Introduction
Angiogenesis, the physiological formation of new
blood vessels from pre-existing ones (1), serves a central role in human physiology
during fetal development, wound healing, tissue repair following
surgery or trauma, menstruation, cancer, and various ischemic and
inflammatory diseases (2). However,
unregulated angiogenesis may result in angiogenic diseases,
including diabetic retinopathy, rheumatoid arthritis, inflammatory
diseases, or tumor growth and metastasis (3,4). As cancer
growth is associated with angiogenesis, the inhibition of
angiogenesis is a promising therapeutic strategy in cancer
treatment. Furthermore, understanding the mechanisms of
angiogenesis inhibition well enough to manipulate it may lead to
numerous therapeutic possibilities.
Avemar (fermented wheat germ extract) is produced by
the industrial fermentation of wheat germ. Avemar is a completely
natural and non-toxic compound that is used clinically as a dietary
supplement for cancer patients undergoing chemotherapy and
radiotherapy (5–9). It is known to have certain biological
effects due its major components, 2-methoxy-benzoquinone and
2,6-dimethoxy-benzoquinone. Additionally, Avemar has been
demonstrated to be associated with anaerobic glycolysis, the
pentose cycle and ribonucleotide reductase enzymes; to exert
significant anti-proliferative effects in a broad spectrum of tumor
cell lines; and to possess the ability to kill tumor cells by
inducing apoptosis through the caspase-poly ADP-ribose polymerase
pathway (5,10). Furthermore, Avemar was reported to be
an effective adjuvant agent in cancer treatment for several types
of cancer. such as breast, colon, lung and prostate cancer
(11). However, the mechanism of the
anti-angiogenic effect of Avemar is unclear. Numerous studies have
investigated cytotoxic effects of Avemar on nearly all types of
cancers that's why we did not do any cell viability in this
project. Therefore, the Avemar concentrations were determined
according to the literature (11–14). The
main aim of the present study was to determine the effects of
Avemar on angiogenesis. Therefore, the present study focused on
molecular target genes associated with angiogenesis, vascular
endothelial growth factor (VEGF) and cyclooxygenase-2 (Cox-2), to
evaluate the anti-angiogenic effect of Avemar on tumor cells.
Materials and methods
Cell lines and Avemar
The human gastric carcinoma cell line NCI-N87, human
prostate cancer cell line PC3, human cervical carcinoma cell line
HeLa, and human lung adenocarcinoma cell line A549 were purchased
from ATCC (Manassas, VA, USA). All cells were grown in Dulbecco's
modified Eagle's medium (Gibco; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA), supplemented with 10% heat-inactivated fetal
bovine serum (Gibco; Thermo Fisher Scientific, Inc.), 2 mM
glutamine, and 1% penicillin/streptomycin/neomycin in a humidified
incubator (5% CO2 in air at 37°C). Avemar was donated by
Biropharma USA Inc. (New York, NY, USA). The Avemar was stored as
dried powder at 4°C in a bottle until use. Prior to use, it was
freshly prepared in sterile water to a final concentration of 400
µg/ml. The solution was centrifuged to remove indissoluble
materials and then filtered with a 0.22-µm filter.
Determination of VEGF and Cox-2 levels
by ELISA
A549, PC3 and NCI-N87 cells release VEGF protein
constitutively. The augmented release of VEGF protein after 48 h
was determined in response to serum starvation in PC3 and NCI-N87
cells, and in response to 1,000 U/ml tumor necrosis factor α
(TNF-α) in A549 cells (15). In our
previous study, the basal VEGF protein levels were determined at
24, 48 and 72 h following seeding of HeLa cells (5×103
cells/well) without any stimuli (16).
A human VEGF ELISA kit (cat. no., ENZ-KIT156; Enzo
Life Sciences, Inc., Farmingdale, NY, USA) was used according to
the manufacturer's protocols in order to determine the possible
effects of Avemar on VEGF levels in tumor cells. Briefly,
5×103 cells were plated in each well of a 96-well plate
and were treated with various concentrations (400, 800, 1,600 or
3,200 µg/ml) of Avemar for 24 or 48 h. Samples (100 µl) were then
added to the microplates containing VEGF-specific monoclonal
antibodies, and the mixtures were incubated for 2 h at room
temperature. The plates were then washed three times to remove any
unbound substances. Enzyme-linked polyclonal antibodies specific
for VEGF were then added to the wells, and the mixtures were
incubated for 2 h at room temperature, prior to a further wash to
remove any unbound antibody or enzyme reagent. The substrate
solution was subsequently added to the wells, and the reaction
resulted in the development of a blue color, the intensity of which
was proportionate to the amount of VEGF bound in the initial step.
Following quenching to cease color development, the intensity of
the color was measured at 450 nm with a Multiskan GO Microplate
Spectrophotometer (Thermo Fisher Scientific, Inc.) and compared to
a standard curve.
