Introduction
Hepatocellular carcinoma (HCC) is the most common
type of liver cancer which results in ~80% of mortalities
associated with hepatic cancer (1).
Surgical resection is the principal strategy for treatment of HCC.
The results of chemotherapy are often unsatisfactory as liver
cancer typically develops resistance to chemotherapy, which may be
due to a number of reasons; for example, the rapid metabolism of
liver cells may quickly inactivate chemotherapy drugs administered
to patients with HCC (2). In
addition, the factors which enable the progression of HCC remain
unidentified, and non-targeted drugs are less effective at
preventing cancer recurrence and metastasis (3).
Karyopherin subunit-α 2 (KPNA2) is a nuclear
transporter, allowing signal communication between the nucleus and
cytoplasm (4). KPNA2 is required for
cell survival (5); it may be mutated
in cancer to enable disease progression (6). KPNA2 has been associated with the
regulation of carcinogenesis, proliferation and recurrence in
various cancer types, as previously reviewed (7). KPNA2 contributes to the relocation of
DNA damage response proteins and has been associated with the
prognosis of breast cancer (4). KPNA2
recognizes cargo with the nuclear location sequence (NLS) and may
affect cancer cell progression by interacting with a number of
transcriptional factors, including c-Myc and p53 (8). Additionally, the upregulation of KPNA2
increases the expression of octamer-binding transcription factor
gene 4 (OCT4), which is associated with the stem-like properties
and dedifferentiation of cancer cells (9). However, the association between KPNA2
and cancer progression has yet to be characterized and requires
further study.
In the present study, it was identified that KPNA2
may serve a function in the progression of HCC. The knockdown of
KPNA2 expression was associated with the downregulation of
cancer-associated genes in HCC cells, and high KPNA2 expression was
associated with a significantly reduced overall and disease-free
survival time in patients with HCC.
Materials and methods
Tumor samples
The present study was approved by the Ethics
Committee of the Third Affiliated Hospital of Kunming Medical
University (Kunming, China) and written informed consent was
obtained from all patients. The patients had not received radiation
or chemotherapy prior to surgery. HCC tissues were obtained from 6
patients (3 males and 3 females), aged between 55 and 65 years old
(mean age, 62 years old) with non-metastatic disease during
surgical resection from the Department of Oncology of the Third
Affiliated Hospital of Kunming Medical University between September
2013 and May 2014. Adjacent liver tissues were used as controls.
All tissues were frozen with dry ice. RNA was extracted from the
samples with TRIzol (Invitrogen; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA), according to the manufacturer's protocol. The
gene expression profile was analyzed using GeneChip®
Human Transcriptome Array 2.0 microarrays (Affymetrix; Thermo
Fisher Scientific, Inc.) according to the manufacturer's protocol.
The raw data were analyzed by using the affy package (version
1.55.0; Affymetrix; Thermo Fisher Scientific, Inc.) in R (10). The robust multi-array average method
was employed to corrects probe intensity values and the microarray
data was then quantile-normalized (10) for the following comparisons. The
normalized microarray data were analyzed using Multiple Experiment
Viewer (version 4.9.0, http://mev.tm4.org/#/welcome).
Cell culture
The human HCC cell line Huh7 was purchased from the
Cell Bank of Type Culture Collection of Chinese Academy of Sciences
(Shanghai, China). The cells were cultured in Dulbecco's Modified
Eagle's Medium (Invitrogen; Thermo Fisher Scientific, Inc.)
supplemented with 5% fetal bovine serum (Gibco; Thermo Fisher
Scientific, Inc.). Cells were maintained in 37°C in an atmosphere
containing 5% CO2 throughout the study.
RNA interference
Short interfering (si)RNA was synthesized by Jima
Biotechnology Co., Ltd. (Shanghai, China); the si-KPNA2-1 sequence
is UUACGAUUAUGUGGUCGACGG, si-KPNA2-2 is UUGUUUGGUUCCGACACCAUC and
si-CTR is a commercial small RNA target with no sequence on known
human gene. A total of 20 pmol siRNA was transfected using 3 µl
RNAiMAX reagent/well (cat. no. 13778-100; Thermo Fisher Scientific,
Inc.) in 24-well plates (cells grown up to 70% confluence). Further
analysis, as subsequently described, was performed at 48 h after
transfection.
