Introduction
Osteosarcoma is the most common non-haematological
primary malignant bone tumor that occurs in children and
adolescents and its overall relapse-free survival rate over 5 years
is 65–75% (1). Osteosarcoma is
characterized by a highly malignant and metastatic potential, and
the leading cause of death of osteosarcoma patients is distant
metastases (2). However, at present,
the pathogenesis of osteosarcoma remains unclear.
Increasing evidences have showed that tumor stem
cells are considered to be responsible for the metastasis of tumors
(3), and some stem cell-related genes
are involved in tumorgenesis (4,5). MSI1 is a
RNA-binding protein of the Musashi family involved in early
asymmetric divisions generating differentiated cells from neural
stem cells or progenitor cells. MSI1 is highly enriched in the
nervous system and has been found to be related with the grade of
the malignancy in glioma (6).
Additionally, MSI1 drives progenitor cell expansion along the
luminal and myoepithelial lineages in mammary glands and regulates
the proliferation and apoptosis of mesenchymal stem cells.
Recently, high MSI1 expression has been found in various types
tumors, including medulloblastoma (7), colon cancer (8), lung cancer (9), cervical cancer (10) and breast cancer (11), and appeared to be a maker of poor
prognosis. Moreover, MSI1 has been found to activate the Notch and
Wnt signaling pathways in several types of normal and cancerous
cells (12,13). However, the role of MSI1 in
osteosarcoma progression is currently unclear.
Here, we investigated the function of MSI1 and its
mechanism in the progression of osteosarcoma. In this study, we
found that MSI1 expression is increased in osteosarcoma tissue
compared with paraneoplastic tissue. Knockdown of MSI1 resulted in
the decreased cell proliferation and slow growth of the tumor
xenografts. Furthermore, knockdown of MSI1 resulted in the arrest
of cell cycle and up-regulation of p21 and p27 protein expression.
These results supported that MSI1 functions as an oncogene in
osteosarcoma.
Materials and methods
Tissue samples
A total of 30 Fresh frozen specimens of matched
osteosarcoma tissues and paraneoplastic tissues were collected from
Shandong Jining No. 1 People's Hospital (Jining, China) from 2011
to 2014. None of the patients had received chemotherapy,
immunotherapy or radiotherapy before the specimen collection. The
study was approved by Institutional Research Ethics Committee, and
patients gave their informed consent before sample collection.
Cell lines and cell culture
Human osteosarcoma cell lines, MG-63 and HOS, were
purchased from the American Type Culture Collection (Rockville, MD,
USA) and were cultured in Dulbecco's modified Eagle's medium (DMEM;
Sigma-Aldrich, St. Louis, MO, USA) supplemented with 10% fetal
bovine serum (FBS; Invitrogen; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) and 1% penicillin-streptomycin. All cells were
cultured at 37°C in an atmosphere 5% CO2.
Vector construction
The small interfering RNA expression vector that
expresses MSI1-specific short hairpin RNA (shRNA) was purchased
from GenePharma Co., Ltd. (Shanghai, China). To construct reporter
vector containing the 3′UTR of p21 and p27, the fragments of the
3′UTR of p21 and p27 mRNA were separately extracted from MG-63
cells and amplified from cDNA by PCR using primers listed below,
then cloned into the pMIR-REPORT luciferase vector. The following
primers were listed in Table I.
 | Table I.The primer sequence of p21 and
p27. |
Table I.
The primer sequence of p21 and
p27.
| Gene | Orientation | Primer sequence |
|---|
| WT p21 | Forward |
5′-ATTGAGCTCTAATCCGCCCACAGGAAG-3′ |
|
| Reverse |
5′-CTCAAGCTTACAAGTAAAGTCACTAAG-3′ |
| MU p21 | Forward |
5′-TGGGAACGACTGTCTTTCCTGGCACTAACGTT-3′ |
|
| Reverse |
5′-AGACAGTCGTTCCCAGCCCCATATGAGCCCAC-3′ |
| WT p27 | Forward | F:
5′-CGCGAGCTCGAATTAAGAATATGTTTC-3′ |
|
| Reverse |
5′-TTGACGCGTATGCAACCTTTTAAGCATAGC-3′ |
| MU p27 | Forward |
5′-CCGCTCGAGTGATCTGCCTCTAAAAGCGT-3′ |
|
| Reverse |
5′-CGGGATCCATTCTTAACATTCAAAACTCCC-3′ |
Western blot analysis
Cells and clinical tissues were lysed on ice in
lysis buffer containing freshly added protease inhibitor cocktail
(Roche Diagnostics, Branchburg, NJ, USA). The protein extracts (15
µg) were separated using SDS-PAGE and transferred into
polyvinylidene difluoride (PVDF) membranes (EMD Millipore,
Billerica, MA, USA). The appropriate primary antibodies were used
after the membranes were blocked in 5% fat-free milk. The primary
antibodies included the following: anti-MSI1 (1:1,000, cat. no.
sc-98845; Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA),
anti-p21 (1:500, cat. no. sc-397; Santa Cruz Biotechnology, Inc.),
anti-p27 (1:500, cat. no. sc-397; Santa Cruz Biotechnology, Inc.)
and anti-β-actin (1:500, cat. no. sc-47778; Santa Cruz
Biotechnology, Inc.). Blots were incubated with a secondary
antibody coupled to horseradish peroxidase (Thermo Fisher
Scientific Inc.), and visualized on X-ray film. Relative
quantitation was measured using the AlphaView system (Cell
Biosciences, Santa Clara, CA, USA).
RT-qPCR analysis
Total RNA was extracted from cells with TRIzol
Reagent (Invitrogen; Thermo Fisher Scientific, Inc.) and reverse
transcription reactions were performed using RT-PCR kit (Takara
Biotechnology Co., Ltd., Dalian, China). Relative mRNA levels were
evaluated by quantitative PCR using SYBR-Green Mastermix (Takara
Biotechnology Co., Ltd.). The primer sequences were listed in
Table II. The results were analyzed
according to the Cq (ΔΔCq) method using GAPDH as the normalizing
gene.
| Table II.The primer sequence of p21 and
p27. |
Table II.
