Introduction
Astrocytoma is the most common primary tumor of the
central nervous system. The tumor malignancy has been classified
according to the histopathological and clinical criteria
established by the World Health Organization (WHO). WHO grade IV
astrocytoma (glioblastoma; GBM), which is the most invasive form,
has a poor prognosis (1). The median
survival time of patients with GBM following standard radiation and
chemotherapy is 15 months (2)
Under physiological conditions, ATP co-exists with
the classical neurotransmitters in vacuoles of synapses, and is
released into the extracellular space during signal transmission
(3). Exonucleases rapidly degrade
extracellular ATP to maintain low physiological concentrations (nM
level) (4). However, in a variety of
pathological conditions, particularly in tumors, tissue damage
accompanied with invasive growth or following surgical removal,
chemotherapy and radiotherapy, causes ATP to be released in large
quantities from the damaged membranes or directly via toxic
transportation (5). The rapid
increase in the levels of extracellular ATP (mM level) (5) and activation of purinergic receptors on
cell membrane (6) trigger a variety
of biological effects.
p2X purinoceptor (P2X7R), activated by
high extracellular ATP concentrations, is an ion channel purinergic
receptor (7). It is activated into a
trimer and forms a membrane pore measuring 4 µm in diameter to
allow the passage of 400–900 D ions, including Ca2+,
K+ and Na+ (8).
The inflow of a large number of basic ions, particularly
Ca2+, leads to mitochondrial damage (9), and activates caspases 9, 7 and 3, which
mediate cellular apoptosis (10–12).
P2X7R-mediated apoptosis controls cell growth under
physiological conditions. P2X7R expression in tumors has
attracted considerable attention for its unique biological
features, since defective apoptosis serves an important role in the
development of cancers. In the present study, the expression of
P2X7R in astrocytoma was determined to elucidate the
mechanisms underlying tumorigenesis and development of
astrocytoma.
Materials and methods
Tumor specimens
Human astrocytoma samples were obtained from the
Department of Neuropathology, Huashan Hospital, Fudan University
(Shanghai, China) for use in this retrospective study.
Paraffin-embedded tumor samples (from 100 individuals) included
diffuse astrocytomas (grade II; 26 cases), anaplastic astrocytomas
(grade III; 28 cases) and GBMs (grade IV; 46 cases). The median age
± standard deviation of the patients was 42.67±17.33 (range, 43–79)
years, with a male to female ratio of 3:2 (male, 60 cases; female,
40 cases). Fresh tissues included 7 cases of GBM tumors and the
surrounding peripheral brain tissue. All the samples were acquired
from individuals who had not received chemotherapy or radiotherapy
prior to surgical resection. The procedures associated with the
acquisition of samples from human subjects were approved by the
Ethics Committee of Fudan University.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA from glioma samples were extracted using
the QIAamp® RNA Mini kit (Qiagen China Co., Ltd.,
Shanghai, China) according to the manufacturer's protocol. For the
reverse transcription reaction, Takara PrimeScript™ RT Master mix
(cat. no. RR036A; Takara Biotechnology Co., Ltd., Dalian, China)
was used according to the manufacturer's protocol. qPCR was then
performed using SYBR Premix Ex Taq™ (Takara Biotechnology Co.,
Ltd.) according to the manufacturer's protocol. Equal amounts of
each cDNA (100 ng) were amplified in at least 35 cycles of 30 sec
at 95°C, 30 sec at 58°C, and 1 min at 72°C. Subsequent to qPCR, a
melting curve analysis was performed by gradually increasing the
temperature to 95°C. Data acquisition was performed during the
elongation step. The following primers were used:
P2X7R forward,
5′-TTTAAGCTTATGCCGGCCTGCTGC-AGCTG-3′ and reverse,
5′-TTTTTGCGGCCGCTCAGTAAGGACTCT-TGAAGCC-3′; GAPDH forward,
5′-GCACCGTCAAGGCTGAGAAC-3′ and reverse, 5′-TGGTGAAGACGCCAGTGGA-3′.
GAPDH was used as an endogenous control. Relative quantification
was calculated using the 2−ΔΔCq method (13). There were ≥3 replicates performed of
each qPCR.
