Introduction
The primary cause of cervical carcinoma (CC) is
persistent infection with high-risk human papillomavirus (HR-HPV)
(1). The infecting HPV type, viral
persistence, viral integration status and viral load contribute to
the pathogenesis of CC (1). Among the
HR-HPVs, HPV 16 and 18 are the most common in cases of CC globally
(2).
CC was the fourth most common type of cancer among
women globally in 2012 (3). In
Mexico, it is the second most frequent cancer type amongst women
and the most common female cancer type in women aged 15–44 years,
with ~9.4% of women in the general population estimated to harbor
cervical HPV infection, and 70.2% of all CC cases being attributed
to HPV 16 or 18 (4). In the state of
Guerrero in Southern Mexico, CC is the most common type of cancer
in women. This state also has the fifth highest mortality rate from
CC in the country (5). HPV 16 is the
most common genotype in CC, followed by HPV 18 (6).
Cytological screening of Papanicolaou smears has
reduced the incidence and mortality rates of CC. However, the low
sensitivity and low reproducibility of cytological tests, despite
the availability of HPV DNA testing as an adjunct procedure, has
resulted in a lack of discrimination between transient and
persistent infections (7).
HPV genomes replicate as episomal DNA during
productive infections, and the integration of HR-HPV genomes into
the host chromosome has been associated with neoplastic progression
(8). The E2 HPV gene has been
revealed to be broken more frequently compared with other sites
(9–11). This break results in a loss of the
negative-feedback control for the transcription of oncogenes E6 and
E7 (12), enabling tumor development
and progression (13,14). Therefore, HPV integration into the
human genome is considered to be a notable event in cervical
carcinogenesis. Although certain studies suggest that integration
is a late event (14,15), others have proposed that integration
may be an early event in carcinogenesis (16–20).
Based on this information, a quantitative polymerase
chain reaction (qPCR) analysis method that uses the PCR
amplification of the E2 and E6 open reading frames (ORFs) and a
calculation of E2/E6 ratios in order to measure integration was
developed (20). Identification of
the integrated HPV has been proposed as a useful biomarker for
lesion progression. Nevertheless, qPCR is a technically difficult
method for use in clinical practice.
HPV integration may render viral and cellular genes
susceptible to epigenetic changes, which may in turn regulate their
expression (21). The L1 gene in HPV
16 and 18 has been observed to be frequently hypermethylated in
cancer, compared with the infrequent methylation observed in
premalignant lesions (18,22–26). It
has been reported that the methylation of the HPV 16 L1 gene
indicates integration and is associated with the grade of
premalignant lesions and cancer types, as well as being a potential
clinical marker of CC progression (18,27–29).
HPV integration and the grade of cervical neoplasia
may also be identified using a biotinyl tyramide-based in
situ hybridization (ISH) method. Punctate signals within the
nucleus are consistent with the integration of HPV into host cell
chromosomes, whereas diffuse signal patterns throughout the nucleus
are consistent with episomal DNA (30–33).
The use of novel tools alongside the cytological
diagnosis of premalignant lesions may improve the sensitivity of
this method, increasing its effectiveness, which would reduce the
number of misdiagnosed cases and prevent unnecessary invasive
treatments in women diagnosed with premalignant lesions. As HPV
integration is a viral marker for lesion progression, it is
important to use a technically simple and affordable method for the
detection of HPV integration in clinical practice.
The aim of the present study was to analyze L1 gene
methylation in cervical lesions of various grades, and to perform a
comparative analysis between L1 methylation, ISH and RT-qPCR as
methods to determine the physical status of HPV 16 and 18.
Additionally, the ease of use of these methods was compared.
Materials and methods
Clinical specimens
A total of 187 female patients between 28–51 years
of age were enrolled in the present study, selected from the 1,115
female residents of the state of Guerrero in Southern Mexico. The
patients were admitted to the Unit for Early Diagnosis of Cervical
Cancer and HPV at the Autonomous University of Guerrero
(Chilpancingo, Mexico) and the Cancer Institute of the State of
Guerrero (Acapulco, Mexico) between November 2013 and August 2015.
Cervical scrapes or biopsies were collected for cytomorphological
examination using liquid cytology or histological examination.
Papanicolau smears were reviewed by a certified cytopathologist and
biopsies by a certified histopathologist. DNA was extracted from
the cervical scrapes or biopsies using the phenol-chloroform
isoamyl alcohol method (34) for HPV
detection and genotyping with an INNO-LiPA genotyping kit
(Fujirebio Europe, Gent, Belgium) (35), without amendment and according the
manufacturer's protocol.
All patients included in the current study presented
with HPV 16 (n=154) or HPV 18 (n=33), and CC (n=33), high-grade
squamous intraepithelial lesion (HSIL; n=13), low-grade squamous
intraepithelial lesion (LSIL; n=102) or no intraepithelial lesion
(non-IL; n=39). Cytological diagnosis was performed according to
the Bethesda System (36), and
histological diagnosis was performed according to the
classification system of the International Federation of Gynecology
and Obstetrics (37). Clinical data
from the patients was obtained from the database of the Molecular
Biomedicine and Cytopathology Laboratories at the School of
Chemistry and Biology, Autonomous University of Guerrero.
The present study was approved by the Bioethics
Committee of the Autonomous University of Guerrero. Written
informed consent was obtained from all patients prior to the
study.
The samples were studied to analyze L1 gene
methylation in the different grades of cervical lesion and to
compare L1 methylation, ISH and qPCR as methods for determining the
physical status of HPV 16 and 18.
Analysis of the methylation of the HPV
16 and HPV 18 L1 gene
Methylation of the L1 gene from HPV 16 and 18 was
analyzed using a bisulfite conversion reaction, wherein
unmethylated cytosine is converted into uracil, followed by PCR
amplification. DNA (400–1,200 ng) from cervical samples was treated
with bisulfite for 150 min at 64°C using an EZ DNA
Methylation-Gold™ kit (Zymo Research Corp., Irvine, CA, USA),
according to the manufacturer's protocol.
