Introduction
Ewing sarcoma (ES) is the second most common sarcoma
of bone in children and young adults (1). It is an aggressive and highly metastatic
tumor. In total, ~1/3 of patients with ES present with metastasis
at diagnosis, with lung tissue and bone marrow being the most
common sites of metastasis, resulting in poor prognosis (2). Treatment and prognosis of patients with
ES are determined by the presence of metastases, among other
factors. The 5-year survival rate of patients with metastases
ranges between 20 and 45%, depending on location, compared with
between 60 and 70% in those with localized disease (2). Thus, novel therapeutic targets,
innovative approaches to therapy and improved understanding of the
metastatic mechanism are necessary to improve the outcome for
patients with metastatic ES.
Autophagy is a highly conserved process that
contributes to maintaining cellular homeostasis via quality control
of proteins and organelles. Under conditions of metabolic stress,
autophagy provides nutrients and energy essential for cell survival
(3–5).
Beclin-1 is a B-cell lymphoma 2 (Bcl-2) homology 3 domain-only
protein that is required for the formation of autophagosomes, which
are utilized in the initiation of autophagy (6–8). It has
been reported that cell autophagy is associated with tumor
initiation and progression, and serves a function in cell signal
regulation in tumors (9–11). To date, the exact effects of autophagy
on the biological behavior of ES cells have not been fully
resolved.
In the present study, SK-ES-1 cells were transfected
with small interfering (si)RNA against Beclin-1 in order to
investigate the effects of Beclin-1 knockdown on cell
proliferation, invasion and migration, and to determine the
underlying molecular mechanisms. To the best of our knowledge, this
is the first study to report the effects of Beclin-1 knockdown on
the behavior of ES cells.
Materials and methods
Materials and reagents
The SK-ES-1 and RD-ES human ES cell lines were
obtained from the American Type Culture Collection (ATCC; Manassas,
VA, USA). RPMI-1640 medium, fetal bovine serum (FBS), PBS, dimethyl
sulfoxide and Cell Counting Kit-8 (CCK-8) were provided by Beijing
Transgen Biotech Co., Ltd. (Beijing, China). Antibodies against
Beclin-1 (ab207612, 1:1,000), matrix metalloproteinase (MMP)-2
(ab92536, 1:1,000), MMP-9 (ab194314, 1:1,000) and β-actin (ab8227,
1:1,000) were all purchased from Abcam (Cambridge, UK). Goat
anti-rabbit IgG (H+L), horseradish peroxidase-conjugated secondary
antibodies (HS101-01, 1:2,000) were both purchased from Beijing
Transgen Biotech Co., Ltd. Lipofectamine® 2000 and
OPTI-MEM were both purchased from Invitrogen (Thermo Fisher
Scientific, Inc., Waltham, MA, USA). Matrigel was purchased from BD
Biosciences (San Jose, CA, USA). The Transwell invasion chambers
were purchased from Costar (Cambridge, MA, USA). Crystal violet
staining solution was purchased from Beyotime Institute of
Biotechnology (Haimen, China). si-Beclin-1 against the
BECLIN-1 gene (NM_003766) and control siRNA (si-CON) were
constructed by Shanghai GeneChem Co., Ltd. (Shanghai, China).
Cell culture and transfection
SK-ES-1 and RD-ES cells were cultured in RPMI-1640
medium supplemented with 10% (v/v) FBS, 100 U/ml penicillin and 100
µg/ml streptomycin. Cells were incubated in a humidified atmosphere
containing 5% CO2 at 37°C. All cells used in the present
study were subjected to <20 cell passages. SK-ES-1 cells at
logarithmic phase were seeded at a density of 3×105
cells/well in a 6-well plate for 24 h prior to transfection.
Lipofectamine 2000 (10 µl diluted in 250 µl OPTI-MEM) was used for
the transfection of 4 µg si-Beclin-1 or empty vector diluted in 250
µl OPTI-MEM, followed by incubation of the samples for 20 min at
room temperature. The plasmid DNA-Lipofectamine 2000 complex was
then added into 500 µl OPTI-MEM and incubated at 37°C with 5%
CO2 in an incubator for 6 h. Subsequently, the medium
was replaced and the cells were incubated for 24 or 48 h in
RPMI-1640 medium supplemented with 10% FBS prior to use in the
corresponding experiments, which included a blank control group
(non-transfected SK-ES-1 cells), a negative control group (SK-ES-1
cells transfected with blank plasmid, i.e., si-CON) and an
experimental group (SK-ES-1 cells transfected with si-Beclin-1).