DNA methylation of human glioma
samples
Genomic DNA was isolated from fresh tissue using the
QIAamp DNA Mini kit (Qiagen China Co., Ltd.) according to the
manufacturer's protocol. Bisulfite treatment of genomic DNA was
performed using the EpiTect Bisulfite kit (Qiagen China Co., Ltd.)
according to the manufacturer's protocol. The
cytosine-phosphodiester-guanosine (CpG) island located in the
+26/+573 nt region of P2X7R was determined using the
Sequenom MassARRAY® system (Sequenom, San Diego, CA,
USA) using the following primers: Forward,
5′-AGGAAGAGAGTATTTTTGTGTAGGTATTTGGGGG-3′ and reverse,
5′-CAGTAATACGACTCACTATAGGGAGAAGGCTACATAATAACAACCTCCCTCCCTAC-3′. The
PCR products were directly sequenced using an ABI BigDye Terminator
Cycle Sequencing kit (PE Applied Biosystems; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) on an ABI 3730 DNA sequencer
(Applied Biosystems; Thermo Fisher Scientific, Inc.) according to
the manufacturer's protocol. At the corresponding CpG site, the
sequencing trace was read as fully or partially methylated (C) and
unmethylated (q). Quantitative analysis of CpG methylation was
performed using MassCLEAVE base-specific cleavage combined with
matrix-assisted laser-desorption ionization-time-of-flight mass
spectrometry using EpiTyper software version 4.0 (Sequenom).
Western blot analysis
Tissues were lysed on ice in
radioimmunoprecipitation assay buffer (Tris 50 mM, NaCl 0.15 nM,
EDTA 10 mM pH 7.4, β-mercaptoethanol 0.1%, Tween-20 0.1% and
anti-protease cocktail 1:100) with protease inhibitors and
quantified using the bicinchoninic acid method. Protein lysates (50
µg) were resolved using 10% SDS-PAGE and electrotransferred to
polyvinylidene fluoride membranes (EMD Millipore, Billerica, MA,
USA). Membranes were blocked for 3 h at room temperature with 5%
skimmed milk in Tris-buffered saline with Tween-20 prior to
immunoblotting overnight at 4°C with anti-P2X7R (cat.
no. ab93354, Abcam, Cambridge, UK; dilution, 1:300) or anti-β actin
(clone AC-40; dilution, 1:50,000; Sigma-Aldrich; Merck KGaA),
followed by treatment with the respective secondary antibodies 1 h
at room temperature, including horseradish peroxidase
(HRP)-conjugated horse anti-mouse IgG (cat. no. PI200) and
HRP-conjugated goat anti-rabbit IgG (cat. no. PI1000; dilution,
1:1,000; both Vector Laboratories, Inc., Burlingame, CA, USA).
Signals were detected using an enhanced chemiluminescence reaction
(ProteinSimple; Bio-Techne, Minneapolis, MN, USA). The intensity of
bands was quantified using Gel-Pro Image Analysis software version
32 (Media Cybernetics, Inc., Rockville, MD, USA).
Immunohistochemical analysis
Formalin-fixed, paraffin embedded astrocytoma
sections (thickness, 4 µm) were used for immunohistochemistry (IHC)
staining using the labeled streptavidin-biotin method (Dako;
Agilent Technologies, Inc., Santa Clara, CA, USA). Endogenous
peroxidase activity was blocked in deparaffinized slides by
incubating sections in 3% H2O2 methanol
solution at room temperature for 10 min. Then antigen retrieval was
performed with 10 mM citrate buffer (pH 6.0) at 95–100°C for 10
min. The slides were blocked with 10% goat serum (Abcam) in
phosphate-buffered saline for 20 min at room temperature. Primary
antibodies included the previously described anti-P2X7R
(dilution, 1:200), p53 (cat. no. P9249; dilution, 1:500;
Sigma-Aldrich; Merck KGaA), epidermal growth factor receptor (EGFR;
cat. no. PB0039; Shanghai Changdao Biotech Co., Ltd., Shanghai,
China; dilution, 1:200) and MIB-1 (marker of proliferation, Ki-67;
cat. no. sc-101861; dilution, 1:1,000; Santa Cruz Biotechnology,
Inc., Dallas, TX, USA). Sections were developed with the
rabbit/mouse peroxidase/3,3′-diaminobenzidine EnVision™ Detection
kit (cat. no. GK500705; Dako; Agilent Technologies, Inc.)
containing the secondary antibody and 3,3′-diaminobenzidine
according to the manufacture's protocol, and the nuclei were
counterstained with eosin.