MTT assay
A CellTiter 96® non-radioactive cell
proliferation assay (cat. no. G4000; Promega Corporation, Madison,
WI, USA) was used to analyze cell proliferation, according to the
manufacturer's protocol. Briefly, 8,000 Huh7 cells were seeded in
96-well plates. Alterations in cell confluence were observed at 24
and 48 h, followed by the addition of 15 µl Dye Solution to each
well. Following a further incubation of 2–3 h, 100 µl Solubilizing
Solution/Stop mix was added to each well and the absorbance was
determined at wavelength of 570 nm on a 96-well plate reader.
Reverse transcription-quantitative
polymerase chain reaction
RNA was extracted from Huh7 cells using TRIzol, and
reverse transcription was performed using the PrimeScript 1st
strand cDNA Synthesis kit (cat. no. 6110A; Takara Bio, Inc., Otsu,
Japan), according to the manufacturer's protocols. The SsoFast
EvaGreen Supermix with Low ROX kit (cat. no. 1572-5211; Bio-Rad
Laboratories, Inc., Hercules, CA, USA) was applied to quantify the
mRNA expression level, according to the manufacturer's protocol.
The primers used in qPCR were as follow: KPNA2 forward,
CTGCAGGAAAACCGCAACAA and reverse, CCTGGCAGCTTGAGTAGCTT; CDK1
forward, TTTCTTTCGCGCTCTAGCCA and reverse, CAATCGGGTAGCCCGTAGAC;
CCNB2 forward, GGCTGGTACAAGTCCAC-TCC and reverse,
CTTCTTCCGGGAAACTGGCT; β-actin forward, GTCATTCC-AAATATGAGATGCGT and
reverse, GCTATCACCTCCCCTGTGTG. Thermocycling protocol included
pre-denatured at 95°C for 3 min, denaturation at 95°C for 15 sec,
annealing at 60°C for 15 sec and extension at 72°C for 1 min for 40
cycles. The 2−ΔΔCq method was used for quantification of
gene relative expression (11).
Cell cycle analysis
After cell transfection with siRNA for 24 h, the
cells were dissociated using 0.25% trypsin in a 37°C cell incubator
for 5 min. The detached cells were washed with PBS once and then
fixed with cold 70% ethanol for 30 min at room temperature.
Briefly, the cells were washed twice with PBS. The cells were
treated with a final concentration of 5 µg/ml ribonuclease for 10
min at room temperature. Propidium iodide (PI) was added (10 µg/ml)
and the cells were stained for 15 min at room temperature. The
cells were analyzed using FACSCanto (BD Biosciences, Inc., San
Jose, CA, USA). A wavelength of 605 nm was selected to determine
the fluorescence of PI.
Bioinformatic data mining
Cancer Genome Atlas (TCGA) data was used. TCGA raw
data was obtained and analyzed using the cBioportal (http://www.cbioportal.org/) and Oncomine (https://www.oncomine.org/) online tools. A total of
440 mRNA expression profiles for HCC (440 adjacent liver tissues
were used as control tissues) were selected. The KPNA2 gene was
input, with the cut-off for high/low expression level set at >1
standard deviation. To evaluate the hazard ratio of KPNA2, the
OncoLnc (http://www.oncolnc.org/) was used to
investigate the Cox coefficients of KPNA2 in HCC patients.
Statistical analysis
Pearson and Spearman correlation analysis were
respectively performed to investigate the correlation between the
KPNA2 and CDK1 as well as CCNB2. Cox's proportional hazards model
was used for survival rate analysis. Kaplan-Meier estimator curves
were plotted for survival analysis and a log-rank test was
performed to determine the statistical significance of differences
in survival between groups. GraphPad prism 7 (GraphPad Software,
Inc., La Jolla, CA, USA) was used for data analysis. Data are
presented as the mean ± standard error of the mean. Student's
t-test was used to compare data between groups.