Transfected cells were collected at 24 or 48 h post-transfection
and used in subsequent experiments.
Cell proliferation assay
Cell growth was determined using the CCK-8 assay. In
brief, cells infected with si-Beclin-1 or si-CON and
non-transfected SK-ES-1 cells were incubated in 96-well plates at a
density of 3×103 cells/well. Cells were treated with 10
µl CCK-8 reagent at 24, 48 and 72 h and then measured at 450 nm
using a Universal Microplate reader (EL800; Bio-Tek Instruments
Inc., Winooski, VT, USA).
Boyden chamber Transwell assays
The invasive capacity of SK-ES-1 cells was detected
via Matrigel-coated Transwell cell culture chambers (8 µm pore
size). Following transfection for 24 h, SK-ES-1 cells of the three
different groups were collected and suspended in serum-free medium.
Isolated cells were then added to the upper chamber of the
Transwell insert at a density of 4×104 cells/well and
the lower wells were filled with complete growth medium
supplemented with 10% FBS. All samples were incubated for 24 h in a
CO2 incubator. Non-invading cells (on the upper membrane
surface) were removed using a cotton swab and invading cells (on
the lower membrane surface) were fixed with 95% ethanol for 15 min
at 25°C, stained with 0.1% crystal violet staining solution for 20
min at 25°C, then counted under a phase-contrast microscope in
three random fields (magnification, ×200).
Wound healing assays
Migration of SK-ES-1 cells was measured using wound
healing assays. After 24 h of transfection, SK-ES-1 cells of the
three different groups were seeded at a density of 5×105
cells/well in a 6-well culture plate to form a confluent monolayer.
Cells were wounded with a sterile 100 µl pipette tip. All cells in
the plates were incubated in fresh RPMI-1640 medium with 10% FBS
for 24 h. Then scratch wounds were observed using a phase-contrast
microscope and images were captured of each wound.
Western blot analysis
Following transfection, SK-ES-1 cells of the three
different groups were seeded in 6-well plates at a concentration of
3×105 cells/well and incubated in RPMI-1640 medium with
10% FBS for 48 h. The cells were collected and lysed in
radioimmunoprecipitation assay buffer containing phenylmethane
sulfonyl fluoride and phosphatase inhibitor cocktail (Sigma
Aldrich; Merck KGaA, Darmstadt, Germany). Each sample was
centrifuged at 17,105.6 × g for 10 min at 4°C using a Universal
320R centrifuge (Andreas Hettich GmbH & Co. KG, Tuttlingen,
Germany), to remove cell debris and collect the supernatant for
immunoblotting. Protein concentrations were calculated using a
bicinchoninic acid assay kit (Beijing Transgen Biotech Co., Ltd.)
according to the manufacturer's instructions with bovine serum
albumin as the relative standard. Proteins (10 µl) were loaded and
separated using SDS-PAGE (10% gel, 100 V for 2 h under reducing
conditions). Following electrophoresis, the proteins were
transferred to polyvinylidene fluoride (PVDF) membranes in a
tris-glycine transfer buffer and incubated with antibodies against
β-actin, Beclin-1, MMP-2 and MMP-9 overnight at 4°C. The PVDF
membranes were washed in Tris-buffered saline Tween-20 (TBST) three
times. Secondary HRP-conjugated antibodies were added at 1:2,000
dilution and incubated for 2 h at 25°C. The PVDF membranes were
washed a further three times in TBST. Immunoreactive proteins were
detected using an enhanced chemiluminescence system (GE Healthcare,
Chicago, IL, USA) according to the manufacturer's instructions
followed by exposure to X-ray films. Western blotting data was
quantified using ImageJ software (version 7.0; National Institutes
of Health, Bethesda, MD, USA).
Statistical analysis
Data were analyzed using the SPSS package for
Windows (version 19.0; IBM Corp., Armonk, NY, USA). Quantitative
data are expressed as mean ± standard deviation. Statistical
analysis was performed using a one-way analysis of variance with
the Student-Newman-Keuls method as a post hoc test. P<0.05 was
considered to indicate a statistically significant difference.