The P2X7R and EGFR immunoreactivity scores (IRS)
were measured. The fraction (intensity percent; IP) of stained
cells was estimated and scored as follows: 0, 0–1%; 1, 2–10%; 2,
11–30%; 3, 31–60%; and 4, 61–100%. The staining intensity (SI) was
scored as follows: 0, no staining; 1, weak but definite; 2,
moderate; and 3, intense. The IRS was then calculated using the
formula: IRS=IP × SI.
The percentages of MIB-1 are presented as the
proliferation index (PI). The PI of tumor tissues was expressed as
follows: PI% = A × 100/(A + C), where A is the number of
MIB-positive cells, and C is the number of counterstained unlabeled
cells.
Statistical analysis
Statistical analysis was performed using SPSS 17.0
statistical package (SPSS, Inc., Chicago, IL, USA). Statistical
methods included unpaired t-test or one-way analysis of variance,
followed by Scheffe's test. Linear correlation analysis was used to
examine the association between P2X7R and other
parameters of glioma. P<0.05 was considered to indicate a
statistically significant difference. Data are expressed as the
mean ± standard error of mean.
Results
P2X7R expression is
significantly lower in astrocytoma compared with the surrounding
normal brain tissue
The role of P2X7R in astrocytoma was
determined by histologically and immunohistochemically analyzing
its expression in normal brain tissue, and astrocytoma.
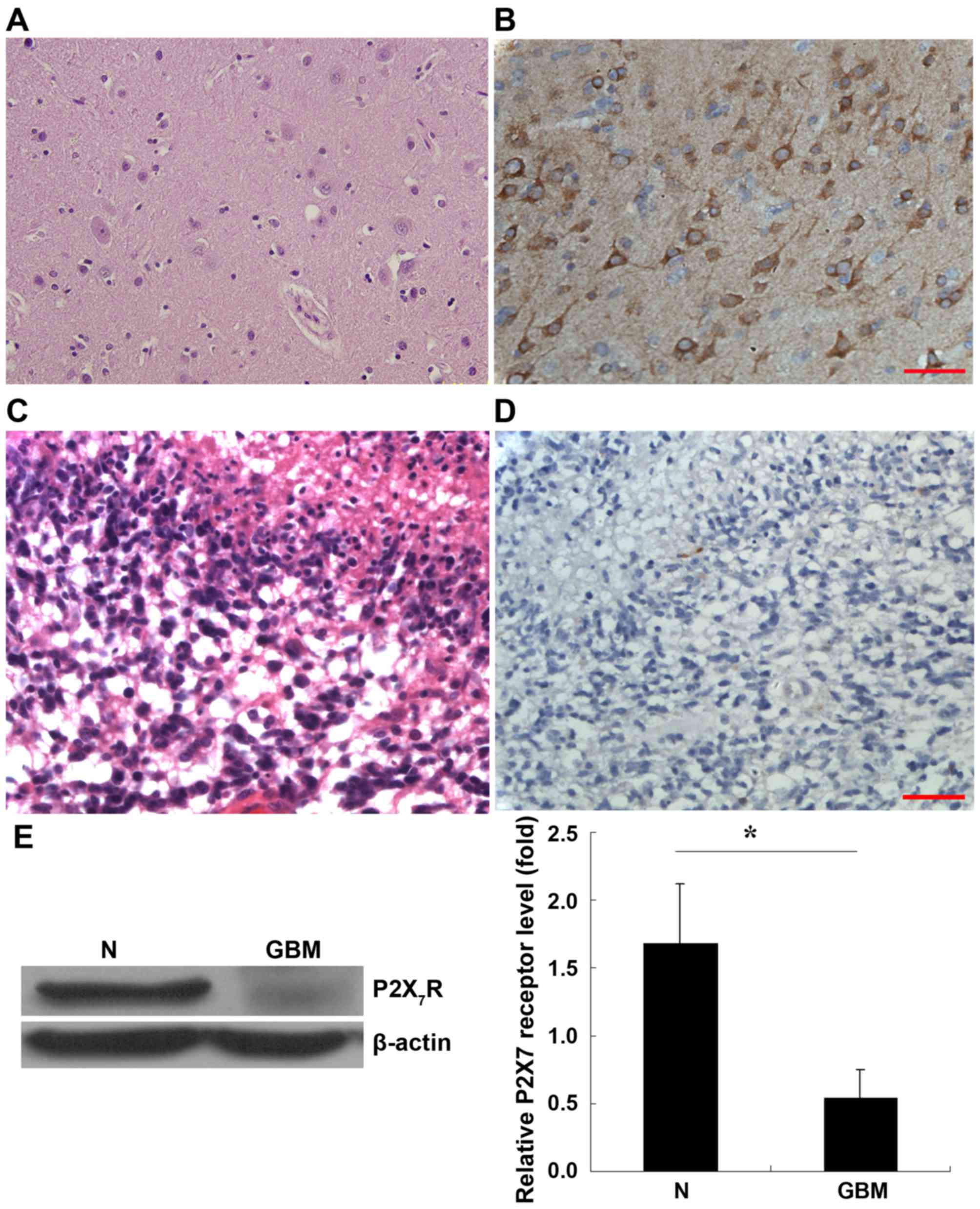
Histological staining of normal brain tissue is presented in
Fig. 1A. IHC staining with
anti-P2X7R antibody revealed that the cells were
positive for P2X7R in the normal brain tissue (Fig. 1B) whereby P2X7R was
expressed in the cytoplasm and membrane of these cells.
