Introduction
Retinoblastoma is the most common type of malignant
intraocular tumor in children, and it affects ~1 in 16,000–18,000
births, with an incidence of 7,000–8,000 new cases per year
worldwide (1–3). A total of ~80% of children with
retinoblastoma are diagnosed prior to the age of 3 (4). Retinoblastoma treatment is administered
according to the International Classification of Retinoblastoma
(ICRB) (5). In developing countries,
although current laser photocoagulation, thermotherapy and
chemotherapy exhibit high survival rates, enucleation remains
inevitable for children with advanced tumors (ICRB group D, E),
including subretinal seedings and vitreous seedings (6). Therefore, combined treatments with
increased efficiency & reduced toxicity are required for
advanced retinoblastoma.
Chemotherapy serves an important function in the
management of retinoblastoma. Carboplatin is a platinum-based
chemotherapy drug that stops tumor growth by directly attacking DNA
repair (7). Since the late 1980s,
this drug has been used to treat numerous types of solid cancer,
including retinoblastoma (8). Current
systemic chemotherapy for retinoblastoma consists of 6–10 cycles of
carboplatin, vincristine and etoposide. To maximize the
effectiveness and minimize the toxicity caused by exposure to
systemic chemotherapy, local chemotherapeutic regimens can be
delivered intra-arterially, transclerally or intravitreally. In
1996, Harbour et al (9)
demonstrated that intravitreous injection of carboplatin caused
retinoblastoma tumor regression in transgenic mice; later that
year, Murray et al (10)
obtained similar results. However, the rate of tumor control with
monochemotherapy for advanced retinoblastoma in the clinic is low
(11). Additional laboratory studies
are required to combine other agents that act against tumorigenesis
with carboplatin to optimize the therapeutic schedule for advanced
retinoblastoma.
Tumorigenesis is a complex process involving
angiogenesis. In the 1970s, Folkman et al (12) revealed anti-angiogenic therapy as a
potential treatment for tumors. Compared with other angiogenic
factors, several studies have demonstrated that vascular
endothelial growth factor (VEGF) and its receptors (VEGFR) perform
central roles in a variety of cancers. For intraocular tumors,
intraocular VEGF levels were increased in the patients of uveal
melanoma and retinoblastoma (13,14). Based
on these findings, a variety of anti-angiogenic drugs, including
anti-VEGF antibodies such as bevacizumab and VEGF-receptor kinase
inhibitors, have been developed and approved for the treatment of
several types of cancer. Bevacizumab is a recombinant humanized
monoclonal antibody and is the first commercially available
angiogenesis inhibitor that blocks angiogenesis by inhibiting all
forms of VEGF-A (15).
To date, there have been limited studies on combined
chemotherapy and anti-angiogenic therapy for retinoblastoma.
Therefore, the purpose of the present study was to investigate the
effect of carboplatin (chemotherapy) in combination with
bevacizumab (anti-angiogenic therapy) on human retinoblastoma in
vitro and in vivo.
Materials and methods
Cell culture and hypoxic
treatment
The human retinoblastoma tumor Y79 cell line was
obtained from the American Tissue Culture Collection (Manassas, VA,
USA) and was cultured in RPMI-1640 with 10% fetal bovine serum
(FBS; Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA), 100
U/ml penicillin and 100 µg/ml streptomycin at 37°C in 5%
CO2 and 95% humidified air. For drug treatment, the
cells were cultured at 37°C for an additional 24, 48 or 72 h in a
humidified-atmosphere incubator (1% O2 and 5%
CO2).
Cell proliferation assay
Y79 cells were plated in 96-well plates at a density
of 4×104 cells per well with 100 µl of complete culture
medium. Following proliferation for 24 h, the cells were
independently treated with 5 concentrations (1, 12.5, 25, 50 and
100 µg/ml) of carboplatin (Bristol-Myers Squibb GmbH & Co.
kGaA, München, Germany) and 3 concentrations (0.1, 1 and 2 mg/ml)
of bevacizumab (Roche Pharmaceutical, Shanghai, China), and were
cultured at 37°C for another 24, 48 or 72 h under hypoxic
conditions; these experiments represented the monotherapy groups.
For combined therapy, 5 concentrations (1, 12.5, 25, 50 and 100
µg/ml) of carboplatin in combination with 1 or 2 mg/ml of
bevacizumab were added to cells, which were then cultured at 37°C
for 48 h under hypoxic conditions. Wells with culture medium served
as negative controls. At each time point, 10 µl of Cell Counting
Kit-8 (CCK-8) working solution (Dojindo Molecular Technologies,
Inc., Shanghai, China) was added to each well prior to additional
incubation for 2 h at 37°C. The absorbance was recorded at 450 nm
with an ELISA plate reader (Finstruments Multiskan Model 347; MTX
Lab Systems, Inc., Vienna, VA, USA). All experiments were performed
in 3 wells and were replicated at least 3 times.
