There were an estimated ~1 million incident
diagnoses of gastric cancer globally in 2012, half of which
occurred in Eastern Asia, particularly in China (1). Although the incidence of gastric cancer
has reduced in the past few decades, it remains the 5th most
frequently diagnosed type of cancer and the 3rd leading cause of
cancer-associated mortality globally (1). Peritoneal dissemination is the typical
metastatic pattern in gastric cancer and data obtained from between
1969 and 1988 in Japan indicated that peritoneal recurrence
accounted for >40% of recurrence cases (2,3). To date,
there is no standard treatment for peritoneal metastasis.
Therefore, investigating the underlying mechanisms of peritoneal
dissemination is required for efficient treatment and to further
improve the prognosis of patients. Paget's ‘seed and soil’ theory
has been established as the fundamental theory of peritoneal
dissemination in gastric cancer (4,5). This
hypothesis compares the viable cancer cells to ‘seeds’ and the
suitable microenvironment of the metastatic sites to ‘soil’
(6). The current review will
thoroughly discuss the underlying mechanisms of peritoneal
dissemination in gastric cancer, based on the ‘seed and soil’
theory.
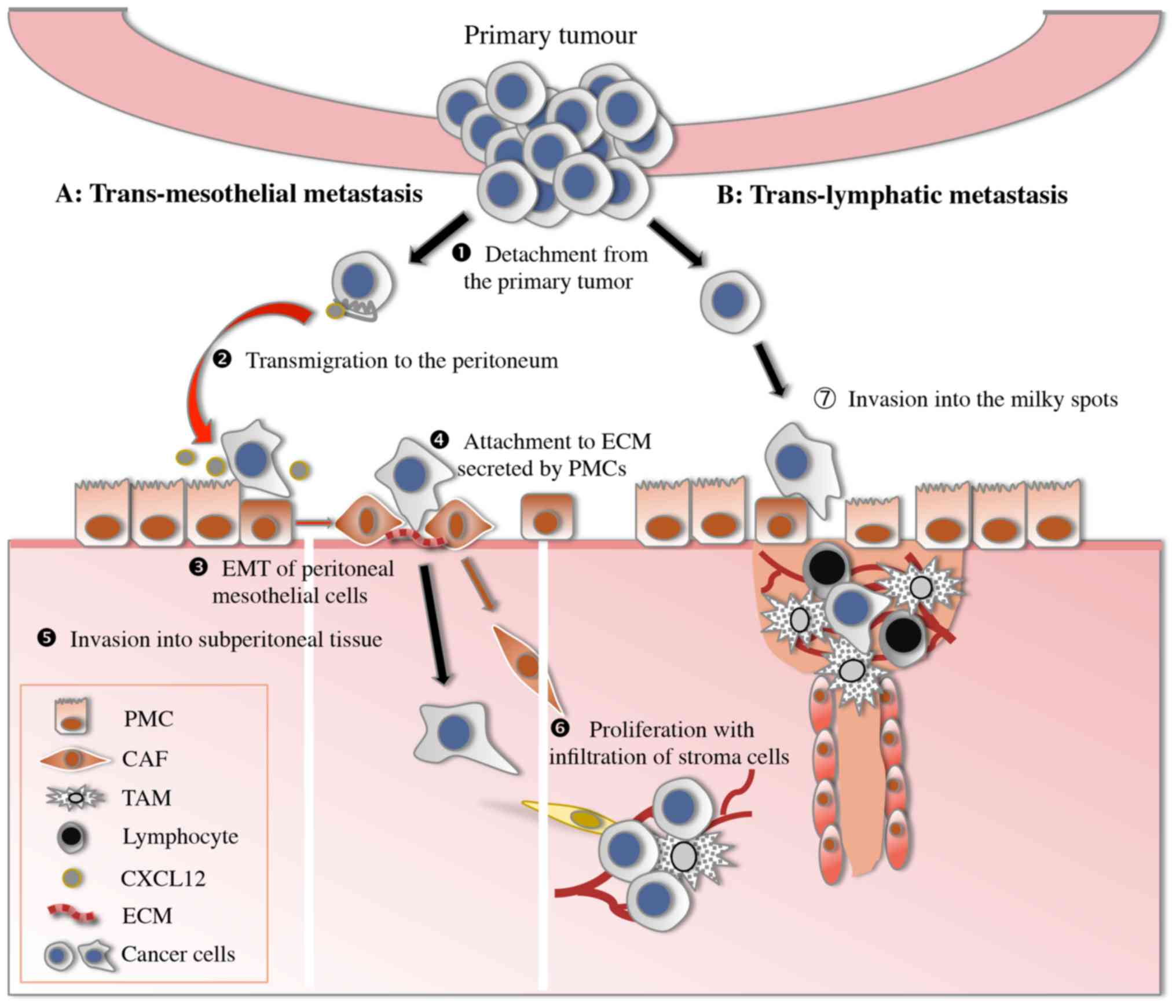
The ‘seeding’ theory demonstrates that several
consecutive steps participate in the development of peritoneal
dissemination, including detachment from the primary tumour,
transmigration to the peritoneum, attachment to peritoneal
mesothelial cells or lymphatic stomata, invasion through the
basement membrane to subperitoneal tissue and proliferation with
blood vascular neogenesis (Fig. 1)
(7). Finally, the progress of these
processes is discussed.
Detachment of cancer cells from the primary tumour
is considered the 1st step of peritoneal dissemination. These
detached cancer cells are designated peritoneal-free cancer cells
and the positive rate of cytological examination is associated with
the depth of tumour invasion and degree of differentiation
(8). With the recent rapid
development of molecular biology techniques, various novel tumour
markers have been identified to be predictors of subclinical
peritoneal dissemination in gastric cancer. Epithelial (E)-cadherin
mediates the adherens junction between homophilic cells, inhibiting
the ability of gastric cancer cells to detach from the primary
tumour. S100 calcium binding protein A4 (S100A4) affects cancer
cell motility via the alteration of cytoskeletal dynamics. Gastric
cancer cells with downregulated epithelial (E)-cadherin and
upregulated S100A4 expression have an increased probability of
undergoing serosal involvement and peritoneal dissemination
(9).
Primary tumours are the principal source of
peritoneal-free cancer cells; two other sources are metastatic
lymph nodes and surgical dissection (10,11). For
patients without serosal invasion, Marutsuka et al (10) demonstrated that lymph node dissection
opens lymphatic channels, which spreads cancer cells into the
abdominal cavity. A recent study performed cytology analysis of
peritoneal wash fluid via reverse transcription-quantitative
polymerase chain reaction (RT-PCR) prior to and following a
gastrectomy, and demonstrated that, of 57 peritoneal wash samples
with no carcinoembryonic antigen (CEA) or cytokeratin (CK)20 prior
to gastrectomy, 35 samples had CEA or CK20 positivity following the
gastrectomy (11). These results
confirmed that surgery may induce viable cancer cell efflux into
the peritoneal cavity during a curative gastrectomy (11). Therefore, it is important for surgeons
to adhere to surgical standards and perform the required actions to
eliminate these viable cancer cells during surgery using modalities
such as hyperthermic intraperitoneal chemotherapy.