The primer sequence of p21 and
p27.
| Genes | Orientation | Primer sequence |
|---|
| p21 | Forward |
5′-GAGCAGTGCCCGAGTTAAGG-3′ |
|
| Reverse |
5′-TGGAACAGGTCGGACATCAC-3′ |
| p27 | Forward |
5′-GGTGCCTTCAATTGGGTCTC-3′ |
|
| Reverse |
5′-GCTTCCTCATCCCTGGACAC-3′ |
MTT assays
Cell viability was assessed every other day using
3-(4,5-dimethylthiazole-yl) 2,5-diphenyl tetrazolium bromide (MTT,
Sigma-Aldrich, USA) dye according to standard protocol.
Approximately 2×103 cells/well were seeded in a 96-well
plate and incubated for 7 days. 20 µl of MTT solution was added to
200 µl of culture media and incubated for 4 h, and then dissolved
in 100 µl of dimethylsulphoxide (DMSO; Sigma-Aldrich). Cell
proliferation was determined by measuring the absorbance at 490
nm.
Flow cytometry analysis
Cells (2×106) were harvested and fixed
with 70% cold ethanol at 4°C overnight. After washed twice in PBS,
the cells were suspended in PBS with 50 µg/ml propidium iodide and
10 µg/ml RNaseA, and then incubated at room temperature for 30 min
in the dark. Then, the cells were measured by FACSCalibur flow
cytometry (BD Biosciences, Franklin Lakes, NJ, USA), and the cell
cycle distributions were analyzed.
In addition, the cells were harvested, washed in PBS
and stained in duplicate with APC annexin V and propidium iodide
(BD Pharmingen, Franklin Lakes, NJ, USA) for 10 min in the dark to
characterize cell apoptosis using flow cytometry.
Animal and tumor xenograft assay
Tumor cells collected from stable transfectants were
bilaterally injected into subcutis on the dorsum of 6- to
7-week-old Balb/c nude mice. Tumor size was measured with calipers
once every week, and volumes (cm3) were calculated
according to the standard formula: V=length × width2/2.
At the end of the experiment, tumors were dissected out, and the
net weight was measured. All animals received humane treatment in
accordance with institutional policies, and all experimental
protocols were approved by the Animal Care and Use Committee of
Jining First people's Hospital.
Luciferase reporter assay
In brief, plasmids containing firefly luciferase
reporters were cotransfected into cells using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.). Then, the cells were lysed in 100 ul of passive
lysis buffer (Promega Corporation, Madison, WI, USA) at 48 h after
transfection and determined with a dual-luciferase assay according
to the manufacturer's instructions. Luciferase activities were
detected in a luminometer (Promega Corporation) and expressed at
the ratio of Firefly to Renilla luciferase activity.
Statistical analysis
All results were confirmed in three independent
experiments, and data were expressed as the mean ± standard
deviation. Student's t-test or one-way ANOVA test were performed
using SPSS 16.0 (SPSS, Inc., Chicago, IL, USA). P<0.05 was
considered to indicate a statistically significant difference.
Results
The expression of MSI1 was upregulated
in osteosarcoma
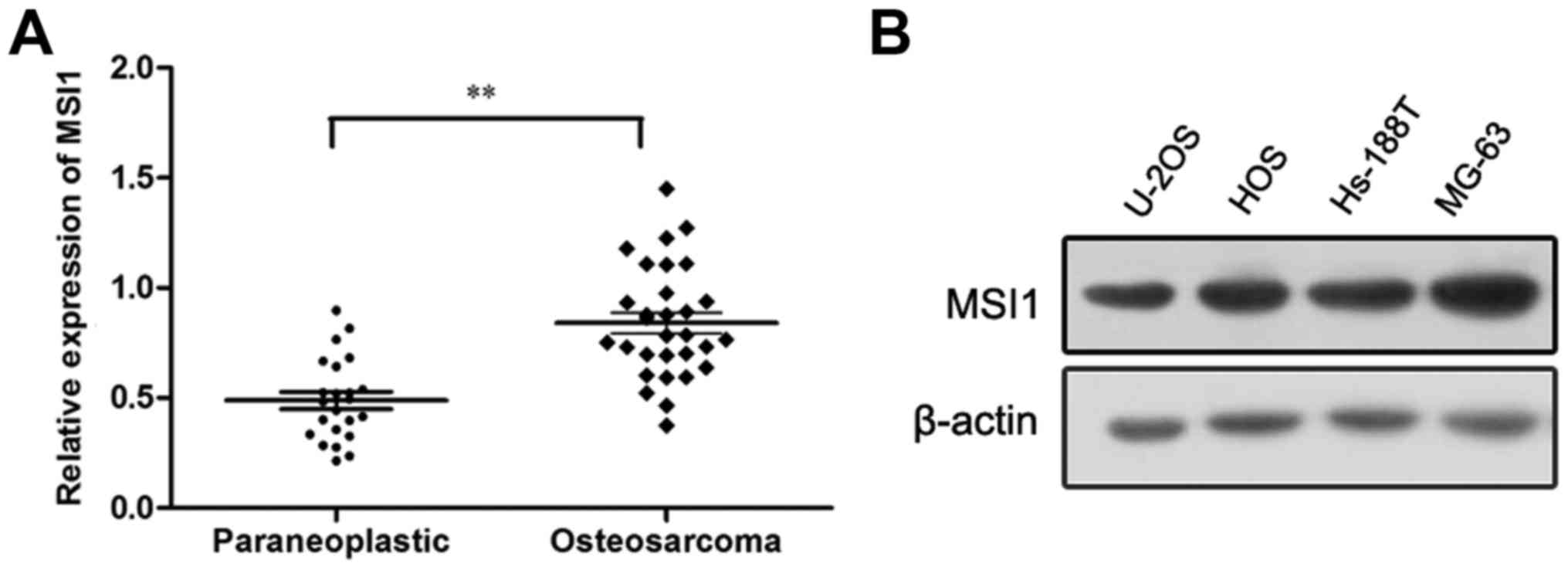
To determine the expression of MSI1 in osteosarcoma,
Western blot analysis was conducted. We found the expression of
MSI1 in osteosarcoma tissues was markedly higher than that in the
paraneoplastic tissues (P<0.01, Fig.