Results
Endogenous expression of Beclin-1 in
the human ES cell lines
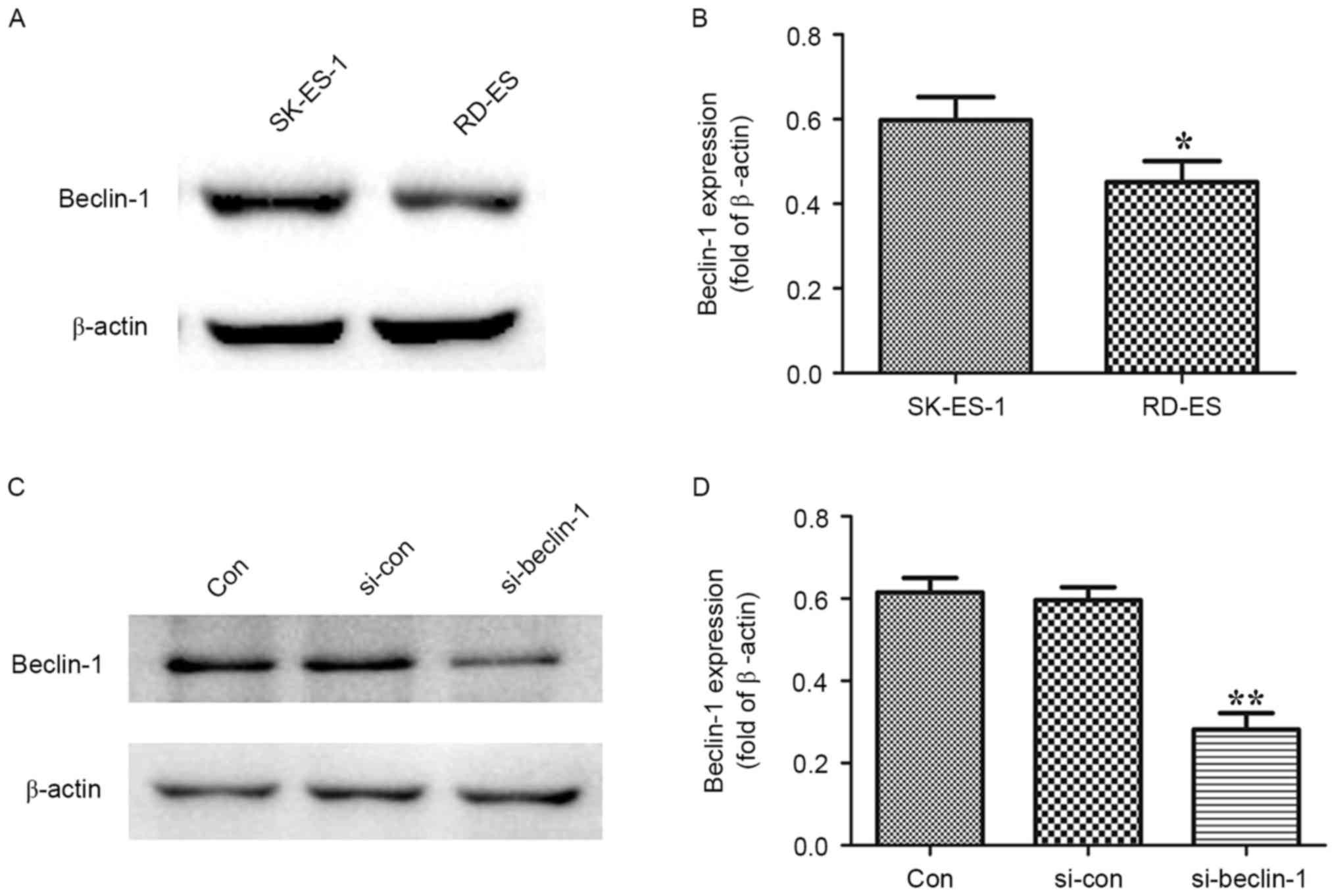
Endogenous expression of Beclin-1 in the human ES
SK-ES-1 and RD-ES cell lines was evaluated using western blot
analysis. As presented in Fig. 1A and
B, expression of Beclin-1 was significantly increased in the
SK-ES-1 cell line compared with the RD-ES cell line (P<0.05).