Histological staining of GBM is presented in Fig. 1C. However, IHC revealed that
P2X7R expression was almost negative in the tumor
(Fig. 1D).
To confirm the expression of P2X7R in
these tissues, western blot analysis was performed. As presented in
Fig. 1E, normal brain tissue and
astrocytoma expressed b-actin at similar levels, whereas the
expression of P2X7R was diminished in GBM samples when
compared with peripheral brain tissue. Statistical analysis using 7
paired GBM tissues and their corresponding samples of normal
peripheral brain tissue revealed that P2X7R expression
was decreased significantly in the GBM (Fig 1E; P<0.05).
Hypermethylation of P2X7R
promoter in GBM
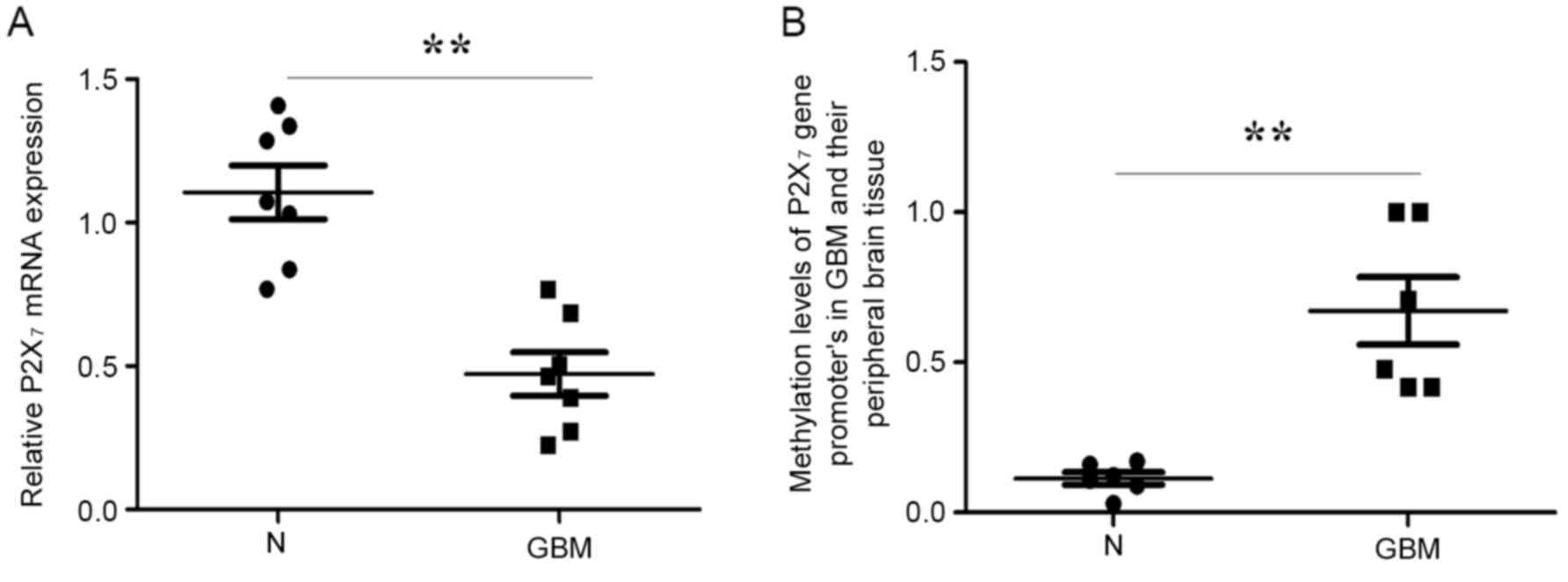
P2X7R mRNA was determined using RT-qPCR
in the 7 fresh GBM and peripheral brain tissue samples. Consistent
with the P2X7R protein expression, the P2X7R
mRNA level in GBM was significantly lower compared with that of the
peripheral brain tissue (Fig.
