Introduction
Urinary bladder cancer is a common malignancy of the
urinary tract and responsible for 150,000 deaths annually. Urinary
bladder cancer is the seventh most prevalent type of cancer
worldwide (1). In Korea, the number
of cases of urinary bladder cancer increased from 2,180 in 1999 to
3,549 in 2011 (2). The incidence of
urinary bladder cancer is four times higher in men compared with
women (3).
Cisplatin-based combination chemotherapy is widely
used against advanced and metastatic bladder cancer. Such drugs,
however, yield poor oncological outcomes and show high toxicity
(4), and there is growing interest in
other types of therapeutic agents for the treatment of advanced
bladder cancer (4). The development
of more effective and less toxic therapeutic regimens is vital for
the improvement of survival among patients with bladder cancer.
Recent studies showed that natural bioactive
compounds, including isothiocyanate and amygdalin, have anticancer
effects on bladder cancer cells (5,6). Garlic, a
species of the Allium genus, has been utilized for medicinal
purposes in recorded history (7–9).
Organosulfur compounds from Allium vegetables have been
reported to have possible preventive and therapeutic properties
against some types of cancer (7).
Sulfur-containing compounds in garlic can be broadly categorized as
oil-soluble (diallylsulphide, diallyldisulphide and
diallyltrisulfide) or water-soluble [S-allyl-L-cysteine (SAC)]
(8). SAC is the most abundant
organosulfur compound in aged-garlic extract and was reported to
have antioxidant, and neuroprotective properties as well as
anticancer activity (9–11). Several studies showed that SAC has
anticancer effects by suppressing the cellular proliferation and
metastasis and induces apoptosis in a number of cancer models,
including ovarian, prostate and hepatocellular carcinoma (7–9).
Nevertheless, whether SAC has anticancer effects in bladder cancer
remains an open question.
In the present study, in vitro effects of SAC
were evaluated by Cell Counting Kit-8 (CCK-8) assay, colony
formation assay, cell cycle analysis and by measuring the
expression of proteins associated with apoptosis and cell cycle in
human bladder cancer cell lines, T24 and T24R2.
Materials and methods
Cell lines and reagents
Human bladder cancer cell lines HTB5, HTB9, J82,
JON, UMUC14 and T24 were obtained from the American Type Culture
Collection (Manassas, VA, USA). The cisplatin-resistant cell line
T24R2 was generated by serial desensitization, as previously
described (12). The cell lines
(HTB5, HTB9, T24, and T24R2) were cultured in RPMI-1640 and the
cell lines (J82, JON, and UMUC14) were cultured in Dulbecco's
modified Eagle's medium supplemented with 10% fetal bovine serum
and 1% penicillin/streptomycin (all from Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) at 37°C in a humidified
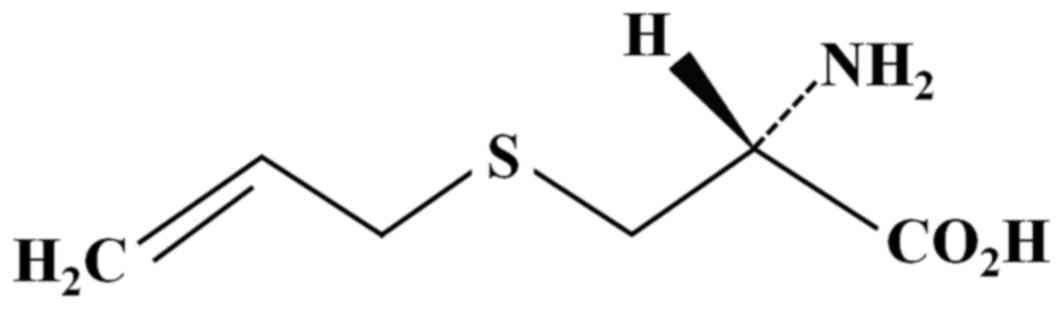
atmosphere incubator containing 5% CO2. SAC (PharmaKing
Co., Ltd., Gyeonggi-do, Korea; Fig.
1) was diluted in dimethyl sulfoxide (DMSO; Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany) to obtain a 100 µM stock solution
and was then diluted to obtain the working concentration. The final
concentration of DMSO in the culture media did not exceed 0.1%
(v/v). Media containing 0.1% DMSO was used as a control.