The 5′ and middle regions of the HPV 16 L1 gene, and
the 5′ and 3′ regions of the HPV 18 L1 gene were amplified using
PCR. Amplifications were performed using 100–150 ng
bisulfite-treated genomic DNA, 6 µl Amplitaq Gold® 360
Master mix (Applied Biosystems; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA), 1 µmol sense primer and 1 µmol antisense primer
in a 15-µl final reaction volume. The following primers were used
for amplification: HPV 16 L1 5′ region forward,
5′-TTGTTGATGTAGGTGATTTTTATTTATATTTTAGTTCCA-3′ and reverse,
5′-CTAATACCCACACCTAATAACTAACC-3′ as previously described by Oka
et al (28) at 59°C, HPV 16 L1
middle region forward, 5′-ATTTAGATTATATTAAAATGGTGTTAGAAT-3′ and
reverse, 5′-AAATAATTAATTACCCCAACAAATACC-3′ as previously described
by Brandsma et al (27) at
54°C. HPV 18 L1 5′ region forward, 5′-GGTTAGTTTTTAGGTGTTGGT-3′ and
reverse, 5′-CACACAACTACCAAATAAAACA-3′ (Tm, 54°C); and HPV 18 L1 3′
region forward, 5′-TTATTAGTTTGGTGGATATATATTG-3′ and reverse,
5′-AAAACATACAAACACAACAATAAATA-3′ as previously described by Badal
et al (38), at 59°C. The
thermocycling conditions were as follows: 10 min at 96°C, followed
by 40 cycles for 30 sec at 96°C, 30 sec at 54°C or 59°C, depending
on the primer used, and 7 min at 72°C. The PCR-amplified fragments
were sequenced using the Big Dye Terminator version 3.1 Cycle
Sequencing in an ABI PRISM 310 sequencer (Applied Biosystems;
Thermo Fisher Scientific, Inc.). The methylation of 12 (5602 to
6580) and 18 (6015 to 7123) CpG dinucleotides in the HPV 16 and HPV
18 L1 gene, respectively, was analyzed using LALIGN online platform
(http://embnet.vital-it.ch/software/LALIGN_form.html).
For the analysis of the L1 gene methylation variation in the
samples, the L1 gene methylation ratio (L1MR) was calculated using
the formula L1MR (%)=(number of methylated CpGs in the analyzed
region of the L1 gene)/(number of all CpGs in the analyzed region
of the L1 gene) ×100. Oka et al (28) suggested that the L1MR is associated
with the grade of cervical intraepithelial neoplasia (CIN) and
indicates the HPV genome status in the cell, as follows: A high
L1MR value indicates HPV genome integration, whereas a low L1MR
indicates episomal HPV genomes. Complete methylation (>80%) is
considered to indicate HPV integration, intermediate methylation
(20–80%) is considered to indicate a mixed status and low
methylation (<20%) is considered to indicate an episomal HPV
genome (39).
Analysis of HPV 16 and 18 integrations
using qPCR
The physical state of HPV 16 was analyzed using
qPCR, amplifying two regions of the E2 gene and a region of the E6
gene using the following primers and TaqMan probes (Applied
Biosystems; Thermo Fisher Scientific, Inc.): E2A region
(transactivation domain) forward, 5′-TGGATATACAGTGGAAGTGCAGTTTG-3′
and reverse, 5′-CCACAGTTACTGATGCTTCTTCACA-3′ primers, and
5′FAM-ATGGAGACATATGCAATACAAT-NFQ probe; E2B region (hinge) forward,
5′-CAGCAACGAAGTATCCTCTCCT-3′ and reverse, 5′-GCGACGGCTTTGGTATGG-3′
primers, and 5′FAM-AAGTGCTGCCTAATAATTTC-NFQ probe; and E6 region
forward, 5′-ACCGTTGTGTGATTTGTTAATTAGGTGTA-3′ and reverse,
5′-GCTTTTTGTCCAGATGTCTTTGCTT-3′ primers, and
5′FAM-AAAGCCACTGTGTCCTG-NFQ probe.
Similarly, the physical state of HPV 18 was analyzed
by amplifying two regions of the E2 gene and a region of the E6
gene using the following primers and TaqMan probes (Applied
Biosystems; Thermo Fisher Scientific, Inc.): E2N1 region
(transactivation domain) forward, 5′-GCAACTAATACGTTGGGAAAATGCA-3′
and reverse, 5′-GCTGGCACCACCTGGT-3′ primers, and
5′FAM-CTGTATGCCATGTTCCC-NFQ probe; E2H1 region forward,
5′-GATTGTAATGACTCTATGTGCAGTACCA-3′ and reverse,
5′-CGGTGCCCACGGACA-3′ primers, and FAM-ACGACACGGTATCCGC-NFQ probe;
and E6 region forward, 5′-AAAACTAACTAACACTGGGTTATACAATT-3′ and
reverse, 5′-CTGCTGGATTCAACGGTTTCTG primers, and
5′FAM-CACCGCAGGCACCTTA-NFQ probe.
To calculate the number of cells, a region of the
RNAse P single copy gene was amplified using forward,
5′-AGATTTGGACCTGCGAGCG-3′ and reverse, 5′-GAGCGGCTGTCTCCACAAGT-3′
primers, and a VIC-TTCTGACCTGAAGGCTCTGCGCG-TAMRA RNAse P probe (cat
no. 4403328 Applied Biosystems; Thermo Fisher Scientific,
Inc.).
In all cases, PCR was performed in a final volume of
10 µl containing 50 ng DNA, 5 µl Universal MasterMix II (Applied
Biosystems; Thermo Fisher Scientific, Inc.) and 0.5 µl of a mix
containing specific primers and probes for each gene and region.
DNA amplification by qPCR was performed in an ABI-PRISM 7500
(Applied Biosystems; Thermo Fisher Scientific, Inc.) using the
following thermocycler conditions: 50°C for 2 min, 95°C for 10 min,
followed by 40 cycles of 95°C for 15 sec and 60°C for 1 min as
previously described by Zhang et al (40). All reactions were run in duplicate,
and the results were averaged for analysis. The positive control
consisted of DNA from the SiHa cell line which contains 1–2
integrated copies of the HPV 16 genome as previously described by
Flores et al (41) and Saunier
et al (42), and the HeLa cell
line, which contains integrated HPV 18. The negative controls
consisted of the reaction components without DNA.
A standard curve for the quantification of the RNAse
P gene was generated using the commercial kit Taqman Copy Number
Reference Assay RNAse P (Applied Biosystems; Thermo Fisher
Scientific) with genomic DNA obtained from 10,000, 5,000, 2,500,
1,250 and 625 cells. Standard curves for the E2 and E6 genes were
generated by determining the quantification cycle (Cq) (43) value of detection with concentrations
of 108, 107, 106, 105,
104 and 103 copies/µl of plasmids containing
the HPV 16 or HPV 18 genome as was performed by Zhang et al
(40). All assays were performed in
duplicate. The number of copies of each gene (x) in the sample was
calculated with the formula x=exp[(Y-b)/m] where Y is the Cq value,
and b and m are constant values obtained from the equation graph
for the standard curve of each gene (44).