The Cox-2 concentration was measured using a human
Cox-2 ELISA kit (cat. no., ADI-900-094) provided by Enzo Life
Sciences, Inc. Samples were prepared by extracting Cox-2 from the
cells and stock solutions were prepared according the
manufacturer's protocol. Briefly, the cells were harvested and
medium was removed. The cells were re-suspended in
radioimmunoprecipitation assay (RIPA) buffer (25 mM Tris, pH 7.4,
0.15 M KCl, 1% NP-40, 5 mM EDTA, 0.5% Sodium deoxycholate, 0.1%
SDS). Samples were prepared by sonicating cells in RIPA buffer for
5 cycles of 30 sec, in 1 min intervals on ice. Samples were then
added the microplate. The plate was incubated at 37°C for 1 h, then
washed prior to the addition of a labeled antibody. The plate was
incubated at 4°C for 30 min, washed, and a substrate solution was
added. The reaction was stopped and absorbance was measured at 450
nm using a Multiskan GO Microplate Spectrophotometer. Protein
concentrations were calculated with reference to the standard
curve.
Total RNA isolation and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
All tumor cell lines were plated in 6-well plates
(3×106 cells/well) and allowed to attach for 24 h.
Following the incubation period, cells were treated with 400 or
3,200 µg/ml Avemar and the cells were incubated for a further 48 h.
Subsequently, the medium in each well was aspirated, and the cells
were washed with ice-cold PBS and immediately lysed in
2-mercaptoethanol in RLT buffer from the RNeasy kit (Qiagen GmbH,
Hilden, Germany). Further RNA isolation was performed with the
RNeasy kit as described by the manufacturer. Total isolated RNA was
then quantified, and 100-ng samples of total RNA were
reverse-transcribed into cDNA using a QuantiTect Reverse
Transcription kit (Qiagen GmbH), according to the manufacturer's
protocol, along with random hexamers and oligo-(dT) 16