Cell proliferation assay
Cell proliferation was analyzed using the CCK-8
assay (Dojindo Molecular Technologies, Inc., Rockville, MD, USA)
according to the manufacturers instructions. Bladder cancer cells
were seeded in 96-well plates at the density of
2×103/well. After 24 h, the cells were incubated at 37°C
with SAC for 24, 48, or 72 h. At the end of the drug exposure, 10
µl CCK-8 solution was added to each well, which contains 200 µl of
the culture medium. After 4 h of incubation, the absorbance at 450
nm was measured on a plate reader (Molecular Devices, LLC,
Sunnyvale, CA, USA). Cell viability was calculated as the
percentage of viable cells in the total population. The experiment
was performed in triplicate.
Clonogenic assay
T24 and T24R2 cells were plated at 5×102
per 6-well plate and incubated with various concentrations of SAC
(T24: 0, 10, 25 and 50 mM; T24R2: 0, 1, 5, 10 and 25 mM) for 48 h.
The cells were incubated at 37°C for another 14 days in the
SAC-free medium. The cells were fixed with methanol and stained
with a 0.1% crystal violet solution. The plates were photographed,
and the colonies >0.2 mm in diameter were counted.
Cell cycle analysis
T24 and T24R2 cells were plated at 3×105
per 60 mm dish, grown for 24 h and then incubated at 37°C with
various concentrations of SAC (T24: 0, 10, 25 and 50 mM; T24R2: 0,
1, 5, 10 and 25 mM) for 48 h. The cells were trypsinized and fixed
in 70% ethanol and subsequently stained with a propidium iodide
solution (1 mg/ml) for 30 min at 37°C. The cell cycle distribution
was determined using CellQuest Pro software (version 5.1) on a
FACSCalibur instrument (both from BD Biosciences, San Jose, CA,
USA).
Western blotting
The cells were lysed with radio immunoprecipitation
assay (RIPA) buffer, consisting of 50 mM Tris-HCl (pH 8.0), 150 mM
sodium chloride, 1.0% NP-40, 0.5% sodium deoxycholate, 0.1% SDS and
1 mM phenylmethylsulfonyl fluoride. Protein concentrations were
determined by using the BCA protein assay kit (Pierce; Thermo
Fisher Scientific, Inc.). Equal amounts of total protein (50 µg)
were analyzed by SDS-PAGE (8 or 10% gels) and then transferred onto
polyvinylidene fluoride membranes (EMD Millipore, Billerica, MA,
USA). The blots were blocked for 1 h with 5% (w/v) non-fat dry milk
at room temperature and then incubated with primary antibodies:
Cleaved caspases-3, −8, −9, PARP, poly (ADP-ribose) polymerase,
cytochrome c, Bcl-2, Bax, cyclins B1, D1, and E1,
phosphorylated-AKT and AKT (Cell Signaling Technology, Inc.,
Danvers, MA, USA) overnight at 4°C. After incubation with secondary
antibodies (anti-mouse; catalog no. sc-2055; dilution, 1:10,000;
and anti-rabbit; catalog no. sc-2004; dilution, 1:5,000; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA), protein expression was
detected with enhanced chemiluminescence (ECL) Western blot
substrate kit (Pierce; Thermo Fisher Scientific, Inc.) and
quantified with ImageJ software (National Institute of Health,
Bethesda, MA, USA).
Statistical analysis
The data are presented as mean ± standard deviation
of three independent experiments in triplicate. All calculations
were performed using SPSS statistical software package (version
20.0; IBM Corp., Armonk, NY, USA). Statistical significance
(P<0.05) was determined by Tukey's multiple comparison test.