Thus, the SK-ES-1 cell line was used for Beclin-1 knockdown.
si-Beclin-1 significantly decreases
the expression of Beclin-1 in SK-ES-1 cells
Protein levels of Beclin-1 were determined using
western blotting following transfection of SK-ES-1 cells with
si-Beclin-1 or si-CON vectors for 48 h. As presented in Fig. 1C and D, Beclin-1 expression was
significantly decreased in the si-Beclin-1 group compared with the
blank control group (P<0.01)
Beclin-1 knockdown inhibits the
proliferation of SK-ES-1 cells
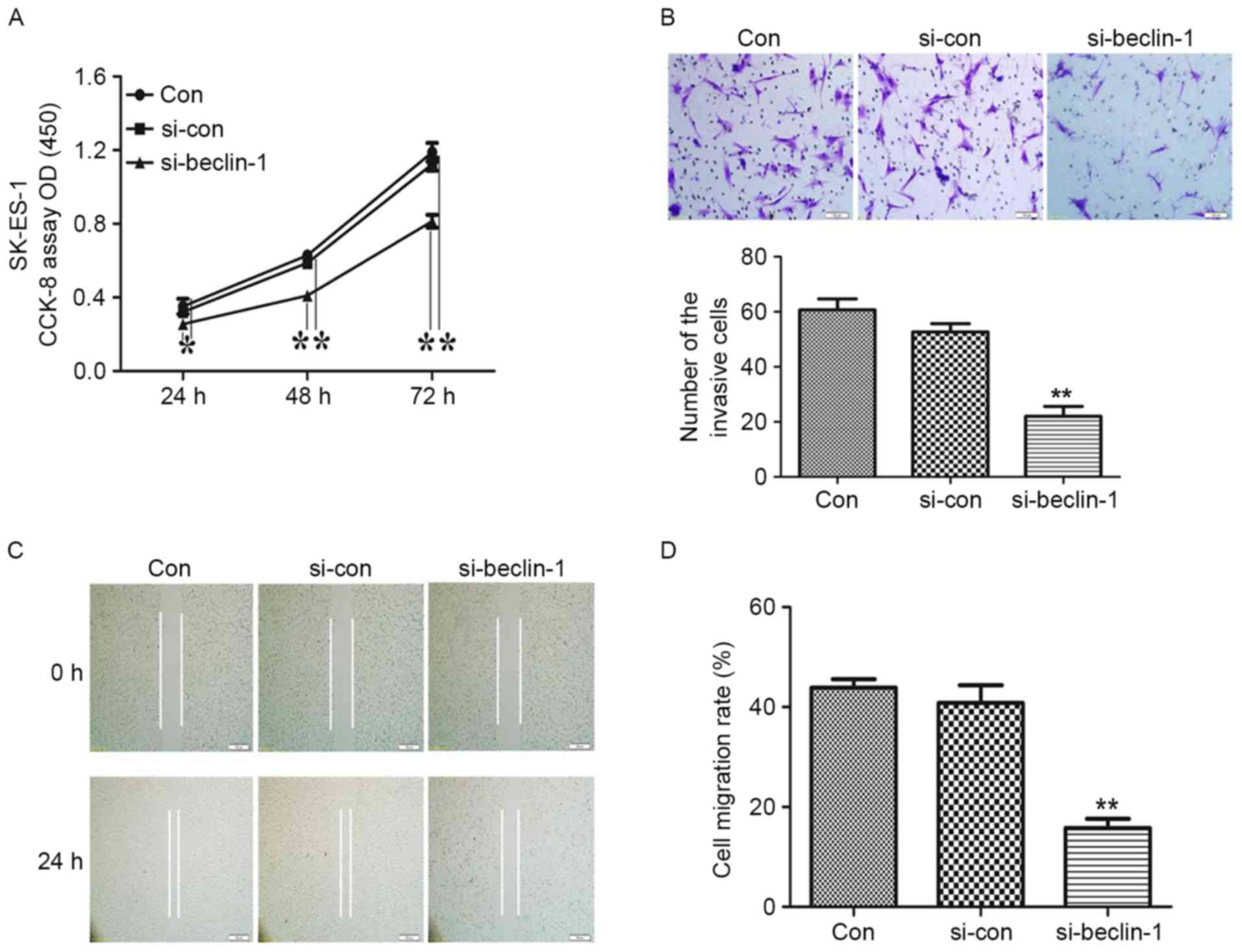
The effect of Beclin-1 knockdown on SK-ES-1 cell
growth was determined using a CCK-8 assay. As presented in Fig. 2A, knockdown of Beclin-1 significantly
suppressed the growth of SK-ES-1 cells (P<0.05 at 24 h and
P<0.01 at 48 h).