2A).
To elucidate the molecular mechanism of the
decrease, a DNA methylation assay was performed. As presented in
Fig. 2B, a significantly higher level
of methylation (mean=0.67±0.1) was observed in GBMs compared with
the surrounding normal brain samples (mean=0.12±0.09) (P=0.046) in
the 547-bp region (+26/+573 nt) of the P2X7R promoter,
containing 18 CpG sites. Thus, decreased
P2X7R expression in astrocytoma may be
associated with hypermethylation of its promoter.
Expression of P2X7R is
negatively associated with malignancy grade in astrocytoma
The expression of P2X7R in different
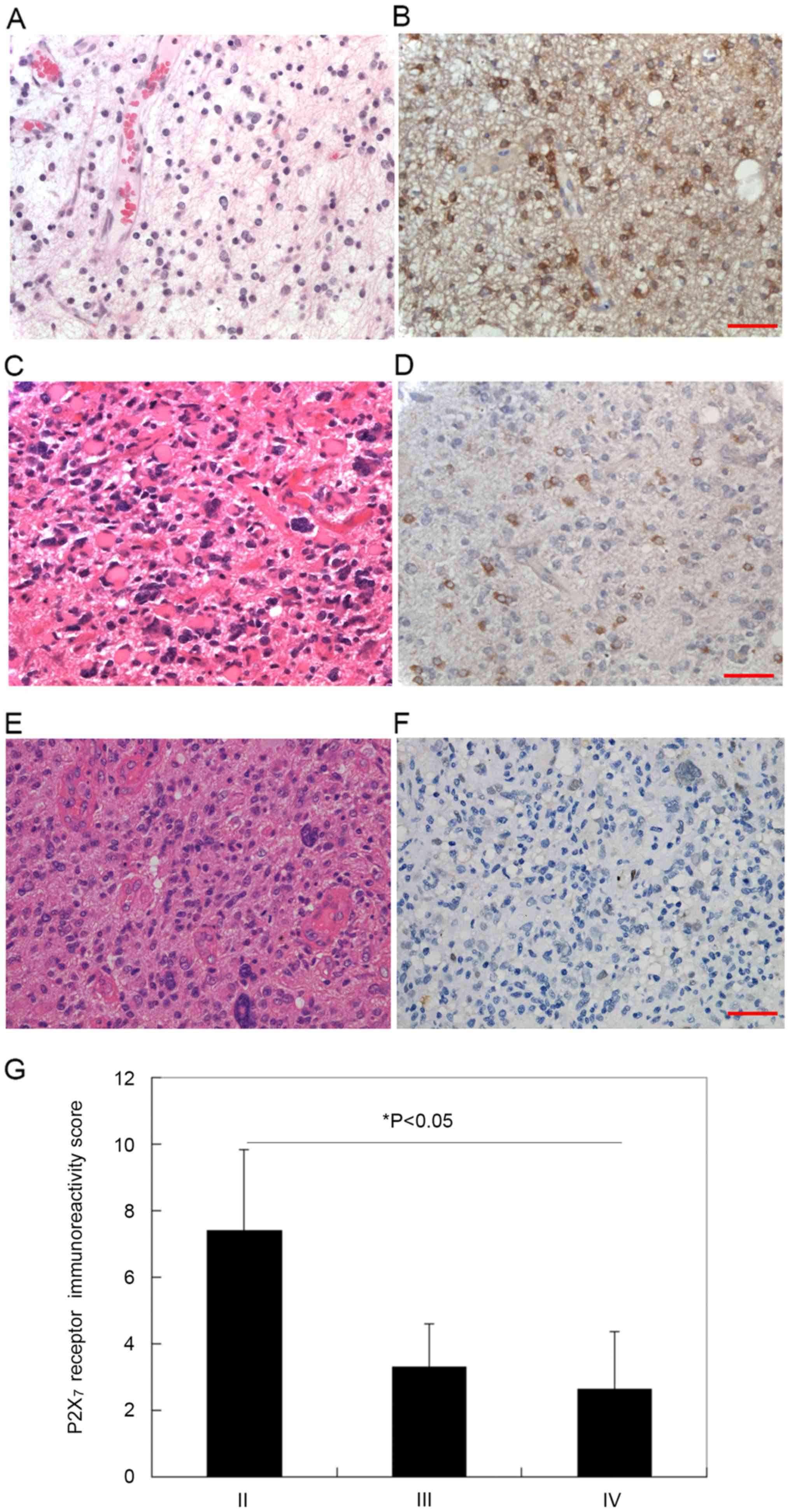
grades of astrocytoma was then examined. Histological grades of
astrocytoma were determined according to nuclear atypia, mitosis,
endothelial proliferation and necrosis (Fig. 3). The malignancy level increased from
WHO grade II to IV. As presented in Fig.