Results
Treatment with SAC suppresses
proliferation of human bladder cancer cells
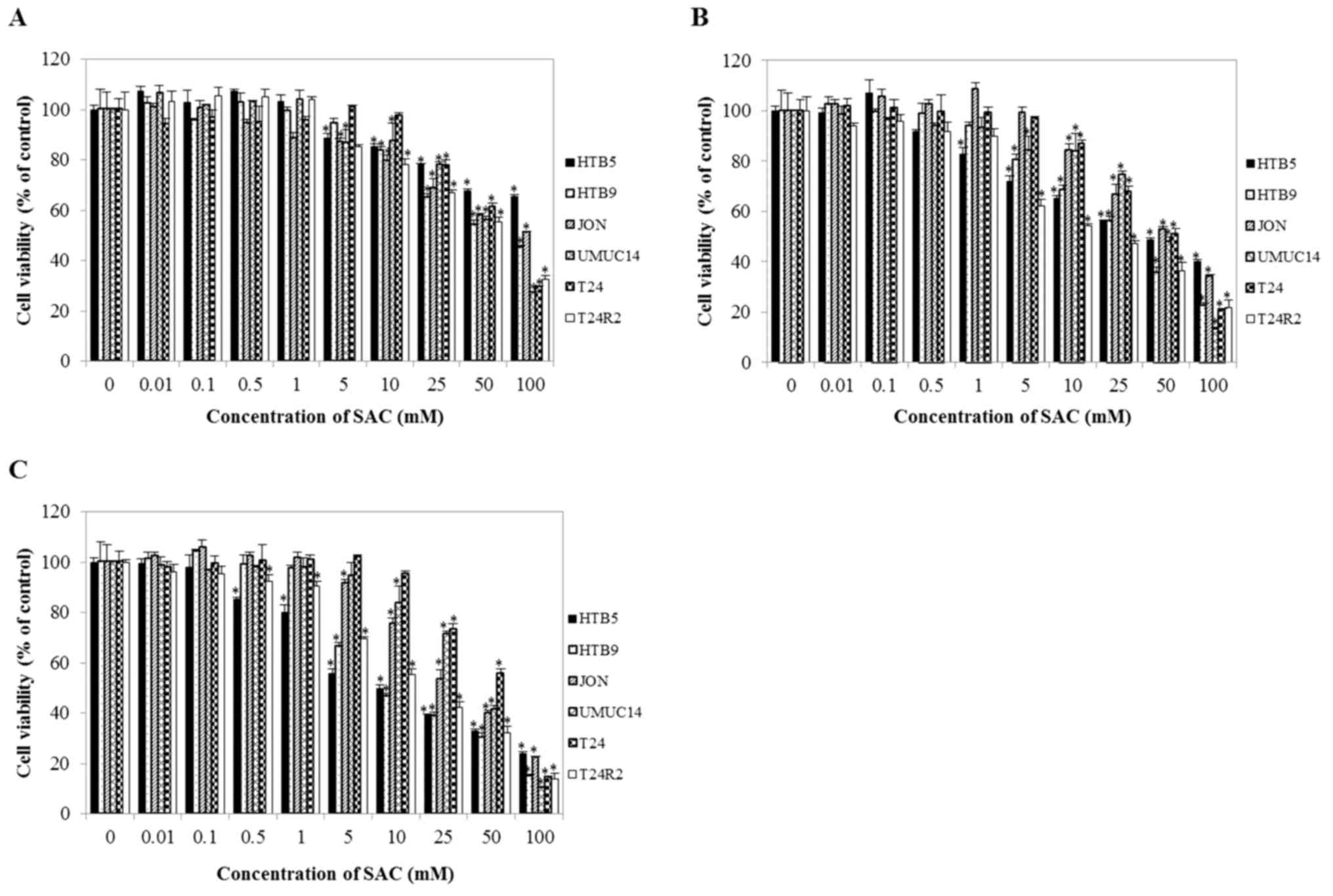
To assess the effect of SAC on the growth of bladder
cancer cells, CCK-8 assays were performed. As shown in Fig. 2, SAC markedly suppressed proliferation
of bladder cancer cells in a time- and dose-dependent manner
compared with untreated cells (control). Compared with the other
cell lines (HTB9, JON, UMUC14 and T24), HTB5 and T24R2 were more
sensitive to SAC treatment. The 50% inhibitory concentration
(IC50) of SAC toward T24 and T24R2 bladder cancer cells
after 48 h of exposure was 52.98 and 19.87 mM, respectively. To
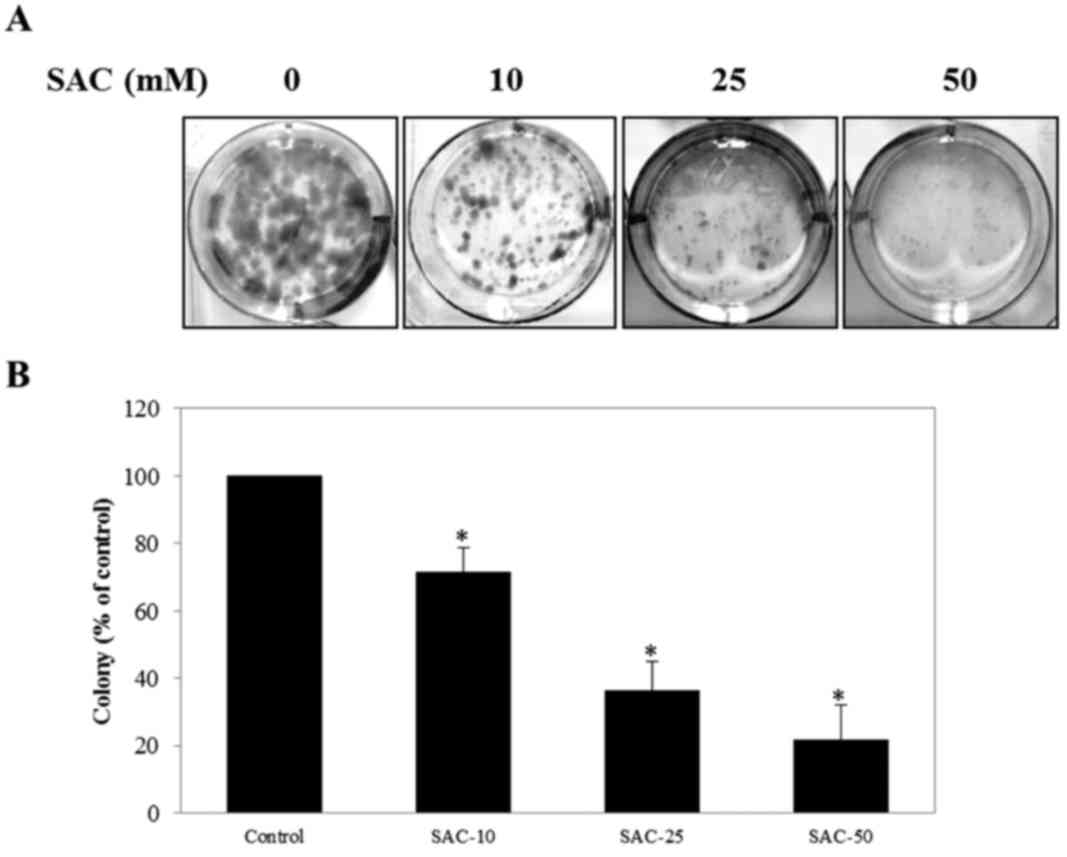
evaluate the anti-proliferative effect of SAC on T24 and T24R2