The physical state of HPV 16 was determined by
calculating the ratios of E2A/E6 and E2B/E6. To ensure the validity
of the method, these ratios were determined using the standard
curves obtained from various dilutions of pUC8 and DNA from SiHa
cells; pUC8 contains the complete HPV 16 genome, representing the
episomal form, and SiHa cells were used as the integrated form
(44). Viral genome integration was
defined by the absence of E2 or an E2/E6 ratio of <0.0010. An
E2A/E6 ratio of 0.0010–0.9270 was defined as a mixture of episomal
and integrated forms. A ratio of >0.9270 was defined as the
episomal form. An E2B/E6 ratio of 0.0010–0.9116 was defined as
mixed form, and an E2B/E6 ratio of >0.9116 was defined as the
episomal form (40).
The physical state of HPV 18 was determined by
calculating the ratios of E2N1/E6 and E2H1/E6. To ensure the
validity of this method, these ratios were determined using
standard curves obtained from various dilutions of a plasmid
containing the HPV 18 genome, representing the episomal form. Viral
genome integration was defined by the absence of E2 or an E2/E6
ratio of <0.0010. An E2N1/E6 ratio of 0.0010–0.9151 was defined
as the mixed form, and >0.9151 was defined as the episomal form.
An E2H1/E6 ratio of 0.0010–0.9206 was defined as the mixed form,
and an E2H1/E6 ratio >0.9206 was defined as the episomal form
(40).
Analysis of HPV 16 and 18 integration
using ISH
Detection of HR-HPV genome integration was performed
using ISH with a GenPoint Tyramide Signal Amplification system
(Dako; Agilent Technologies, Inc., Santa Clara, CA, USA).
Cytological samples were placed in a solution of acetone for 5 min
at room temperature. The monolayer smears were digested for 1 min
at 37°C with proteinase K (Invitrogen; Thermo Fisher Scientific,
Inc.; 1:1,000). Biotinylated viral DNA with probes for 13 HR-HPV
genotypes (types 16, 18, 31, 33, 35, 39, 45, 51, 52, 56, 58, 59 and
68) and individual probes (Dako; Agilent Technologies, Inc.)
directed at HPV 6 and HPV 11 were subsequently applied. The slides
were denatured for 10 min at 95°C, hybridized with a biotinylated
HPV DNA probe (Dako; Agilent Technologies, Inc.) for 20 h at 37°C
and then placed in an astringent solution (1:50) at 55°C for 20
min. Primary streptavidin peroxidase (1:100) was added for 40 min
at room temperature, followed by biotinyl-tyramide for 20 min at
37°C, and horseradish peroxidase conjugated streptavidin (cat no.
N100; Thermo Fisher Scientific, Inc.) undiluted, as the secondary
antibody, was subsequently added for 20 min at 37°C. The chromogen
3,3′-diaminobenzidene (Dako; Agilent Technologies, Inc.) was then
added and Mayer's hematoxylin (Merck KGaA, Darmstadt, Germany) at
37°C for 10 sec was used as a contrast dye. Positive reactions were
visualized as brown inside of the nucleus in a LEICA DM 1000 light
microscope and, according to the type of signal, were classified as
diffuse (episomal state), punctate (integrated state) or diffuse
and punctate (mixed state) (30,31,33). SiHa
cell lines (containing HPV 16) were used as positive controls to
demonstrate an integrated state; the same cell lines without probes
were used as negative controls.
Statistical analysis
L1MR was compared between groups using one-way
analysis of variance and Tukey's multiple comparison test as a
post-hoc test using GraphPad Prism V5-03 software (GraphPad
Software, Inc., La Jolla, CA, USA). P<0.05 was considered to
indicate a statistically significant difference. Odds ratios (ORs)
were calculated to determine the risk of lesion progression due to
the methylation of specific CpG sites using STATA version 11
software (StataCorp LLC, College Station, TX, USA; http://www.stata.com/). Concordance between the
various methods for the detection of viral integration was also
calculated (45).
Results
Methylation of the HPV 16 and HPV 18
L1 gene in cervical specimens from women with CC, SIL and
non-IL
The methylation status of the HPV 16 and HPV 18 L1
gene was evaluated in 187 cervical specimens from women with CC,
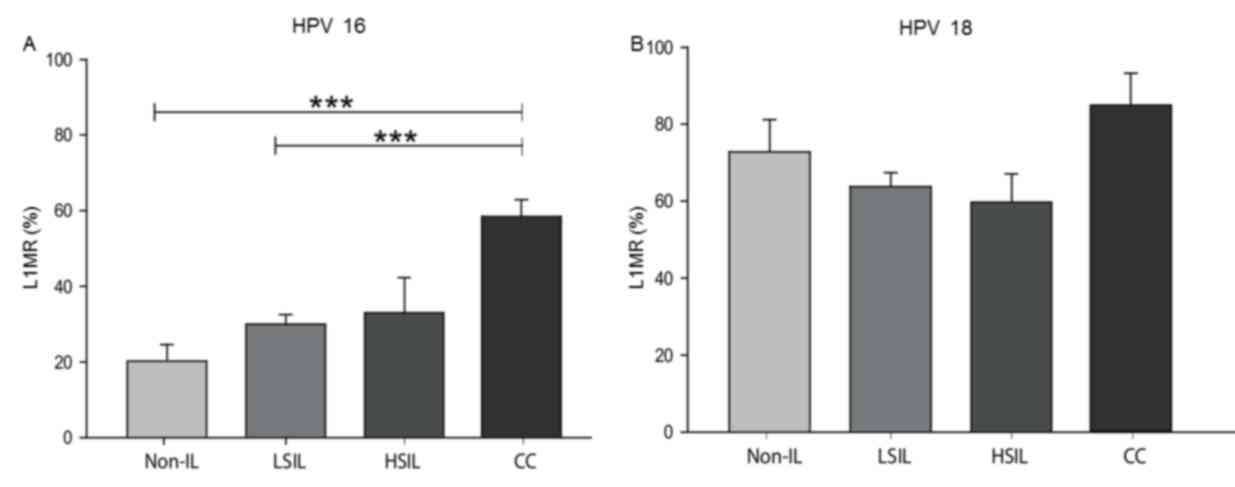
HSIL, LSIL and non-IL. The median HPV 16 L1MRs were 20.2% for
non-IL, 30.0% for LSIL, 33.0% for HSIL and 58.6% for CC. The HPV 16
L1MR was associated with histological grade, as statistically
significant differences were identified in the following
comparisons: LSIL vs. CC (P<0.0001) and non-IL vs. CC
(P<0.0001; Fig. 1A). For HPV 18,
the median L1MRs were 73.3% for non-IL, 64.1% for LSIL, 69.7% for
HSIL and 85.7% for CC; however, these differences between
histological grades were not significant (Fig. 1B).