The next step of peritoneal dissemination is
transmigration of peritoneal-free cancer cells to the distant
peritoneum. The sites where cancer cells adhere are not randomly
selected but are promoted by chemokines and corresponding receptors
(12,13). Chemokines and their receptors have an
important role in promoting the migration of leucocytes (14). However, chemokine receptors have
recently been the focus of attention as the expression of
chemokines receptors on cancer cells, particularly C-X-C motif
chemokine receptor 4 (CXCR4), is associated with cancer cell
migration and metastasis (15). CXCR4
was first reported as a chemokine receptor that participates in the
lung metastasis of breast cancer (16). An increasing number of studies have
demonstrated that the CXC chemokine ligand 12 (CXCL12)/CXCR4 axis
participates in the development of peritoneal dissemination in
gastric cancer (17). It is
hypothesized that CXCL12 is produced by peritoneal mesothelial
cells and attracts gastric cancer cells expressing CXCR4 (17). The positive rate of CXCR4 expression
is significantly higher in primary gastric cancer with peritoneal
metastasis (67%), compared with other types of distant metastasis
(25%) (15). Although 85% of patients
who are positive for CXCR4 develop peritoneal metastasis, the
proportion of patients who are negative for CXCR4 is only 48%
(15). In addition, high
concentrations of CXCL12 are detected in malignant ascitic fluids
(15). These results are concordant
with the CXCL12/CXCR4 axis serving an important function in the
peritoneal dissemination in gastric cancer; therefore, CXCR4 may be
a novel target for treatment.
Adhesion of gastric cancer cells to the peritoneal
lining is a crucial step in peritoneal dissemination. Formation of
peritoneal dissemination from peritoneal-free gastric cancer cells
is considered to occur through two distinct processes:
Transmesothelial and translymphatic metastasis.
Transmesothelial metastasis originates from the
attachment of peritoneal-free cancer cells to the mesothelium
(18). Human peritoneal mesothelial
cells (HPMCs) have a vital role in this process. Normal peritoneum
is composed of a single layer of mesothelial cells closely
connecting with each other, creating an anatomical barrier that
prevents peritoneal-free gastric cancer cells from invading into
submesothelial tissues (19).
However, the morphology of HPMCs is altered by stimulation with a
number of the growth factors secreted by cancer cells. The
alteration in morphology from epithelial-like to mesenchymal-like
is termed the epithelial-mesenchymal transition (EMT) (20). The EMT is characterized by the
attenuation of E-cadherin expression and elevation of α-smooth
muscle actin (SMA) expression (20).
HPMCs that have undergone EMT take on a spindle-like,
fibroblastic-pattern morphology, which promotes a space between
mesothelial cells and exposes the basement membrane to cancer cells
(21). Transforming growth factor β1
(TGF-β1) serves an important role in the progress of EMT (22).
TGF-β1 is part of the TGF-β superfamily that
controls the proliferation and differentiation of the majority of
cells; it is a crucial promoter in several fibrotic diseases
(23). TGF-β1 has been established to
promote the expression of extracellular matrix (ECM) factors,
including collagen, through Smad3 and the conversion of renal
tubular epithelial cells into fibroblasts in the kidney via Smad2
(24). Regarding gastric cancer,
TGF-β1 levels detected using an ELISA with peritoneal wash fluid
samples are associated with peritoneal dissemination and depth of
invasion (18). Furthermore, TGF-β1
induces the EMT of HPMCs in vitro and in vivo
(25). Subsequent previous studies
have further demonstrated that TGF-β1 increases the expression of
collagen I in rat peritoneal mesothelial cells via the c-Jun
N-terminal kinase (JNK)/Smad3 signalling pathway, and of collagen
III and fibronectin of HPMCs via Smad2 (26,27).
Consequently, mesothelial cells stimulated by TGF-β1 promote
peritoneal fibrosis via upregulating the expression of ECM
proteins, including collagen I, collagen III and fibronectin
(18). Cancer cells may anchor to the
ECM via adhesion molecules; therefore peritoneal fibrosis mediated
by TGF-β1 provides an effective ‘soil’ for dissemination.
Several regulatory factors modulate the function of
TGF-β1, including connective tissue growth factor (CTGF) (28), angiotensin II (Ang II) (29) and phytosulphokine (PSK) (30). CTGF is an important downstream
effector of TGF-β1. A previous study demonstrated that CTGF induces
the EMT of HPMCs directly and that EMT that is mediated by TGF-β1
is reversed when the cells are transfected with CTGH small
interfering (si)RNA (28). These
results are concordant with CTGF participating in the progress of
EMT, mediated by TGF-β1. Ang II is a member of the
renin-angiotensin system (RAS) and the identification of local RAS
in cancer tissues brings more attention to the function of Ang II
in the development of cancer. Local RAS indicates that Ang II is
converted directly from Ang I in tumour tissues in an
angiotensin-converting enzyme (ACE)-independent manner. In
vitro studies have demonstrated that Ang II increases the
expression levels of TGF-β1 and is inhibited by candesartan, a type
of angiotensin II type 1 block (ARB) (29). These results indicated that ARBs may
be used to treat fibrosis in peritoneal dissemination (29). Protein-bound polysaccharide K (PSK),
used as an anticancer immunologic adjuvant therapy, is considered
to prolong the survival of patients with gastric cancer (31). PSK has been established to suppress
the TGF signalling pathway (32).
Shinbo et al (30) reported
that PSK inhibits the EMT of HPMCs induced by TGF-β1 in
vitro, and the co-inoculum of gastric cancer cells and HPMCs
significantly inhibits fibrosis in xenograft tumours. These results
suggest that PSK may be used to control tumour fibrosis in gastric
cancer.
Cluster of differentiation (CD)44, a type of
adhesion molecule produced by gastric cancer cells, also
contributes to the contraction of mesothelial cells by
phosphorylating the cellular skeleton (33). All of these factors result in
morphological alteration of HPMCs and peritoneal fibrosis, which
provides suitable ‘soil’ for peritoneal-free cancer cells.
Consequently, cancer cells migrate through a layer of mesothelial
cells and further attach to the ECM expressed by HPMCs to expose
the basement membrane mediated by integrin molecules (34). Integrins are membrane-bound proteins
that mediate the direct contact between cells and the extracellular
matrix (34). Integrins are composed
of two distinct chains, the α and β subunits (35). There are 18 α-integrin subunits in
mammals. Amongst these subunits, α1, α2, α3 and β1 have been
reported to be closely associated with the peritoneal dissemination
of gastric cancer (36,37), particularly α3β1 integrin (34), which mediates gastric cancer cell
adhesion to laminin. A recent study identified that CTGF
effectively blocks adhesion by binding to α3β1 integrin, and the
authors hypothesized that recombinant CTGF may have therapeutic
potential (38).