cells, clonogenic assay was performed. The colony-forming ability
of T24 and T24R2 cells was significantly inhibited with increasing
SAC concentrations in comparison with the control (Fig. 3). These data indicated that the SAC
treatment may suppress the proliferation of bladder cancer cells in
a dose-dependent manner.
Treatment with SAC induces alterations
in cell cycle in bladder cancer cells
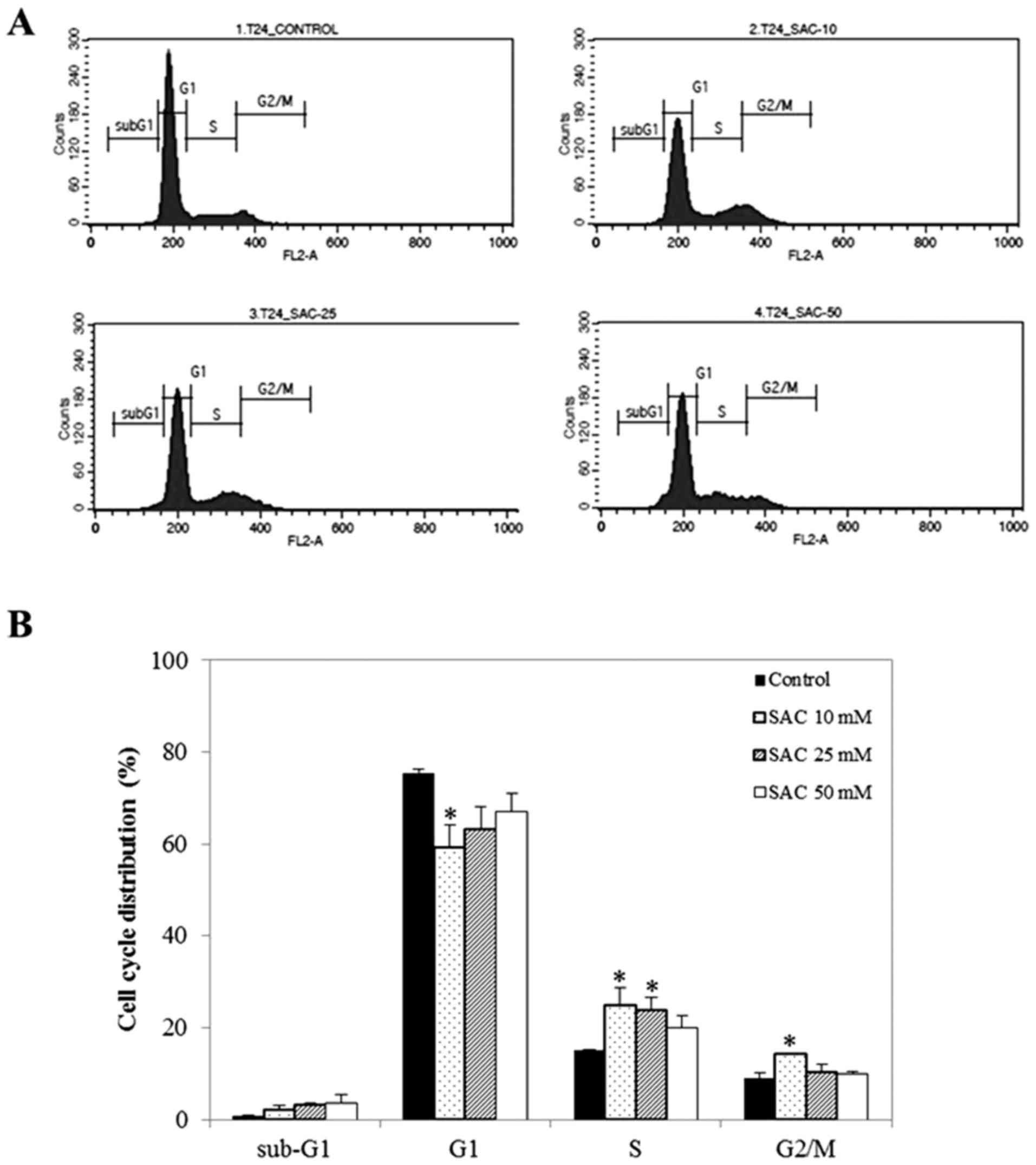
To elucidate the mechanism of SAC-induced
proliferation inhibition, cell cycle distribution was detected by
fluorescence-activated cell sorting (FACS) analysis. It was
demonstrated that SAC markedly altered cell cycle progression of
bladder cancer cell lines. Particularly, SAC markedly increased the
sub-G1 cell population (which corresponds to apoptotic cells) in a
dose-dependent manner (Fig. 4). The
highest concentration of SAC (T24: 50 mM; T24R2: 25 mM) caused a
4.43 and 3.12-fold increase in the number of cells in the sub-G1
population in T24 and T24R2 cells, respectively, compared with the
control. Of note, a statistically significant difference was
observed in T24R2 cells treated with 25 mM SAC compared with the
control. Additionally, the S phase population was noticeably
increased from 16.8 (untreated control) to 38.2% in T24R2 cells
treated with 25 mM SAC. These results showed that SAC increased the
sub-G1 population, which corresponds to apoptotic cells, in T24 and
T24R2 bladder cancer cells.
