Introduction
Prostate cancer is the most common form of cancer in
men. It accounts for 10% of malignant tumours worldwide and 13% of
male cancer deaths in the UK. It is most frequently seen in older
men, with 80% of cases being diagnosed in the over 65 years
(1). It is estimated that
approximately 70% of men will develop some form of prostate cancer
but the majority of cases will not be clinically relevant (2). Prostate cancer is not necessarily
lethal; it is a multifocal and heterogeneous cancer with a wide
range of outcomes. Its heterogeneity provides a real challenge for
accurate prognosis and making appropriate treatment decisions
(3). The difficulty lies in
distinguishing between indolent and aggressive forms of the
disease. The current non-invasive diagnostic test of choice is the
PSA serum test. However, raised PSA levels can be due to factors
other than prostate cancer such as benign prostatic hyperplasia and
inflammation (4,5).
Gleason grading on histopathological examination is
currently the gold standard prognostic test; however it cannot
always give a correct prognosis. Inconsistencies arise due to
differences in sampling procedures, and morphologically similar
cancers can behave very differently. Approximately 70–80% of
Gleason 6 and 20% of Gleason 7 tumours may be non-aggressive and
not require intervention for 15 years or more (6); conversely, an undetected, aggressive
cancer may become lethal within 2–12 years (7). Aggressive prostate cancer can be treated
successfully if caught in early, organ-confined stages. Therefore
it is imperative to develop a method able to discern indolent cases
not requiring immediate intervention from the aggressive ones which
do. Treatment can have a detrimental effect on the patients'
quality of life by causing urinary and sexual dysfunction (8). Furthermore, approximately 30% of
patients suffer from disease recurrence with metastases subsequent
to radical prostatectomy (9).
Prostatic intraepithelial neoplasia (PIN) lesions
are benign alterations thought to appear approximately 10 years
before the development of prostatic carcinoma. PIN is often
unidentified as it does not produce high levels of PSA and can only
be detected by biopsy (3). A marker
which could easily detect PIN would be extremely useful in both
diagnostic and prognostic testing. Much attention has recently
focused on a gene fusion, TMPRSS2:ETS-related gene
(ERG), that is frequently found in aggressive prostate
cancer. The TMPRRSS2 portion of the fusion contains an
androgen-responsive promoter which drives the aberrant expression
of the oncogenic transcription factor ERG (10). ERG is involved in homeostasis,
survival, differentiation, angiogenesis and vasculogenesis
(11).
Phosphatase and tensin homologue (PTEN) is
one of the most studied tumour suppressor genes that influences a
wide range of cellular processes including survival, proliferation,
adhesion, migration, metabolism and differentiation. Loss of
functional PTEN protein accelerates cancer by allowing the PI3K/AKT
pathway to be constitutively switched on, promoting
epithelial-mesenchymal transition (EMT) and metastasis (3,12).
PTEN is lost or mutated in 50–80% of primary prostate cancer
(but not in all cases). Its loss is involved in tumour initiation,
is associated with highly aggressive and metastatic cancer,
predicts poor clinical outcome (13),
and is linked with progression to androgen-independence and
biochemical recurrence (14). Tumours
in which the loss of PTEN protein is observed at biopsy are more
likely to have higher Gleason scores (15). Thus both ERG and PTEN
are especially important players in prostate cancer; but the extent
to which they might interact is not yet clear (16,17). Given
the presence several potential ERG binding sites in the PTEN
promoter, we sought to determine whether or not the transcription
factor ERG might regulate PTEN expression directly.
Materials and methods
Cell culture
The PC3 and LNCaP prostate cancer cell lines, and
the PNT2 normal prostatic epithelium cell lines were maintained in
RPMI-1640 and the DU145 and VCaP prostate cancer cell lines were
maintained in Dulbecco's modified Eagle's medium (DMEM;
Gibco-Invitrogen, Paisley, UK). All media were supplemented with 2
mM L-glutamine + 10% (v/v) Donor Bovine Serum (Sigma-Aldrich,
Poole, UK). The PC3 and VCaP cell lines were obtained from the
Health Protection Agency (HPA)-European Collection of Cell Cultures
(ECACC, Salisbury, UK). The DU145, LNCaP and PNT2 cell lines were
kindly provided by Professor Jeff Holly's group (Department of
Clinical Science at North Bristol, University of Bristol, Bristol,
UK).
RNA extraction and cDNA synthesis
RNA was extracted using the Isolate RNA mini kit
(BioLine, London, UK) according to the manufacturer's protocols.
RNA was quantified using a Nanodrop 1000 (Thermo Fisher Scientific,
Inc., Waltham, MA, USA). A total of 500 ng of RNA was reverse
transcribed using the cDNA Synthesis kit (BioLine) spiked with 0.2
µg of Arabidopsis thaliana RuBisCO RNA exogenous
control.
Quantitative polymerase chain reaction
(qPCR)
qPCR reactions were set up using BioLine's SensiFAST
SYBR Hi-ROX kit consisting of master mix, primers (0.25 nM each),
cDNA (6.25 ng) and run on an ABI (Applied Biosystems; Thermo Fisher
Scientific, Inc.) 7300 qPCR thermal cycler for 95°C for 10 min,
followed by 95°C for 15 sec and 60°C for 1 min for 40 cycles.