Degradation of the ECM and the basement membrane is
beneficial for the invasion of cancer cells. Matrix
metalloproteinases (MMPs) are able to degrade all types of ECM.
MMP7, and MMP14 are synthesized primarily by cancer cells, whilst
other MMPs are synthesized by cancer cells and stromal cells
(39). The level of MMP7 in
peritoneal lavage was detected using RT-PCR and the results
demonstrated that MMP7 expression is associated with peritoneal
metastasis, for which the sensitivity was 53.3% (40). In addition, the expression of MMP14,
as detected using RT-PCR of peripheral blood and bone marrow
samples, is associated with peritoneal dissemination in gastric
cancer (41). MMPs are suppressed by
tissue inhibitor metalloproteinases (TIMPs) (42). Miyagi et al (43) transfected the TIMP-1 gene into gastric
cancer cells using adenoviral vectors and identified that the cell
invasion ability was reduced, as analysed using an in vitro
invasion assay, and the weight of the metastasis peritoneal nodes
was also significantly reduced in in vivo studies, compared
with those in the control group. Notably, when detecting TIMP-1
concentrations in tumour tissues using ELISA, high levels of TIMP-1
are associated with peritoneal dissemination and an unfavourable
prognosis (44). Consequently,
MMP7/MMP14 and TIMP-1 may be used as efficient markers to predict
peritoneal dissemination.
Gastric cancer cells require the ability to migrate
in order to invade the gastric wall and subperitoneal tissue.
Motility factors are expressed by cancer cells to promote cell
invasion and migration. c-MET, also known as hepatocyte growth
factor (HGF) receptor, is a membrane receptor that participates in
the development of certain types of disease, including cancer,
autism, and cardiac dysfunction (45). HGF is the only established ligand of
c-MET (46). High expression levels
of HGF and c-MET are associated with peritoneal dissemination and a
reduced median survival time (47).
Toiyama et al (47)
demonstrated that HGF promotes invasiveness, the migration of
gastric cancer cells and the inhibition of anoikis, which was
inhibited by the kinase inhibitor SU11274.
Brain-derived neurotrophic factor (BDNF) is part of
the neurotrophin family of growth factors and has been demonstrated
to bind to >2 receptors, including tropomyosin-related kinase B
(TrkB) and low affinity nerve growth factor receptor (48). BDNF expression levels in the invasive
cells of tumours are significantly increased compared with those in
the core cells and in the normal mucosa (49). In addition, co-expression of BDNF/TrkB
is associated with the prognosis of gastric cancer (49). This study has also confirmed that BDNF
promotes the proliferation, migration and invasion of gastric
cancer cells that express TrkB in vitro, and that the Trk
antagonist K252 significantly decreased the tumour size and the
number of peritoneal dissemination sites, compared with the control
group, in vivo (49).
When gastric cancer cells invade into subperitoneal
tissue, growth factors secreted by cancer cells and stromal cells,
including vascular endothelial growth factor (VEGF)-A and VEGF-C,
further promote cell proliferation and vascular neogenesis
(53).
It is well established that tumour tissues contain
cancer cells and cancer stroma (54).
Interactions between the cancer cells and cancer stroma induce the
processes of invasion and migration. The stroma is comprised of
numerous types of mesenchymal cells and extracellular matrix. Cells
of the cancer stroma include fibroblasts, immune cells, endothelial
cells and other types of bone marrow-derived cells (55). Carcinoma-associated fibroblasts
(CAFs), which consist of both fibroblasts and myofibroblasts, are
frequently observed in the stroma of human carcinomas (56). Myofibroblasts, as the form of
activated fibroblasts, are characterized by the overexpression of
α-smooth muscle actin (α-SMA) and vimentin, compared with
fibroblasts (56).
CAFs produce numerous types of growth factors,
cytokines, chemokines, MMPs and ECM proteins that promote the
progression of tumours. CAFs are able to stimulate angiogenesis via
the secretion of VEGF, promote tumour cell proliferation via
CXCL12, induce tumour cell invasion via TGF-β1 and decrease tumour
cell death via an anti-apoptosis factor, secreted frizzled-related
protein 1 (55).
As for the sources of CAFs, a previous study that
reported the origin of myofibroblasts in kidney fibrosis could
provide implications (57). The
results suggested that myofibroblasts arise from four sources,
proliferation of resident fibroblasts (50%), differentiation from
bone marrow (35%), and transition from endothelial (10%) and
epithelial (5%) forms (57). However,
the sources of CAFs in peritoneal metastasis nodes from gastric
cancer have yet to be fully elucidated. Fuyuhiro et al
(58) reported that the ratios of
myofibroblasts to fibroblasts in tumour tissue are significantly
higher compared with those in normal gastric tissue. Additionally,
the ability of gastric cancer cells to migrate and invade is
increased by CAFs, a function that may be attenuated using Smad2
siRNA and an anti-TGF-β antibody (58). These results indicated that CAFs have
a crucial role in the growth and invasion of gastric cancer cells
(58). Another previous study
verified that HPMCs are one source of CAFs (25). HPMCs have a spindle-shaped morphology
following the stimulation of TGF-β and these activated HPMCs are
more invasive. The molecular mechanisms by which PMCs are
incorporated into tumour structures have also been clarified
(59). A previous study demonstrated
that PMCs create the invasion front of peritoneal metastasis in a
manner dependent on the activation of a tyrosine kinase substrate
with five SH3 domains, a substrate of Src kinase (59). Furthermore, it has recently been
identified that CAFs originate from bone marrow-derived cells
(60). Bone marrow-derived cells that
adopt a fibroblast-like morphology in the circulation are referred
to as fibrocytes. Fibrocytes express the cell markers of leucocytes
(CD45) as well as the markers of mesenchymal cells (collagen I). In
addition, elevated levels of CXCL12 in gastric cancer cells and of
CXCR4 in fibrocytes can be detected, indicating that CXCL12/CXCR4
may participate in the migration of fibrocytes from the bone marrow
to gastric cancer sites (60).
Taken together, CAFs in gastric cancer originate
from orthotopic fibroblasts, HPMCs and bone marrow-derived cells,
and they promote the progression of gastric cancer in a number of
ways (25,58,60). The
CXCL12/CXCR4 axis may be required in the migration of CAFs. In the
tumour microenvironment, TGF-β1, which originates from cancer cells
and CAFs, has an important role in the interaction between cancer
cells and CAFs (61). TGF-β1 secreted
by cancer cells promotes the transition to myofibroblasts and, in
turn, myofibroblasts can secrete TGF-β1, which further stimulates
the proliferation of cancer cells (61).