Flow cytometry
The Y79 cell line was subject to an apoptosis study
fluorescein isothiocyanate (FITC) Annexin V Apoptosis Detection
kit; BD Biosciences, Franklin Lakes, NJ, USA) and cell cycle
analysis (Cycletest Plus DNA Reagent kit; BD Biosciences), these
were performed according to the manufacturer's protocol. Briefly,
Y79 cells (5×105/ml) with 1,000 µl complete RPMI-1640
were seeded in 6-well plates and were incubated at 37°C for 48 h
under hypoxic conditions with 5 concentrations (1, 12.5, 25, 50 and
100 µg/ml) of carboplatin, carboplatin plus 1 mg/ml bevacizumab, or
with no additives as a control. The samples (1×104
cells) were subsequently analyzed using flow cytometry (FACS
Calibur; BD Biosciences) according to the manufacturer's protocol
and as previously described (16).
The apoptotic rate was calculated as the percentage of early
apoptotic cells [FITC Annexin V positive and propidium iodide (PI)
negative, LR] plus late apoptotic cells (FITC Annexin V positive
and PI positive, UR). The cell cycle distribution of different
groups was analyzed. The experiments were performed in triplicate
and were repeated 3 times.
Transmission electron microscopy
(TEM)
The Y79 cells that were treated with 25 µg/ml of
carboplatin alone or in combination with 1 mg/ml of bevacizumab and
cultured at 37°C for 48 h under hypoxic conditions were fixed with
1.5% glutaraldehyde in 0.1 M PBS (pH 7.0) for 2 h at 4°C. The cells
were then post-fixed in 1% osmic acid in the same buffer,
dehydrated in ethanol and embedded in epoxy resin. Thin sections
(thickness, 70 nm) were cut and were stained with toluidine blue.
Finally, the sections were stained with uranium and lead prior to
examination using TEM (FEI; Thermo Fisher Scientific, Inc.). The
images were captured at magnifications of ×2,550 and ×8,200 and
were processed with graphics software (Adobe Photoshop CS5, Adobe
Systems, Inc., San Jose, CA, USA).
Western blot analysis
The Y79 cells treated with 3 concentrations (1, 25
and 100 µg/ml) of carboplatin alone or in combination with 1 mg/ml
of bevacizumab and cultured for 48 h under hypoxic conditions were
washed with ice-cold PBS and were then prepared using
radioimmunoprecipitation assay protein extraction buffer with a
protease inhibitor cocktail (Pierce; Thermo Fisher Scientific,
Inc.). The lysates were cleared by centrifugation at 15,000 × g for
15 min at 4°C. The supernatant was collected, and the protein
content of each lysate was measured using a bicinchoninic acid
protein assay kit (Pierce; Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol. Equal amounts (50 µg) of
protein were loaded and separated by 10% SDS-PAGE prior to being
transferred electrophoretically onto polyvinylidene difluoride
membranes (GE Healthcare Life Sciences, Chalfont, UK).
Subsequent to blocking nonspecific binding with 5%
skimmed milk, the membranes were incubated with rabbit polyclonal
antibodies against phospho-(p-) extracellular signal-regulated
kinases (ERK) 1/2 (catalog no. 4370; dilution, 1:1,000; Cell
Signaling Technology, Inc., Danvers, MA, USA), ERK1/2 (catalog no.
9102; dilution, 1:1,000; Cell Signaling Technology, Inc.),
p-protein kinase B (Akt; catalog no. 4060; dilution, 1:1,000; Cell
Signaling Technology, Inc.) or Akt (catalog no. 4685; dilution,
1:1,000; Cell Signaling Technology, Inc.) followed by incubation
with a horseradish peroxidase-conjugated goat antibody against
rabbit (catalog no. 7074; dilution, 1:5,000; Cell Signaling
Technology, Inc.). The proteins were visualized using enhanced
chemiluminescence western blot detection reagents (Pierce; Thermo
Fisher Scientific, Inc.). To ensure equal loading of the protein in
each lane, the blots were stripped and re-probed with an antibody
against β-actin (catalog no. 4087; dilution, 1:5,000; Cell
Signaling Technology, Inc.), followed by incubation with a
horseradish peroxidase-conjugated goat antibody against rabbit
(catalog no. 707; dilution, 1:5,000; Cell Signaling Technology,
Inc.). The intensity of protein bands was determined by ImageJ
(version 2.0.0; National Institutes of Health, Bethesda, MA, USA).
Western blot analyses were repeated 3 times, and similar results
were obtained.
Animals and ethics statement
A total of 40 BALB/c nude female mice (Peking
University Health Science Center, Beijing, China) that were 8 weeks
old and weighed 18–20 g were used in the in vivo study. The
left eyes (n=40) were injected for orthotopic retinoblastoma mouse
model, as described below. All mice were housed under specific
pathogen-free conditions in the Animal Facility of Peking
University Health Science Center, with controlled humidity (between
40 and 70%), at 24±1°C, under 12 h light/dark conditions, and with
food and water ad libitum.
This study adhered to The Association for Research
in Vision and Ophthalmology Statement for the Use of Animals in
Ophthalmic and Vision Research and was performed in accordance with
the guidelines provided by the Animal Care Use Committee of Peking
University.
Orthotopic retinoblastoma mouse model
and drug treatment
Y79 cell suspensions (5×105/ml) in PBS
were used for intravitreous injection. Prior to cell injection,
mice were anesthetized using intraperitoneal injections of ketamine
(50 mg/kg) and xylazine (5 mg/kg). Subsequently, 1 drop of atropine
was administered to dilate the pupil. Finally, a hamilton syringe
with 2 µl of cell suspension was inserted until the needle could be
observed in the vitreous of the left eye through the microscope.