Primers were designed using FastPCR software (PrimerDigital Ltd.,
Helsinki, Finland). Data was analysed using the relative standard
curve method. For each experiment a standard curve was generated
for both the gene of interest and the RuBisCO control. Reverse
transcribed control cDNA (including the RuBisCO spike) was serially
diluted over seven points and assigned arbitrary values. These
values were then converted to log base 10 and plotted against the
Ct data points for the target gene or RuBisCO to generate a line
equation y=mx+c. To find the relative log values (x) the following
equation was used: x=Ct-c/m. The antilog was taken to reach the
original relative values. To calculate the relative abundance,
target gene values were normalised to their corresponding RuBisco
values.
Protein extraction and
quantitation
Standard RIPA cell lysis buffer (20 mM Tris-HCl pH
7.4, with 150 mM NaCl, 0.1% SDS, 1% Triton X-100, 1% deoxycholate,
5 mM EDTA) plus protease inhibitors (Pierce A32953; Pierce; Thermo
Fisher Scientific, Inc.) was added to the cells and left to
incubate on ice for 15 min. Wells were then scraped and the lysate
homogenised by aspiration using a needle and syringe. Cells were
pelleted at 14,000 rpm for 10 min at 4°C in a microcentrifuge.
Clarified supernatant was then transferred to a fresh tube and
frozen at −80°C. Protein quantitation was performed using the
Pierce BCA (bicinchoninic acid) assay and a Nanodrop 1000 (Thermo
Fisher Scientific, Inc.).
Western blotting
SDS-PAGE was performed according to standard
procedures (10% acrylamide gels). Immunodetection was undertaken
with 30 µg of protein lysate using the chemiluminescent Luminata
Forte kit (Merck Millipore, Watford, UK). The ERG primary antibody
(rabbit polyclonal, ERG-1/2/3 (C-20) antibody sc-353; Santa Cruz
Biotechnology, Inc., Santa Cruz, CA, USA) was used at a dilution of
1:500. The PTEN antibody (mouse monoclonal 26H9; Cell Signaling
Technology, Inc., Danvers, MA, USA) was used at a dilution 1:1,000.
The secondary horse-radish peroxidise (HRP)-conjugated anti-rabbit
IgG antibody was used at a 1:6,000 dilution. For a loading control
a GAPDH primary antibody [rabbit polyclonal, GAPDH (sc-25778; Santa
Cruz Biotechnology, Inc.) was used at a 1:6,000 dilution.
Immunoblots were developed and imaged using an Amersham Imager 600
(GE Healthcare, Buckinghamshire, UK) or LI-COR Odyssey Fc gel-doc
system. Densitometric analysis of the blots was performed using
ImageJ software.
Two-step chromatin
immunoprecipitation
Putative ETS transcription factor binding sites
within the PTEN promoter were determined by searching for
GGAA or TTCC sequences within the promoter's nucleotide sequence
(NCBI Accession no. AF067844.1). Primers for each gene were
designed using the FastPCR programme (PrimerDigital Ltd.).
Chromatin immunoprecipitation was performed on extracts derived
from the ERG expressing VCaP cell line; 6×106 cells were
seeded into a 100 mm tissue culture dish and left to adhere for 72
h. Chromatin immunoprecipitation was then carried out using the
Champion ChIP Assay kit (SABiosciences; Qiagen, Frederick, MD, USA)
following the manufacturer's recommended protocol but with an
additional protein-protein cross-linking step before formaldehyde
fixation. Cells were fixed in 2 mM disuccinimidyl glutarate (DSG)
with 1 mM MgCl2 in PBS at room temperature for 45 min.
Cells were then washed in PBS and fixed in 1% formaldehyde + 1 mM
MgCl2 in PBS (pH 8) and incubated for 15 min at room
temperature. Sonication was performed using a MSE Soniprep 150 set
at 7 amplitude microns with 4 cycles of 15 sec on, 30 sec off. 4 µg
of ERG antibody (ERG-1/2/3 (C-20): sc-353 for ChIP; Santa Cruz
Biotechnology, Inc.), RNA Pol II antibody (positive control;
Sigma-Aldrich) and mouse IgG (negative control; Sigma-Aldrich) were
used in the immunoprecipitations. DNA was extracted from the
immunoprecipitate using the GenElute Mammalian Genomic DNA
Purification kit (Sigma-Aldrich) and amplified by SYBR-Green qPCR.
Primers corresponding to the target promoters were as follows (all
5′ to 3′): PTEN forward tcaacggctatgtgttcacg, and reverse
gtcttagcacaaagagcaacctgc (163 bp amplicon); IGFBP2 forward
tgctgctactgggcgcgagt, and reverse acaagtgccctcgcccatgaccag (329
bp). Data was analysed using the % input method. Firstly, input Ct
was adjusted to 100% (Ct input-6.64). Results from
immunoprecipitated samples were analysed using the following
calculation: 100*2^ [adjusted input-Ct (IP)]. Fold difference was
calculated against the negative control (mouse IgG).
Knockdown of ERG through
splice-switching oligonucleotides
Splice-switching oligonucleotides (Vivo-Morpholinos)
were designed and provided by Gene Tools, LLC (Philomath, OR, USA).