3A, C and E, histological staining revealed a marked increase
in cell density, nuclear heteromorphism, and caryokinesis between
grade II and IV. By contrast, IHC staining revealed that the
expression of P2X7R decreased correspondingly (Fig. 3B, D and F). Thus, the expression of
P2X7R may be negatively associated with the malignancy
of astrocytoma (Fig. 3G).
In addition, it was confirmed that P2X7R
was negatively correlated with the proliferation index of MIB-1
(r=−0.411; P<0.01). No correlation was identified between
P2X7R, and cumulative p53 or EGFR expression in the
astrocytoma (data not shown).
Discussion
P2X7R-mediated apoptosis depends on
P2X7R expression (14).
The expression of P2X7R did not differ significantly
between normal colon epithelial cells and colon cancer cells
(15). However, thyroid cancer cells
exhibit higher P2X7R expression compared with the normal
cells (16). The P2X7R
expression has be revealed to be decreased in early phases of
neoplasia of the ectoderm (skin and breast) (17,18), the
distal paramesonephric duct (endocervix and endometrium) (19), and the urogenital sinus (bladder and
ectocervix) (20,21). However, the expression of
P2X7R in brain tumor remains elusive.
Diffuse infiltration of glioma is a major
therapeutic challenge for definitive surgical resection. It limits
the efficacy of other local therapies, leading to local recurrence
and short survival. In GBM, areas, including the tumor center
distal to the capillaries often become hypoxic and undergo
necrosis. Such regions are considered to contain higher
concentrations of extracellular ATP (22). Activation of P2X7R in
glioma C6 cells leads to increased mobilization of intracellular
calcium, formation of large pores and enhanced expression of
several pro-inflammatory factors (23). The tumor suppressor role of
P2X7R has been attributed to its pro-inflammatory and
pro-apoptotic function (24).
Consistent with this hypothesis, activation of P2X7R in
astrocytes leads to a state of reversible growth arrest (25). These data were obtained from mouse GBM
or normal astrocyte cells. Using fresh human GBM and surrounding
brain tissue samples, the present study revealed that the
expression of P2X7R mRNA, and protein was significantly
decreased. However, another immunohistochemical study reported the
elevated expression of P2X7R in glioma (24). In the present study, using 100
astrocytoma samples at different malignant grades, P2X7R
expression was identified to be negatively correlated with the
malignancy of astrocytoma.
Generally, methylation at CpG islands in a gene
promoter favors transcriptional repression (25). DNA cytosine methylation is one of the
most consistent epigenetic alterations in various types of human
cancer (26). DNA hypermethylation is
an essential epigenetic mechanism for the silencing of numerous
genes, including those involving cell cycle regulation, receptors,
DNA repair and apoptosis. The present study revealed that
P2X7R genes were hypermethylated at the CpG sites in the
+26/+573 nt region of human astrocytoma samples. These results
indicate that the decrease in P2X7R expression may be
associated with hypermethylation of its gene promoter.
Previous studies have demonstrated that
P2X7R exhibits significant growth-promoting effects
in vivo (27,28), inducing extensive neovascularization
and elevated levels of VEGF, and thereby promoting cell invasion
and migration (29). However, the
exact molecular basis of P2X7R-mediated growth-promoting
activity is unclear. By contrast, P2X7R suppression
induced the growth of glioma by directly promoting cell
proliferation and angiogenesis (30).
Functional P2X7R activation by ATP resulted in rapid
cytotoxicity affecting cell growth and survival, leading to
inhibition of tumor growth in vitro, and in vivo
(31,32). P2X7R expression was also
identified to be negatively correlated with a high proliferation
index of glioma in the present study. Therefore, the decrease in
P2X7R expression may have an important role in the
development and progression of astrocytoma.