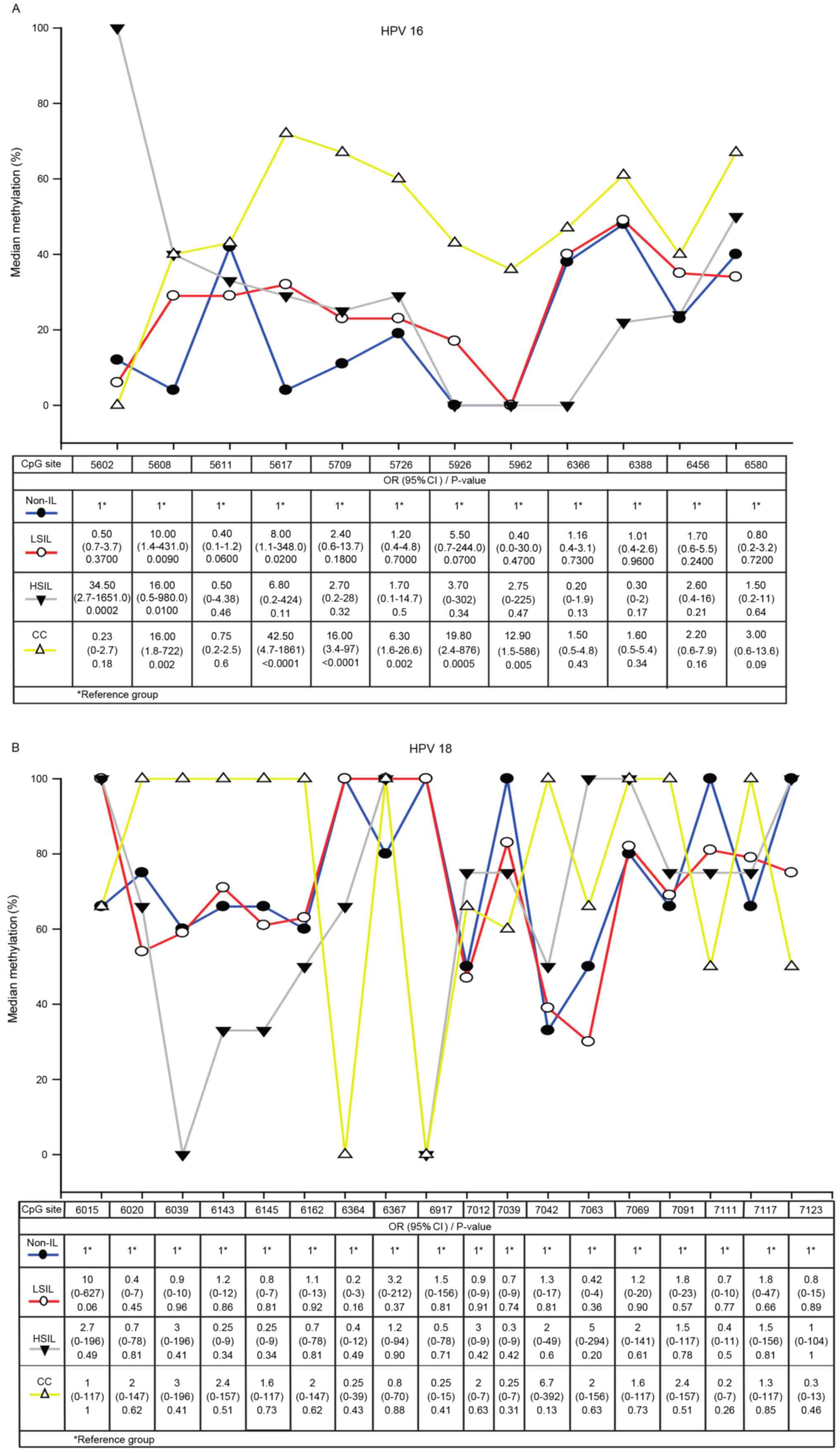
A total of 12 CpG dinucleotides (from 5602 to 6580)
in the HPV 16 L1 gene and 18 CpG sites (from 6015 to 7123) in the
HPV 18 L1 gene were analyzed in specimens from patients with
non-IL, LSIL, HSIL and CC. In HPV 16, the methylation of CpG sites
5608 and 5617 were associated with LSIL, the methylation of CpG
sites 5602 and 5608 were associated with HSIL, and CpG sites 5608,
5617, 5709, 5726, 5926, 5962 were associated with CC. The strongest
association with CC was CpG site 5617 (OR, 42.5; 95% confidence
interval, 4.7–1861; P<0.0001), followed by CpG sites 5926, 5608,
5709 and 5962. Only CpG site 5608 was associated with all grades of
cervical lesions. In CC samples, the highest L1MR was at CpG site
5617 (71.4%) (Fig. 2A).
High L1MRs at CpG sites in HPV 18 were detected in
all grades of cervical lesions. 100% methylation was detected at 10
CpG sites in CC, 5 CpG sites in HSIL, 5 CpG sites in LSIL and 5 CpG
sites in non-IL samples. However, the methylation differences in
CC, HSIL, LSIL compared with non-IL were not significant (Fig. 2B).
Analysis of HPV 16 and HPV 18
integration using qPCR, ISH and L1 methylation
The physical statuses of HPV 16 and HPV 18 were
evaluated using qPCR, ISH and L1 methylation in cervical specimens
from CC, HSIL, LSIL and non-IL. HPV 18 was more frequently
integrated compared with HPV 16; HPV 18 was integrated in 50%
(qPCR), 100% (ISH) and 67% (L1 methylation) of CC specimens, whilst
HPV 16 was integrated in 52% (qPCR), 92% (ISH) and 16% (L1
methylation) of CC specimens. Non-IL and LSIL demonstrated high
rates of integration of HPV 16, as follows: 50 and 39% by qPCR, 33
and 45% by ISH, and only 6 and 7% by L1 methylation, respectively.
HPV 18 was not integrated in non-IL, according to the qPCR and ISH
analyses, but did exhibit integration in LSIL (10% by qPCR, 53% by
ISH and 25% by L1 methylation; Table
I).
 | Table I.Physical status of HPV 16 and HPV 18
determined by qPCR, ISH and L1 methylation. |
Table I.