The cell suspension was then slowly and carefully injected. The
needle was removed 2 min subsequent to the injection, and 1 drop of
erythromycin was administered to each eye. The eyes were then
examined daily for leukocoria. Once tumor growth was detected, the
mice were randomly divided into 2 groups: PBS intravitreous
injection group and 5 µg carboplatin with 1.25 µg bevacizumab
intravitreous injection group. Ocular images were captured at weeks
2, 4, 6 and 8 (n=5) post-treatment for analysis.
Histopathological observation
Mice were euthanized with an overdose of ketamine
and xylazine at different time points post-treatment, and the eyes
were removed and dissected. Paraffin-embedded sections were then
prepared from whole eyes. Serial sections (5 µm) were stained with
conventional hematoxylin and eosin (H&E) staining, and the
slides were then observed and photographed using a microscope
(Olympus Corporation, Tokyo, Japan). All micrographs were taken
under the same microscope with the same parameters (exposure time,
1 sec; resolution ratio, 1,280:960; brightness, 100%) and were
saved as TIFF files. The images were processed with graphics
software (Adobe Photoshop CS5; Adobe Systems, Inc.).
Statistical analysis
Data analysis was performed using the statistical
software Prism 5 (GraphPad Software, Inc., La Jolla, CA, USA). All
data are presented as the mean ± standard error of the mean and
have been evaluated for normality of distribution. Differences
between groups were evaluated with the Student's unpaired t-test
(2-tailed). P<0.05 was considered to indicate a statistically
significant difference.
Results
Inhibition of Y79 cell proliferation
by carboplatin and bevacizumab under hypoxic conditions
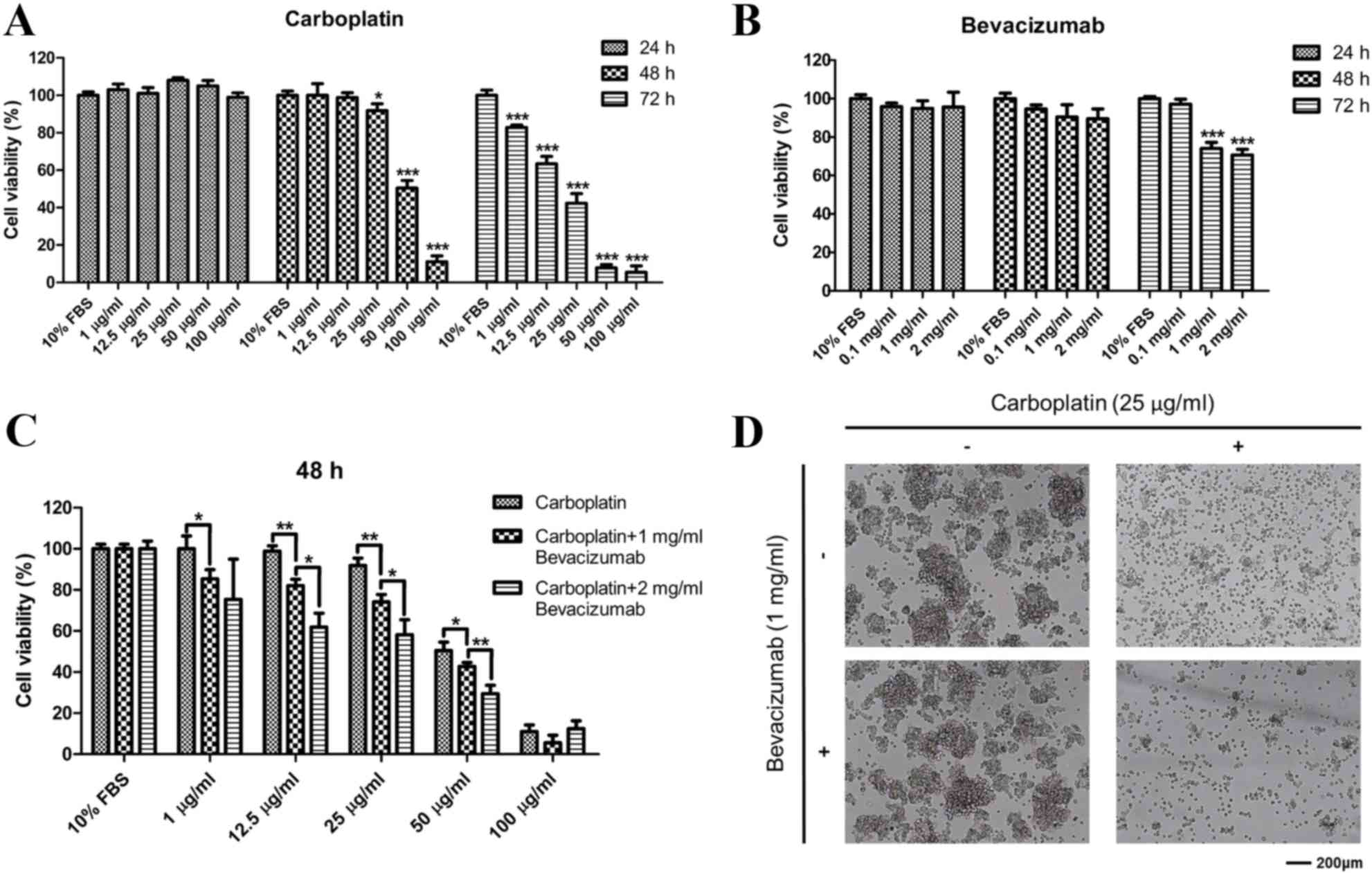
A CCK-8 measuring proliferation was used to evaluate
the inhibitory effects of carboplatin and bevacizumab on Y79 cell
proliferation in vitro under hypoxic culture conditions.