A standard morpholino control (5′-CCTCTTACCTCAGTTACAATTTATA-3′) and
two Vivo-Morpholinos targeting the 3′ and 5′ splice sites of exon 4
of ERG pre-mRNA were used (E43′ and E45′, respectively,
sequences available on request). A total of 700,000 VCaP (ATCC
CRL-2876) cells were seeded into compartments of a 6-well plate and
cultured at 37°C for 72 h. Media (DMEM with 10% FBS) was then
replaced with fresh media containing 0.006% endoporter delivery
agent (Gene Tools, LLC) and one of the following: 6 µM standard
morpholino control; 6 µM E45′ Vivo-Morpholinos; 6 µM E43′
Vivo-Morpholinos. After 72 h of Morpholino treatment cells were
lysed in RIPA buffer (20 mM Tris-HCl pH 7.4, with 150 mM NaCl, 0.1%
SDS, 1% Triton X-100, 1% deoxycholate, 5 mM EDTA) plus protease
inhibitors (Pierce A32953; Pierce; Thermo Fisher Scientific,
Inc.).
Over-expression of ERG
A pCMV-SPORT6 plasmid containing the full length
cDNA clone of ERG variant 1 (accession no: BC040168) in DH10B TonA
cells was purchased from Open Biosystems (Thermo Fisher Scientific,
Inc., Ashford, UK). Cells were revived overnight in 5 ml of
lysogeny broth with 100 µg/ml ampicillin (Sigma-Aldrich). Plasmids
were then purified using the PureYield Plasmid Miniprep Sytem
(Promega Corp., Madison, WI, USA) according to the manufacturer's
instructions. The day before transfection 1.0×106 cells
per well were seeded into a 6-well plate. After 24 h the cells were
starved for two h in Optimem. Cells were then transfected with 2 µg
pCMV-SPORT6-ERG DNA with transfection reagent (Fugene HD; Promega
Corp.) added at a ratio of 1:3 (DNA: reagent). Cells were incubated
for four h and then refreshed in complete media. Cells were
transfected for up to 72 h, after which cells were subjected to
either RNA or protein extraction.
Construction of transcription reporter
plasmids
The promoter constructs for IGF1,
IGFBP2 and PTEN were made as follows. DNA containing
the upstream promoter sequences of IGF1, IGFBP2 and
PTEN was amplified from human cheek cell genomic DNA by PCR
using KOD Hot Start DNA polymerase (Novagen; Merck Biosciences, Bad
Soden, Germany) supplemented with 5% (v/v) DMSO final volume.
Primers were designed to introduce KpnI restriction sites at
the 5′ end of each amplified fragment. The PCR product was cut with
KpnI/NcoI, and ligated into KpnI/NcoI
digested luciferase-expressing pGL3 plasmid (Promega Corp.).
Plasmids were transformed into E. coli (Topo cloning kit;
Invitrogen; Thermo Fisher Scientific, Inc.), amplified and
purified. The promoter sequences were verified by restriction
analysis and sequencing (DNA Sequencing and Services; University of
Dundee, Dundee, Scotland).
Dual-luciferase transcription
assay
DU145 and VCaP cells were transfected using the
method previously described (overexpression of ERG).
Promoter constructs were transfected at 400 ng per well in a
12-well plate along with 40 ng of pRLTk Renilla-expressing plasmid
as an internal control or 400 ng promoter + 40 ng Renilla +
0–1000 ng pCMVSport6-ERG. Untransfected cells were used as a
control for background fluorescence. The dual-luciferase assay kit
(Promega Corp.) was used according to the manufacturer's
instructions and luminescence measured using a Centro XS LB 960
Microplate Luminometer and Microwin2000 software (BERTHOLD
TECHNOLOGIES GmbH & Co. KG, Bad Wildbad, Germany). Read-outs
for untreated cells were subtracted from query sample readings to
remove background noise. Query samples were normalised against
Renilla outputs, normalised query results were compared to control
samples (promoter construct only). Results were calculated as fold
differences.
siRNA-mediated ERG knockdown combined
with DLR assay
At 72 h before transfection VCaP (ATCC CRL-2876)
cells were seeded at 1.0×106 cells per well in a 6-well
plate. On the day of transfection, cells were starved in
reduced-serum medium (Optimem; Invitrogen; Thermo Fisher
Scientific, Inc.) for 2 h, followed by transfection with 100 nM
ERG-targeting siRNA (Qiagen) or non-targeting siRNA
(allstars negative control, Qiagen). Cells were incubated for four
h before removal of the transfection media and replacement with
complete media. Cells were transfected for up to 120 h, then
subjected to dual-luciferase assays, as described above.
Results and Discussion
ERG is a member of the ETS family of transcription
factors that share a DNA-binding domain termed the ETS-binding
domain (EBD). The EBD is an 85 amino-acid domain that forms a
winged helix-turn-helix motif that binds to DNA sequences that
contain a core GGAA/T sequence (18).
ERG has been shown to bind to this core sequence (19).