Physical status of HPV 16 and HPV 18
determined by qPCR, ISH and L1 methylation.
| A, HPV 16 physical
status, n (%) |
|---|
|
|---|
|
| qPCR (n=100) | ISH (n=140) | L1 methylation
(n=154) |
|---|
|
|
|
|
|
|---|
| Diagnosis | Episomal (n=2) | Mixed (n=52) | Integrated
(n=46) | Episomal (n=4) | Mixed (n=66) | Integrated
(n=70) | Episomal
(n=51) | Mixed (n=91) | Integrated
(n=12) |
|---|
| Non-IL | 1 (4) | 12 (46) | 13 (50) | 3 (10) | 17 (57) | 10 (33) | 17 (52) | 16 (48) | 0 (6) |
| LSIL | 1 (2) | 27 (59) | 18 (39) | 1 (1) | 43 (54) | 36 (45) | 28 (34) | 48 (59) | 6 (7) |
| HSIL | 0 (0) | 1 (33) | 2 (67) | 0 (0) | 4 (67) | 2 (33) | 4 (45) | 4 (45) | 1 (10) |
| CC | 0 (0) | 12 (48) | 13 (52) | 0 (0) | 2 (8) | 22 (92) | 2 (7) | 23 (77) | 5 (16) |
|
| B, HPV 18
physical status, n (%) |
|
|
| qPCR
(n=19) | ISH
(n=30) | L1 methylation
(n=33) |
|
|
|
|
|
| Diagnosis | Episomal (n=5) | Mixed (n=12) | Integrated
(n=2) | Episomal (n=1) | Mixed (n=14) | Integrated
(n=15) | Episomal (n=0) | Mixed (n=23) | Integrated
(n=10) |
|
| Non-IL | 2 (67) | 1 (33) | 0 (0) | 1 (17) | 5 (83) | 0 (0) | 0 (0) | 4 (67) | 2 (33) |
| LSIL | 1 (10) | 8 (80) | 1 (10) | 0 (0) | 9 (47) | 10 (53) | 0 (0) | 15 (75) | 5 (25) |
| HSIL | 2 (50) | 2 (50) | 0 (0) | 0 (0) | 0 (0) | 2 (100) | 0 (0) | 3 (75) | 1 (25) |
| CC | 0 (0) | 1 (50) | 1 (50) | 0 (0) | 0 (0) | 3 (100) | 0 (0) | 1 (33) | 2 (67) |
The concordance rate between qPCR and ISH for the
detection of the physical status of HPV 16 and HPV 18 was 96.1%.
The concordance rates between qPCR and L1 methylation, and between
ISH and L1 methylation were 76.7 and 84.8%, respectively (Table II).
| Table II.Concordance rates of qPCR, ISH and
HPV L1 methylation in the detection of the physical status of HPV
16 and HPV 18 in clinical samples. |
Table II.
Concordance rates of qPCR, ISH and
HPV L1 methylation in the detection of the physical status of HPV
16 and HPV 18 in clinical samples.
| A, qPCR/ISH
(n=107) |
|---|
|
|---|
|
| qPCR |
|
|---|
|
|
|
|
|---|
| Physical
status |
Mixed/integrated | Episomal | Positive
concordance, %a |
|---|
| ISH |
|
| 96.1 |
|
Mixed/integrated | 98 | 2 |
|
|
Episomal | 6 | 1 |
|
|
| B, qPCR/L1
methylation (n=120) |
|
|
| qPCR |
|
|
|
|
|
| Physical
status |
Mixed/integrated |
Episomal | Positive
concordance, %a |
|
| L1 methylation |
|
| 76.7 |
|
Mixed/integrated | 74 | 7 |
|
|
Episomal | 38 | 0 |
|
|
| C, ISH/L1
methylation (n=172) |
|
|
| ISH |
|
|
|
|
|
| Physical
status |
Mixed/integrated |
Episomal | Positive
concordance, %a |
|
| L1 methylation |
|
| 84.8 |
|
Mixed/integrated | 123 | 2 |
|
|
Episomal | 42 | 3 |
|
Discussion
The purpose of the present study was to investigate
L1 gene methylation between various grades of cervical lesions
positive for HPV 16 or HPV 18. Additionally, L1 methylation, ISH
and qPCR were compared as methods with which to determine the
integration of HPV 16 and HPV 18 in cervical specimens, including
CC, premalignant lesions and non-cervical lesions.
The results indicated that the HPV 16 L1 gene
undergoes a significant progressive increase in methylation rate
according to the increase of the grade of the cervical lesion, in
concordance with previous studies (23,28,46–48).
However, higher methylation rates in non-IL and LSIL specimens were
observed in the present study compared with those in a previous
study (28). Methylation of L1
increases progressively through the four pathological categories
(non-IL, LSIL, HSIL and CC). However, methylation is also present
in the absence of a cervical lesion in tissue positive for HPV 16
(L1MR, 20.2% in non-IL).
A progressive increase in the methylation of the HPV
18 L1 gene was detected in non-IL, LSIL, HSIL and CC samples, with
no significant differences observed between the different grades of
cervical lesion. There are certain differences from the results
obtained from various previous studies. Badal et al
(38) reported the presence of L1
methylation in HPV 18-positive samples from CC and asymptomatic
patients, whereas Turan et al (25) reported unmethylated HPV 18 L1 genes in
samples from asymptomatic infections and infections with
premalignant lesions, but strong methylation in all CCs. Kalantari
et al (47) reported that
methylation of HPV 16 and HPV 18 is lowest in cases of asymptomatic
infection and increases progressively during the progression into
cancer.
Kalantari et al (47) hypothesized that highly methylated LSIL
and HSIL samples are those that have undergone certain molecular
changes that predispose them to develop into cancer. The present
study concurs with this hypothesis, however, the level of
methylation should be taken into account even in patients without
cervical lesions, as this may indicate the possibility of lesion
progression.
The usefulness of specific sites of methylation in
the HPV genome in identifying women who are at an increased risk of
developing CC has been suggested previously (24,48–50).
In the present study, 12 methylation CpG sites of
the HPV 16 L1 gene were analyzed in samples from non-IL, LSIL, HSIL
and CC tissues. Methylation of the 5608 CpG site was associated
with the development of LSIL, HSIL and CC, whilst the methylation
of the 5617 CpG site was associated with LSIL and CC, and the
methylation of the 5709, 5726, 5926 and 5962 CpG sites was
associated exclusively with CC. Mirabello et al (48) identified a strong association between
methylation at the 6457 CpG site and CIN2 (HSIL), and in a
subsequent study a strong association between methylation at the
5602, 5608, 5611 and 5617 CpG sites and cervical precancerous
tissue was revealed (50). Sun et
al (29) revealed increased
methylation of the CpG sites in HPV 16 L1 associated with CIN1,
CIN2 and CIN3 (LSIL/HSIL). Lorincz et al (24) reported a higher methylation of the
6367 and 6389 L1 CpG sites in women with CIN2/3. Brandsma et
al (51) reported that the
frequency of methylation was the highest for CIN3, followed by CIN2
and CIN1 at the 5602, 5608, 5709, 5726, 6367, 6389, 6581 (referred
to as 5600, 5606, 5707, 5724, 6365, 6387 and 6579) L1 CpG sites.