Results
KPNA2 mRNA expression is deregulated
in hepatocellular carcinoma tissues and associated with cell
regulators
A total of 6 samples from patients with HCC were
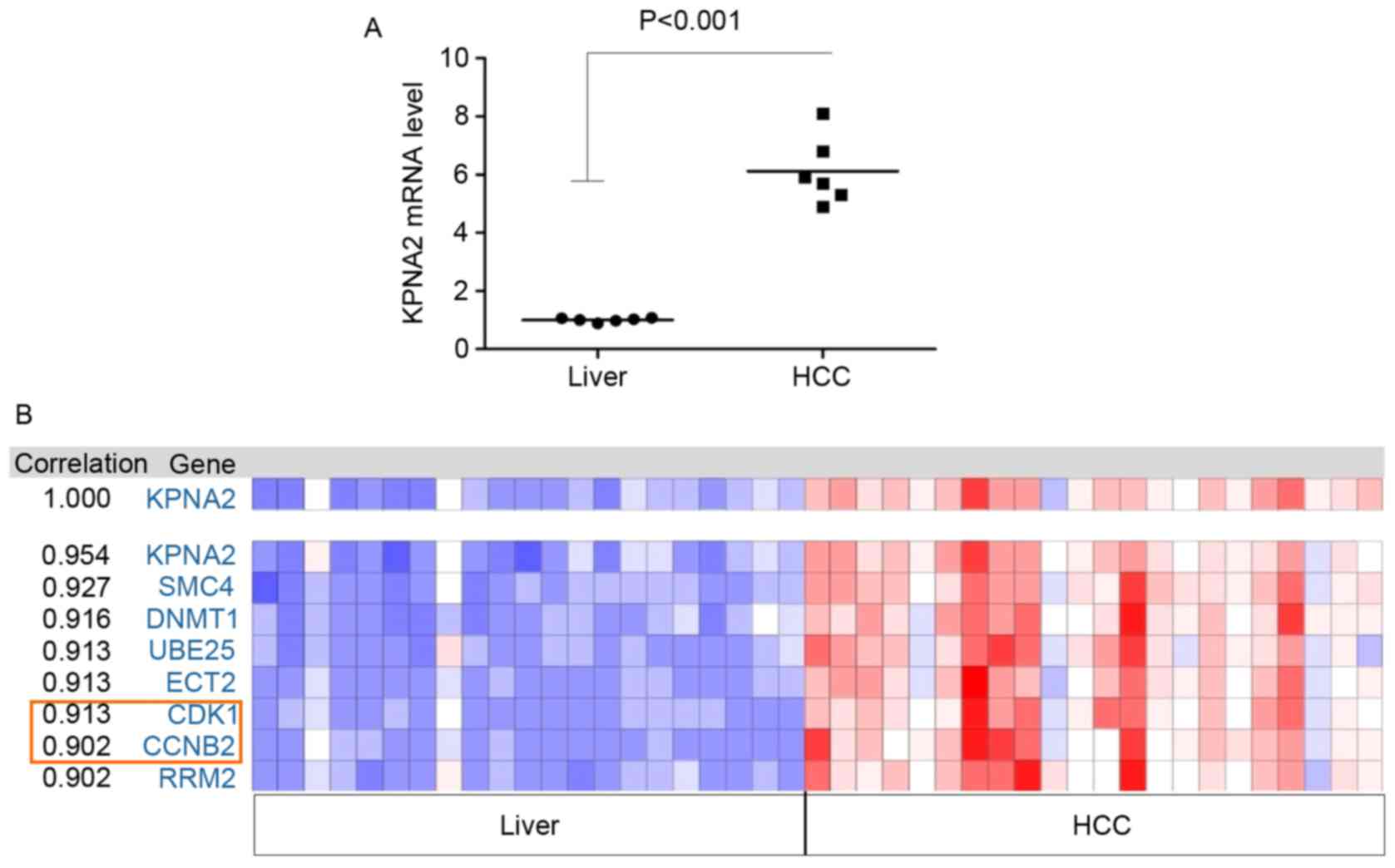
analyzed using microarrays. The mRNA expression of certain genes
was upregulated compared with adjacent liver tissues in all tumor
tissue samples; KPNA2 was the most significantly upregulated of all
the differentially expressed genes (P<0.001; Fig. 1A). Therefore, the function of KPNA2 in
the progression of malignant HCC was selected for further study.