The cell line VCaP, established from a vertebral
metastasis, retains PTEN expression and is positive for the
TMPRSS2:ERG fusion (20). We
first confirmed that ERG is expressed exclusively in VCaP cells and
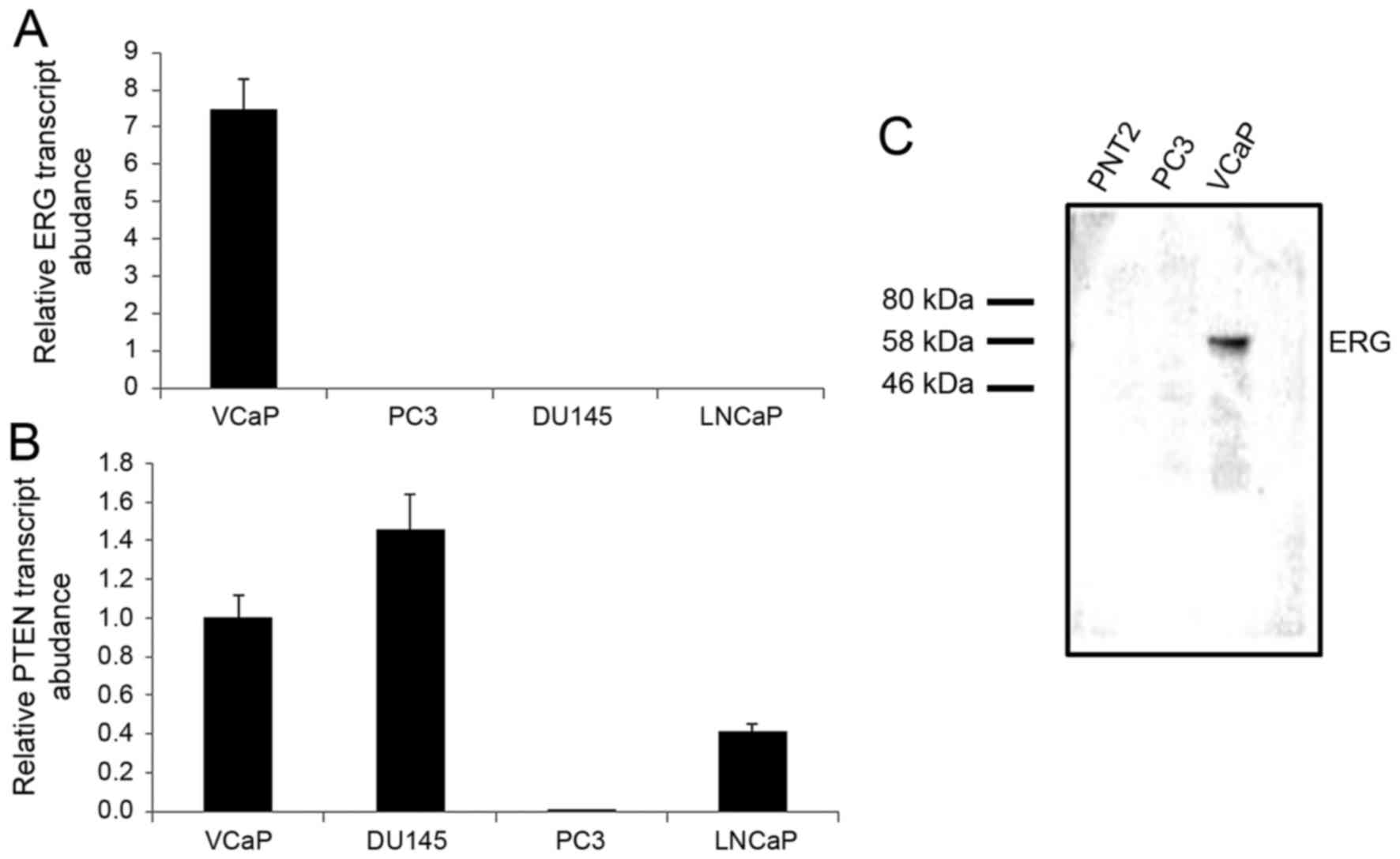
that PTEN is expressed in VCaP, DU145 and LNCaP cells (Fig. 1). We examined the PTEN promoter
and identified several clusters of GGAA sequences ranging from
1,300 bp upstream to 500 downstream of the transcription start site
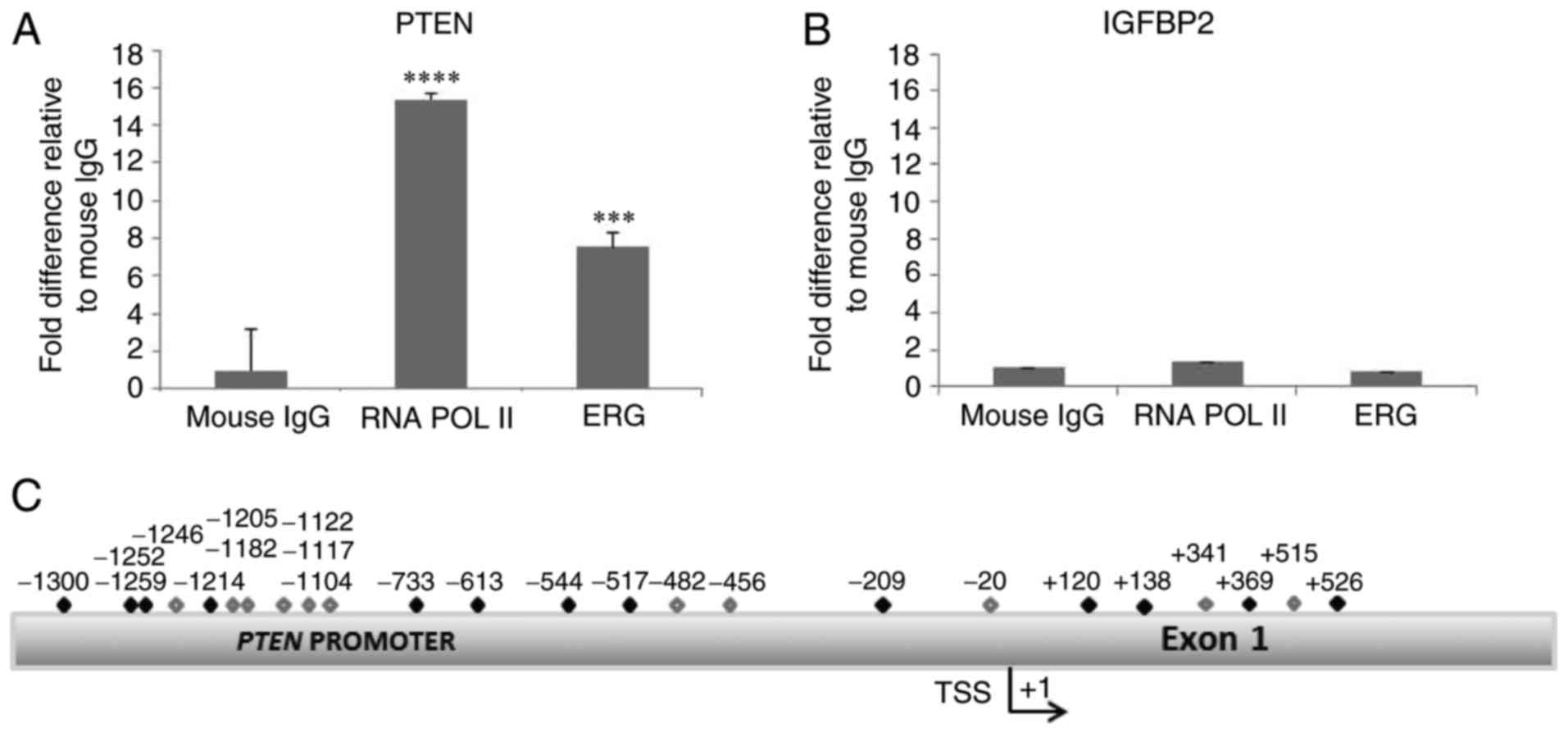
(Fig. 2C). We performed a ChIP
(chromatin immunoprecipitation) assay using extracts from VCaP
cells and observed that ERG interacts with the PTEN promoter
(Fig. 2). As a positive control an
antibody against RNA polymerase II co-precipitated with the
PTEN promoter (as PTEN is expressed in VCaP cells).
We also looked at whether the ERG antibody could co-precipitate
with another promoter that contains putative ETS binding sites, and
examined IGFBP2. Only the PTEN promoter
co-precipitated with ERG.