Carboplatin inhibited Y79 proliferation in a time and
dose-dependent manner (Fig. 1A,
P<0.05). Bevacizumab significantly inhibited cell proliferation
at high concentrations (≥1 mg/ml) following treatment for 72 h when
compared with the 10% FBS group (Fig.
1B, P<0.001). Since bevacizumab did not inhibit cell
proliferation at other concentrations (0.1, 1 and 2 mg/ml) at 24
and 48 h, the present study chose 1 mg/ml bevacizumab in
combination with different concentrations of carboplatin at 48 h
for the following experiments.
Inhibition of Y79 cell proliferation
by carboplatin in combination with bevacizumab under hypoxic
conditions
A total of 5 concentrations (1, 12.5, 25, 50 and 100
µg/ml) of carboplatin or carboplatin in combination with 1 or 2
mg/ml of bevacizumab were cocultured with Y79 cells for 48 h under
hypoxic conditions. As shown in Fig.
1C, the combination group exhibited higher inhibition under
hypoxic conditions in a time and dose-dependent manner (P<0.05).
Subsequent to 48 h of exposure, an increased number of cells were
swollen in the combined treatment group compared with either the
carboplatin group or the PBS group (Fig.
1D).
Combination of carboplatin and
bevacizumab induces Y79 apoptosis, cell cycle arrest and
morphological changes under hypoxic conditions
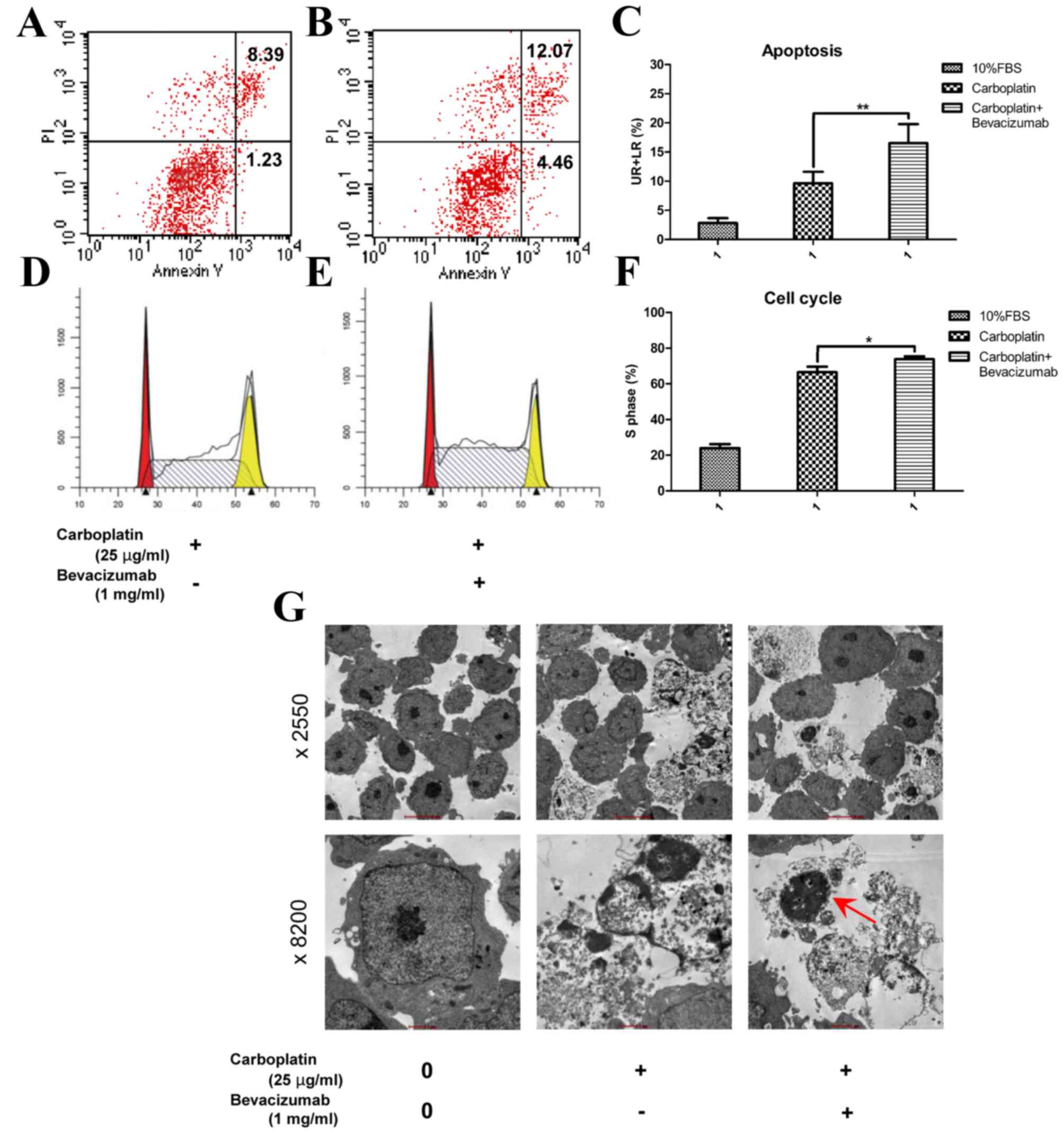
Flow cytometry tests were used to evaluate the
effects of carboplatin alone and carboplatin plus bevacizumab on
Y79 cell apoptosis (early apoptosis and late apoptosis) and on
cell-cycle arrest (S phase) under hypoxic conditions. In the
present study, as presented in Fig.