In the present study, compared with the tumor
tissue, the surrounding normal brain tissue demonstrated high
P2X7R immunoreactivity. Thus, we hypothesize that the
normal brain cells may become apoptotic following P2X7R
activation, thus accommodating tumor cells. As a consequence,
P2X7R expression in normal brain cells enhances the
infiltration and proliferation of the glioma. Contrary to this, a
higher P2X7R expression in astroglioma may increase the
response to radiotherapy (33).
Therefore, increasing the malignancy and decreasing
P2X7R expression of astroglioma may lead to higher
therapeutic resistance.
In conclusion, the results indicated that the levels
of P2X7R were lower in astrocytoma tissues compared with
the normal surrounding brain tissue. Inhibition of the
P2X7R-mediated apoptosis in the peripheral normal brain
tissue surrounding the tumor may limit glioma infiltration and
growth. Therefore, it is important to understand P2X7R
expression and its function in glioma, and normal brain cells to
develop appropriate chemotherapeutic interventions in
vivo.
Acknowledgements
The present study was supported by the Chinese
National Science Foundation (grant no. 81272796) and the Natural
Sciences Foundation of Shanghai Project (grant no.
12ZR1403200).
References
|
1
|
Cavenee W K, Leung S, Hawkins C, et al:
WHO Classification of Tumours of the Central Nervous System,
Revised. Fourth Edition. WHO Press; Geneva, Switzerland: pp. 28–45.
2016
|
|
2
|
Johnson DR and O'Neill BP: Glioblastoma
survival in the United States before and during the temozolomide
era. J Neurooncol. 107:359–364. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cotrina ML, Lin JH, López-García JC, Naus
CC and Nedergaard M: ATP-mediated glia signaling. J Neurosci.
20:2835–2844. 2000.PubMed/NCBI
|
|
4
|
Zimmermann H: Extracellular metabolism of
ATP and other nucleotides. Naunyn Schmiedebergs Arch Pharmacol.
362:299–309. 2003. View Article : Google Scholar
|
|
5
|
Ciccarelli R, Ballerini P, Sabatino G,
Rathbone MP, D'Onofrio M, Caciagli F and Di Lorio P: Involvement of
astrocytes in purine-mediated reparative processes in the brain.
Int J Dev Neurosci. 19:395–414. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ralevic V and Burnstock G: Receptors for
purines and pyrimidines. Pharmacol Rev. 50:413–492. 1998.PubMed/NCBI
|
|
7
|
Di Virgilio F: Dr. Jekyll/Mr. Hyde: The
dual role of extracellular ATP. J Auton Nerv Syst. 81:59–63. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kim M, Jiang LH, Wilson HL, North RA and
Surprenant A: Proteomic and functional evidence for a P2X7 receptor
signalling complex. EMBO J. 20:6347–6358. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang L, Feng YH and Gorodeski GI:
Epidermal growth factor facilitates epinephrine inhibition of
P2X7-receptor-mediated pore formation and apoptosis: A novel
signaling network. Endocrinology. 146:164–174. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Schneider EM, Vorlaender K, Ma X, Du W and
Weiss M: Role of ATP in trauma-associated cytokine release and
apoptosis by P2X7 ion channel stimulation. Ann NY Acad Sci.
1090:245–252. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bulanova E, Budagian V, Orinska Z, Hein M,
Petersen F, Thon L, Adam D and Bulfone-Paus S: Extracellular ATP
induces cytokine expression and apoptosis through P2X7 receptor in
murine mast cells. J Immunol. 174:3880–3890. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Said T, Dutot M, Christon R, Beaudeux JL,
Martin C, Warnet JM and Rat P: Benefits and side effects of
different vegetable oil vectors on apoptosis, oxidative stress, and
P2X7 cell death receptor activation. Invest Ophthalmol Vis Sci.
48:5000–5006. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Feng YH, Wang L, Wang Q, Li X, Zeng R and
Gorodeski GI: ATP stimulates GRK-3 phosphorylation and
beta-arrestin-2-dependent internalization of P2X7 receptor. Am J
Physiol Cell Physiol. 288:C1342–C1356. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li X, Qi X, Zhou L, Fu W, Abdul-Karim FW,
Maclennan G and Gorodeski GI: P2X(7) receptor expression is
decreased in epithelial cancer cells of ectodermal, uro-genital
sinus and distal paramesonephric duct origin. Purinergic Signal.