1A). Additionally, the expression of MSI1 in osteosarcoma cell
lines was also measured by western blot, and the high expression of
MSI1 in U-2OS, HOS, Hs-188T and MG-63 cell lines were observed
(Fig. 1B). These data suggested MSI1
may contribute to the progression of osteosarcoma.
The effect of knockdown of MSI1 on the
proliferation and apoptosis of osteosarcoma cells
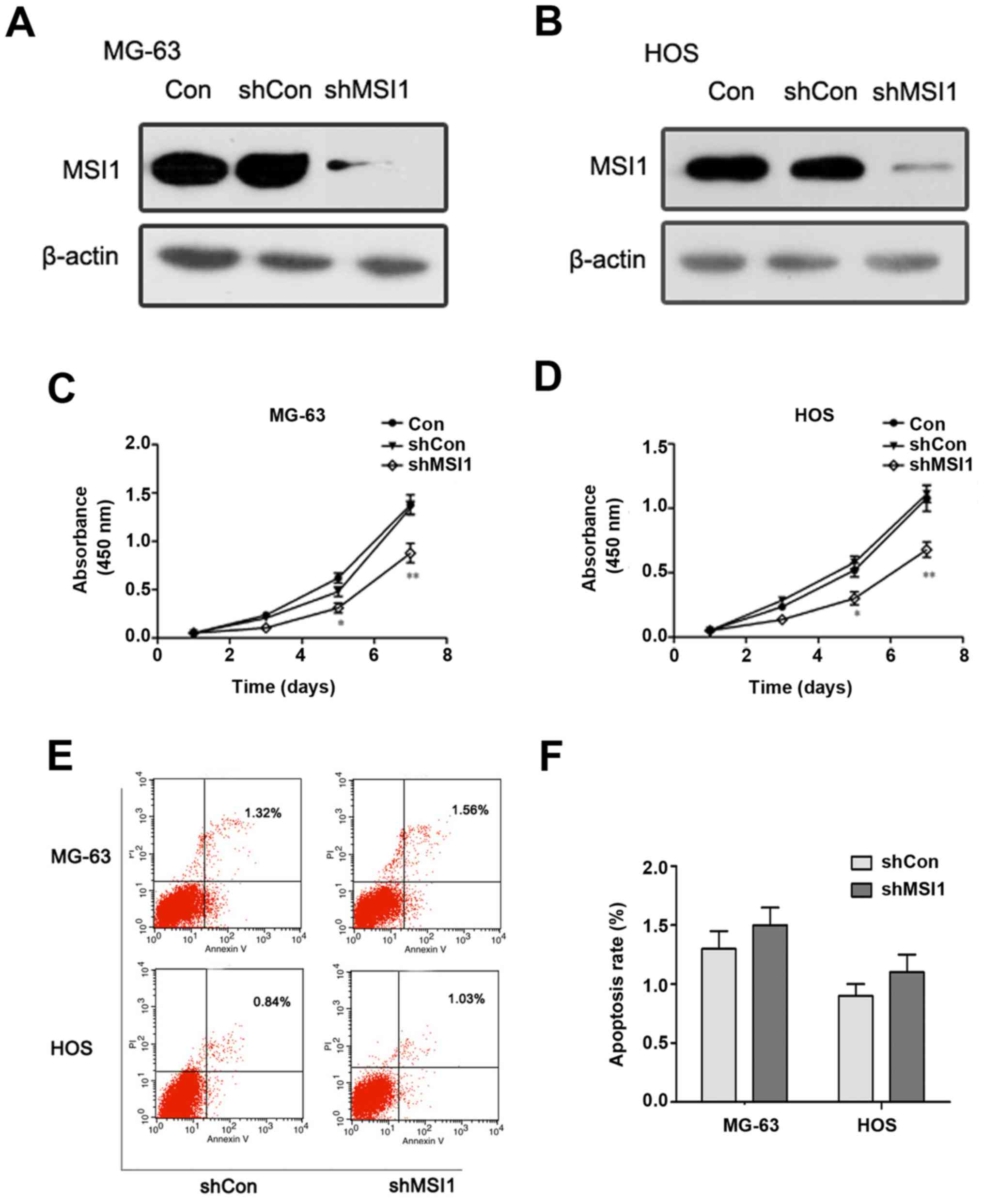
To assess the effect of MSI1 on the proliferation
and apoptosis of osteosarcoma cells, we established MG-63 and HOS
osteosarcoma cells with MSI1 knockdown by shRNA, respectively
(Fig. 2A and B). MTT assay revealed
that MSI1 knockdown MG-63 cells had much lower proliferation
ability than the corresponding control cells (Fig. 2C), and similar results were observed
in MSI1 knockdown HOS cells (Fig.
2D). These findings suggested that knockdown of MSI1 suppressed
the proliferation of osteosarcoma cells.
However, the apoptosis analysis of MG-63 and HOS
cells using flow cytometry showed that there was no significant
difference in the proportion of apoptotic cells between MSI1
knockdown cells and the control (Fig. 2E
and F), which suggested that MSI1 had no significant influence
on apoptosis in the osteosarcoma cells.