2, there was a significant difference between the carboplatin
plus bevacizumab group (Fig. 2B) and
the carboplatin-alone group (Fig. 2A)
in the apoptosis test (Fig. 2C;
P<0.01). Additionally, combination treatment (Fig. 2E) induced a greater degree of Y79 cell
cycle arrest at S phase than the control group (Fig. 2D), and the difference was significant
(Fig. 2F; P<0.05). A typical
apoptotic body could be observed in the combined group (Fig. 2G, red arrow).
Combination of carboplatin and
bevacizumab inhibited ERK 1/2 and Akt signaling pathway
phosphorylation
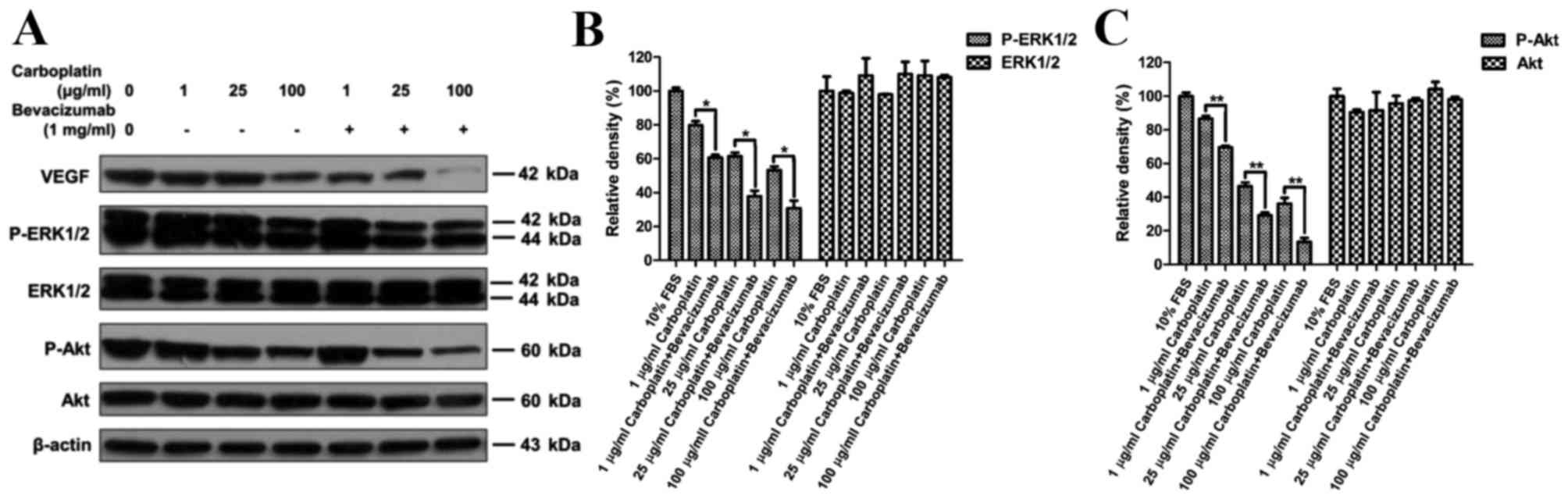
To examine the kinetics of ERK and Akt activation,
cells were exposed to 3 concentrations (1, 25 and 100 µg/ml) of
carboplatin alone or in combination with 1 mg/ml of bevacizumab for
48 h under hypoxic conditions. As shown in Fig. 3, the expression levels of
phospho-ERK1/2 and phospho-Akt decreased significantly in the
combination treatment group when compared to the carboplatin group,
no change in the total levels of ERK1/2, Akt and β-actin was
observed (Fig. 3A). Compared with the
carboplatin group, significantly decreased expression of
phospho-ERK1/2 and phospho-Akt in the combination treatment was
also revealed by quantitative analysis of P-ERK1/2 and ERK1/2
expression relative to the controls (Fig.
3B; P<0.05), and quantitative analysis of P-Akt and Akt
expression relative to the controls (Fig.
3B; P<0.01). These observations indicate that blockage of
ERK1/2 and Akt activation partly contributed to the death of Y79
cells following treatment.