The Oncomine online tool was used to identify the correlation
between the expression of KPNA2 and other identified genes from the
TCGA tumor mRNA microarray data. In addition, the correlation in
expression between KPNA2 and cell cycle-associated genes, including
the cyclin family and CDKs, was analyzed. The results demonstrated
that the expression of CCNB2 and CDK1 was correlated with KPNA2
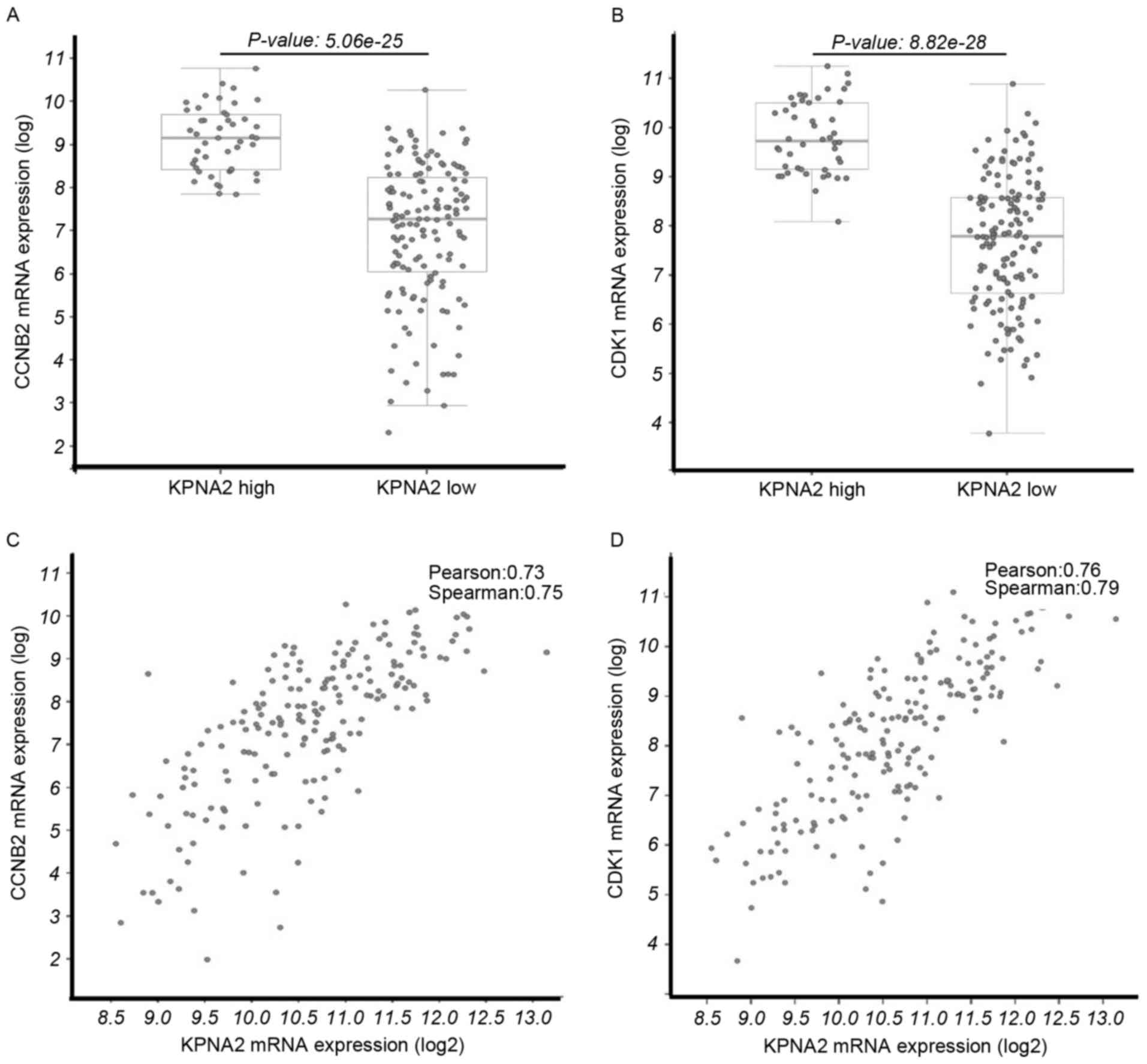
(Fig. 1B). The mRNA expression levels
of CCNB2 (P=5.06×10−25; Fig.