Knockdown of MSI1 inhibited the tumor
formation of osteosarcoma cells
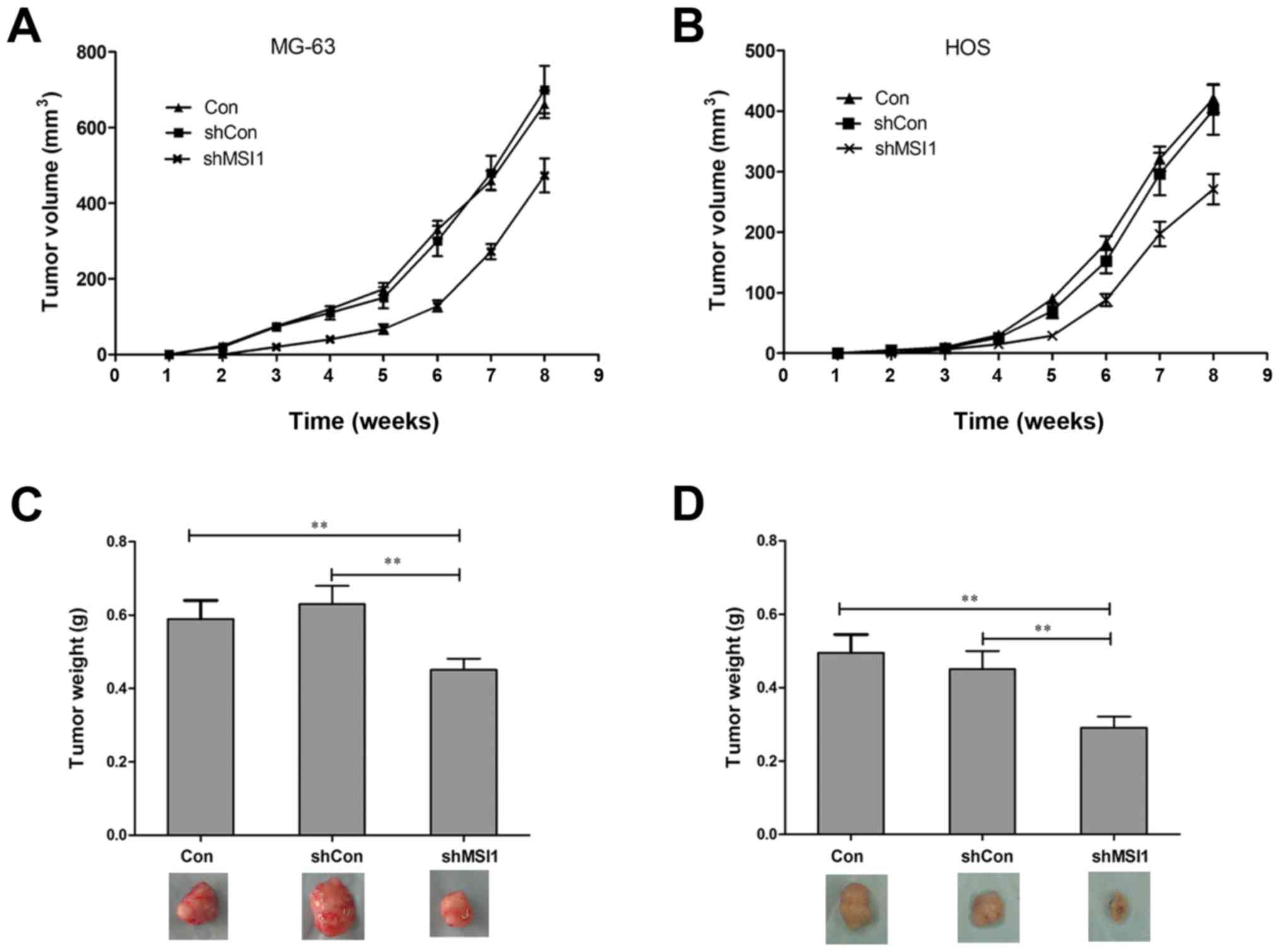
To further assess the effect of MSI1 on the tumor
formation, nude mice were injected subcutaneously with MSI1
knockdown MG-63 and HOS cells, and the growth of tumors was
monitored in terms of tumor volume every three days. At the end of
the experiment, the mice were sacrificed, and the tumors were
excised and weighed. The tumors formed by the MSI1 knockdown MG-63
and HOS cells grew much slower than those formed by the Control
(Fig. 3A and B, P<0.01). In
addition, the weights of the tumors formed by the MSI1 knockdown
cells were significantly reduced compared to the Control (Fig. 3C and D, P<0.01). All these data
suggested that knockdown of MSI1 inhibited the tumor formation
ability of osteosarcoma cells in vivo.