Beclin-1 knockdown represses the
invasion and migration of SK-ES-1 cells
Transwell and wound healing assays were conducted to
confirm the effect of Beclin-1 knockdown on the invasion and
migration of SK-ES-1 cells. Representative micrographs of Transwell
filters are presented in Fig. 2B. The
invasive cell count, also presented in Fig. 2B, demonstrated that the invasive
potential was significantly decreased in the si-Beclin-1 group
relative to the blank control group (P<0.01). Furthermore,
Beclin-1 knockdown resulted in a decrease in migration capability,
as presented in Fig. 2C and D
(P<0.01).
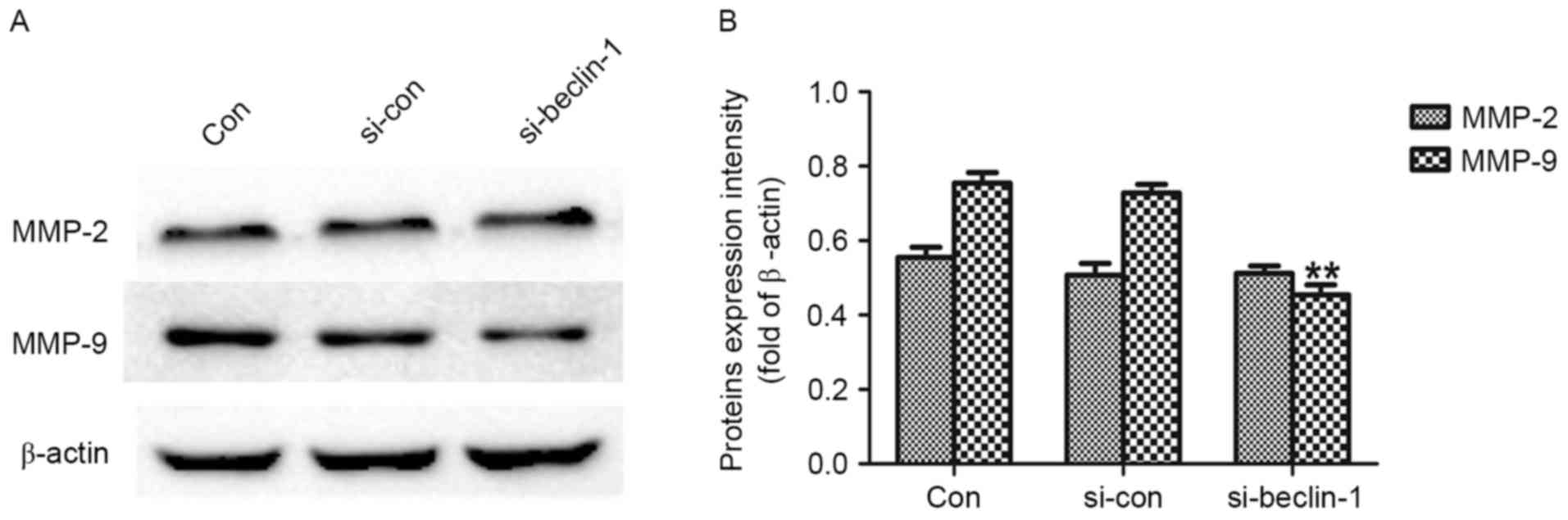
A western blot assay was performed to investigate
the effect of Beclin-1 knockdown on the expression of MMP-2 and
MMP-9, since it is generally acknowledged that they serve functions
in tumor invasion, and metastasis (12–14). As
presented in Fig. 3A and B, MMP-9
expression was significantly decreased in the si-Beclin-1 group
compared with the Con group (P<0.01); however, no significant
difference in the expression of MMP-2 was observed between the two
groups. These results suggest that Beclin-1 knockdown may inhibit
invasion and metastasis of SK-ES-1 cells via downregulation of the
expression of MMP-9.