2A) and CDK1 (P=8.82×10−28; Fig. 2B) were significantly upregulated in
the KPNA2 mRNA high expression group and the expression levels of
CDK1 and CCNB2 were positively associated with the KPNA mRNA
expression level (Fig. 2C and D).
Knockdown of KPNA2 affects the
expression levels of CDK1 and cyclin B2, induces G2/M
cell cycle arrest and inhibits cell proliferation
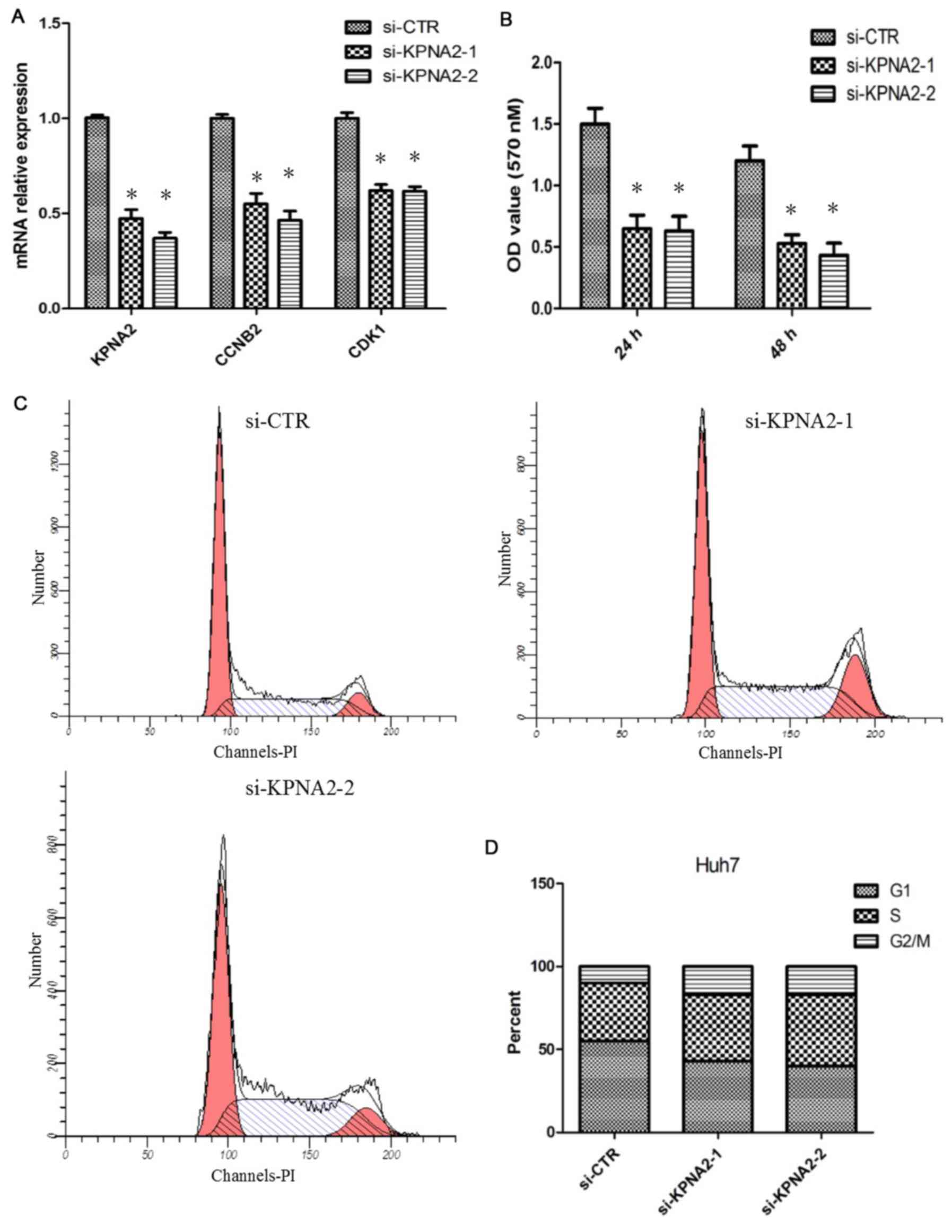
To identify the function of KPNA2 in hepatic tumor
cell proliferation, siRNA was used to knockdown KPNA2 expression in
Huh7 cells. The efficiency of the knockdown of KPNA2 in Huh7 cells
was analyzed with RT-qPCR. It was demonstrated that the siRNA
significantly reduced the KPNA2 mRNA level; the expression level of
CCNB2 and CDK1 mRNA was simultaneously decreased (P<0.05;
Fig. 3A). An MTT assay was performed
to determine the effect of KPNA2 knockdown on Huh7 cell
proliferation. The result demonstrated that KPNA2 knockdown
significantly decreased the proliferative abilities of cells
(P<0.05; Fig. 3B). As CDK1 and
CCNB2 are associated with the regulation of the G2/M
transition in the cell cycle, the effect of KPNA2 knockdown on cell
cycle distribution was investigated using flow cytometry. The
results suggested that knockdown of KPNA2 resulted in
G2/M arrest in Huh7 cells (Fig. 3C and D).
Expression of KPNA2 in tumor tissues
is negatively associated with the prognosis