primers included in the kit. RT was performed at 42°C for 30 min.
The total volume for each reverse transcription reaction was 20 µl.
The generated cDNA was then used as the template for qPCR in a
StepOnePlus™ Real-Time PCR System (Thermo Fisher Scientific, Inc.,
Waltham, MA, USA). The qPCR was performed under the following
conditions: 5 min initial denaturation at 94°C, then 35 cycles of
denaturation (30 sec at 94°C), annealing (45 sec at 58°C),
extension (45 sec at 72°C), and a final extension at 72°C for 5
min.
The reaction mixtures consisted of SYBR Green I
(QuantiTect SYBR Green PCR kit; Qiagen GmbH), 300 µM forward and
reverse primers (final concentrations optimized during the assay
setup), 100 nM human UPL probe, and 2.5 µl cDNA template. The
sequences of the primer pairs are listed in Table I. The expression levels of the tested
genes were normalized to that of GAPDH and presented as the fold
change compared with the control group (17). All experiments were repeated three
times.
 | Table I.Primer pairs used for reverse
transcription-quantitative polymerase chain reaction. |
Table I.
Primer pairs used for reverse
transcription-quantitative polymerase chain reaction.
|
| Primer sequences
(5′→3′) |
|---|
|
|
|
|---|
| Gene name | Forward | Reverse |
|---|
| Cox-2 |
TCACGCATCAGTTTTTCAAGA |
TCACCGTAAATATGATTTAAGTCCAC |
| VEGF |
AGGCCAGCACATAGGAGAGA |
TTTCCCTTTCCTCGAACTGA |
| GAPDH |
TCACCAGGGCTGCTTTTAACCT |
CACGCCACAGTTTCCCGGAG |
Statistical analysis
All values are presented as the mean ± standard
error of mean. Data were analyzed using a one-way analysis of
variance (ANOVA) followed by Dunnett's multiple comparisons test.
Analyses were performed with GraphPad InStat v.10.0 software
(GraphPad Software, Inc., La Jolla, CA, USA). P<0.05 was
considered statistically significant.
Results
VEGF is affected by Avemar in tumor
cell lines
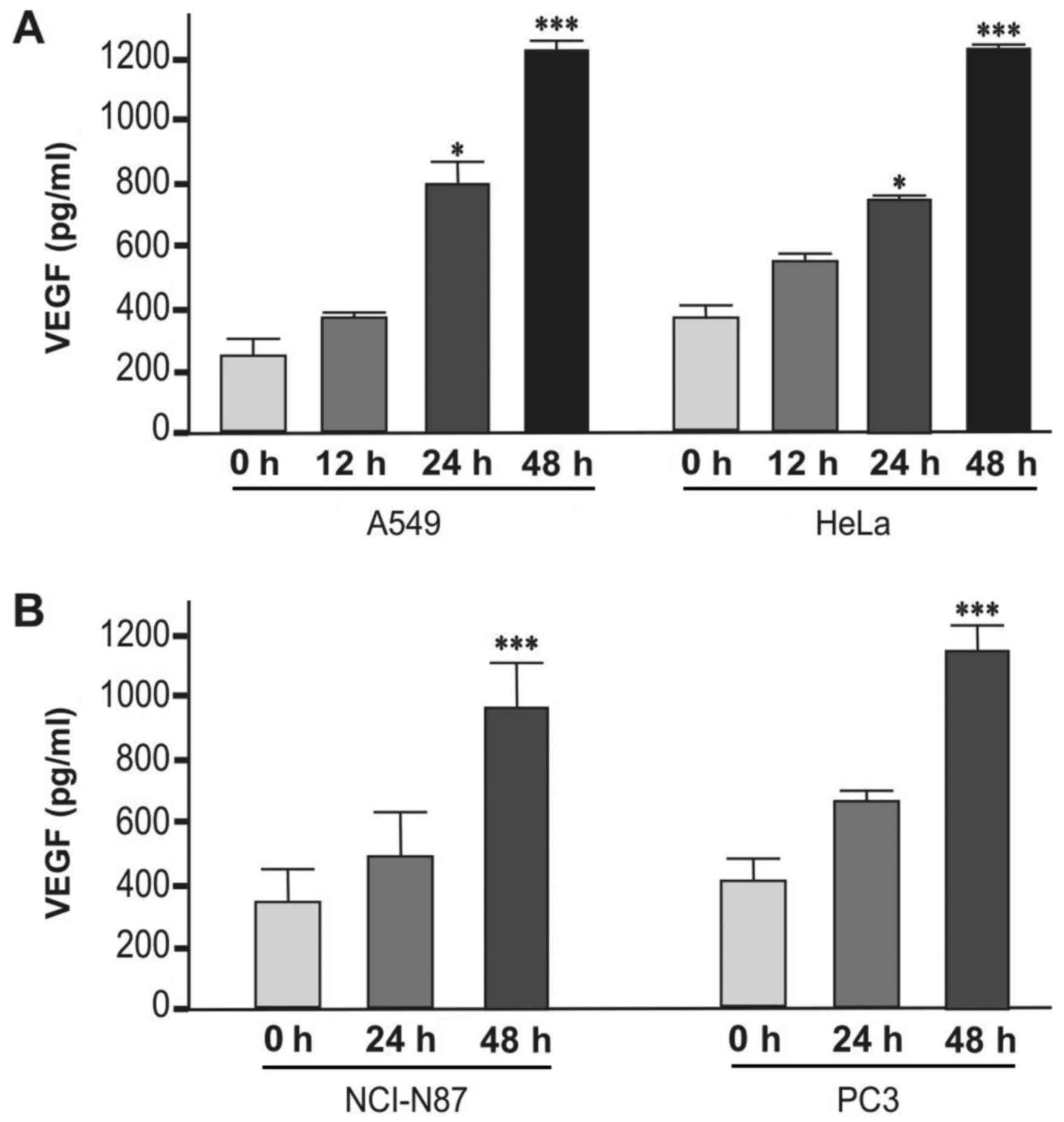
