X, Wo L, Gu X, Hu Y,
Xie X, Chang H, Lou L and Shen X: miR-145 suppresses breast cancer
cell migration by targeting FSCN-1 and inhibiting
epithelial-mesenchymal transition. Am J Transl Res. 8:3106–3114.
2016.PubMed/NCBI
|
|
11
|
Tian J, Hu X, Gao W, Zhang J, Chen M,
Zhang X, Ma J and Yuan H: Identification a novel tumor-suppressive
hsa-miR-599 regulates cells proliferation, migration and invasion
by targeting oncogenic MYC in hepatocellular carcinoma. Am J Transl
Res. 8:2575–2584. 2016.PubMed/NCBI
|
|
12
|
Joyce DP, Kerin MJ and Dwyer RM:
Exosome-encapsulated microRNAs as circulating biomarkers for breast
cancer. Int J Cancer. 139:1443–1448. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yang Z, He N and Zhou Q: Brd4 recruits
P-TEFb to chromosomes at late mitosis to promote G1 gene expression
and cell cycle progression. Mol Cell Biol. 28:967–976. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang YH, Sui YN, Yan K, Wang LS, Wang F
and Zhou JH: BRD4 promotes pancreatic ductal adenocarcinoma cell
proliferation and enhances gemcitabine resistance. Oncol Rep.
33:1699–1706. 2015.PubMed/NCBI
|
|
15
|
Segura MF, Fontanals-Cirera B,
Gaziel-Sovran A, Guijarro MV, Hanniford D, Zhang G, González-Gomez
P, Morante M, Jubierre L, Zhang W, et al: BRD4 sustains melanoma
proliferation and represents a new target for epigenetic therapy.
Cancer Res. 73:6264–6276. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Puissant A, Frumm SM, Alexe G, Bassil CF,
Qi J, Chanthery YH, Nekritz EA, Zeid R, Gustafson WC, Greninger P,
et al: Targeting MYCN in neuroblastoma by BET bromodomain
inhibition. Cancer Discov. 3:308–323. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cheng Z, Gong Y, Ma Y, Lu K, Lu X, Pierce
LA, Thompson RC, Muller S, Knapp S and Wang J: Inhibition of BET
bromodomain targets genetically diverse glioblastoma. Clin Cancer
Res. 19:1748–1759. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang YH, Sui XM, Sui YN, Zhu QW, Yan K,
Wang LS, Wang F and Zhou JH: BRD4 induces cell migration and
invasion in HCC cells through MMP-2 and MMP-9 activation mediated
by the Sonic hedgehog signaling pathway. Oncol Lett. 10:2227–2232.
2015.PubMed/NCBI
|
|
19
|
Lockwood WW, Zejnullahu K, Bradner JE and
Varmus H: Sensitivity of human lung adenocarcinoma cell lines to
targeted inhibition of BET epigenetic signaling proteins. Proc Natl
Acad Sci USA. 109:19408–19413. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Patel AJ, Liao CP, Chen Z, Liu C, Wang Y
and Le LQ: BET bromodomain inhibition triggers apoptosis of
NF1-associated malignant peripheral nerve sheath tumors through Bim
induction. Cell Rep. 6:81–92. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Andrieu G, Tran AH, Strissel KJ and Denis
GV: BRD4 regulates breast cancer dissemination through
Jagged1/Notch1 signaling. Cancer Res. 76:6555–6567. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Pritchard CC, Cheng HH and Tewari M:
MicroRNA profiling: Approaches and considerations. Nat Rev Genet.
13:358–369. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xie B, Zhang C, Kang K and Jiang S:
miR-599 inhibits vascular smooth muscle cells proliferation and
migration by targeting TGFB2. PLoS One. 10:e01415122015. View Article : Google Scholar : PubMed/NCBI
|