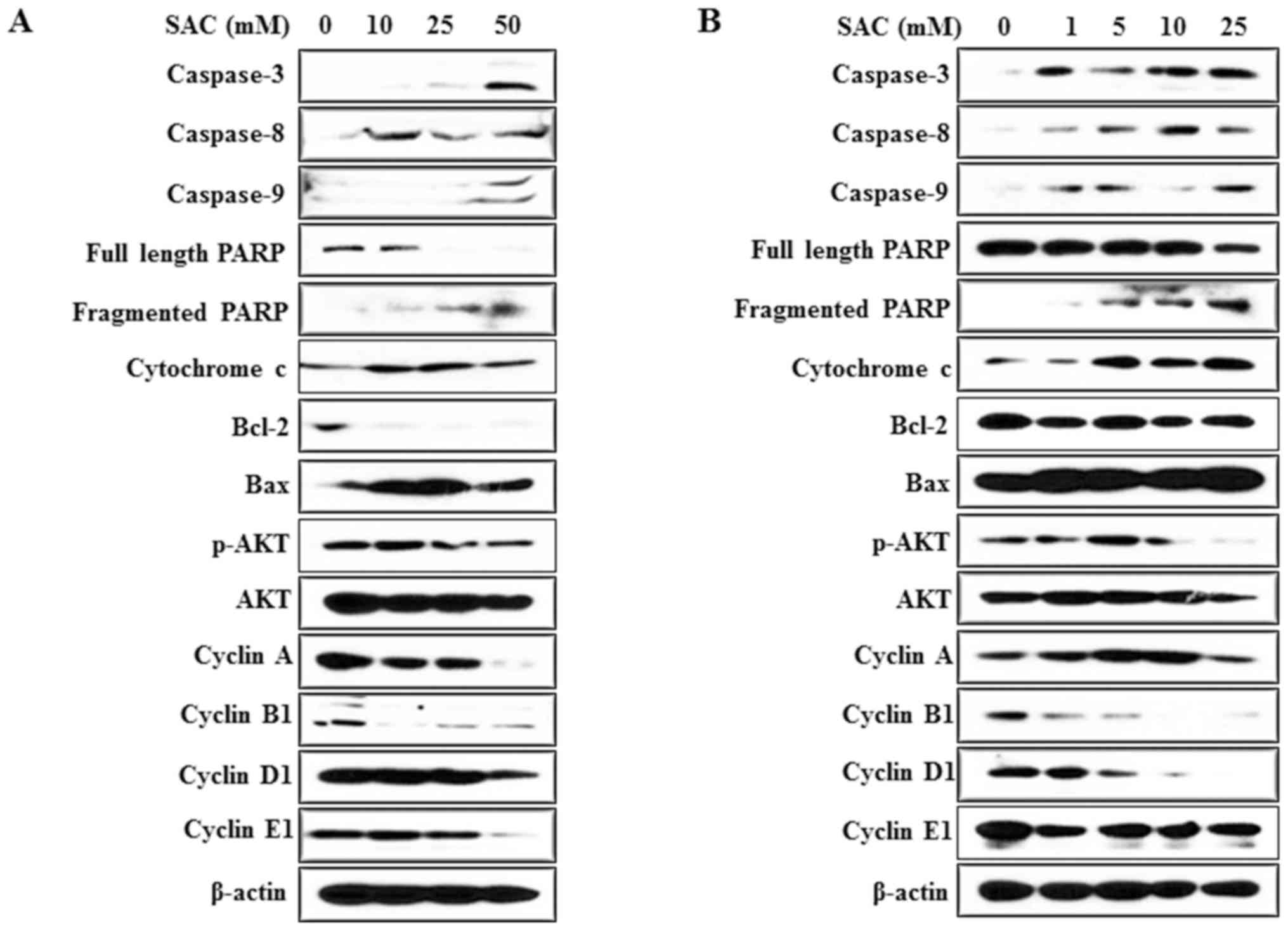
Treatment with SAC alters expression
of apoptosis and cell cycle-associated proteins in bladder cancer
cells
To validate the effects of SAC on apoptosis and the
cell cycle, western blot analysis was performed. The expression of
caspases-3, −8, and −9; fragmented PARP and cytochrome c was
markedly increased with increasing SAC concentrations in T24 and
T24R2 cells (Fig. 5). Additionally,
the expression of the anti-apoptotic protein Bcl-2 was decreased
following SAC treatment in T24 and T24R2 cells, whereas the
expression of the pro-apoptotic protein Bax was increased.
In addition, as an index of S phase arrest (13), the expression of cyclins was measured.
Treatment with SAC markedly downregulated cyclin B1, D1 and E1. AKT
phosphorylation was also markedly suppressed by SAC in the bladder
cancer cells (Fig. 5). These results
suggested that SAC inhibited the growth of bladder cancer cells via
an apoptosis-associated mechanism and disrupted cell survival by
inhibiting the AKT signaling pathway.
Discussion
Nowadays, cancer patients are interested in
complementary and alternative medicine (CAM) because they do not
like adverse effects of conventional medical treatment, and because
of the need for personal control over treatment as well as, the
perceived safety of a natural product (14,15). In
Europe, >50% of cancer patients use CAM instead of or in
combination with conventional medical treatment (6). CAM includes diet and nutritional
interventions, music therapy and aromatherapy. Among these
therapies, diet and nutritional therapies are most commonly used by
cancer patients (14).