An increasing number of studies have focused on the
underlying mechanisms by which CAFs promote the progression of
gastric cancer. The CAFs in gastric cancer have high expression
levels of HGF and are one of the sources of secreted HGF in the
microenvironment (62). HGF enhances
the proliferation and migration of gastric cancer cells via
paracrine signalling (62). A number
of ECM proteins secreted from CAFs also have a role in the
promotion of gastric cancer, including asporin and periostin
(63,64). Asporin belongs to a family of small
leucine-rich proteoglycans and is a primary non-collagen component
of ECM (65). Asporin is typically
secreted by CAFs into the microenvironment of gastric cancer
(63). Asporin in the
microenvironment promotes invasion of CAFs and surrounding cancer
cells through the CD44/Rac1 pathway (63). Periostin (POSTN), produced by
fibroblasts, participates in the adhesion and migration of
epithelial cells (66). Kikuchi et
al (64) reported that POSTN is
secreted by CAFs and enhances the proliferation of gastric cancer
cells via the activation of ERK. In vivo investigation has
further demonstrated that co-inoculation of gastric cancer cells
with fibroblasts expressing POSTN facilitates tumour formation
(64). The ECM proteins secreted by
CAFs provide an optimal microenvironment for gastric cancer.
Furthermore, a multitude of molecules, including growth,
transcription and inflammatory factors, are also highly expressed
in CAFs and stimulate the invasion of gastric cancer cells; these
include fibroblast growth factor 9 (67), Twist 1 (68), interleukin 22 (69), galectin-1 (70) and transgelin (71). Consequently, CAFs, as important
components of stromal cells, secrete numerous molecules that
further promote tumourigenesis.
Another metastatic process of peritoneal
dissemination is translymphatic metastasis. Lymphatic orifices,
also known as stomata, are the gates on the peritoneal surface to
the underside of capillary lymphatic vessels (72). Furthermore, there are gaps amongst the
mesothelial cells covering stomata; therefore, peritoneal-free
cancer cells are able to invade the submesothelial space through
these stomata (73). The lack of the
physical barrier presented by the compact HPMC monolayer indicates
that translymphatic metastasis does not require numerous stages,
unlike transmesothelial metastasis. Therefore translymphatic
metastasis is frequently developed at an earlier stage compared
with transmesothelial metastasis (73). Stomata are primarily located in the
greater omentum, diaphragm, small bowel mesentery, pelvic
peritoneum and falciform ligament, whilst the anterior abdominal
wall, liver capsule, surface of the gastric wall and small bowel do
not typically have lymphatic orifices (7). Hagiwara et al (73) initially identified milky spots on the
peritoneum and malignant cells were revealed to implant into these
sites. Milky spots are located in the greater omentum, mesenterium
and pelvic floor in humans (7).
However, there are no milky spots on the anterior parietal
peritoneum, liver capsule or serosal surface of the small bowel
(73). Therefore, these peritoneal
regions are not invaded until the late stages of peritoneal
metastasis with massive malignant ascites (73). Milky spots are specific to lymphoid
tissue on the peritoneum and consist primarily of macrophages, as
well as a number of lymphocytes and mastocytes (74). There are numerous lymphatic orifices
on the surface of milky spots and macrophages migrate into the
peritoneal cavity via these stomata (74).
Recently, the C-C motif chemokine ligand 22
(CCL22)/CC chemokine receptor 4 (CCR4) axis has been the focus of
more studies as it participates in the infiltration of gastric
cancer cells into milky spots. A previous study established that
gastric cancer cells express CCR4, and that CCL22 may also be
detected within omental milky spots (80). CCL22 increases the proliferation and
migration of gastric cancer cells in vitro (80). These results indicate that CCL22/CCR4
may participate in this selective infiltration process (80). Furthermore, milky spots are rich in
capillaries and the surrounding vascular density is high (81). CD105+ vessels and vascular
sprouts have been also located within milky spots, indicating that
angiogenesis is active (81).
Additionally, milky spots also serves as a hypoxic niche, which
enhances the self-renewing capacity of gastric cancer
stem/progenitor cells regulated by hypoxia-inducible factor-1α
(HIF-1α) (82).
Consequently, milky spots, as a lymphatic apparatus,
participate in the immune defence of the peritoneal cavity.
However, stomata on the milky spots provide a gate via which
peritoneal-free cancer cells may migrate into the submesothelial
space.
Microarray analysis has been used to systematically
search aberrantly expressed microRNA (miRNA) that is associated
with human cancer. miRNAs are small non-coding RNAs that have a
role in the post-transcriptional regulation of gene expression. An
increasing number of studies have demonstrated that miRNAs are
associated with tumour development and progression (83,84).
Certain miRNAs have been established to be associated with
peritoneal dissemination in gastric cancer (85–87). Zheng
et al (85) reported that
microRNA (miR)-409-3p inhibited the ability of gastric cancer cells
to invade and metastasise via targeting radixin. Another previous
study demonstrated that miR-495 and miR-551a suppressed gastric
cancer cell migration and invasion by targeting PRL-3, which
exhibits significantly increased expression levels in primary
gastric cancer cells with peritoneal dissemination (86). miR-25 was identified to enhance
gastric cancer progression by targeting Erb-B2 receptor tyrosine
kinase 2 both in vitro and in vivo (87). Other miRNAs have been reported to
correlate with peritoneal dissemination in gastric cancer,
including miR-200b (88), miR-125a-3q
(89) and let-7 miRNAs (90). These studies, regarding miRNAs,
present a novel therapeutic target for suppressing peritoneal
dissemination in gastric cancer. Takei et al (91) established an orthotopic implantation
model for the peritoneal metastasis of gastric cancer and
demonstrated that inoculation with mi-516a-3q cells resulted in
significantly increased survival periods, compared with inoculation
with control cells.
Paget's ‘seed and soil’ theory is the mainstream
theory for the formation of peritoneal dissemination in gastric
cancer. However, a number of studies have questioned the validity
of this theory and have proposed certain alternative mechanisms to
explain the progress of peritoneal dissemination. Miyake et
al (92) suggested a hypothesis
wherein peritoneal dissemination is a type of transvessel
metastasis mediated by HIF-1α. HIF-1α is a subunit of the
heterodimeric transcription factor HIF-1 and is an important
regulator of the cellular response to hypoxia. Recent studies have
implied that hypoxia participates in each step of the cancer
metastasis cascade, including EMT, invasion, intravasation, homing
and proliferation to secondary organs (93–95). All
of these results have indicated that HIF-1α may be a vital
regulator in cancer metastasis. Miyake et al (92) established orthotopic implantation
(o.i.) and conventional intraperitoneal injection (i.p.) models to
mimic the peritoneal dissemination of gastric cancer. Results
showed that HIF-1α was crucial for the development of peritoneal
dissemination in the o.i. model, whereas it had an inhibitory role
in the peritoneal dissemination in the i.p. model (92). These results confirm the angiogenesis
function of HIF-1α and indicate the ‘seeding’ theory may not be
correct (92). In addition, another
previous study has suggested that the subperitoneal space comprises
blood vessels, the lymphatics, lymph nodes and fatty tissue, which
provide an important passageway for cancer cells within the
peritoneal cavity (96). This
previous study supports the transvessel metastasis hypothesis to a
certain extent. However, further studies are required to validate
the accuracy of this theory.