Having obtained evidence that ERG interacts with the
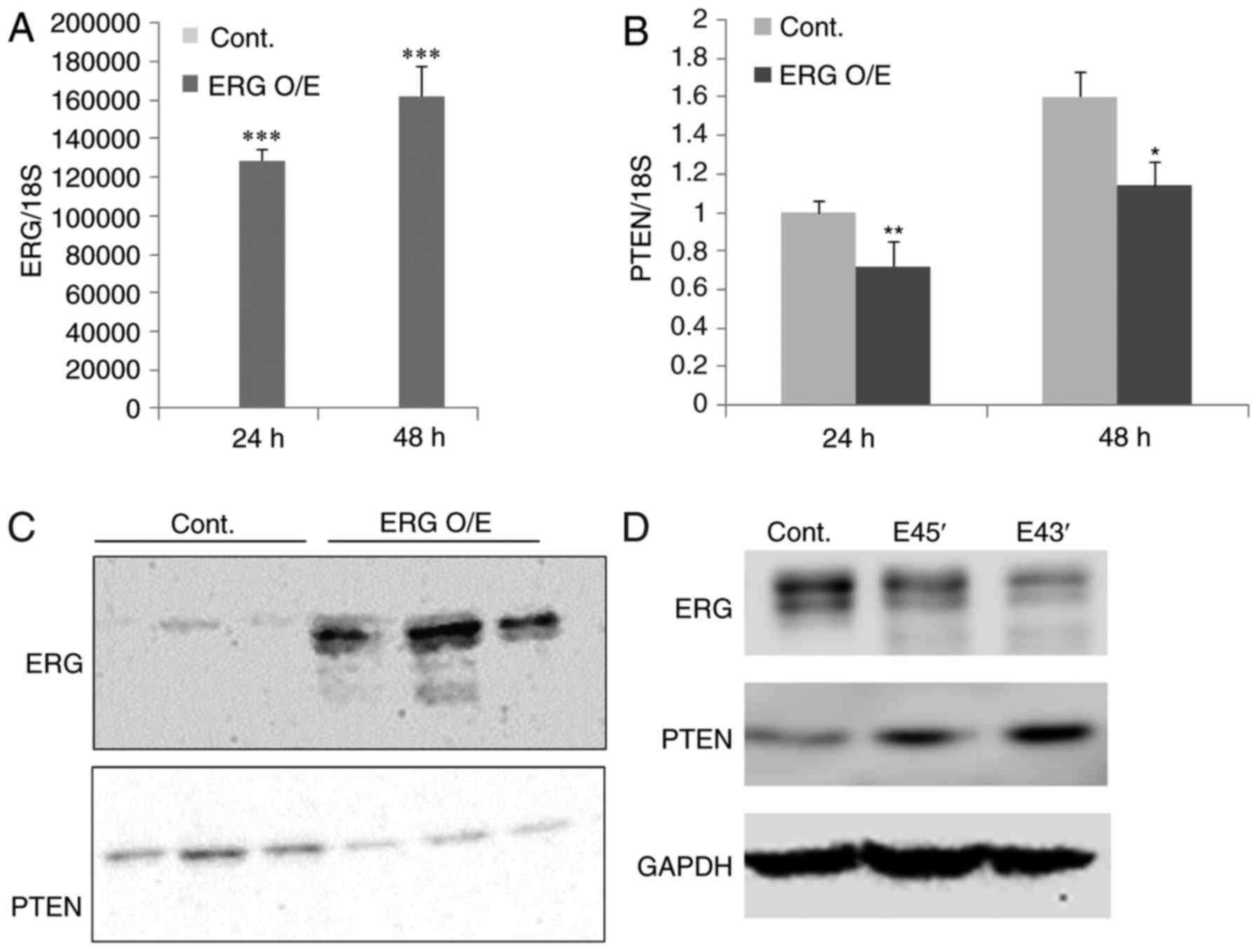
PTEN promoter, we transfected DU145 prostate cancer cells
(ERG negative and PTEN positive) with a plasmid that expresses full
length ERG. After 24–48 h we observed a significant increase in ERG
expression; this was confirmed by qPCR and western blotting
(Fig. 3). At 48 h post-transfection
we observed a significant reduction in PTEN expression both at the
RNA and protein level in cells transfected with the ERG expressing
plasmid compared to the control. Next we determined whether the
knockdown of ERG resulted in an upregulation of PTEN. To knockdown
ERG in VCaP cells we used a splice switching oligonucleotide (SSO)
approach. Morpholino SSOs were generated against both the 5′ and 3′
splice sites of ERG's exon 4. Transfection of the SSOs results in
exon 4 skipping (data not shown). Exon 4 skipping creates a
premature stop codon which leads to nonsense mediated decay and a