Knockdown of MSI1 led to the cell
cycle arrest of osteosarcoma cells
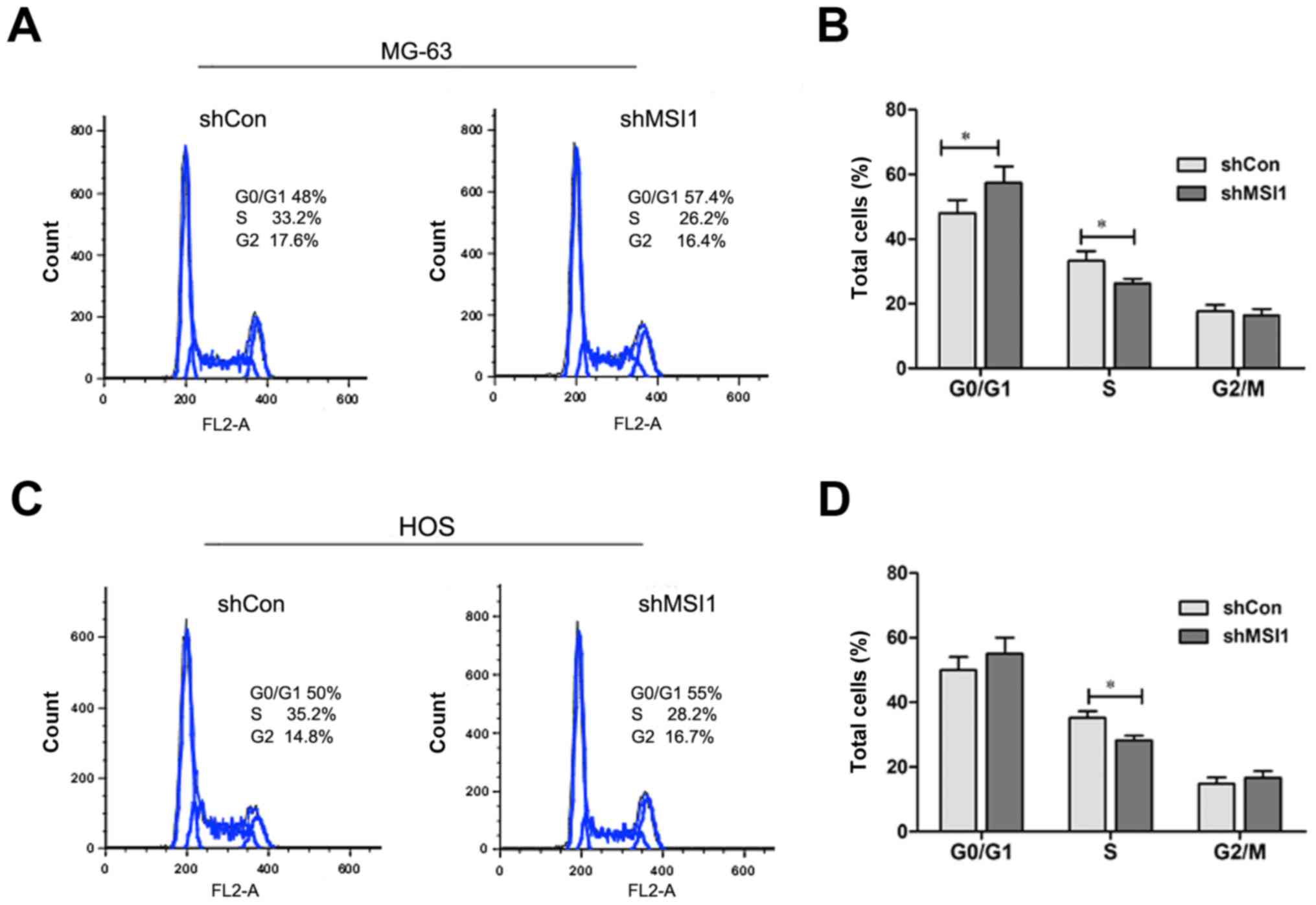
We all know that cell proliferation is usually
associated with the modulation of the cell cycle, then, we
characterized cell cycles by flow cytometry analysis. As shown in
Fig. 4A and B, the proportion of MSI1
knockdown MG-63 cells in G0/G1 phase was markedly increased, while
proportion in S phase was significantly decreased compared to the
control (P<0.05), and a similar result was observed in MSI1
knockdown HOS cells (Fig. 4C and D).
These data indicated that the knockdown of MSI1 resulted in the
cell cycle arrest.
Knockdown of MSI1 increased the
expression of p21 and p27 protein
Previous research reported that MSI1 regulated cell
cycle by negative regulation of p21 expression in bladder carcinoma
(14) and breast cancer (15). To explore the molecular mechanism of
MSI1 on the regulation of cell cycle of osteosarcoma cells, the
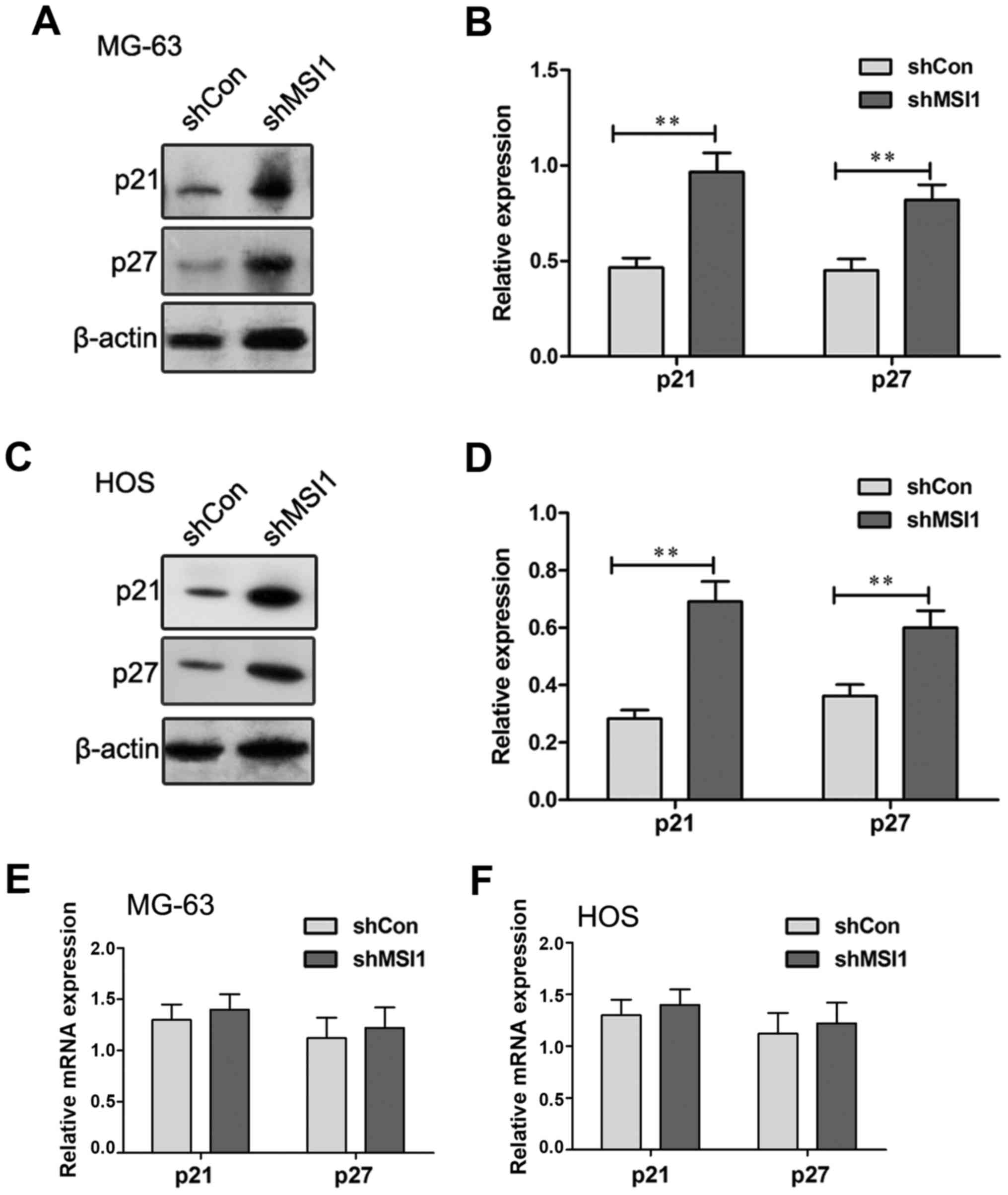
expression of p21 and p27 were examined by Western blot (Fig. 5A-D). A significant increase in the
levels of p21 and p27 protein expression were observed in the MSI1
knockdown MG-63 and HOS cells compared to their control cells,