First, the maximum level of VEGF expression was
determined in A549 cells stimulated by TNF-α and in PC3 and NCI-N87
cells stimulated by serum starvation. Basal VEGF protein levels
were measured at 12, 24 and 48 h in HeLa cells, and the maximum
induction of released VEGF protein was observed after 48 h without
any stimuli (Fig. 1).
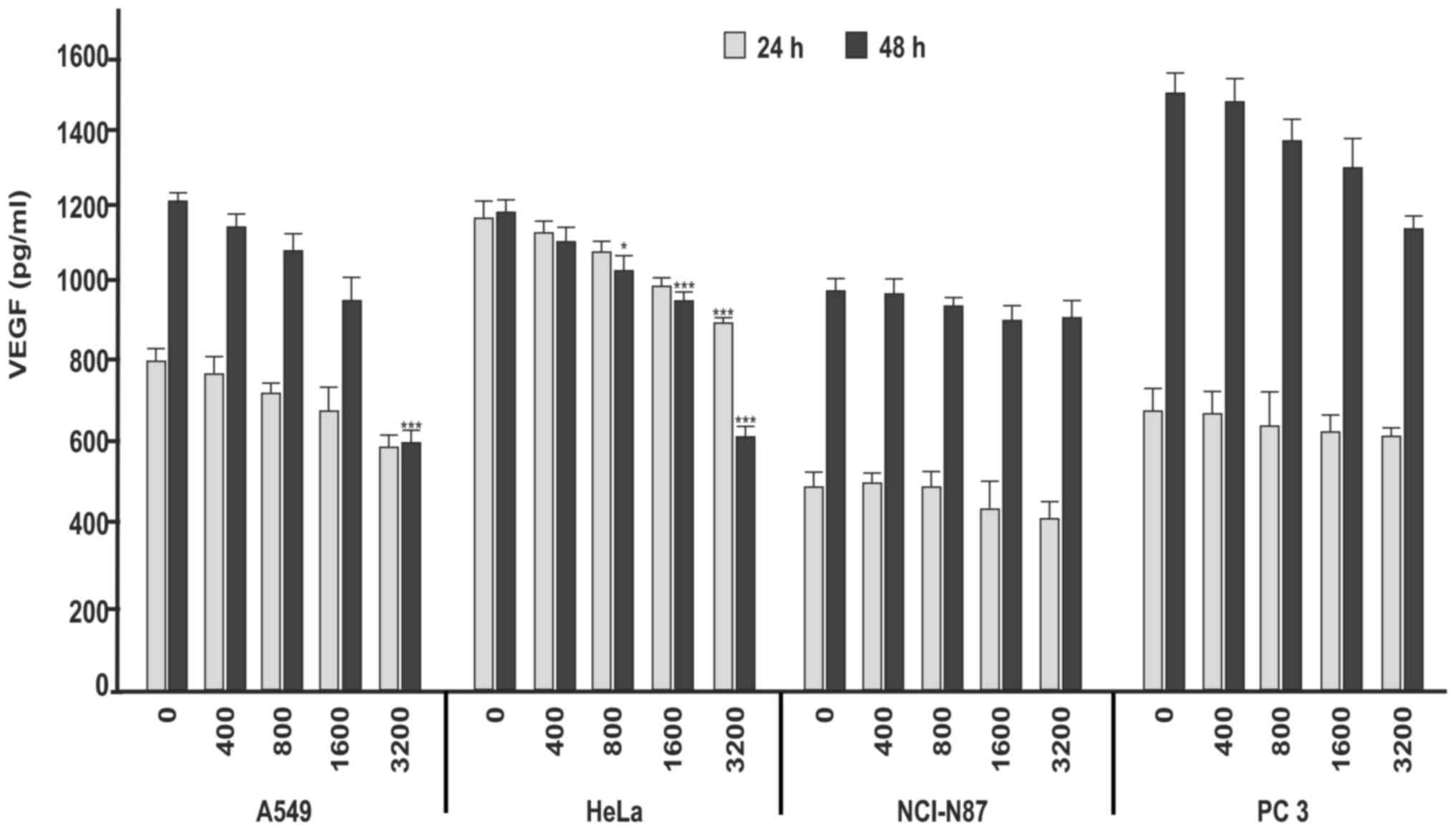
To determine the possible effects of Avemar on VEGF
levels induced by the different stimuli for each cell line, all
tumor cells were treated with 400, 800, 1,600 and 3,200 µg/ml
Avemar for 24 and 48 h. In all cell lines, the inhibition of
induced VEGF levels increased in a dose-dependent manner (Fig. 2). The increase in inhibition, as
compared with the control group, was statistically significant in
the HeLa and A549 cells that were treated with 3,200 µg/ml Avemar
for 48 h (P<0.001). No significant changes in VEGF inhibition
were observed in the NCI-N87 and PC3 cells at either time point
(P>0.05). In A549 and HeLa cells, RT-qPCR was performed to
investigate whether the inhibition of VEGF levels occurred due to
alterations at the transcript level. The results revealed
consistent alterations in the mRNA (Fig.
3) and protein levels of VEGF (P<0.001). As shown in
Fig. 3, Avemar treatment (3,200
µg/ml) yielded 3.2- and 3.4-fold decreases in VEGF mRNA levels in
A549 and HeLa cells, respectively (P<0.001).
Cox-2 may be responsible for the
inhibition by Avemar of VEGF induction
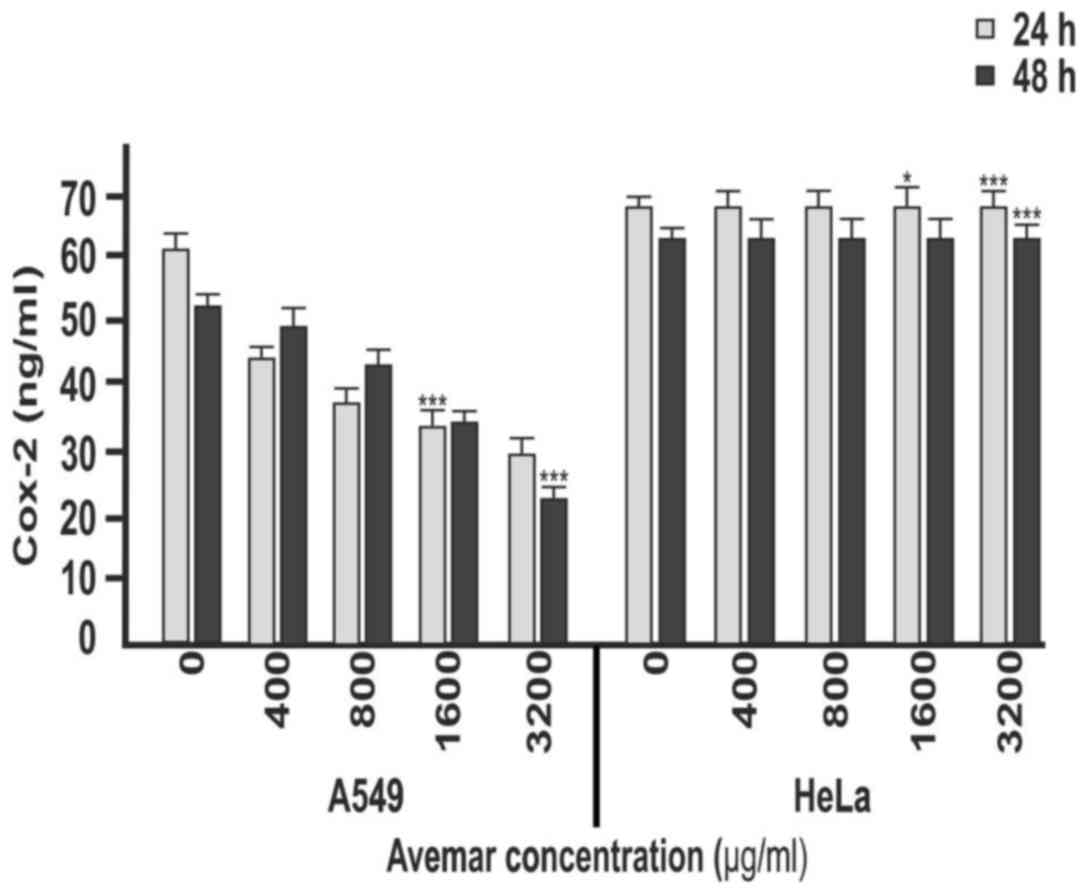
It was next assessed whether Cox-2 was responsible
for the VEGF inhibition following Avemar treatment in HeLa and A549
cells. Unstimulated HeLa cells and TNF-α-stimulated A549 cells were
treated with 400, 800, 1,600 and 3,200 µg/ml Avemar for 24 and 48
h, and Cox-2 protein levels were measured by ELISA. The inhibition
of Cox-2 levels following Avemar treatment in HeLa and A549 cells