The results of the present study coincide with the aforementioned
studies, and the observed differences in what the sites of CpG
methylation in the HPV 16 L1 gene have been identified to be may be
due to the circulating HPV 16 variants in each geographical region.
A previous study performed in the state of Guerrero in Southern
Mexico revealed 27 variants of HPV 16 that belong to European and
Asian-American sub-lineages (52).
Nucleotide differences between variants may determine changes in
HPV 16 methylation.
There is increasing evidence that the elevated
methylation of specific HPV 16 CpG sites in the L1 gene ORF may be
a biomarker of progression from premalignant lesions to CC. The
present study concurs with Mirabello et al (48) in that a methylation assay in one or a
small number of CpG sites would be easier to perform, minimize
costs and could potentially serve as a prognostic biomarker to
predict the development of premalignant lesions and CC.
In HPV 18, analysis of 18 CpG sites was performed in
all grades of cervical lesions, revealing that none of the sites
were significantly associated with the development of CC or
premalignant lesions. However, high L1MRs in a number of CpG sites
were detected in non-IL, LSIL, HSIL and CC, which may indicate that
potentially certain changes have occurred that predispose the
tissue to develop into cancer, as proposed by Kalantari et
al (46), considering that viral
methylation appears to occur years prior to the detection of
premalignant lesions (48).
The integration of HR-HPV genomes is a key event in
cervical carcinogenesis, as it contributes to neoplastic
transformation. The integration of viral genomes is a consequence
of chromosomal instability induced by deregulated E6 and E7
oncogene expression (53). There are
several methods that may be used in order to detect HPV integration
in the human genome, including qPCR (20), ISH (30,31) and L1
methylation (18). However, few
studies have compared such methods by which to determine the
physical state of HPV. Fujii et al (54) compared ISH with qPCR using SIL and CC
samples, reporting an 86% concordance rate between the two methods.
Biesaga et al (55)
additionally compared ISH with qPCR to assess the physical viral
genome state in CC samples and revealed no significant difference
between these methods, concluding that ISH may be used for the
assessment of viral genome status.
To the best of our knowledge, no previous studies
comparing the use of qPCR, ISH and L1 gene methylation assay
methods have been performed in order to evaluate HPV 16
integration. In the present study, these three methods were
compared. Good concordance was identified between them, and each
method possessed certain advantages. qPCR is a useful technique for
detecting integrated and episomal forms of HPV. This method has the
advantage of additionally calculating the viral copy numbers for
episomal and integrated HPV, making it a prognostic tool for
determining the risk of CC (20). L1
methylation is a frequent result of integration. Therefore, it has
been proposed that L1 gene methylation indicates integration, and
may be used as a potential biomarker of cancer progression
(18,56). ISH with signal amplification by
biotinyl tyramide to demonstrate HPV DNA has the advantage of
preserving the histological context of a lesion and allows the
detection of low-copy DNA (57). It
is generally accepted that a diffuse signal indicates an episomal
status and that punctate signals indicate integrated HPV (31). The ISH assay with signal amplification
by biotinyl tyramide is convenient for clinical purposes (58). According to the results of the present
study, it has a high concordance rate with qPCR (96.1%) and also
with the HPV L1 gene methylation assay (84.8%).
In conclusion, methylation of the HPV 16 L1 gene
increases significantly according to the grade of cervical lesion.
CpG site 5608 is associated with premalignant lesions and CC,
whereas CpG site 5617 had the strongest association with CC;
therefore, these two sites may be useful as prognostic biomarkers.
Furthermore, ISH and L1 methylation analyses have good concordance
with qPCR and are, therefore, useful methods for determining the
physical state of HPV. However, the combination of these techniques
may provide more reliable results.
Acknowledgements
The present study was supported by the National
Council of Science and Technology (grant no. 201579). The authors
would like to thank Dr Marco Antonio Jiménez-López of the Cancer
Institute of the State of Guerrero ‘Dr. Arturo Beltrán Ortega’ for
the histopathological diagnosis, and the technicians of the
Cytopathology and Histochemistry Laboratory (School of Chemical and
Biological Sciences, Autonomous University of Guerrero) for their
assistance.
References
|
1
|
zur Hausen H: Papillomaviruses and cancer:
From basic studies to clinical application. Nat Rev Cancer.
2:342–350. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Muñoz N, Bosch FX, de Sanjosé S, Herrero
R, Castellsagué X, Shah KV, Snijders PJ and Meijer CJ:
International Agency for Research on Cancer Multicenter Cervical
Cancer Study Group: Epidemiologic classification of human
papillomavirus types associated with cervical cancer. N Engl J Med.
348:518–527. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ferlay J, Soerjomataram I, Ervik M,
Dikshit R, Eser S, Mathers C, Rebelo M, Parkin DM, Forman D and
Bray F: GLOBOCAN 2012 v1.0, Cancer Incidence and Mortality
Worldwide: IARC Cancer Base No. 11[Internet]. Lyon, France:
International Agency for Research on Cancer; 2013, http://globocan.iarc.fr17–09. 2017
|
|
4
|
Bruni LB-R L, Serrano B, Brotons M, Cosano
R, Munoz J, Bosch FX, de Sanjosé S and Castellsagué X: ICO
Information centre on HPV and cancer (HPV Information Centre).
Human Papillomavirus and Related Diseases in México. Summary
Reports. 2014.
|
|
5
|
de Secretaría Salud: Información
estadística. Estadísticas de cáncer cervicouterino. https://www.gob.mx/cms/uploads/attachment/file/12951/MortalidadCaCu2000a2013.pdf30–05.