A number of previous studies have focused on the
molecules involved in the peritoneal dissemination of gastric
cancer, and each step of the process demonstrates the stimulation
of various molecules, including growth factors, cytokines and
chemokines. Furthermore, microarray analysis has been increasingly
used to systematically identify metastasis-associated genes,
including their associated mRNAs and miRNAs. Additionally, it may
be possible to predict, prevent and treat peritoneal metastasis by
targeting these molecules. Previous studies have primarily focused
on the characteristics of malignant cancer cells, but an increasing
number of studies have focused on the tumour microenvironment,
particularly milky spots and the tumour stroma (54,64). Milky
spots are specific sites for peritoneal metastasis and numerous
studies have demonstrated that milky spots are the first metastasis
location for free cancer cells. Stroma, as a component of tumour
tissue, interacts with cancer cells and promotes cell invasion and
migration. However, the most frequently used peritoneal
dissemination model was established using the i.p. model of gastric
cancer cells. This model does not include the initial steps of
dissemination, such as invasion in the gastric wall and the
detachment from the primary tumour. Therefore, a more persuasive
orthotopic implantation model would mimic the formation of
peritoneal dissemination. The underlying mechanisms of peritoneal
metastasis remain in dispute. The transvessel metastasis theory has
been recently proposed by Miyake et al (92) and provides more information and the
novel therapeutic target HIF-1α. However, further investigation is
required in order to validate this hypothesis.
The present study was supported by the National
Natural Science Foundation of China (grant no. 81372364).
|
1
|
Stewart BW and Wild CP: World cancer
report 2014. World. 2014.
|
|
2
|
Japanese Gastric Cancer Association
Registration Committee, . Maruyama K, Kaminishi M, Hayashi K, Isobe
Y, Honda I, Katai H, Arai K, Kodera Y and Nashimoto A: Gastric
cancer treated in 1991 in Japan: Data analysis of nationwide
registry. Gastric Cancer. 9:51–66. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Maehara Y, Hasuda S, Koga T, Tokunaga E,
Kakeji Y and Sugimachi K: Postoperative outcome and sites of
recurrence in patients following curative resection of gastric
cancer. Br J Surg. 87:353–357. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yashiro M, Chung YS, Nishimura S, Inoue T
and Sowa M: Fibrosis in the peritoneum induced by scirrhous gastric
cancer cells may act as ‘soil’ for peritoneal dissemination.
Cancer. 77 8 Suppl:S1668–S1675. 1996. View Article : Google Scholar
|
|
5
|
Li Z, Miao Z, Jin G, Li X, Li H, Lv Z and
Xu HM: big-h3 supports gastric cancer cell adhesion, migration and
proliferation in peritoneal carcinomatosis. Mol Med Rep. 6:558–564.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Paget S: The distribution of secondary
growths in cancer of the breast. Cancer Metastasis Rev. 8:98–101.
1989.PubMed/NCBI
|
|
7
|
Yonemura Y, Kawamura T, Bandou E,
Tsukiyama G, Endou Y and Miura M: The natural history of free
cancer cells in the peritoneal cavity. Recent Results Cancer Res.
169:11–23. 2007.PubMed/NCBI
|
|
8
|
Bando E, Yonemura Y, Takeshita Y,
Taniguchi K, Yasui T, Yoshimitsu Y, Fushida S, Fujimura T,
Nishimura G and Miwa K: Intraoperative lavage for cytological
examination in 1,297 patients with gastric carcinoma. Am J Surg.
178:256–262. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yonemura Y, Endou Y, Kimura K, Fushida S,
Bandou E, Taniguchi K, Kinoshita K, Ninomiya I, Sugiyama K,
Heizmann CW, et al: Inverse expression of S100A4 and E-cadherin is
associated with metastatic potential in gastric cancer. Clin Cancer
Res. 6:4234–4242. 2000.PubMed/NCBI
|
|
10
|
Marutsuka T, Shimada S, Shiomori K,
Hayashi N, Yagi Y, Yamane T and Ogawa M: Mechanisms of peritoneal
metastasis after operation for non-serosa-invasive gastric
carcinoma: An ultrarapid detection system for intraperitoneal free
cancer cells and a prophylactic strategy for peritoneal metastasis.
Clin Cancer Res. 9:678–685. 2003.PubMed/NCBI
|
|
11
|
Takebayashi K, Murata S, Yamamoto H,
Ishida M, Yamaguchi T, Kojima M, Shimizu T, Shiomi H, Sonoda H,
Naka S, et al: Surgery-induced peritoneal cancer cells in patients
who have undergone curative gastrectomy for gastric cancer. Ann
Surg Oncol. 21:1991–1997. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zlotnik A: Chemokines and cancer. Int J
Cancer. 119:2026–2029. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tanaka T, Bai Z, Srinoulprasert Y, Yang
BG, Hayasaka H and Miyasaka M: Chemokines in tumor progression and
metastasis. Cancer Sci. 96:317–322. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yoshie O: Immune chemokines and their
receptors: The key elements in the genesis, homeostasis and
function of the immune system. Springer Semin Immunopathol.
22:371–391. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yasumoto K, Koizumi K, Kawashima A, Saitoh
Y, Arita Y, Shinohara K, Minami T, Nakayama T, Sakurai H, Takahashi
Y, et al: Role of the CXCL12/CXCR4 axis in peritoneal
carcinomatosis of gastric cancer. Cancer Res. 66:2181–2187. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Müller A, Homey B, Soto H, Ge N, Catron D,
Buchanan ME, McClanahan T, Murphy E, Yuan W, Wagner SN, et al:
Involvement of chemokine receptors in breast cancer metastasis.
Nature. 410:50–56. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Koizumi K, Hojo S, Akashi T, Yasumoto K
and Saiki I: Chemokine receptors in cancer metastasis and cancer
cell-derived chemokines in host immune response. Cancer Sci.
98:1652–1658. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lv ZD, Na D, Liu FN, Du ZM, Sun Z, Li Z,
Ma XY, Wang ZN and Xu HM: Induction of gastric cancer cell adhesion
through transforming growth factor-beta1-mediated peritoneal
fibrosis. J Exp Clin Cancer Res. 29:1392010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kiyasu Y, Kaneshima S and Koga S:
Morphogenesis of peritoneal metastasis in human gastric cancer.