resulting drop in ERG protein. Reduction of ERG protein achieved
with the exon 4 SSOs resulted in a clear increase of PTEN
protein.
Taken together these results clearly suggested that
ERG transcriptionally represses PTEN. To obtain further
evidence to confirm this we fused the PTEN promoter to a
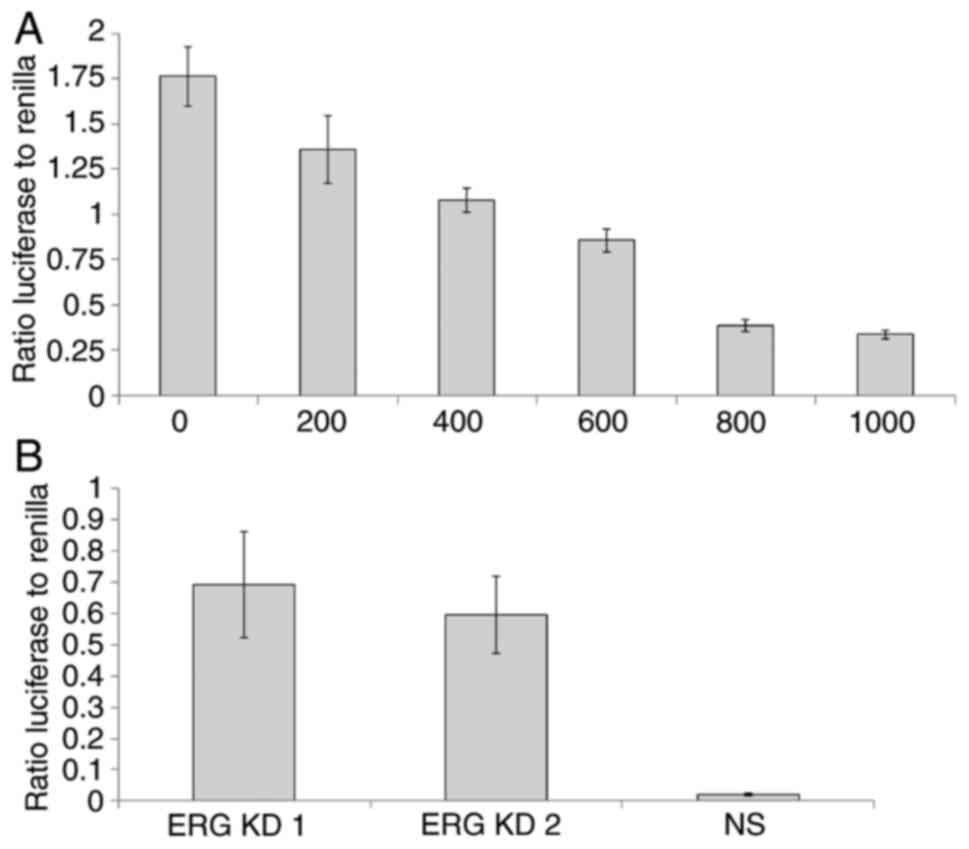
luciferase reporter plasmid for use in a dual-luciferase:Renilla
(DLR) transcription assay. We co-transfected increasing amounts of
the ERG expressing plasmid (0-1,000 ng) with 1 µg of the
PTEN promoter:luciferase construct in DU145 cells. We
observed a progressive reduction in PTEN promoter activity
with increasing amounts of co-transfected ERG expressing plasmid
(Fig. 4A). To extend these findings,
we also transfected the PTEN promoter:luciferase construct
into VCaP cells (which express both ERG and PTEN) and
then knocked-down ERG in VCaP cells using two independent siRNAs,
observing in each case a significant increase in the activity of
the PTEN promoter (Fig.