increased in a time- and dose-dependent manner (Fig. 4). Compared with the control group, the
amount of Cox-2 protein decreased by ~60% in HeLa and A549 cells
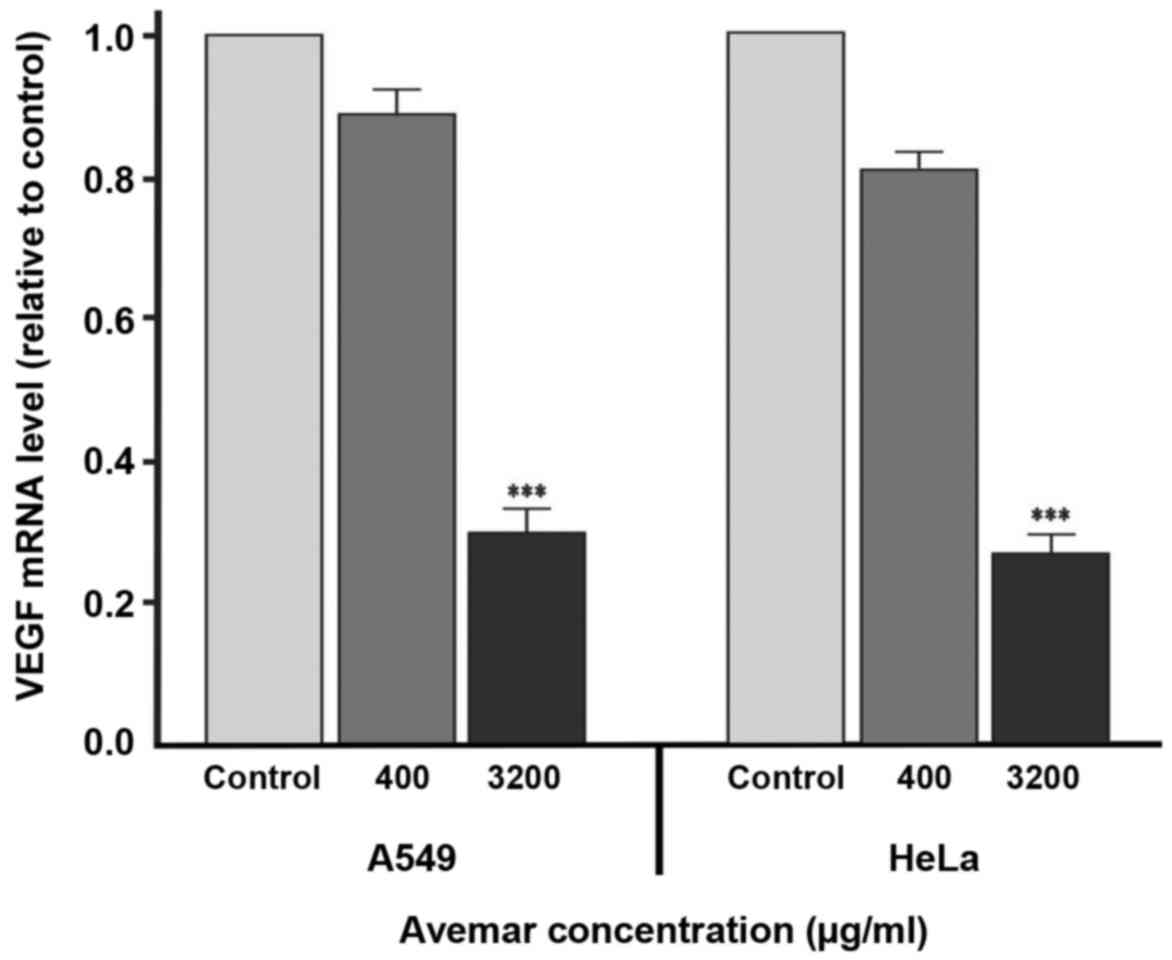
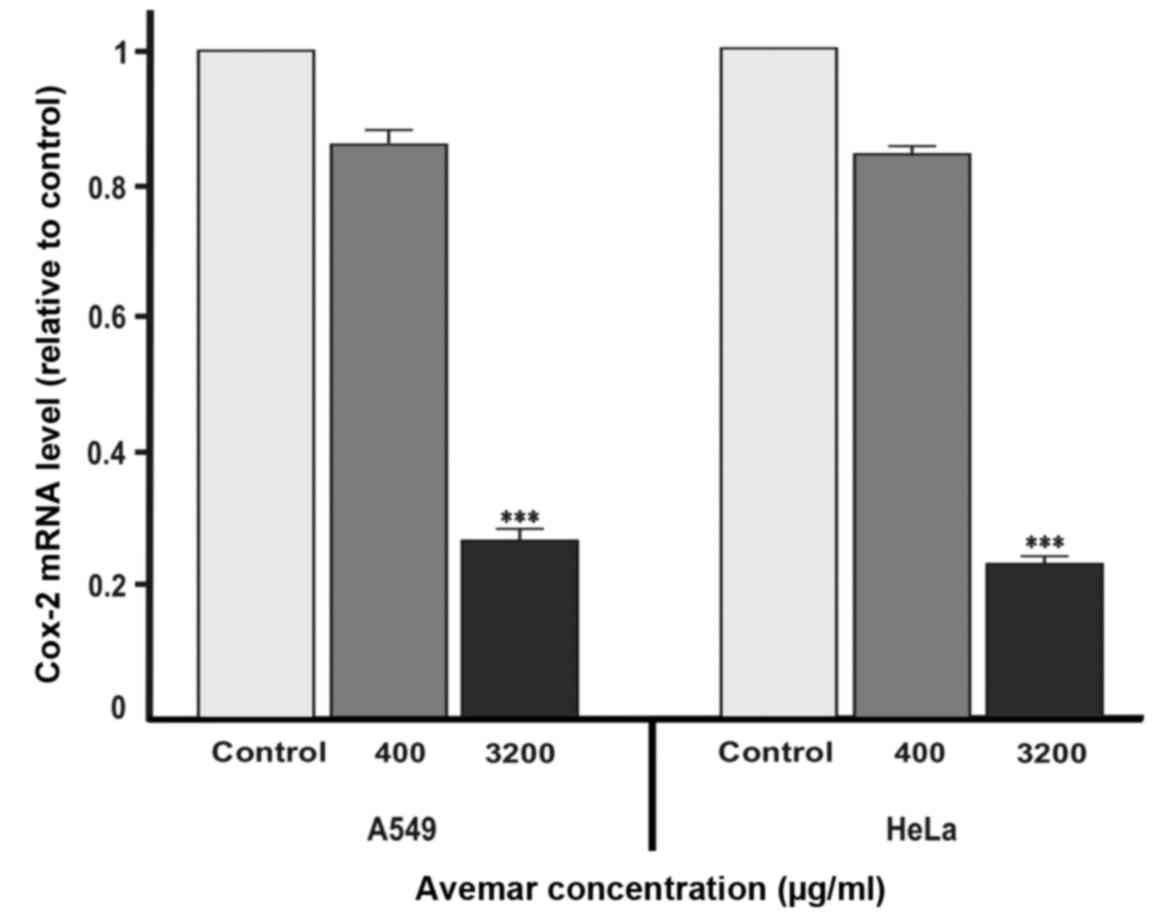
(P<0.001). RT-qPCR was also performed to examine whether Avemar
treatment was also associated with the inhibition of Cox-2 at the
transcript level. RT-qPCR verified the ELISA results for Cox-2
levels, with Avemar treatment (3,200 µg/ml) yielding 4.3- and
4.6-fold decreases in Cox-2 mRNA levels in A549 and HeLa cells,
respectively (Fig. 5).
Discussion
Avemar has been approved as a medical dietary
supplement for cancer patients by Hungary's National Institute of
Food Safety and Nutrition. It has been demonstrated to
synergistically increase the effects of anticancer agents such as
5-fluorouracil (5-FU) and dacarbazine (18), and has also been shown to reduce the
formation of metastases in certain animal models and to increase
survival rates by inhibiting the proliferation of skin grafts
(19,20). In addition, the oral use of Avemar in
patients with advanced colorectal cancer or high-risk melanoma
inhibits the formation of tumor metastases and prolongs survival
following chemotherapy and surgery (13,14). The
present study showed that Avemar exerts anti-angiogenic effects by
inhibiting VEGF and Cox-2 gene expression in PC3, NCI-N87, HeLa and
A549 cells. These findings highlight the anti-angiogenic effect of
Avemar on cancer cell lines, and, to the best of our knowledge,
this is the first report confirming this possible anti-angiogenic
mechanism.
Angiogenesis is an important feature of malignant
tumors. Newly formed tumor blood vessels provide the necessary
nutrition for tumor growth as well as a means to spread to other
organs. VEGF, the most widely accepted stimulator of angiogenesis,
promotes the formation of new vessels by binding to VEGF receptors