Discussion
ES is an aggressive bone and soft tissue malignant
tumor that primarily affects children, and young adults (15). In previous years, the overall survival
rate has risen markedly for patients who present with localized
disease, owing to development of multi-agent systemic chemotherapy
and aggressive local control methods, which have resulted in
five-year event-free survival rates of 70–80% in these patients
(15,16). However, for the ~25% of patients who
present with metastatic disease, the prognosis is poor and
event-free survival rate for these patients remains <25%
(15). Thus, novel therapeutic
targets and increased understanding of the metastatic mechanism of
ES are required to achieve an improved outcome for these
patients.
The function of autophagy in cancer has been
highlighted in previous years. Autophagy serves a function in cell
cycle regulation, apoptosis, angiogenesis and other aspects of
tumor initiation and progression (17). Increasing evidence suggests that
autophagy contributes to the malignant phenotype in a number of
tumors, including lung adenocarcinoma, ovarian carcinoma and
esophageal squamous cell carcinoma (18–20).
Autophagy-related protein six, known as Beclin-1, the first
confirmed mammalian autophagic gene, has been demonstrated to
initiate autophagosome formation through binding to a class III
phosphatidylinositol-3-OH kinase (Vps34). Beclin-1 upregulates
autophagy by combining with Vps34, and other positive and negative
co-factors, including Bcl-2/B-cell lymphoma extra-large, Vps15,
Beclin-1-associated autophagy-related key regulator, Autophagy and
Beclin-1 regulator 1, High mobility group box 1, and Survivin to
form the Beclin-1 interactome (21).
Beclin-1 dysfunction has been identified in a number of disorders,
including cancer, aging and degenerative diseases; for example,
overexpression of Beclin-1 markedly promotes autophagic cell death
in leukemia cells (22).
Additionally, Beclin-1 knockdown using siRNA protects cells from
sorafenib-induced autophagic cell death in hepatocellular carcinoma
cells (23). On the contrary,
autophagy promotes tumor survival and progression in pancreatic
cancer (24). However, the exact
function of Beclin-1 in the proliferation and malignant phenotype
of ES cells remains unclear.
In the present study, it was demonstrated that
Beclin-1 knockdown inhibited proliferation, invasion and migration
in SK-ES-1 cells. The simultaneous decline in the expression of
MMP-9, which is generally established to be closely associated with
tumor invasion and metastasis, suggested that Beclin-1 knockdown
may inhibit invasion and migration of SK-ES-1 cells through
downregulating the expression of MMP-9.
Taken together, the results of the present study
suggest that Beclin-1 knockdown may suppress the growth and
malignant phenotype of SK-ES-1 cells by inhibition of MMP-9. Thus
Beclin-1 is a promising therapeutic target for treatment of ES,
particularly in patients that present with metastasis. Further
experiments on the in vitro effects of Beclin-1 knockdown on
apoptosis of ES cells may further resolve its viability as a
therapeutic target.
Acknowledgements
The present study was supported by The Foundation of
Health Department of Jiangxi Province (grant no. 2016A073) and
Gan-Po Talents Project 555 of Jiangxi Province.
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gaspar N, Hawkins DS, Dirksen U, Lewis IJ,
Ferrari S, Le Deley MC, Kovar H, Grimer R, Whelan J, Claude L, et
al: Ewing sarcoma: Current management and future approaches through
collaboration. J Clin Oncol. 33:3036–3046. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Levine B and Klionsky DJ: Development by
self-digestion: Molecular mechanisms and biological functions of
autophagy. Dev Cell. 6:463–477. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mizushima N: Autophagy: Process and
function. Genes Dev. 21:2861–2873. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mathew R, Karantza-Wadsworth V and White
E: Role of autophagy in cancer. Nat Rev. 7:961–967. 2007.
View Article : Google Scholar
|
|
6
|
Liang XH, Kleeman LK, Jiang HH, Gordon G,
Goldman JE, Berry G, Herman B and Levine B: Protection against
fatal Sindbis virus encephalitis by beclin, a novel
Bcl-2-interacting protein. J Virol. 72:8586–8596. 1998.PubMed/NCBI
|
|
7
|
Oberstein A, Jeffrey PD and Shi Y: Crystal
structure of the Bcl-XL-Beclin 1 peptide complex: Beclin 1 is a
novel BH3-only protein. J Biol Chem. 282:13123–13132. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu B, Bao JK, Yang JM and Cheng Y:
Targeting autophagic pathways for cancer drug discovery. Chin J
Cancer. 32:113–120. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Meijer AJ and Codogno P: Regulation and
role of autophagy in mammalian cells. Int J Biochem Cell Biol.