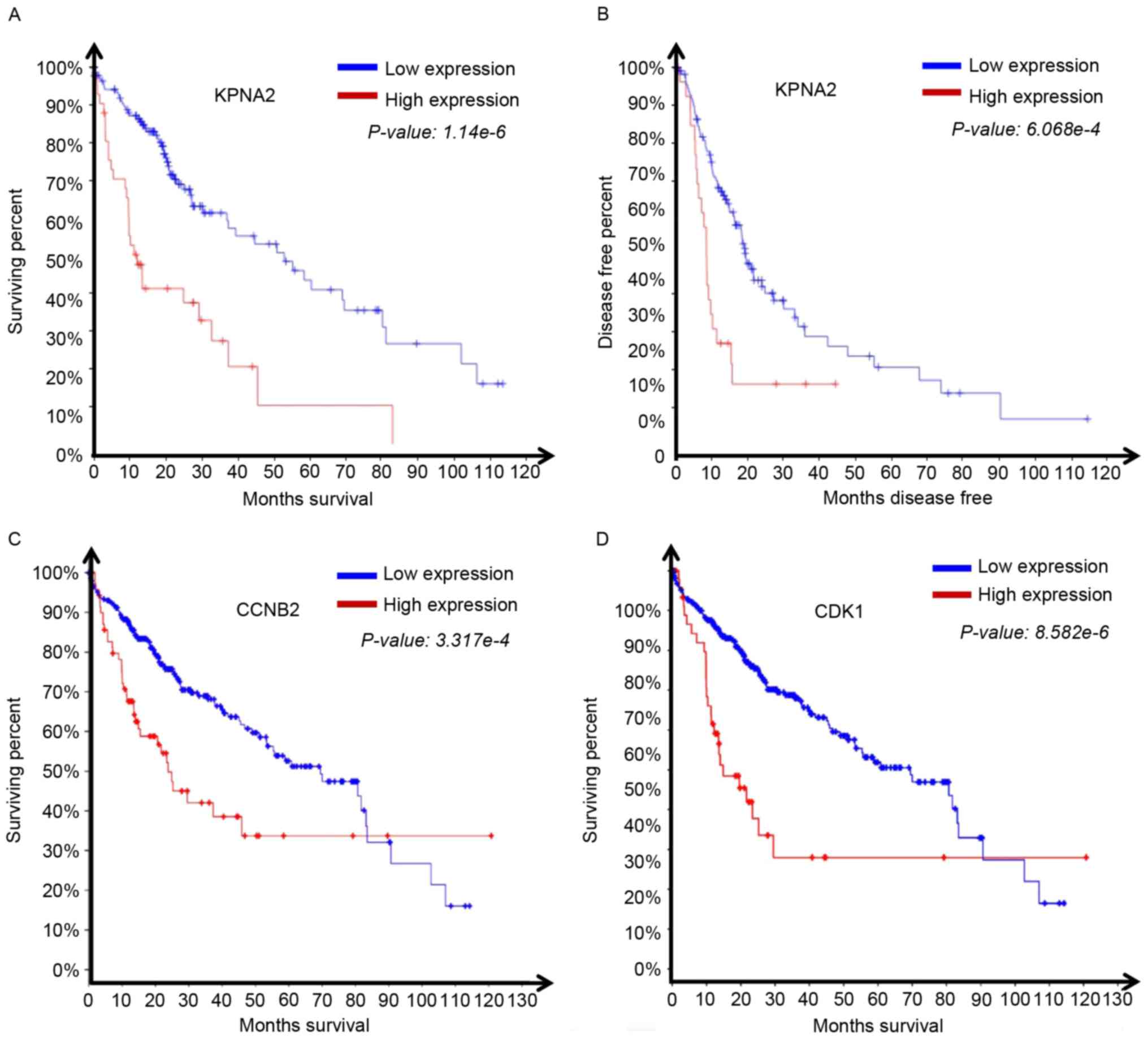
The survival data associated with HCC expression
profiles from TCGA was analyzed. The results demonstrated that
patients with high KPNA2 expression exhibited significantly reduced
overall (1.14×10−6; Fig.
4A) and disease-free survival time (6.07×10−4;
Fig. 4B) compared with the patients
with low KPNA2 expression. We submitted in OncoLnc (http://www.oncolnc.org). The hazard ratio for high
KPNA2 expression in HCC was 0.617. It was then assessed whether the
expression of the KPNA2-associated genes CDK1 and CCNB2 affected
overall survival time. The overall survival time of patients
exhibiting a relatively high level of CCNB2
(P=3.32×10−4; Fig. 4C) or
CDK1 (P=8.58×10−6; Fig.
4D) was significantly reduced. The results revealed a role for
KPNA2, CDK1 and CCNB2 expression in the progression of HCC.
Discussion
HCC occurs worldwide, and the diversity in the
genetic backgrounds of patients with HCC may cause difficulty in
identifying the genes that promote the progression of HCC (12). In the present study, tumor tissues
were selected from 6 patients with HCC and analysis was performed
using microarrays; it was identified that KPNA2 was the most
significantly upregulated gene. Therefore, it was hypothesized that
KPNA2 may promote cancer progression.
TCGA is a public cancer research database containing
high-throughput genome sequencing and transcriptome data to enable
the study of cancer with bioinformatics tools. TCGA data was
selected using the cBioportal and Oncomine online tools (13). To identify the genes associated with
the KPNA2 gene in a wider HCC population. In the present study, the
expression level of genes in the cyclin B family were demonstrated
to be associated with the expression of KPNA2.
Unrestricted cell growth and proliferation is a
hallmark of cancer cells (14). In a
normal cell, cell proliferation is precisely controlled by cell
cyclins and CDKs (15). In the
present study, CCNB2 and CDK1 were relatively abundant in tumor
tissues and their expression was significantly correlated with
KPNA2 expression. CDK1, also known as CDC2, binds cyclin B1 and B2
to promote G2/M transition (16). The
dimerization of CCNB2 with CDK1 is an essential component of the
cell cycle regulatory machinery (17). A previous study demonstrated that
KPNA2 affected cancer progression by regulating DNA damage response
protein subcellular location (4). The
CDK1 amino acid sequence was analyzed and an NLS was identified
(18), which may be recognized by
KPNA2 to result in CDK1 nuclear transportation. Further study is
required to characterize the process of CCNB2/CDK1 transcriptional
activation, the effect on cell cycle progression and how this is
associated with KPNA2.