Recently, many studies showed that natural
compounds, including (−)-epigallocatechin-3-gallate and
resveratrol, induce cell death and cell cycle arrest in bladder
cancer cells (16–18). In the present study, incubation of
bladder cancer cell lines with SAC markedly reduced cell
proliferation according to CCK-8 and clonogenic assays in a time-
and dose-dependent manner. SAC inhibited the proliferation of
bladder cancer cells at least in part by inducing apoptosis, as
evidenced by increase in the sub-G1 population. The data in the
present study are consistent with the findings of another study,
which showed that SAC exhibits its anticancer activity on prostate
cancer cells and ovarian cancer cells by inducing apoptosis
(8,9).
Altogether, these studies indicate that SAC may account for
apoptosis in multiple cancer cells, including bladder cancer cells
and contribute to suppressed cancer cell proliferation.
Cell cycle checkpoints are a key factor in cancer
pathogenesis, and their dysregulation may affect the effectiveness
of cancer chemotherapy. Numerous studies have shown that cancer
progression involves the loss of checkpoint controls that regulate
the passage through the cell cycle (19). Flow cytometry was used in the present
study to identify the mechanism underlying the SAC-mediated
inhibition of cell growth. Cell cycle arrest was detected in the S
phase following treatment with SAC in bladder cancer cells,
consistent with the findings of Nq et al (7) in hepatic cancer cells. In support of
this hypothesis, it was also identified that SAC inhibits
expression of cyclins B1, D1 and E1 in the bladder cancer cells.
Taken together, the results in the present study indicated that SAC
may inhibit cell growth in part by affecting apoptosis and the
regulation of the cell cycle.
Overexpression of anti-apoptotic proteins including
Bcl-2 and Bcl-xl is often associated with tumor recurrence, poor
prognosis and resistance to cancer treatment (20). Generally, cancer cells can inactivate
apoptotic signaling pathways by overexpressing such anti-apoptotic
proteins and via the down-regulation of pro-apoptotic proteins
(including Bax and Bad) and the caspase family (20,21). On
the other hand, inhibition of the intrinsic apoptotic pathways is
linked with activities of the AKT and NF-κB signaling pathways. The
activated AKT pathway inhibits the intrinsic apoptotic pathway by
upregulating anti-apoptotic proteins while downregulating
pro-apoptotic proteins (21). To
identify the possible mechanisms underlying the apparent anticancer
effect of SAC, the expression of apoptosis-associated proteins,
including PARP, caspases, Bcl-2, Bax, and AKT was examined. As
expected, SAC treatment reduced phosphorylation of AKT.
Furthermore, SAC markedly decreased the expression of Bcl-2,
whereas the expression of Bax was increased in a dose-dependent
manner, further confirming the promotion of apoptosis. There was
also a change in the expression of PARP and caspases, in favor of
apoptosis.
The results in the present study are consistent with
the findings of Xu et al (8).
Treatment of A2780 ovarian cancer cells with SAC induces S phase
arrest and apoptosis, accompanied by reduced expression of Bcl-2
and increased expression of caspase-3 and Bax (8). Accordingly, the authors of the present
study hypothesize that SAC suppresses proliferation of T24 and
T24R2 cells through the induction of apoptosis.
In conclusion, SAC inhibits cell proliferation and
clonogenicity through the induction of apoptosis and cell cycle
arrest by altering the expression of apoptosis and cell cycle
regulators in bladder cancer cells. The findings of the present
study suggest that SAC is a promising agent for the treatment of
bladder cancer.
Acknowledgements
The present study was funded by a research grant
from Ildong Pharmaceutical Co., Ltd. (grant no. 06-2015-028). SAC
was supplied by PharmaKing Co., Ltd. (Gyeonggi-do, Republic of
Korea).
References
|
1
|
Yun SJ and Kim WJ: Role of the
epithelial-mesenchymal transition in bladder cancer: From prognosis
to therapeutic target. Korean J Urol. 54:645–650. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Song W and Jeon HG: Incidence of kidney,
bladder, and prostate cancers in Korea: An update. Korean J Urol.