Combination of carboplatin and
bevacizumab inhibits intravitreous tumorigenesis in mice
Following the present in vitro investigation
of the antitumor effect of the combination of carboplatin and
bevacizumab on Y79 cells and its mechanism, cells were injected
into the vitreous cavity of BALB/c nude mice as an orthotopic
retinoblastoma model. The present study used 2 groups of mice: PBS
and the combination treatment. Drugs were administered every 2
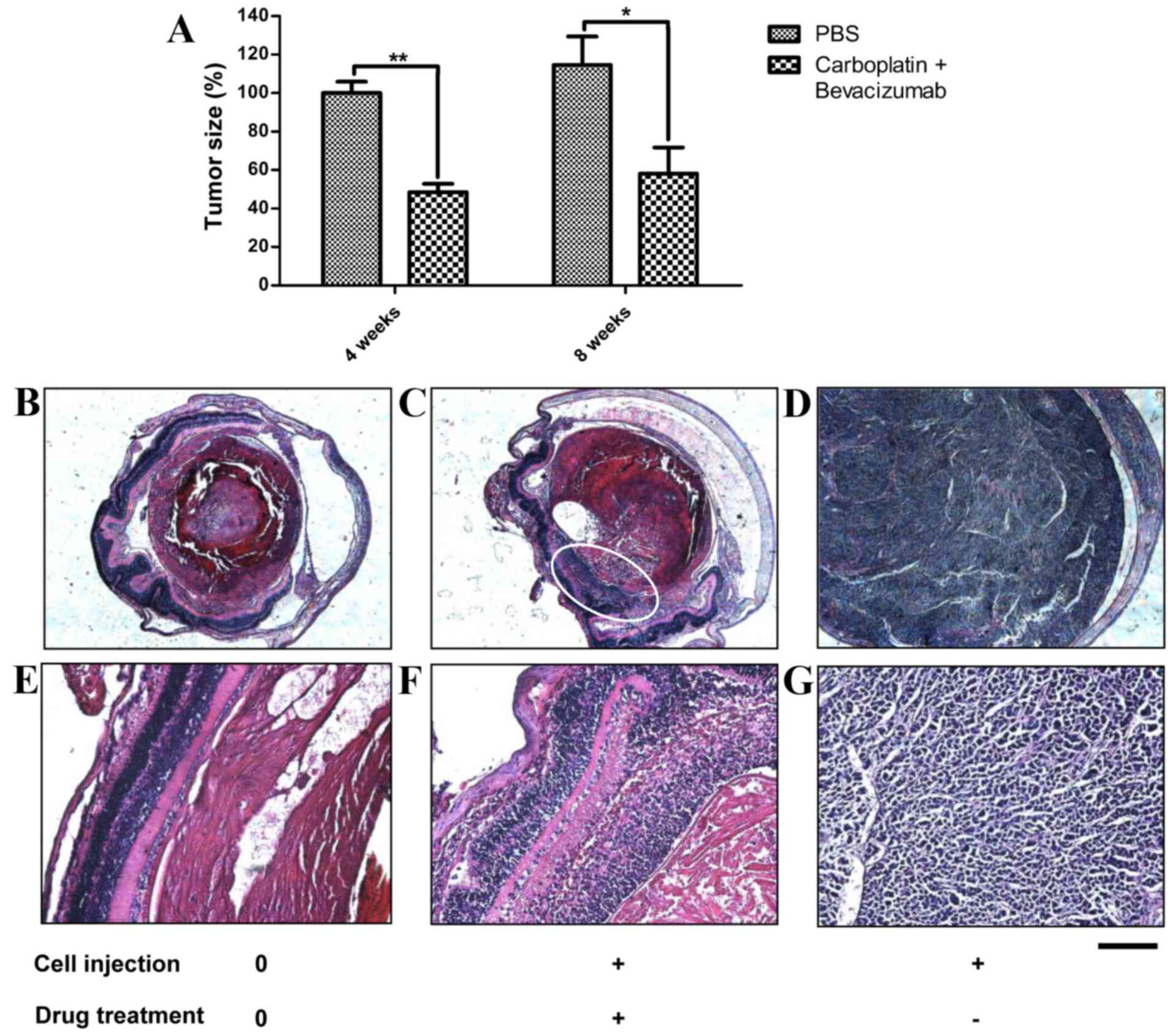
weeks, and the longest diameter of the PBS-treated tumor (4 weeks:
8.9±0.34 mm; 8 weeks: 10.2±0.85 mm) and drug-treated tumor (4
weeks: 4.3±0.25 mm; 8 weeks: 5.2±0.78 mm) were determined. Similar
to the in vitro experiments, the combination of carboplatin
and bevacizumab substantially suppressed tumor volume (Fig. 4A; P<0.05), whereas the average
tumor volume in the PBS-treated eyes increased gradually with age
(Fig. 4A).
Histological examination
To identify the tumor growth in the PBS-treated and
drug-treated eyes, H&E staining of orthotopic retinoblastoma
model was performed. Compared with normal eyes at ×4 magnification
(Fig. 4B) and ×20 magnification
(Fig. 4E), the histological studies
suggested that the drug-treated eyes at ×4 magnification (Fig. 4C) and ×20 magnification (Fig. 4F) exhibited a decreased tumor size;
whereas the untreated eyes at ×4 magnification (Fig. 4D) and ×20 magnification (Fig. 4G) contained numerous tumor cells with
deeply stained nuclei and condensed cytoplasm.
Discussion
Retinoblastoma, the most frequent intraocular tumor
in children, is treated according to its classification and
severity. For children with advanced tumors, subretinal and
vitreous seeding makes enucleation common for survival; therefore
novel combined treatments are required.