respectively (P<0.01), suggesting that MSI1 may reduce the
levels of p21 and p27 protein expression. However, the examination
using RT-qPCR assay showed that no significant differences in the
expression of p21 and p27 mRNA were observed in the MSI1 knockdown
MG-63 and HOS cells and their control (Fig. 5E and F). Therefore, these data
indicated that MSI1 could influence the expression of p21 and p27
at protein level, not mRNA level in MG-63 and HOS cells.
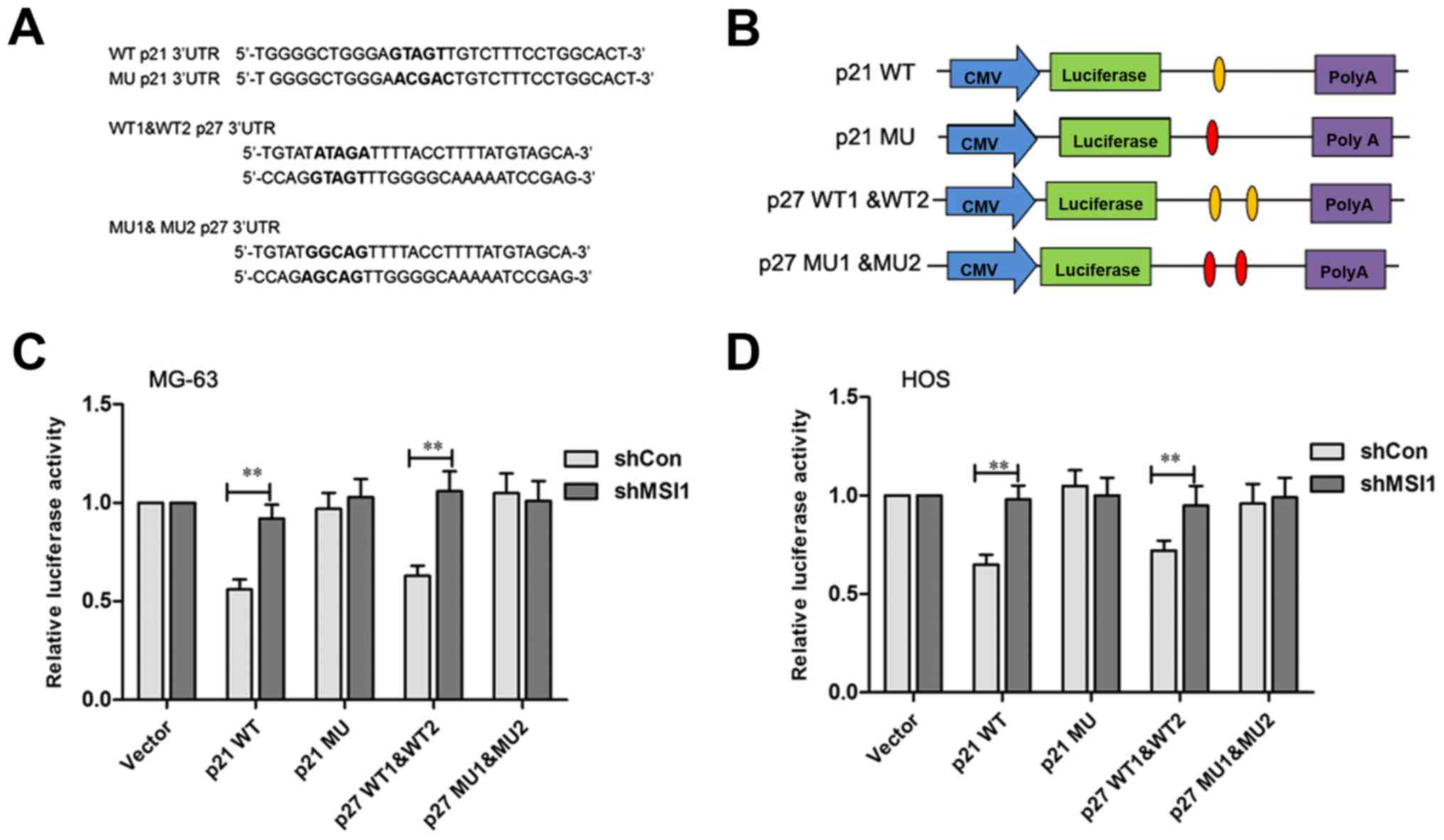
It has been reported that p21 and p27 were the MSI1
target genes in which there is a binding sequence region in their
3′UTR (16,17). The MSI1 binding sites in p21 and p27
protein are shown in Fig. 6A,
respectively. To test the direct interaction between MSI1 and its
putative target site, the wild-type and mutant MSI1 binding
sequence was inserted downstream of the luciferase vectors
(Fig. 6B). After transfected by p21
wild-type vector, the relative luciferase activity of MSI1
knockdown MG-63 and HOS cells was significantly increased compared
to the control (P<0.01), however, no significant difference in
the luciferase activities between the MSI1 knockdown MG-63 and HOS
cells and their control cells transfected with the mutant vector
was observed (Fig. 6C and D).