5:351–368. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Solini A, Cuccato S, Ferrari D, Santini E,
Gulinelli S, Callegari MG, Dardano A, Faviana P, Madec S, Di
Virgilio F and Monzani F: Increased P2X7 receptor expression and
function in thyroid papillary cancer: A new potential marker of the
disease? Endocrinology. 149:389–396. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fu W, McCormick T, Qi X, Luo L, Zhou L, Li
X, Wang BC, Gibbons HE, Abdul-Karim FW and Gorodeski GI: Activation
of P2X(7)-mediated apoptosis Inhibits DMBA/TPA-induced formation of
skin papillomas and cancer in mice. BMC Cancer. 9:1142009.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Greig AV, Linge C, Healy V, Lim P, Clayton
E, Rustin MH, McGrouther DA and Burnstock G: Expression of
purinergic receptors in non-melanoma skin cancers and their
functional roles in A431 cells. J Invest Dermatol. 121:315–327.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li X, Qi X, Zhou L, Catera D, Rote NS,
Potashkin J, Abdul-Karim FW and Gorodeski GI: Decreased expression
of P2X7 in endometrial epithelial pre-cancerous and cancer cells.
Gynecol Oncol. 106:233–243. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lee HY, Bardini M and Burnstock G:
Distribution of P2X receptors in the urinary bladder and the ureter
of the rat. J Urol. 163:2002–2007. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li X, Zhou L, Feng YH, Abdul-Karim FW and
Gorodeski GI: The P2X7 receptor: A novel biomarker of uterine
epithelial cancers. Cancer Epidemiol Biomarkers Prev. 15:1906–1913.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wei W, Ryu JK, Choi HB and McLarnon JG:
Expression and function of the P2X(7) receptor in rat C6 glioma
cells. Cancer Lett. 260:79–87. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Fang KM, Wang YL, Huang MC, Sun SH, Cheng
H and Tzeng SF: Expression of macrophage inflammatory protein-1α
and monocyte chemoattractant protein-1 in glioma-infiltrating
microglia: Involvement of ATP and P2X7 receptor. J Neurosci Res.
89:199–211. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ryu JK, Jantaratnotai N, Serrano-Perez MC,
McGeer PL and McLarnon JG: Block of purinergic P2X7R inhibits tumor
growth in a C6 glioma brain tumor animal model. J Neuropathol Exp
Neurol. 70:13–22. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zilberman D and Henikoff S: Genome-wide
analysis of DNA methylation patterns. Development. 134:3959–3965.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Esteller M: CpG island hypermethylation
and tumor suppressor genes: A booming present, a brighter future.
Oncogene. 21:5427–5440. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Gómez-Villafuertes R, García-Huerta P,
Díaz-Hernández JI and Miras-Portugal MT: PI3K/Akt signaling pathway
triggers P2X7 receptor expression as a pro-survival factor of
neuroblastoma cells under limiting growth conditions. Sci Rep.
5:184172015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Braganhol E, Kukulski F, Lévesque SA,
Fausther M, Lavoie EG, Zanotto-Filho A, Bergamin LS, Pelletier J,
Bahrami F, Ben Yebdri F, et al: Nucleotide receptors control
IL-8/CXCL8 and MCP-1/CCL2 secretions as well as proliferation in
human glioma cells. Biochim Biophys Acta. 1852:120–130. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Qiu Y, Li WH, Zhang HQ, Liu Y, Tian XX and
Fang WG: P2X7 mediates ATP-driven invasiveness in prostate cancer
cells. PLoS One. 9:e1143712014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Fang J, Chen X, Zhang L, Chen J, Liang Y,
Li X, Xiang J, Wang L, Guo G, Zhang B and Zhang W: P2X7R
suppression promotes glioma growth through epidermal growth factor
receptor signal pathway. Int J Biochem Cell Biol. 45:1109–1120.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
White N, Butler PE and Burnstock G: Human
melanomas express functional P2X(7) receptors. Cell Tissue Res.
321:411–418. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Bian S, Sun X, Bai A, Zhang C, Li L,
Enjyoji K, Junger WG, Robson SC and Wu Y: P2X7 integrates PI3K/AKT
and AMPK-PRAS40-mTOR signaling pathways to mediate tumor cell
death. PLoS One. 8:e601842013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Gehring MP, Kipper F, Nicoletti NF,
Sperotto ND, Zanin R, Tamajusuku AS, Flores DG, Meurer L, Roesler
R, Filho AB, et al: P2X7 receptor as predictor gene for glioma
radiosensitivity and median survival. Int J Biochem Cell Biol.
68:92–100. 2015. View Article : Google Scholar : PubMed/NCBI
|