on the endothelial cells surrounding the blood vessels (21). In recent years, drugs that can block
the VEGF pathway have become important components of cancer
treatment. Another key regulator of angiogenesis is Cox-2, and
there is a close association between the Cox-2 and VEGF pathways in
the regulation of angiogenesis (22).
Stimulating Cox-2 induces the activation and production of VEGF
(23). Downregulation of the Cox-2
and VEGF pathways has been associated with the anti-angiogenic
activity of polyphenols and polyphenol-rich foods in in
vitro and in vivo models of angiogenesis (24–26).
Therefore, it was hypothesized that Avemar, which includes a number
of components, could reduce angiogenic responses in tumor cells and
that this reduction may be mediated by decreases in Cox-2 and VEGF
expression.
Tsujii et al (27) demonstrated that the overexpression of
Cox-2 may lead to dedifferentiation, adhesion to the extracellular
matrix and inhibition of programmed cell death in intestinal cells.
Although the precise mechanisms by which Cox-2 promotes tumor cell
growth remain to be fully elucidated, it has been demonstrated that
Cox-2 stimulates endothelial cell motility and tube formation in
Caco-2 human colorectal cancer and HCA-7 human colonic
adenocarcinoma cells by upregulating the production of
pro-angiogenic factors, including VEGF. In a study performed by
Sawaoka et al (28), which
utilized an MKN45 xenograft model, it was demonstrated that the
inhibition of Cox-2 by NS-398 (a selective Cox-2 inhibitor)
suppressed the protein levels of VEGF and basic fibroblast growth
factor, and that the angiogenic and apoptotic indexes were
significantly associated. Furthermore, in a preclinical study by
Mueller et al (11), the
cytotoxic activity of Avemar was screened in a large panel of human
tumor cell lines to evaluate its potential antitumor properties.