Cancer Res. 41:1236–1239. 1981.PubMed/NCBI
|
|
20
|
Lamouille S, Xu J and Derynck R: Molecular
mechanisms of epithelial-mesenchymal transition. Nat Rev Mol Cell
Biol. 15:178–196. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelial-mesenchymal transitions in development and disease.
Cell. 139:871–890. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Margetts PJ, Oh KH and Kolb M:
Transforming growth factor-beta: Importance in long-term peritoneal
membrane changes. Perit Dial Int. 25 Suppl 3:S15–S17.
2005.PubMed/NCBI
|
|
23
|
Massagué J: TGFβ signalling in context.
Nat Rev Mol Cell Biol. 13:616–630. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gordon KJ and Blobe GC: Role of
transforming growth factor-beta superfamily signaling pathways in
human disease. Biochim Biophys Acta. 1782:197–228. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tsukada T, Fushida S, Harada S, Yagi Y,
Kinoshita J, Oyama K, Tajima H, Fujita H, Ninomiya I, Fujimura T
and Ohta T: The role of human peritoneal mesothelial cells in the
fibrosis and progression of gastric cancer. Int J Oncol.
41:476–482. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu Q, Mao H, Nie J, Chen W, Yang Q, Dong
X and Yu X: Transforming growth factor {beta}1 induces
epithelial-mesenchymal transition by activating the JNK-Smad3
pathway in rat peritoneal mesothelial cells. Perit Dial Int. 28
Suppl 3:S88–S95. 2008.PubMed/NCBI
|
|
27
|
Lv ZD, Wang HB, Li FN, Wu L, Liu C, Nie G,
Kong B, Qu HL and Li JG: TGF-β1 induces peritoneal fibrosis by
activating the Smad2 pathway in mesothelial cells and promotes
peritoneal carcinomatosis. Int J Mol Med. 29:373–379.
2012.PubMed/NCBI
|
|
28
|
Jiang CG, Lv L, Liu FR, Wang ZN, Na D, Li
F, Li JB, Sun Z and Xu HM: Connective tissue growth factor is a
positive regulator of epithelial-mesenchymal transition and
promotes the adhesion with gastric cancer cells in human peritoneal
mesothelial cells. Cytokine. 61:173–180. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Okazaki M, Fushida S, Harada S, Tsukada T,
Kinoshita J, Oyama K, Tajima H, Ninomiya I, Fujimura T and Ohta T:
The angiotensin II type 1 receptor blocker candesartan suppresses
proliferation and fibrosis in gastric cancer. Cancer Lett.
355:46–53. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Shinbo T, Fushida S, Tsukada T, Harada S,
Kinoshita J, Oyama K, Okamoto K, Ninomiya I, Takamura H, Kitagawa
H, et al: Protein-bound polysaccharide K suppresses tumor fibrosis
in gastric cancer by inhibiting the TGF-b signaling pathway. Oncol
Rep. 33:553–558. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tanaka H, Muguruma K, Ohira M, Kubo N,
Yamashita Y, Maeda K, Sawada T and Hirakawa K: Impact of adjuvant
immunochemotherapy using protein-bound polysaccharide-K on overall
survival of patients with gastric cancer. Anticancer Res.
32:3427–3433. 2012.PubMed/NCBI
|
|
32
|
Ono Y, Hayashida T, Konagai A, Okazaki H,
Miyao K, Kawachi S, Tanabe M, Shinoda M, Jinno H, Hasegawa H, et
al: Direct inhibition of the transforming growth factor-β pathway
by protein-bound polysaccharide through inactivation of Smad2
signaling. Cancer Sci. 103:317–324. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Nakashio T, Narita T, Akiyama S, Kasai Y,
Fujiwara M, Ito K, Takagi H and Kanngi R: Adhesion of human gastric
and pancreatic cancer cells to peritoneal mesothelial cells is
mediated by CD44 and beta(1) integrin. Int J Oncol. 10:183–188.
1997.PubMed/NCBI
|
|
34
|
Takatsuki H, Komatsu S, Sano R, Takada Y
and Tsuji T: Adhesion of gastric carcinoma cells to peritoneum
mediated by alpha3beta1 integrin (VLA-3). Cancer Res. 64:6065–6070.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Giancotti FG and Ruoslahti E: Integrin
signaling. Science. 285:1028–1032. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Fukuda K, Saikawa Y, Yagi H, Wada N,
Takahashi T and Kitagawa Y: Role of integrin a1 subunits in gastric
cancer patients with peritoneal dissemination. Mol Med Rep.
5:336–340. 2012.PubMed/NCBI
|
|
37
|
Nishimura S, Chung YS, Yashiro M, Inoue T
and Sowa M: Role of alpha 2 beta 1- and alpha 3 beta 1-integrin in
the peritoneal implantation of scirrhous gastric carcinoma. Br J
Cancer. 74:1406–1412. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chen CN, Chang CC, Lai HS, Jeng YM, Chen
CI, Chang KJ, Lee PH and Lee H: Connective tissue growth factor
inhibits gastric cancer peritoneal metastasis by blocking integrin
a3b1-dependent adhesion. Gastric Cancer. 18:504–515. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yonemura Y, Endo Y, Fujita H, Kimura K,
Sugiyama K, Momiyama N, Shimada H and Sasaki T: Inhibition of
peritoneal dissemination in human gastric cancer by MMP-7-specific
antisense oligonucleotide. J Exp Clin Cancer Res. 20:205–212.
2001.PubMed/NCBI
|
|
40
|
Li Z, Zhang D, Zhang H, Miao Z, Tang Y,
Sun G and Dai D: Prediction of peritoneal recurrence by the mRNA
level of CEA and MMP-7 in peritoneal lavage of gastric cancer
patients. Tumour Biol. 35:3463–3470. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Mimori K, Fukagawa T, Kosaka Y, Ishikawa
K, Iwatsuki M, Yokobori T, Hirasaki S, Takatsuno Y, Sakashita H,
Ishii H, et al: A large-scale study of MT1-MMP as a marker for
isolated tumor cells in peripheral blood and bone marrow in gastric
cancer cases. Ann Surg Oncol. 15:2934–2942. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Visse R and Nagase H: Matrix
metalloproteinases and tissue inhibitors of metalloproteinases:
Structure, function, and biochemistry. Circ Res. 92:827–839. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Miyagi M, Aoyagi K, Kato S and Shirouzu K:
The TIMP-1 gene transferred through adenovirus mediation shows a
suppressive effect on peritoneal metastases from gastric cancer.