Similarly, the luciferase activity levels in MSI1 knockdown MG-63
and HOS cells were found to be higher than their control cells
after being transfected with the p27 1&2 wild-type vector
(P<0.01), while, the differences were not significant between
the MSI1 knockdown MG-63 and HOS cells and their control cells
after being transfected by p27 1&2 mutant vector. These data
suggested that MSI1 might inhibit p21 and p27 translation by
specifically binding site.
Discussion
Increasing evidence showed that RNA binding proteins
play an important role in the cell proliferation and apoptosis
involved in tumorigenesis (18–20). MSI1,
a multifunction RNA binding protein of MSI family, was initially
identified as a marker of neuronal stem cell (21,22) and
more recently identified as a putative marker of intestinal stem
cell (23), was overexpressed in
several types of cancers, such as glioma (13), medulloblastoma (24), and breast cancer (15).
In this study, we found that the expression of MSI1
protein in osteosarcoma tissues was significantly higher than that
of the paraneoplastic tissues, suggesting MSI1 was likely to play
an important role in osteosarcomagenesis. To further seek for the
function of MSI1 involved in osteosarcomagenesis, the functional
characterization was conducted by knockdown of MSI1 in osteosarcoma
cell lines (MG-63 and HOS cells). We found that knockdown of MSI1
significantly inhibited the proliferation of MG-63 and HOS cells
in vitro. Moreover, knockdown of MSI1 also significantly
suppressed the tumor growth in vivo. These results suggested
that MSI1 played an important role in the cell proliferation and
tumor growth. In addition, the analysis of cell cycle and apoptosis
showed that knockdown of MSI1 induced cell cycle arrest in the
G0/G1 phase, but no markedly change of apoptosis, suggesting that
MSI1 might have important impact on the cell cycle, not apoptosis.
It is well known that cell proliferation is usually regulated by
cell cycle progression. Therefore, our data suggested that the
inhibitory effect of MSI1 knockdown on cell proliferation and tumor
formation of osteosarcoma cells were probably associated with the
regulation of cell cycle. Our finding was in agreement with
previous findings in other types of tumors (25–27),
suggesting that MSI1 related signaling may have a similar
regulatory effect on the growth of different types of human
malignancies.
It has been reported that increased expression of
p21 and p27 leads to apoptosis and cell cycle arrest in various
cancer cells (16,28,29). In
this study, we found the expression of p21 and p27 protein levels
were up-regulated in the MSI1 knockdown MG-63 and HOS cells
compared to the control. As p21 and p27 is universal
cyclin-dependent kinase (CDK) inhibitor that directly inhibits the
activity of cyclin-CDK complexes (30,31),
resulting in cell cycle arrest at G0/G1 phase, these results
suggested that the cell cycle arrest caused by MSI1 knockdown in
osteosacoma cells might be associated with the overexpression of
p21 and p27. Furthermore, luciferase assay revealed that MSI1 could
directly bind to the consensus sequence of p21 and p27 3′UTR in
osteosarcoma cells, indicating that knockdown of MSI1 could lead to
the up-regulation of the expression of p21 and p27.
In summary, our study demonstrated that MSI1 was
high expressed in osteosarcoma tissue, and knockdown of MSI1
inhibited the cell proliferation and tumor formation by the arrest
of cell cycle involved the activation of p21 and p27. Our study
provided a potential target for clinical therapy of
osteosarcoma.
References
|
1
|
Lewis IJ, Nooij MA, Whelan J, Sydes MR,
Grimer R, Hogendoorn PC, Memon MA, Weeden S, Uscinska BM, van
Glabbeke M, et al: Improvement in histologic response but not
survival in osteosarcoma patients treated with intensified
chemotherapy: A randomized phase III trial of the European
osteosarcoma intergroup. J Natl Cancer Inst. 99:112–128. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Buddingh EP, Anninga JK, Versteegh MI,
Taminiau AH, Egeler RM, van Rijswijk CS, Hogendoorn PC, Lankester
AC and Gelderblom H: Prognostic factors in pulmonary metastasized
high-grade osteosarcoma. Pediatr Blood Cancer. 54:216–221.
2010.PubMed/NCBI
|
|
3
|
Kapoor A and Kumar S: Cancer stem cell: A
rogue responsible for tumor development and metastasis. Indian J
Cancer. 51:282–289. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Liu XF, Yang WT, Xu R, Liu JT and Zheng
PS: Cervical cancer cells with positive Sox2 expression exhibit the
properties of cancer stem cells. PLoS One. 9:e870922014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wu XS, Xi HQ and Chen L: Lgr5 is a
potential marker of colorectal carcinoma stem cells that correlates
with patient survival. World J Surg Oncol. 10:2442012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Toda M, Iizuka Y, Yu W, Imai T, Ikeda E,
Yoshida K, Kawase T, Kawakami Y, Okano H and Uyemura K: Expression
of the neural RNA-binding protein Musashi1 in human gliomas. Glia.
34:1–7. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sanchez-Diaz PC, Burton TL, Burns SC, Hung
JY and Penalva LO: Musashi1 modulates cell proliferation genes in
the medulloblastoma cell line Daoy. BMC Cancer. 8:2802008.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fan LF, Dong WG, Jiang CQ, Xia D, Liao F
and Yu QF: Expression of putative stem cell genes Musashi-1 and
beta1-integrin in human colorectal adenomas and adenocarcinomas.