2016
|
|
6
|
Illades-Aguiar B, Alarcón-Romero Ldel C,
Antonio-Véjar V, Zamudio-López N, Sales-Linares N, Flores-Alfaro E,
Fernández-Tilapa G, Vences-Velázquez A, Muñoz-Valle JF and
Leyva-Vázquez MA: Prevalence and distribution of human
papillomavirus types in cervical cancer, squamous intraepithelial
lesions, and with no intraepithelial lesions in women from Southern
Mexico. Gynecol Oncol. 117:291–296. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Pinto AP, Degen M, Villa LL and Cibas ES:
Immunomarkers in gynecologic cytology: The search for the ideal
‘biomolecular Papanicolaou test’. Acta Cytol. 56:109–121. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Pett M and Coleman N: Integration of
high-risk human papillomavirus: A key event in cervical
carcinogenesis? J Pathol. 212:356–367. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Badaracco G, Venuti A, Sedati A and
Marcante ML: HPV16 and HPV18 in genital tumors: Significantly
different levels of viral integration and correlation to tumor
invasiveness. J Med Virol. 67:574–582. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kalantari M, Karlsen F, Kristensen G, Holm
R, Hagmar B and Johansson B: Disruption of the E1 and E2 reading
frames of HPV 16 in cervical carcinoma is associated with poor
prognosis. Int J Gynecol Pathol. 17:146–153. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Romanczuk H and Howley PM: Disruption of
either the E1 or the E2 regulatory gene of human papillomavirus
type 16 increases viral immortalization capacity. Proc Natl Acad
Sci USA. 89:3159–3163. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jeon S and Lambert PF: Integration of
human papillomavirus type 16 DNA into the human genome leads to
increased stability of E6 and E7 mRNAs: Implications for cervical
carcinogenesis. Proc Natl Acad Sci USA. 92:1654–1658. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kalantari M, Blennow E, Hagmar B and
Johansson B: Physical state of HPV16 and chromosomal mapping of the
integrated form in cervical carcinomas. Diagn Mol Pathol. 10:46–54.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kalantari M, Villa LL, Calleja-Macias IE
and Bernard HU: Human papillomavirus-16 and −18 in penile
carcinomas: DNA methylation, chromosomal recombination and genomic
variation. Int J Cancer. 123:1832–1840. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
zur Hausen H: Papillomaviruses causing
cancer: Evasion from host-cell control in early events in
carcinogenesis. J Natl Cancer Inst. 92:690–698. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gallo G, Bibbo M, Bagella L, Zamparelli A,
Sanseverino F, Giovagnoli MR, Vecchione A and Giordano A: Study of
viral integration of HPV-16 in young patients with LSIL. J Clin
Pathol. 56:532–536. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Huang LW, Chao SL and Lee BH: Integration
of human papillomavirus type-16 and type-18 is a very early event
in cervical carcinogenesis. J Clin Pathol. 61:627–631. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kalantari M, Chase DM, Tewari KS and
Bernard HU: Recombination of human papillomavirus-16 and host DNA
in exfoliated cervical cells: A pilot study of L1 gene methylation
and chromosomal integration as biomarkers of carcinogenic
progression. J Med Virol. 82:311–320. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kulmala SM, Syrjänen SM, Gyllensten UB,
Shabalova IP, Petrovichev N, Tosi P, Syrjänen KJ and Johansson BC:
Early integration of high copy HPV16 detectable in women with
normal and low grade cervical cytology and histology. J Clin
Pathol. 59:513–517. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Peitsaro P, Johansson B and Syrjänen S:
Integrated human papillomavirus type 16 is frequently found in
cervical cancer precursors as demonstrated by a novel quantitative
real-time PCR technique. J Clin Microbiol. 40:886–891. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Das P, Thomas A, Mahantshetty U,
Shrivastava SK, Deodhar K and Mulherkar R: HPV genotyping and site
of viral integration in cervical cancers in Indian women. PLoS One.
7:e410122012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Bird A: DNA methylation patterns and
epigenetic memory. Genes Dev. 16:6–21. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kalantari M, Calleja-Macias IE, Tewari D,
Hagmar B, Lie K, Barrera-Saldana HA, Wiley DJ and Bernard HU:
Conserved methylation patterns of human papillomavirus type 16 DNA
in asymptomatic infection and cervical neoplasia. J Virol.
78:12762–12772. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lorincz AT, Brentnall AR, Vasiljević N,
Scibior-Bentkowska D, Castanon A, Fiander A, Powell N, Tristram A,
Cuzick J and Sasieni P: HPV16 L1 and L2 DNA methylation predicts
high-grade cervical intraepithelial neoplasia in women with mildly
abnormal cervical cytology. Int J Cancer. 133:637–644. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Turan T, Kalantari M, Calleja-Macias IE,
Cubie HA, Cuschieri K, Villa LL, Skomedal H, Barrera-Saldaña HA and
Bernard HU: Methylation of the human papillomavirus-18 L1 gene: A
biomarker of neoplastic progression? Virology. 349:175–183. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Turan T, Kalantari M, Cuschieri K, Cubie
HA, Skomedal H and Bernard HU: High-throughput detection of human
papillomavirus-18 L1 gene methylation, a candidate biomarker for
the progression of cervical neoplasia. Virology. 361:185–193. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Brandsma JL, Sun Y, Lizardi PM, Tuck DP,
Zelterman D, Haines GK III, Martel M, Harigopal M, Schofield K and
Neapolitano M: Distinct human papillomavirus type 16 methylomes in
cervical cells at different stages of premalignancy. Virology.
389:100–107. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Oka N, Kajita M, Nishimura R, Ohbayashi C
and Sudo T: L1 gene methylation in high-risk human papillomaviruses
for the prognosis of cervical intraepithelial neoplasia. Int J
Gynecol Cancer. 23:235–243. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sun C, Reimers LL and Burk RD: Methylation
of HPV16 genome CpG sites is associated with cervix precancer and
cancer. Gynecol Oncol. 121:59–63. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Evans MF, Aliesky HA and Cooper K:
Optimization of biotinyl-tyramide-based in situ hybridization for
sensitive background-free applications on formalin-fixed,
paraffin-embedded tissue specimens. BMC Clin Pathol. 3:22003.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Evans MF, Mount SL, Beatty BG and Cooper
K: Biotinyl-tyramide-based in situ hybridization signal patterns
distinguish human papillomavirus type and grade of cervical
intraepithelial neoplasia. Mod Pathol. 15:1339–1347. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Vega-Peña A, Illades-Aguiar B,
Flores-Alfaro E, López-Bayghen E, Leyva-Vázquez MA,
Castañeda-Saucedo E and Alarcón-Romero Ldel C: Risk of progression
of early cervical lesions is associated with integration and
persistence of HPV-16 and expression of E6, Ki-67, and telomerase.
J Cytol. 30:226–232. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zubillaga-Guerrero MI, Illades-Aguiar B,
Leyva-Vazquez MA, Flores-Alfaro E, Castañeda-Saucedo E, Muñoz-Valle
JF and Alarcón-Romero LC: The integration of HR-HPV increases the
expression of cyclins A and E in cytologies with and without
low-grade lesions. J Cytol. 30:1–7. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ausubel F, Brent R, Kingston R, Moore D,
Seidman JG, Smith JA and Struhl K: Short Protocols in Molecular
Biology. 2–3. 1995.