Int J Clin Oncol. 12:17–24. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Yoshikawa T, Tsuburaya A, Kobayashi O,
Sairenji M, Motohashi H, Yanoma S and Noguchi Y: Intratumoral
concentrations of tissue inhibitor of matrix metalloproteinase 1 in
patients with gastric carcinoma a new biomartker for invasion and
its impact on survival. Cancer. 91:1739–1744. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Gentile A, Trusolino L and Comoglio PM:
The met tyrosine kinase receptor in development and cancer. Cancer
Metastasis Rev. 27:85–94. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Trusolino L, Bertotti A and Comoglio PM:
MET signalling: Principles and functions in development, organ
regeneration and cancer. Nat Rev Mol Cell Biol. 11:834–848. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Toiyama Y, Yasuda H, Saigusa S, Matushita
K, Fujikawa H, Tanaka K, Mohri Y, Inoue Y, Goel A and Kusunoki M:
Co-expression of hepatocyte growth factor and c-Met predicts
peritoneal dissemination established by autocrine hepatocyte growth
factor/c-Met signaling in gastric cancer. Int J Cancer.
130:2912–2921. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Binder DK and Scharfman HE: Brain-derived
neurotrophic factor. Growth Factors. 22:123–131. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Okugawa Y, Tanaka K, Inoue Y, Kawamura M,
Kawamoto A, Hiro J, Saigusa S, Toiyama Y, Ohi M, Uchida K, et al:
Brain-derived neurotrophic factor/tropomyosin-related kinase B
pathway in gastric cancer. Br J Cancer. 108:121–130. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Perretti M and D'Acquisto F: Annexin A1
and glucocorticoids as effectors of the resolution of inflammation.
Nat Rev Immunol. 9:62–70. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Leoni G, Alam A, Neumann PA, Lambeth JD,
Cheng G, McCoy J, Hilgarth RS, Kundu K, Murthy N, Kusters D, et al:
Annexin A1, formyl peptide receptor, and NOX1 orchestrate
epithelial repair. J Clin Invest. 123:443–454. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Cheng TY, Wu MS, Lin JT, Lin MT, Shun CT,
Huang HY, Hua KT and Kuo ML: Annexin A1 is associated with gastric
cancer survival and promotes gastric cancer cell invasiveness
through the formyl peptide receptor/extracellular signal-regulated
kinase/integrin beta-1-binding protein 1 pathway. Cancer.
118:5757–5767. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Aoyagi K, Kouhuji K, Yano S, Miyagi M,
Imaizumi T, Takeda J and Shirouzu K: VEGF significance in
peritoneal recurrence from gastric cancer. Gastric Cancer.
8:155–163. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Mueller MM and Fusenig NE: Friends or
foes-bipolar effects of the tumour stroma in cancer. Nat Rev
Cancer. 4:839–849. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Shimoda M, Mellody KT and Orimo A:
Carcinoma-associated fibroblasts are a rate-limiting determinant
for tumour progression. Semin Cell Dev Biol. 21:19–25. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Gabbiani G, Ryan GB and Majne G: Presence
of modified fibroblasts in granulation tissue and their possible
role in wound contraction. Experientia. 27:549–550. 1971.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
LeBleu VS, Taduri G, O'Connell J, Teng Y,
Cooke VG, Woda C, Sugimoto H and Kalluri R: Origin and function of
myofibroblasts in kidney fibrosis. Nat Med. 19:1047–1053. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Fuyuhiro Y, Yashiro M, Noda S, Matsuoka J,
Hasegawa T, Kato Y, Sawada T and Hirakawa K: Cancer-associated
orthotopic myofibroblasts stimulates the motility of gastric
carcinoma cells. Cancer Sci. 103:797–805. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Satoyoshi R, Aiba N, Yanagihara K, Yashiro
M and Tanaka M: Tks5 activation in mesothelial cells creates
invasion front of peritoneal carcinomatosis. Oncogene.
34:3176–3187. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Terai S, Fushida S, Tsukada T, Kinoshita
J, Oyama K, Okamoto K, Makino I, Tajima H, Ninomiya I, Fujimura T,
et al: Bone marrow derived ‘fibrocytes’ contribute to tumor
proliferation and fibrosis in gastric cancer. Gastric Cancer.
18:306–313. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Ostman A and Augsten M: Cancer-associated
fibroblasts and tumor growth-bystanders turning into key players.
Curr Opin Genet Dev. 19:67–73. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Wu X, Chen X, Zhou Q, Li P, Yu B, Li J, Qu
Y, Yan J, Yu Y, Yan M, et al: Hepatocyte growth factor activates
tumor stromal fibroblasts to promote tumorigenesis in gastric
cancer. Cancer Lett. 335:128–135. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Satoyoshi R, Kuriyama S, Aiba N, Yashiro M
and Tanaka M: Asporin activates coordinated invasion of scirrhous
gastric cancer and cancer-associated fibroblasts. Oncogene.
34:650–660. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Kikuchi Y, Kunita A, Iwata C, Komura D,
Nishiyama T, Shimazu K, Takeshita K, Shibahara J, Kii I, Morishita
Y, et al: The niche component periostin is produced by
cancer-associated fibroblasts, supporting growth of gastric cancer
through ERK activation. Am J Pathol. 184:859–870. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Nakajima M, Kizawa H, Saitoh M, Kou I,
Miyazono K and Ikegawa S: Mechanisms for asporin function and
regulation in articular cartilage. J Biol Chem. 282:32185–32192.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Liu Y and Liu BA: Enhanced proliferation,
invasion, and epithelial-mesenchymal transition of
nicotine-promoted gastric cancer by periostin. World J
Gastroenterol. 17:2674–2680. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Sun C, Fukui H, Hara K, Zhang X, Kitayama
Y, Eda H, Tomita T, Oshima T, Kikuchi S, Watari J, et al: FGF9 from
cancer-associated fibroblasts is a possible mediator of invasion
and anti-apoptosis of gastric cancer cells. BMC Cancer. 15:3332015.
View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Sung CO, Lee KW, Han S and Kim SH: Twist1
is up-regulated in gastric cancer-associated fibroblasts with poor
clinical outcomes. Am J Pathol. 179:1827–1838. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Fukui H, Zhang X, Sun C, Hara K, Kikuchi
S, Yamasaki T, Kondo T, Tomita T, Oshima T, Watari J, et al: IL-22
produced by cancer-associated fibroblasts promotes gastric cancer
cell invasion via STAT3 and ERK signaling. Br J Cancer.
111:763–771. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
He XJ, Tao HQ, Hu ZM, Ma YY, Xu J, Wang
HJ, Xia YJ, Li L, Fei BY, Li YQ and Chen JZ: Expression of
galectin-1 in carcinoma-associated fibroblasts promotes gastric
cancer cell invasion through upregulation of integrin b1. Cancer
Sci. 105:1402–1410. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Yu B, Chen X, Li J, Qu Y, Su L, Peng Y,
Huang J, Yan J, Yu Y, Gu Q, et al: Stromal fibroblasts in the
microenvironment of gastric carcinomas promote tumor metastasis via
upregulating TAGLN expression. BMC Cell Biol. 14:172013. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Shimotsuma M, Shields JW, Simpson-Morgan
MW, Sakuyama A, Shirasu M, Hagiwara A and Takahashi T:
Morpho-physiological function and role of omental milky spots as
omentum-associated lymphoid tissue (OALT) in the peritoneal cavity.