4B).
The interest in ERG as a potential biomarker
of prostate cancer has grown since the first report of its
over-expression in clinical prostate cancer samples (10). The fusion with the TMPRSS2
promoter allows ERG expression to be driven by androgens. At
the same time the literature indicates clearly that loss of
expression of the PTEN tumour suppressor gene is also linked
to the progression of prostate cancer. Mice that overexpress ERG
and lack PTEN expression develop prostate tumours by the age of six
months, further confirming the synergy between these two genetic
alterations (21). PTEN
deletions tend to occur after ERG activation-it has also been
suggested that ERG might itself drive the development of PTEN
aberrations (22). However it is also
clear that the upregulation of ERG and the deletion of PTEN
expression can occur independently and do not always occur in the
same tumour (23).
In the current study we sought to look into the
possibility that the transcription factor ERG affects the
expression of PTEN directly. We present evidence that ERG
represses PTEN transcription in DU145 and VCaP prostate
cancer cells. By repressing the transcription of PTEN, ERG
could help cancer development by activating the AKT/PI3K pathway,
increasing angiogenesis, proliferation, invasion, motility and
metastasis. This finding may have broader significance because ERG
is not exclusively associated with prostate cancer. ERG is also
implicated in leukaemia where it is linked to chemoresistance
(24,25). The ability of ERG to repress
PTEN transcription in leukaemia, or in other types of
cancer, remains to be investigated.
ERG, like the vast majority of human genes,
is alternatively spliced. We recently reported in advanced prostate
cancer an increased inclusion rate of exons that encode the CAE
domain in the middle of ERG (26).
The CAE domain is thought to modulate ERG's transcriptional
activities. Future experiments will compare the extent to which ERG
splice isoforms can repress PTEN transcription and whether
or not ERG's repression of PTEN plays a role in the development of
aggressive prostate cancer.
In summary, the ability of ERG to repress the
transcription of a critically important tumour suppressor such as
PTEN further implicates ERG in carcinogenesis and underlines
its clear potential as a diagnostic marker and therapeutic
target.
Acknowledgements
We would like to thank Professor Jeff Holly for
providing cell lines and reagents. This study was supported by a
grant from the Bristol Urological Institute (no. BUI 256), the
Rotary Club of Bristol, Funds for Women Graduates, and Prostate
Cancer UK (no. RIA15-ST2-030).
References
|
1
|
Thompson JC, Wood J and Feuer D: Prostate
cancer: Palliative care and pain relief. Br Med Bull. 83:341–354.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Taichman RS, Loberg RD, Mehra R and Pienta
KJ: The evolving biology and treatment of prostate cancer. J Clin
Invest. 1179:2351–2361. 2007. View
Article : Google Scholar
|
|
3
|
Abate-Shen C and Shen MM: Molecular
genetics of prostate cancer. Genes Dev. 14:2410–2434. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chu DC, Chuang CK, Liou YF, Tzou RD, Lee
HC and Sun CF: The use of real-time quantitative PCR to detect
circulating prostate-specific membrane antigen mRNA in patients
with prostate carcinoma. Ann N Y Acad Sci. 1022:157–162. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Parekh DJ, Ankerst DP, Troyer D,
Srivastava S and Thompson IM: Biomarkers for prostate cancer
detection. J Urol. 178:2252–2259. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kumar-Sinha C and Chinnaiyan AM: Molecular
markers to identify patients at risk for recurrence after primary
treatment for prostate cancer. Urology. 62 Suppl 1:S19–S35. 2003.