The highest activity was found in neuroblastoma and ovarian cancer
cell lines. The half-maximal inhibitory concentrations of Avemar
were 0.5 mg/ml in M2 (a gastric cancer cell line) and 0.4 mg/ml in
A549 (a lung adenocarcinoma cell line). According to this, the
results of the current study are in agreement with previous studies
of Avemar that have demonstrated antitumor activity. We conclude
that Avemar may exert anti-angiogenic activity by reducing VEGF and
Cox-2 in tumor cells. However, further studies on how the products
of Cox-2 mediate the transcriptional regulation of VEGF are
required.
There are several limitations to the present study.
First, the protein and gene levels of VEGF and Cox-2 were the only
direct markers used in the assessment of angiogenesis. The extent
and activity of other genes related to angiogenesis, such as matrix
metalloproteinases (MMPs) and interleukins, requires further
evaluation to better assess angiogenesis. The roles of MMP-2 and
MMP-9 in angiogenesis have been explored in a variety of studies
(29–31). Recently, Yang et al (32) demonstrated that Avemar significantly
decreased the expression of MMP-2, but not MMP-1 or MMP-9, in SCC-4
oral cancer cells. In addition, the authors assayed the expression
levels of urokinase-type plasminogen activator (uPA), an upstream
activator of MMPs, which are typically associated with metastasis.
The results of RT-PCR indicated that Avemar inhibited MMP-2 and uPA
due to altering the transcript levels of these genes (31).
Future investigations are warranted on the
combination of Avemar with other anticancer drugs, such as 5-FU and
cisplatin, which may enhance its in vivo and in vitro
anti-angiogenic and anti-proliferative activities.
In conclusion, the present study indicates that
Avemar diminishes angiogenic activity in various tumor cell lines.
This is mediated through decreased Cox-2 and VEGF levels. To the
best of our knowledge, this is the first report examining the
mechanism of the anti-angiogenic effect of Avemar on gastric,
prostatic, cervical and lung cancer cells. We hope that this study
will encourage further studies into the anti-angiogenic effects of
Avemar alone and in combination with other anticancer agents.
Acknowledgements
The present study was supported by the Scientific
Research Project Coordination Unit of Akdeniz University (grant no.
2014.01.0110.001). All experiments were carried out in the Cancer
Molecular Biology Laboratory of the Biology Department of the
Faculty of Science, Akdeniz University. The authors wish to thank
Biropharma USA Inc. for donating the Avemar used in the study.
References
|
1
|
Folkman J: Angiogenesis in cancer,
vascular, rheumatoid and other disease. Nat Med. 1:27–31. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Adams RH and Alitalo K: Molecular
regulation of angiogenesis and lymphangiogenesis. Nat Rev Mol Cell
Biol. 8:464–478. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bar J and Onn A: Combined
anti-proliferative and anti-angiogenic strategies for cancer.
Expert Opin Pharmacother. 9:701–715. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cao Y: Angiogenesis: What can it offer for
future medicine? Exp Cell Res. 316:1304–1308. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mueller T and Voigt W: Fermented wheat
germ extract-nutrional supplement or anticancer drug? Nutrition.
10:892011. View Article : Google Scholar
|
|
6
|
Saiko P, Ozsvar-Kozma M, Madlener S,
Bernhaus A, Lackner A, Grusch M, Horvath Z, Krupitza G, Jaeger W,
Ammer K, et al: Avemar, a nontoxic fermented wheat germ extract,
induces apoptosis and inhibits ribonucleotide reductase in human
HL-60 promyelocytic leukemia cells. Cancer Lett. 250:323–328. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hidvégi M, Ráso E, Tömösközi-Farkas R,
Paku S, Lapis K and Szende B: Effect of Avemar and Avemar C vitamin
C on tumor growth and metastasis in experimental animals.
Anticancer Res. 18:2353–2358. 1998.PubMed/NCBI
|
|
8
|
Demidov LV, Manziuk LV, Kharkevitch GY,
Pirogova NA and Artamonova EV: Adjuvant fermented wheat germ
extract (Avemar) nutraceutical improves survival of high-risk skin
melanoma patients: A randomized, pilot, phase II clinical study
with a 7-year follow-up. Cancer Biother Radiopharm. 23:477–482.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Heimbach JT, Sebestyen G, Semjen G and
Kennepohl E: Safety studies regarding a standardized extract of
fermented wheat germ. Int J Toxicol. 26:253–259. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Comin-Anduix B, Boros LG, Marin S, Boren
J, Callol-Masot C, Centelles JJ, Torres JL, Angell N, Bassilian S
and Cascante M: Fermented wheat germ extract inhibits
glycolysis/pentose cycle enzymes and induces apoptosis through
Poly(ADP-riboz) polymerase activation in Jurkat T-cell leukemia
tumor cells. J Biol Chem. 277:46408–46414. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mueller T, Jordan K and Voigt W: Promising
cytotoxic profile of fermented wheat germ extract
(Avemar®) in human cancer cell lines. J Exp Clin Cancer
Res. 30:422011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Garami M, Schuler D, Babosa M, Borgulya G,
Hauser P, Müller J, Paksy A, Szabó E, Hidvégi M and Fekete G:
Fermented wheat germ extract reduces chemotherapy-induced febrile
neutropenia in pediatric cancer patients. J Pediatr Hematol Oncol.