36:2445–2462. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sun Y, Liu JH, Jin L, Lin SM, Yang Y, Sui
YX and Shi H: Over-expression of the Beclin1 gene upregulates
chemosensitivity to anti-cancer drugs by enhancing therapy-induced
apoptosis in cervix squamous carcinoma CaSki cells. Cancer Lett.
294:204–210. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jin S and White E: Role of autophagy in
cancer: Management of metabolic stress. Autophagy. 3:28–31. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li H, Zhang K, Liu LH, Ouyang Y, Bu J, Guo
HB and Xiao T: A systematic review of matrix metalloproteinase 9 as
a biomarker of survival in patients with osteosarcoma. Tumour Biol.
35:5487–5491. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang J, Shi Q, Yuan TX, Song QL, Zhang Y,
Wei Q, Zhou L, Luo J, Zuo G, Tang M, et al: Matrix
metalloproteinase 9 (MMP-9) in osteosarcoma: Review and
meta-analysis. Clin Chim Acta. 433:225–231. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Shang HS, Chang JB, Lin JH, Lin JP, Hsu
SC, Liu CM, Liu JY, Wu PP, Lu HF, Au MK and Chung JG: Deguelin
inhibits the migration and invasion of U-2 OS human osteosarcoma
cells via the inhibition of matrix metalloproteinase-2/-9 in vitro.
Molecules. 19:16588–16608. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Balamuth NJ and Womer RB: Ewing's sarcoma.
Lancet Oncol. 11:184–192. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gorlick R, Janeway K, Lessnick S, Randall
RL and Marina N; COG Bone Tumor Committee, : Children's Oncology
Group's 2013 blueprint for research: Bone tumors. Pediatr Blood
Cancer. 60:1009–1015. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Filippi-Chiela EC, Villodre ES, Zamin LL
and Lenz G: Autophagy interplay with apoptosis and cell cycle
regulation in the growth inhibiting effect of resveratrol in glioma
cells. PLoS One. 6:e208492011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Pan B, Chen D, Huang J, Wang R, Feng B,
Song H and Chen L: HMGB1-mediated autophagy promotes docetaxel
resistance in human lung adenocarcinoma. Mol Cancer. 13:1652014.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bao LJ, Jaramillo MC, Zhang ZB, Zheng YX,
Yao M, Zhang DD and Yi XF: Nrf2 induces cisplatin resistance
through activation of autophagy in ovarian carcinoma. Int J Clin
Exp Pathol. 7:1502–1513. 2014.PubMed/NCBI
|
|
20
|
Chen Y, Li X, Wu X, He C, Guo L, Zhang S,
Xiao Y, Guo W and Tan B: Autophagy-related proteins LC3 and
Beclin-1 impact the efficacy of chemoradiation on esophageal
squamous cell carcinoma. Pathol Res Pract. 209:562–567. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kang R, Zeh HJ, Lotze MT and Tang D: The
Beclin 1 network regulates autophagy and apoptosis. Cell Death
Differ. 18:571–580. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tong Y, You L, Liu H, Li L, Meng H, Qian Q
and Qian W: Potent antitumor activity of oncolytic adenovirus
expressing beclin-1 via induction of autophagic cell death in
leukemia. Oncotarget. 4:860–874. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tai WT, Shiau CW, Chen HL, Liu CY, Lin CS,
Cheng AL, Chen PJ and Chen KF: Mcl-1-dependent activation of Beclin
1 mediates autophagic cell death induced by sorafenib and SC-59 in
hepatocellular carcinoma cells. Cell Death Dis. 4:e4852013.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ko YH, Cho YS, Won HS, Jeon EK, An HJ,
Hong SU, Park JH and Lee MA: Prognostic significance of
autophagy-related protein expression in resected pancreatic ductal
adenocarcinoma. Pancreas. 42:829–835. 2013. View Article : Google Scholar : PubMed/NCBI
|