Due to the limited number of retinoblastoma cases,
cell lines and animal models have performed an important role in
understanding retinoblastoma tumorigenesis and developing
treatments. For advanced retinoblastoma, once cells are seeded into
the vitreous cavity, they are kept in a suspended state. Therefore,
the present study utilized suspended retinoblastoma Y79 cells. The
Y79 cell line was the first retinoblastoma cell line and was
derived from the tumor of a 2.5-year-old Caucasian female with a
strong maternal history of retinoblastoma. It was also the first
cell line transplanted into the anterior chamber of the nude mouse
eye to establish a xenograft model of human retinoblastoma
(17).
Angiogenesis is required for tumor growth and
expansion. Previous studies have demonstrated that VEGF levels are
significantly elevated in patients with retinoblastoma (18–20).
Bevacizumab, an anti-VEGF agent, has been proven to affect the
growth and differentiation of retinoblastoma cells in vitro
(21,22). In advanced retinoblastoma, the
disseminated tumor cells are exposed to a hypoxic and nutrient-poor
environment, which stimulates the tumor cells to secrete more
angiogenic factors. Therefore, it was hypothesized that anti-VEGF
therapy may have an important therapeutic role in advanced
retinoblastoma. The present study treated Y79 cells (cultured under
hypoxic conditions in vitro to mimic the advanced tumor
microenvironment) with carboplatin plus bevacizumab. It was found
that the carboplatin plus bevacizumab group inhibited Y79 cell
proliferation and caused morphological changes. To further
understand this inhibitory effect, the present study used flow
cytometry to detect cellular apoptosis and analyze the cell cycle
in the carboplatin-alone and combined-treatment groups. It was
revealed that combination treatment induced greater cellular
apoptosis and increased frequent cell-cycle arrest compared with
monotherapy. These experimental results indicate that the
carboplatin-and-bevacizumab combination results in a strengthened
antitumor effect against human retinoblastoma cells in
vitro.
Currently, there are limited studies on the
mechanism of carboplatin and bevacizumab combination therapy. A
previous study identified that VEGF-VEGFR2 is a key mediator for
tumor angiogenesis and that VEGFR2 is expressed in Y79 cells
(22). The present study analyzed
common intracellular signaling pathways involved in VEGF-VEGFR2
signaling. Previous studies have demonstrated that the expression
of phosphorylated ERK1/2 is necessary for cellular proliferation,
and activation of the mitogen-activated protein kinase (MAPK)/ERK
pathway is one of the hallmarks of cancer cells (23,24). The
PI3K/Akt pathway also serves a central role in cell survival and
proliferation and is aberrantly activated in numerous types of
cancer (25). For these reasons, the
MAPK/ERK and PI3K/Akt pathways are potential therapeutic targets
for retinoblastoma. The present study revealed that the combined
treatment reduced the increases of the expression of VEGF,
phosphorylated ERK1/2 and Akt in the cells in vitro.
Compared to in vitro experiments, preclinical
models have great advantages for tumor environment simulation and
therapeutic evaluation. For additional in vivo study, the
present study established an orthotopic retinoblastoma model by
injecting cells into the vitreous cavity to mimic the advanced
tumor microenvironment. The dose of drugs used in the present study
was identical to that used for human patients, in association with
body size. In 1993, Kim et al (26) confirmed that blood vessels are
important for tumor growth and metastasis and that anti-VEGF
treatment could inhibit the growth of human rhabdomyosarcoma,
glioblastoma multiforme and leiomyosarcoma in nude mice models.
Similarly, other studies have demonstrated that intraperitoneal
injections of the anti-VEGF monoclonal antibody inhibits tumor
growth in tumor-bearing nude mice and that VEGF-VEGFR2 signals
perform a dominant role in tumor angiogenesis (27). The present results are consistent with
these observations; in the present intravitreous model, carboplatin
and bevacizumab combination therapy significantly inhibited the
expression of VEGF and suppressed tumor growth in a time-dependent
manner when compared with the control therapy.
Intraocular drug injection may cause secondary
vitreous seeding, although the incidence is low. However, local
application may significantly maximize the effectiveness of
treatment and minimize the toxicity caused by exposure to systemic
chemotherapy. Additional studies are required to optimize local
therapy for retinoblastoma.
In summary, the present study was the first to
confirm that a combination of carboplatin and bevacizumab results
in a strengthened antitumor effect against human advanced
retinoblastoma in vitro and in vivo and that the
effect likely occurs via inhibition of the PI3K/Akt and MAPK/ERK
pathways. Therefore, the present findings demonstrate that
anti-angiogenic therapy may perform an adjuvant role in the
treatment of advanced retinoblastoma.
Acknowledgements
The present study was supported by The Peking
University People's Hospital Research and Development Fund (grant
no. RDC-2014-24) and the National Basic Research Program of China
(973 Program; grant no. 2011CB510200).
References
|
1
|
Broaddus E, Topham A and Singh AD:
Incidence of retinoblastoma in the USA: 1975–2004. Br J Ophthalmol.
93:21–23. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kivelä T: The epidemiological challenge of
the most frequent eye cancer: Retinoblastoma, an issue of birth and
death. Br J Ophthalmol. 93:1129–1131. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Seregard S, Lundell G, Svedberg H and
Kivelä T: Incidence of retinoblastoma from 1958 to 1998 in Northern
Europe: Advantages of birth cohort analysis. Ophthalmology.