|
|
35
|
Kleter B, van Doorn LJ, Schrauwen L,
Molijn A, Sastrowijoto S, Schegget Ter J, Lindeman J, Harmsel Ter
B, Burger M and Quint W: Development and clinical evaluation of a
highly sensitive PCR-reverse hybridization line probe assay for
detection and identification of anogenital human papillomavirus. J
Clin Microbiol. 37:2508–2517. 1999.PubMed/NCBI
|
|
36
|
Solomon D, Davey D, Kurman R, Moriarty A,
O'Connor D, Prey M, Raab S, Sherman M, Wilbur D, Wright T Jr, et
al: The 2001 Bethesda system: Terminology for reporting results of
cervical cytology. JAMA. 287:2114–2119. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Benedet JL, Bender H, Jones H III, Ngan HY
and Pecorelli S: FIGO staging classifications and clinical practice
guidelines in the management of gynecologic cancers. FIGO Committee
on Gynecologic Oncology. Int J Gynaecol Obstet. 70:209–262. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Badal S, Badal V, Calleja-Macias IE,
Kalantari M, Chuang LS, Li BF and Bernard HU: The human
papillomavirus-18 genome is efficiently targeted by cellular DNA
methylation. Virology. 324:483–492. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Reuschenbach M, Huebbers CU, Prigge ES,
Bermejo JL, Kalteis MS, Preuss SF, Seuthe IM, Kolligs J, Speel EJ,
Olthof N, et al: Methylation status of HPV16 E2-binding sites
classifies subtypes of HPV-associated oropharyngeal cancers.
Cancer. 121:1966–1976. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhang D, Zhang Q, Zhou L, Huo L, Zhang Y,
Shen Z and Zhu Y: Comparison of prevalence, viral load, physical
status and expression of human papillomavirus-16, −18 and −58 in
esophageal and cervical cancer: A case-control study. BMC Cancer.
10:6502010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Flores R, Papenfuss M, Klimecki WT and
Giuliano AR: Cross-sectional analysis of oncogenic HPV viral load
and cervical intraepithelial neoplasia. Int J Cancer.
118:1187–1193. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Saunier M, Monnier-Benoit S, Mauny F,
Dalstein V, Briolat J, Riethmuller D, Kantelip B, Schwarz E, Mougin
C and Prétet JL: Analysis of human papillomavirus type 16 (HPV16)
DNA load and physical state for identification of HPV16-infected
women with high-grade lesions or cervical carcinoma. J Clin
Microbiol. 46:3678–3685. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Chang L, He X, Yu G and Wu Y:
Effectiveness of HPV 16 viral load and the E2/E6 ratio for the
prediction of cervical cancer risk among Chinese women. J Med
Virol. 85:646–654. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Kwiecien R, Kopp-Schneider A and Blettner
M: Concordance analysis: Part 16 of a series on evaluation of
scientific publications. Dtsch Arztebl Int. 108:515–521.
2011.PubMed/NCBI
|
|
46
|
Fernandez AF, Rosales C, Lopez-Nieva P,
Graña O, Ballestar E, Ropero S, Espada J, Melo SA, Lujambio A,
Fraga MF, et al: The dynamic DNA methylomes of double-stranded DNA
viruses associated with human cancer. Genome Res. 19:438–451. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Kalantari M, Osann K, Calleja-Macias IE,
Kim S, Yan B, Jordan S, Chase DM, Tewari KS and Bernard HU:
Methylation of human papillomavirus 16, 18, 31, and 45 L2 and L1
genes and the cellular DAPK gene: Considerations for use as
biomarkers of the progression of cervical neoplasia. Virology.
448:314–321. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Mirabello L, Schiffman M, Ghosh A,
Rodriguez AC, Vasiljevic N, Wentzensen N, Herrero R, Hildesheim A,
Wacholder S, Scibior-Bentkowska D, et al: Elevated methylation of
HPV16 DNA is associated with the development of high grade cervical
intraepithelial neoplasia. Int J Cancer. 132:1412–1422. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Clarke MA, Wentzensen N, Mirabello L,
Ghosh A, Wacholder S, Harari A, Lorincz A, Schiffman M and Burk RD:
Human papillomavirus DNA methylation as a potential biomarker for
cervical cancer. Cancer Epidemiol Biomarkers Prev. 21:2125–2137.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Mirabello L, Frimer M, Harari A, McAndrew
T, Smith B, Chen Z, Wentzensen N, Wacholder S, Castle PE,
Raine-Bennett T, et al: HPV16 methyl-haplotypes determined by a
novel next-generation sequencing method are associated with
cervical precancer. Int J Cancer. 136:E146–E153. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Brandsma JL, Harigopal M, Kiviat NB, Sun
Y, Deng Y, Zelterman D, Lizardi PM, Shabanova VS, Levi A, Yaping T,
et al: Methylation of twelve CpGs in human papillomavirus type 16
(HPV16) as an informative biomarker for the triage of women
positive for HPV16 infection. Cancer Prev Res (Phila). 7:526–533.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Ortiz-Ortiz J, Alarcón-Romero Ldel C,
Jiménez-Lopez MA, Garzón-Barrientos VH, Calleja-Macías I,
Barrera-Saldaña HA, Leyva-Vázquez MA and Illades-Aguiar B:
Association of human papillomavirus 16 E6 variants with cervical
carcinoma and precursor lesions in women from Southern Mexico.
Virol J. 12:292015. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Vinokurova S, Wentzensen N, Kraus I, Klaes
R, Driesch C, Melsheimer P, Kisseljov F, Dürst M, Schneider A and
von Knebel Doeberitz M: Type-dependent integration frequency of
human papillomavirus genomes in cervical lesions. Cancer Res.
68:307–313. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Fujii T, Masumoto N, Saito M, Hirao N,
Niimi S, Mukai M, Ono A, Hayashi S, Kubushiro K, Sakai E, et al:
Comparison between in situ hybridization and real-time PCR
technique as a means of detecting the integrated form of human
papillomavirus 16 in cervical neoplasia. Diagn Mol Pathol.
14:103–108. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Biesaga B, Szostek S, Klimek M, Jakubowicz
J and Wysocka J: Comparison of the sensitivity and specificity of
real-time PCR and in situ hybridization in HPV16 and 18 detection
in archival cervical cancer specimens. Folia Histochem Cytobiol.
50:239–247. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Bryant D, Tristram A, Liloglou T, Hibbitts
S, Fiander A and Powell N: Quantitative measurement of Human
Papillomavirus type 16 L1/L2 DNA methylation correlates with
cervical disease grade. J Clin Virol. 59:24–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Bryan JT, Taddeo F, Skulsky D, Jansen KU,
Frain BM, Qadadri B and Brown DR: Detection of specific human
papillomavirus types in paraffin-embedded sections of cervical
carcinomas. J Med Virol. 78:117–124. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
De Marchi Triglia R, Metze K, Zeferino LC
and De Angelo Andrade Lucci LA: HPV in situ hybridization signal
patterns as a marker for cervical intraepithelial neoplasia
progression. Gynecol Oncol. 112:114–118. 2009. View Article : Google Scholar : PubMed/NCBI
|