View Article : Google Scholar
|
|
7
|
Tryggvadóttir L, Vidarsdóttir L,
Thorgeirsson T, Jonasson JG, Olafsdóttir EJ, Olafsdóttir GH, Rafnar
T, Thorlacius S, Jonsson E, Eyfjord JE and Tulinius H: Prostate
cancer progression and aurvival in BRCA2 mutation carriers. J Natl
Cancer Inst. 99:929–935. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liatsikos EN, Assimakopoulos K and
Stolzenburg JU: Quality of life after radical prostatectomy. Urol
Int. 80:226–230. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gopalkrishnan RV, Kang DC and Fisher PB:
Molecular markers and determinants of prostate cancer metastasis. J
Cell Physiol. 189:245–256. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tomlins SA, Rhodes DR, Perner S,
Dhanasekaran SM, Mehra R, Sun XW, Varambally S, Cao X, Tchinda J,
Kuefer R, et al: Recurrent fusion of TMPRSS2 and ETS transcription
factor genes in prostate cancer. Science. 310:644–648. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Birdsey GM, Dryden NH, Amsellem V,
Gebhardt F, Sahnan K, Haskard DO, Dejana E, Mason JC and Randi AM:
Transcription factor Erg regulates angiogenesis and endothelial
apoptosis through VE-cadherin. Blood. 111:3498–3506. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Squire JA: TMPRSS2-ERG and PTEN loss in
prostate cancer. Nat Genet. 41:509–510. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mithal P, Allott E, Gerber L, Reid J,
Welbourn W, Tikishvili E, Park J, Younus A, Sangale Z, Lanchbury
JS, et al: PTEN loss in biopsy tissue predicts poor clinical
outcomes in prostate cancer. Int J Urol. 21:1209–1214. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chaux A, Peskoe SB, Gonzalez-Roibon N,
Schultz L, Albadine R, Hicks J, De Marzo AM, Platz EA and Netto GJ:
Loss of PTEN expression is associated with increased risk of
recurrence after prostatectomy for clinically localized prostate
cancer. Mod Pathol. 25:1543–1549. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lotan TL, Carvalho FL, Peskoe SB, Hicks
JL, Good J, Fedor HL, Humphreys E, Han M, Platz EA, Squire JA, et
al: PTEN loss is associated with upgrading of prostate cancer from
biopsy to radical prostatectomy. Mod Pathol. 28:128–137. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chetram MA, Odero-Marah V and Hinton CV:
Loss of PTEN permits CXCR4-mediated tumorigenesis through ERK1/2 in
prostate cancer cells. Mol Cancer Res. 9:90–102. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Singareddy R, Semaan L, Conley-Lacomb MK,
St John J, Powell K, Iyer M, Smith D, Heilbrun LK, Shi D, Sakr W,
et al: Transcriptional regulation of CXCR4 in prostate cancer:
Significance of TMPRSS2-ERG fusions. Mol Cancer Res. 11:1349–1361.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shore P, Whitmarsh AJ, Bhaskaran R, Davis
RJ, Waltho JP and Sharrocks AD: Determinants of DNA-binding
specificity of ETS-domain transcription factors. Mol Cell Biol.
16:3338–3349. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Nhili R, Peixoto P, Depauw S, Flajollet S,
Dezitter X, Munde MM, Ismail MA, Kumar A, Farahat AA, Stephens CE,
et al: Targeting the DNA-binding activity of the human ERG
transcription factor using new heterocyclic dithiophene diamidines.
Nucleic Acids Res. 41:125–138. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
He J, Sun X, Shi T, Schepmoes AA, Fillmore
TL, Petyuk VA, Xie F, Zhao R, Gritsenko MA, Yang F, et al:
Antibody-independent targeted quantification of TMPRSS2-ERG fusion
protein products in prostate cancer. Mol Oncol. 8:1169–1180. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Srivastava A, Price DK and Figg WD:
Prostate tumor development and androgen receptor function
alterations in a new mouse model with ERG overexpression and PTEN
inactivation. Cancer Biol Ther. 15:1293–1295. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Krohn A, Freudenthaler F, Harasimowicz S,
Kluth M, Fuchs S, Burkhardt L, Stahl PC, Tsourlakis M, Bauer M,
Tennstedt P, et al: Heterogeneity and chronology of PTEN deletion
and ERG fusion in prostate cancer. Mod Pathol. 27:1612–1620. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Heselmeyer-Haddad KM, Garcia Berroa LY,
Bradley A, Hernandez L, Hu Y, Habermann JK, Dumke C, Thorns C,
Perner S, Pestova E, et al: Single-cell genetic analysis reveals
insights into clonal development of prostate cancers and indicates
loss of PTEN as a marker of poor prognosis. Am J Pathol.
184:2671–2686. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bock J, Mochmann LH, Schlee C,
Farhadi-Sartangi N, Göllner S, Müller-Tidow C and Baldus CD: ERG
transcriptional networks in primary acute leukemia cells implicate
a role for ERG in deregulated kinase signaling. PLoS One.
8:e528722013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Mochmann LH, Neumann M, von der Heide EK,
Nowak V, Kühl AA, Ortiz-Tanchez J, Bock J, Hofmann WK and Baldus
CD: ERG induces a mesenchymal-like state associated with
chemoresistance in leukemia cells. Oncotarget. 5:351–362. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hagen RM, Adamo P, Karamat S, Oxley J,
Aning JJ, Gillatt D, Persad R, Ladomery MR and Rhodes A:
Quantitative analysis of ERG expression and its splice isoforms in
formalin-fixed, paraffin-embedded prostate cancer samples:
Association with seminal vesicle invasion and biochemical
recurrence. Am J Clin Pathol. 142:533–540. 2014. View Article : Google Scholar : PubMed/NCBI
|