111:1228–1232. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chintagumpala M, Chevez-Barrios P, Paysse
EA, Plon SE and Hurwitz R: Retinoblastoma: Review of current
management. Oncologist. 12:1237–1246. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Shields CL, Mashayekhi A, Au AK, Czyz C,
Leahey A, Meadows AT and Shields JA: The International
classification of retinoblastoma predicts chemoreduction success.
Ophthalmology. 113:2276–2280. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Balmer A, Zografos L and Munier F:
Diagnosis and current management of retinoblastoma. Oncogene.
25:5341–5349. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wheate NJ, Walker S, Craig GE and Oun R:
The status of platinum anticancer drugs in the clinic and in
clinical trials. Dalton Trans. 39:8113–8127. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Shields CL, Kaliki S, Rojanaporn D,
Al-Dahmash S, Bianciotto CG and Shields JA: Intravenous and
intra-arterial chemotherapy for retinoblastoma: What have we
learned? Curr Opin Ophthalmol. 23:202–209. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Harbour JW, Murray TG, Hamasaki D,
Cicciarelli N, Hernández E, Smith B, Windle J and O'Brien JM: Local
carboplatin therapy in transgenic murine retinoblastoma. Invest
Ophthalmol Vis Sci. 37:1892–1898. 1996.PubMed/NCBI
|
|
10
|
Murray TG, Roth DB, O'Brien JM, Feuer W,
Cicciarelli N, Markoe AM, Hernández E, Smith BJ and Windle JJ:
Local carboplatin and radiation therapy in the treatment of murine
transgenic retinoblastoma. Arch Ophthalmol. 114:1385–1389. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gombos DS, Kelly A, Coen PG, Kingston JE
and Hungerford JL: Retinoblastoma treated with primary chemotherapy
alone: The significance of tumour size, location, and age. Br J
Ophthalmol. 86:80–83. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Folkman J: Tumor angiogenesis: Therapeutic
implications. N Engl J Med. 285:1182–1186. 1971. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Shibuya M: VEGF-VEGFR signals in health
and disease. Biomol Ther (Seoul). 22:1–9. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Missotten GS, Schlingemann RO and Jager
MJ: Angiogenesis and vascular endothelial growth factors in
intraocular tumors. Dev Ophthalmol. 46:123–132. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bevacizumab, . Anti-VEGF monoclonal
antibody, avastin, rhumab-VEGF. Drugs R D. 3:28–30. 2002.PubMed/NCBI
|
|
16
|
Bai YJ, Huang LZ, Xu XL, Du W, Zhou AY, Yu
WZ and Li XX: Polyethylene glycol-modified pigment
epithelial-derived factor: New prospects for treatment of retinal
neovascularization. J Pharmacol Exp Ther. 342:131–139. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Reid TW, Albert DM, Rabson AS, Russell P,
Craft J, Chu EW, Tralka TS and Wilcox JL: Characteristics of an
established cell line of retinoblastoma. J Natl Cancer Inst.
53:347–360. 1974. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Areán C, Orellana ME, Abourbih D, Abreu C,
Pifano I and Burnier MN Jr: Expression of vascular endothelial
growth factor in retinoblastoma. Arch Ophthalmol. 128:223–229.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Youssef NS and Said AM:
Immunohistochemical expression of CD117 and vascular endothelial
growth factor in retinoblastoma: Possible targets of new therapies.
Int J Clin Exp Pathol. 7:5725–5737. 2014.PubMed/NCBI
|
|
20
|
Cheng Y, Zheng S, Pan CT, Yuan M, Chang L,
Yao Y, Zhao M and Liang J: Analysis of aqueous humor concentrations
of cytokines in retinoblastoma. PLoS One. 12:e01773372017.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lee SY, Kim DK, Cho JH, Koh JY and Yoon
YH: Inhibitory effect of bevacizumab on the angiogenesis and growth
of retinoblastoma. Arch Ophthalmol. 126:953–958. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Heo JW and Kim JH, Cho CS, Jun HO, Kim DH,
Yu YS and Kim JH: Inhibitory activity of bevacizumab to
differentiation of retinoblastoma cells. PloS one. 7:e334562012.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shilo A, Ben Hur V, Denichenko P, Stein I,
Pikarsky E, Rauch J, Kolch W, Zender L and Karni R: Splicing factor
hnRNP A2 activates the Ras-MAPK-ERK pathway by controlling A-Raf
splicing in hepatocellular carcinoma development. RNA. 20:505–515.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hu Y, Yang H, Lu XQ, Xu F, Li J and Qian
J: ARHI suppresses pancreatic cancer by regulating MAPK/ERK 1/2
pathway. Pancreas. 44:342–343. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Georgakis GV and Younes A: From Rapa Nui
to rapamycin: Targeting PI3K/Akt/mTOR for cancer therapy. Expert
Rev Anticancer Ther. 6:131–140. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kim KJ, Li B, Winer J, Armanini M, Gillett
N, Phillips HS and Ferrara N: Inhibition of vascular endothelial
growth factor-induced angiogenesis suppresses tumour growth in
vivo. Nature. 362:841–844. 1993. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Mordenti J, Thomsen K, Licko V, Chen H,
Meng YG and Ferrara N: Efficacy and concentration-response of
murine anti-VEGF monoclonal antibody in tumor-bearing mice and
extrapolation to humans. Toxicol Pathol. 27:14–21. 1999. View Article : Google Scholar : PubMed/NCBI
|