56:422–428. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Arantes-Rodrigues R, Pinto-Leite R,
Fidalgo-Gonçalves L, Palmeira C, Santos L, Colaço A and Oliveira P:
Synergistic effect between cisplatin and sunitinib malate on human
urinary bladder-cancer cell lines. Biomed Res Int. 2013:7914062013.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Pliarchopoulou K, Laschos K and Pectasides
D: Current chemotherapeutic options for the treatment of advanced
bladder cancer: A review. Urol Oncol. 31:294–302. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sávio AL, da Silva GN and Salvadori DM:
Inhibition of bladder cancer cell proliferation by allyl
isothiocyanate (mustard essential oil). Mutat Res. 771:29–35. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Makarević J, Rutz J, Juenqel E, Kaulfuss
S, Reiter M, Tsaur I, Bartsch G, Haferkamp A and Blaheta RA:
Amygdalin blocks bladder cancer cell growth in vitro by diminishing
cyclin A and cdk2. PLos One. 9:e1055902014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Nq KT, Guo DY, Cheng Q, Geng W, Ling CC,
Li CX, Liu XB, Ma YY, Lo CM, Poon RT, et al: A garlic derivative,
S-allylcysteine (SAC), suppresses proliferation and metastasis of
hepatocellular carcinoma. PLoS One. 7:e316552012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Xu YS, Feng JG, Zhang D, Zhang B, Luo M,
Su D and Lin NM: S-allylcysteine, a garlic derivative, suppresses
proliferation and induces apoptosis in human ovarian cancer cells
in vitro. Acta Pharmacol Sin. 35:267–274. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liu Z, Li M, Chen K, Yang J, Chen R, Wang
T, Liu J, Yang W and Ye Z: S-allylcysteine induces cell cycle
arrest and apoptosis in androgen-independent human prostate cancer
cells. Mol Med Rep. 5:439–443. 2012.PubMed/NCBI
|
|
10
|
Chung LY: The antioxidant properties of
garlic compounds: Allyl cysteine, alliin, allicin, and allyl
disulfide. J Med Food. 9:205–213. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Garcia E, Santana-Martinez R, Silva-Islas
CA, Colín-González AL, Galván-Arzate S, Heras Y, Maldonado PD,
Sotelo J and Santamaría A: S-allyl cysteine protects against
MPTP-induced striatal and nigral oxidative neurotoxicity in mice:
Participation of Nrf2. Free Radic Res. 48:159–167. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Byun SS, Kim SW, Choi H, Lee C and Lee E:
Augmentation of cisplatin sensitivity in cisplatin-resistant human
bladder cancer cells by modulating glutathione concentrations and
glutathione-related enztme activities. BJU Int. 95:1086–1090. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yeo EJ, Ryu JH, Chun YS, Cho YS, Jang IJ,
Cho H, Kim J, Kim MS and Park JW: YC-1 induces S cell cycle arrest
and apoptosis by activating checkpoint kinases. Cancer Res.
66:6345–6352. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yu JS, Kim CB, Kim KK, Lee JE and Kim MY:
Behaviors of providers of traditional korean medicine therapy and
complementary and alternative medicine therapy for the treatment of
cancer patients. J Pharmacopuncture. 18:27–35. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Stubbe CE and Valero M: Complementary
strategies for the management of radiation therapy side effects. J
Adv Pract Oncol. 4:219–231. 2013.PubMed/NCBI
|
|
16
|
Philippou Y, Hadjipavlou M, Khan S and
Rane A: Complementary and alternative medicine (CAM) in prostate
and bladder cancer. BJU Int. 112:1073–1079. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen JJ, Ye ZQ and Koo MW: Growth
inhibition and cell cycle arrest effects of epigallocatechin
gallate in the NBT-II bladder tumour cell line. BJU Int.
93:1082–1086. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bai Y, Mao QQ, Qin J, Zheng XY, Wang YB,
Yang K, Shen HF and Xie LP: Resveratrol induces apoptosis and cell
cycle arrest of human T24 bladder cancer cells in vitro and
inhibits tumor growth in vivo. Cancer Sci. 101:488–493. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang K, Wu J, Wu X, Wang X, Wang Y, Zhou
N, Kuo ML, Liu X, Zhou B, Chang L, et al: p53R2 inhibits the
proliferation of human cancer cells in association with cell-cycle
arrest. Mol Cancer Tehr. 10:269–278. 2011. View Article : Google Scholar
|
|
20
|
Goldar S, Khaniani MS, Derakhshan SM and
Baradaran B: Molecular mechanisms of apoptosis and roles in cancer
development and treatment. Asian Pac J Cancer Prev. 16:2129–2144.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Fiandalo MV and Kyprianou N: Caspase
control: Protagonists of cancer cell apoptosis. Exp Oncol.
34:165–175. 2012.PubMed/NCBI
|