Int J Colorectal Dis. 25:17–23. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang XY, Yu H, Linnoila RI, Li L, Li D, Mo
B, Okano H, Penalva LO and Glazer RI: Musashi1 as a potential
therapeutic target and diagnostic marker for lung cancer.
Oncotarget. 4:739–750. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ye F, Zhou C, Cheng Q, Shen J and Chen H:
Stem-cell-abundant proteins Nanog, Nucleostemin and Musashi1 are
highlyexpressed in malignant cervical epithelial cells. BMC Cancer.
8:1082008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang XY, Penalva LO, Yuan H, Linnoila RI,
Lu J, Okano H and Glazer RI: Musashi1 regulates breast tumor cell
proliferation and is a prognostic indicator of poor survival. Mol
Cancer. 9:2212010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang XY, Yin Y, Yuan H, Sakamaki T, Okano
H and Glazer RI: Musashi1 modulates mammary progenitor cell
expansion through proliferin-mediated activation of the Wnt and
Notch pathways. Mol Cell Biol. 28:3589–3599. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Muto J, Imai T, Ogawa D, Nishimoto Y,
Okada Y, Mabuchi Y, Kawase T, Iwanami A, Mischel PS, Saya H, et al:
RNA-binding protein Musashi1 modulates glioma cell growth through
the post-transcriptional regulation of Notch and PI3 kinase/Akt
signaling pathways. PLoS One. 7:e334312012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Nikpour P, Baygi ME, Steinhoff C, Hader C,
Luca AC, Mowla SJ and Schulz WA: The RNA binding protein
Musashi1regulates apoptosis, gene expression and stress granule
formation in urothelial carcinoma cells. J Cell Mol Med.
15:1210–1224. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang XY, Penalva LO, Yuan H, Linnoila RI,
Lu J, Okano H and Glazer RI: Musashi1 regulates breast tumor cell
proliferation and is a prognostic indicator of poor survival. Mol
Cancer. 9:2212010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu X, Yang WT and Zheng PS: Msi1 promotes
tumor growth and cell proliferation by targeting cell cycle
checkpoint proteins p21, p27 and p53 in cervical carcinomas.
Oncotarget. 5:10870–10885. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Götte M, Greve B, Kelsch R, Müller-Uthoff
H, Weiss K, Masouleh Kharabi B, Sibrowski W, Kiesel L and Buchweitz
O: The adult stem cell marker Musashi-1 modulates endometrial
carcinoma cell cycle progression and apoptosis via Notch-1 and p21
WAF1/CIP1. Int J Cancer. 129:2042–2049. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Abdelmohsen K, Srikantan S, Kuwano Y and
Gorospe M: miR-519 reduces cell proliferation by lowering
RNA-binding protein HuR levels. Proc Natl Acad Sci USA.
105:20297–20302. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Busà R, Paronetto MP, Farini D,
Pierantozzi E, Botti F, Angelini DF, Attisani F, Vespasiani G and
Sette C: The RNA-binding protein Sam68 contributes to proliferation
and survival of human prostate cancer cells. Oncogene.
26:4372–4382. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sutherland LC, Rintala-Maki ND, White RD
and Morin CD: RNA binding motif (RBM) proteins: A novel family of
apoptosis modulators? J Cell Biochem. 94:5–24. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kaneko Y, Sakakibara S, Imai T, Suzuki A,
Nakamura Y, Sawamoto K, Ogawa Y, Toyama Y, Miyata T and Okano H:
Musashi1: An evolutionally conserved marker for CNS progenitor
cells including neural stem cells. Dev Neurosci. 22:139–153. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Okano H, Kawahara H, Toriya M, Nakao K,
Shibata S and Imai T: Function of RNA-binding protein Musashi-1 in
stem cells. Exp Cell Res. 306:349–356. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Rezza A and Plateroti M: M1621 Study of
the RNA binding protein Musashi1, An intestinal epithelial stem
cell marker. Gastroenterology. 136:A–396. 2009. View Article : Google Scholar
|
|
24
|
Vo DT, Subramaniam D, Remke M, Burton TL,
Uren PJ, Gelfond JA, de Sousa Abreu R, Burns SC, Qiao M, Suresh U,
et al: The RNA-binding protein Musashi1 affects medulloblastoma
growth via a network of cancer-related genes and is an indicator of
poor prognosis. Am J Pathol. 181:1762–1772. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Shi C and Zhang Z: miR-761 inhibits tumor
progression by targeting MSI1 in ovarian carcinoma. Tumor Biol.
37:5437–5443. 2016. View Article : Google Scholar
|
|
26
|
Akasaka Y, Saikawa Y, Fujita K, Kubota T,
Ishikawa Y, Fujimoto A, Ishii T, Okano H and Kitajima M: Expression
of a candidate marker for progenitor cells, Musashi-1, in the
proliferative regions of human antrum and its decreased expression
in intestinal metaplasia. Histopathology. 47:348–356. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wei JG, Zhao F and Sun AJ: Effect of Msi1
siRNA on proliferation of SW-480 cell line. Chin J Clin Exp Pathol.
2012.
|
|
28
|
Wang YF, Chen NS, Chung YP, Chang LH,
Chiou YH and Chen CY: Sodium butyrate induces apoptosis and cell
cycle arrest in primary effusion lymphoma cells independently of
oxidative stress and p21 CIP1/WAF1 induction. Mol Cell Biochem.
285:51–59. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Møller MB: P27 in cell cycle control and
cancer. Leuk Lymphoma. 39:19–27. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liu DF, Ferguson K, Cooper GS, Grady WM
and Willis J: p27 cell-cycle inhibitor is inversely correlated with
lymph node metastases in right-sided colon cancer. J Clin Lab Anal.
13:291–295. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Coqueret O: New roles for p21 and p27
cell-cycle inhibitors: A function for each cell compartment? Trends
in Cell Biol. 13:65–70. 2003. View Article : Google Scholar
|