Lymphology. 26:90–101. 1993.PubMed/NCBI
|
|
73
|
Hagiwara A, Takahashi T, Sawai K,
Taniguchi H, Shimotsuma M, Okano S, Sakakura C, Tsujimoto H, Osaki
K, Sasaki S, et al: Milky spots as the implantation site for
malignant cells in peritoneal dissemination in mice. Cancer Res.
53:687–692. 1993.PubMed/NCBI
|
|
74
|
Beelen RH, Fluitsma DM and Hoefsmit EC:
The cellular composition of omentum milky spots and the
ultrastructure of milky spot macrophages and reticulum cells. J
Reticuloendothel Soc. 28:585–599. 1980.PubMed/NCBI
|
|
75
|
Liebermann-Meffert D: The greater omentum.
Anatomy, embryology, and surgical applications. Surg Clin North Am.
80(275–293): xii2000.
|
|
76
|
Oosterling SJ, van der Bij GJ, Bögels M,
van der Sijp JR, Beelen RH, Meijer S and van Egmond M: Insufficient
ability of omental milky spots to prevent peritoneal tumor
outgrowth supports omentectomy in minimal residual disease. Cancer
Immunol Immunother. 55:1043–1051. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Cui L, Johkura K, Liang Y, Teng R, Ogiwara
N, Okouchi Y, Asanuma K and Sasaki K: Biodefense function of
omental milky spots through cell adhesion molecules and leukocyte
proliferation. Cell Tissue Res. 310:321–330. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Liu XY, Miao ZF, Zhao TT, Wang ZN, Xu YY,
Gao J, Wu JH, You Y, Xu H and Xu HM: Milky spot macrophages
remodeled by gastric cancer cells promote peritoneal mesothelial
cell injury. Biochem Biophys Res Commun. 439:378–383. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Bingle L, Brown NJ and Lewis CE: The role
of tumour-associated macrophages in tumour progression:
Implications for new anticancer therapies. J Pathol. 196:254–265.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Cao L, Hu X, Zhang J, Huang G and Zhang Y:
The role of the CCL22-CCR4 axis in the metastasis of gastric cancer
cells into omental milky spots. J Transl Med. 12:2672014.
View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Gerber SA, Rybalko VY, Bigelow CE, Lugade
AA, Foster TH, Frelinger JG and Lord EM: Preferential attachment of
peritoneal tumor metastases to omental immune aggregates and
possible role of a unique vascular microenvironment in metastatic
survival and growth. Am J Pathol. 169:1739–1752. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Miao ZF, Wang ZN, Zhao TT, Xu YY, Gao J,
Miao F and Xu HM: Peritoneal milky spots serve as a hypoxic niche
and favor gastric cancer stem/progenitor cell peritoneal
dissemination through hypoxia-inducible factor 1a. Stem Cells.
32:3062–3074. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Esquela-Kerscher A and Slack FJ: Oncomirs
- microRNAs with a role in cancer. Nat Rev Cancer. 6:259–269. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Zhang B, Pan X, Cobb GP and Anderson TA:
microRNAs as oncogenes and tumor suppressors. Dev Biol. 302:1–12.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Zheng B, Liang L, Huang S, Zha R, Liu L,
Jia D, Tian Q, Wang Q, Wang C, Long Z, et al: MicroRNA-409
suppresses tumour cell invasion and metastasis by directly
targeting radixin in gastric cancers. Oncogene. 31:4509–4516. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Li Z, Cao Y, Jie Z, Liu Y, Li Y, Li J, Zhu
G, Liu Z, Tu Y, Peng G, et al: miR-495 and miR-551a inhibit the
migration and invasion of human gastric cancer cells by directly
interacting with PRL-3. Cancer Lett. 323:41–47. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Li BS, Zuo QF, Zhao YL, Xiao B, Zhuang Y,
Mao XH, Wu C, Yang SM, Zeng H, Zou QM and Guo G: MicroRNA-25
promotes gastric cancer migration, invasion and proliferation by
directly targeting transducer of ERBB2, 1 and correlates with poor
survival. Oncogene. 34:2556–2565. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Kurashige J, Mima K, Sawada G, Takahashi
Y, Eguchi H, Sugimachi K, Mori M, Yanagihara K, Yashiro M, Hirakawa
K, et al: Epigenetic modulation and repression of miR-200b by
cancer-associated fibroblasts contribute to cancer invasion and
peritoneal dissemination in gastric cancer. Carcinogenesis.
36:133–141. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Hashiguchi Y, Nishida N, Mimori K, Sudo T,
Tanaka F, Shibata K, Ishii H, Mochizuki H, Hase K, Doki Y and Mori
M: Down-regulation of miR-125a-3p in human gastric cancer and its
clinicopathological significance. Int J Oncol. 40:1477–1482.
2012.PubMed/NCBI
|
|
90
|
Ohshima K, Inoue K, Fujiwara A, Hatakeyama
K, Kanto K, Watanabe Y, Muramatsu K, Fukuda Y, Ogura S, Yamaguchi K
and Mochizuki T: Let-7 microRNA family is selectively secreted into
the extracellular environment via exosomes in a metastatic gastric
cancer cell line. PLoS One. 5:e132472010. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Takei Y, Takigahira M, Mihara K, Tarumi Y
and Yanagihara K: The metastasis-associated microRNA miR-516a-3p is
a novel therapeutic target for inhibiting peritoneal dissemination
of human scirrhous gastric cancer. Cancer Res. 71:1442–1453. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Miyake S, Kitajima Y, Nakamura J, Kai K,
Yanagihara K, Tanaka T, Hiraki M, Miyazaki K and Noshiro H: HIF-1a
is a crucial factor in the development of peritoneal dissemination
via natural metastatic routes in scirrhous gastric cancer. Int J
Oncol. 43:1431–1440. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Krishnamachary B, Zagzag D, Nagasawa H,
Rainey K, Okuyama H, Baek JH and Semenza GL: Hypoxia-inducible
factor-1-dependent repression of E-cadherin in von Hippel-Lindau
tumor suppressor-null renal cell carcinoma mediated by TCF3,
ZFHX1A, and ZFHX1B. Cancer Res. 66:2725–2731. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Funasaka T and Raz A: The role of
autocrine motility factor in tumor and tumor microenvironment.
Cancer Metastasis Rev. 26:725–735. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Lu X and Kang Y: Hypoxia and
hypoxia-inducible factors: Master regulators of metastasis. Clin
Cancer Res. 16:5928–5935. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Silverman PM: The subperitoneal space:
Mechanisms of tumour spread in the peritoneal cavity, mesentery,
and omentum. Cancer Imaging. 4:25–29. 2003. View Article : Google Scholar : PubMed/NCBI
|