26:631–635. 2004. View Article : Google Scholar
|
|
13
|
Jakab F, Shoenfeld Y, Balogh A, Nichelatti
M, Hoffmann A, Kahán Z, Lapis K, Mayer A, Sápy P, Szentpétery F, et
al: A medical nutriment has supportive value in the treatment of
colorectal cancer. Br J Cancer. 89:465–469. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang JY, Xiao X, Dong Y, Wu J, Yao F and
Zhou XH: Effect of fermented wheat germ extract with lactobacillus
plantarum dy-1 on HT-29 cell proliferation and apoptosis. J Agric
Food Chem. 63:2449–2457. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Koyama S, Sato E, Tsukadaira A, Haniuda M,
Numanami H, Kurai M, Nagai S and Izumi T: Vascular endothelial
growth factor mRNA and protein expression in airway epithelial cell
lines in vitro. Eur Respir J. 20:1449–1456. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Şimşek E, Aydemir EA, İmir N, Koçak O,
Kuruoğlu A and Fışkın K: Dimethyl sulfoxide-caused changes in pro-
and anti-angiogenic factor levels could contribute to an
anti-angiogenic response in HeLa cells. Neuropeptides. 53:37–43.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-delta delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hidvégi M, Rásό E, Tömösközi-Farkas R,
Lapis K and Szende B: Effect of MSC on the immune response of mice.
Immunopharmacology. 41:183–186. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Szende B, Rásό E, Hidvégi M, Tömösköziné
FR, Paku S, Prónai L, Bocsi J and Lapis K: A new
benzoquinone-containing antimetastatic product. Orv Hetil.
139:2893–2897. 1998.(In Hungarian). PubMed/NCBI
|
|
20
|
Hidvégi M, Rásό E, Tömösközi-Farkas R,
Szende B, Paku S, Prónai L, Bocsi J and Lapis K: MSC, a new
benzoquinone-containing natural product with antimetastatic effect.
Cancer Biother Radiopharm. 14:277–289. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kerbel RS: Tumor angiogenesis. N Engl J
Med. 358:2039–2049. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Taleb H, Morris RK, Withycombe CE,
Maddocks SE and Kanekanian AD: Date syrup derived polyphenols
attenuate angiogenic responses and exhibits anti-inflammatory
activity mediated by VEGF and COX-2 expression in endothelial
cells. Nutr Res. 36:636–647. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Costa C, Incio J and Soares R:
Angiogenesis and chronic inflammation: Cause or consequence?
Angiogenesis. 10:149–166. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Scoditti E, Calabriso N, Massaro M,
Pellegrino M, Storelli C, Martines G, De Caterina R and Carluccio
MA: Mediterranean diet polyphenols reduce inflammatory angiogenesis
through MMP-9 and Cox-2 inhibition in human vascular endothelial
cells: A potentially protective mechanism in atherosclerotic
vascular disease and cancer. Arch Biochem Biophys. 527:81–89. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Corona G, Deiana M, Incani A, Vauzour D,
Dessí MA and Spencer JP: Inhibition of p38/CREB phosphorylation and
Cox-2 expression by olive oil polyphenols underlies their
anti-proliferative effects. Biochem Biophys Res Commun.
362:606–611. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Medda R, Lyros O, Schmidt JL, Jovanovic N,
Nie L, Link BJ, Otterson MF, Stoner GD, Shaker R and Rafiee P:
Anti-inflammatory and anti-angiogenic effect of black raspberry
extract on human esophageal and intestinal microvascular
endothelial cells. Microvasc Res. 97:167–180. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tsujii M, Kawano S and DuBois RN:
Cyclooxygenase-2 expression in human colon cancer cells increases
metastatic potential. Proc Natl Acad Sci USA. 94:3336–3340. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sawaoka H, Tsuji S, Tsujii M, Gunawan ES,
Sasaki Y, Kawano S and Hori M: Cyclooxygenase inhibitors suppress
angiogenesis and reduce tumor growth in vivo. Lab Invest.
79:1469–1477. 1999.PubMed/NCBI
|
|
29
|
Braunhut SJ and Moses MA: Retinoids
modulate endothelial cell production of matrix-degrading proteases
and tissue inhibitors of metalloproteinases (TIMP). J Biol Chem.
269:13472–13479. 1994.PubMed/NCBI
|
|
30
|
Vacca A, Moretti S, Ribatti D, Pellegrino
A, Pimpinelli N, Bianchi B, Bonifazi E, Ria R, Serio G and Dammacco
F: Progression of mycosis fungoides is associated with changes in
angiogenesis and expression of the matrix metalloproteinases 2 and
9. Eur J Cancer. 33:1685–1692. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Itoh T, Tanioka M, Yoshida H, Yoshioka T,
Nishimoto H and Itohara S: Reduced angiogenesis and tumor
progression in gelatinase A-deficient mice. Cancer Res.
58:1048–1051. 1998.PubMed/NCBI
|
|
32
|
Yang MD, Chang WS, Tsaia CW, Wang MF, Chan
YC, Chan KC, Lu MC, Kao AW, Hsu CM and Bau DT: Inhibitory effects
of AVEMAR on proliferation and metastasis of oral cancer cells.
Nutr Cancer. 68:473–480. 2016. View Article : Google Scholar : PubMed/NCBI
|