To validate an interaction between KPNA2, CCNB2 and
CDK1 in HCC, Huh7 cells were selected. KPNA2 expression was
silenced with siRNA. It was demonstrated that KPNA interference
directly decreased the expression of cyclin B2 and CDK1 at the
mRNA. In addition, it was revealed that cell proliferation was
inhibited by KPNA interference. Therefore, the prediction of genes
associated with genes, including KPNA2, using the cBioportal tool
may be reliable, as the prediction result could be experimentally
verified. Additionally, the present study aimed to determine
whether KPNA2 was associated with the clinical prognosis for
patients with HCC. Significant differences in overall and
disease-free survival time were identified between patients with
high and low KPNA2 expression; patients with high KPNA2 expression
exhibited a relatively poor prognosis.
In conclusion, the results of the present study
revealed that high KPNA2 expression is a risk factor in HCC and
that KPNA2 regulated the cell cycle checkpoint-associated proteins
cyclin B2 and CDK1. Therefore, KPNA2 may be a novel therapeutic
target in HCC.
Acknowledgements
The authors thank Dr Xi Lan Shi (Department of
Pharmacology, Kunming Medical University) for his critical comments
on data analysis and presentation.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Housman G, Byler S, Heerboth S, Lapinska
K, Longacre M, Snyder N and Sarkar S: Drug resistance in cancer: An
overview. Cancers (Basel). 6:1769–1792. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sukowati CH, Rosso N, Crocè LS and
Tiribelli C: Hepatic cancer stem cells and drug resistance:
Relevance in targeted therapies for hepatocellular carcinoma. World
J Hepatol. 2:114–126. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Alshareeda AT, Negm OH, Green AR, Nolan
CC, Tighe P, Albarakati N, Sultana R, Madhusudan S, Ellis IO and
Rakha EA: KPNA2 is a nuclear export protein that contributes to
aberrant localisation of key proteins and poor prognosis of breast
cancer. Br J Cancer. 112:1929–1937. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Huang L, Wang HY, Li JD, Wang JH, Zhou Y,
Luo RZ, Yun JP, Zhang Y, Jia WH and Zheng M: KPNA2 promotes cell
proliferation and tumorigenicity in epithelial ovarian carcinoma
through upregulation of c-Myc and downregulation of FOXO3a. Cell
Death Dis. 4:e7452013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Teng SC, Wu KJ, Tseng SF, Wong CW and Kao
L: Importin KPNA2, NBS1, DNA repair and tumorigenesis. J Mol
Histol. 37:293–299. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Christiansen A and Dyrskjøt L: The
functional role of the novel biomarker karyopherin α 2 (KPNA2) in
cancer. Cancer Lett. 331:18–23. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kim IS, Kim DH, Han SM, Chin MU, Nam HJ,
Cho HP, Choi SY, Song BJ, Kim ER, Bae YS and Moon YH: Truncated
form of importin alpha identified in breast cancer cell inhibits
nuclear import of p53. J Biol Chem. 275:23139–23145. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li XL, Jia LL, Shi MM, Li X, Li ZH, Li HF,
Wang EH and Jia XS: Downregulation of KPNA2 in non-small-cell lung
cancer is associated with Oct4 expression. J Transl Med.
11:2322013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gautier L, Cope L, Bolstad BM and Irizarry
RA: affy-analysis of Affymetrix GeneChip data at the probe level.
Bioinformatics. 20:307–315. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Maass T, Sfakianakis I, Staib F, Krupp M,
Galle PR and Teufel A: Microarray-based gene expression analysis of
hepatocellular carcinoma. Curr Genomics. 11:261–268. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cerami E, Gao J, Dogrusoz U, Gross BE,
Sumer SO, Aksoy BA, Jacobsen A, Byrne CJ, Heuer ML, Larsson E, et
al: The cBio cancer genomics portal: An open platform for exploring
multidimensional cancer genomics data. Cancer Discov. 2:401–404.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Malumbres M and Barbacid M: Cell cycle,
CDKs and cancer: A changing paradigm. Nat Rev Cancer. 9:153–166.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Dorée M and Hunt T: From Cdc2 to Cdk1:
When did the cell cycle kinase join its cyclin partner? J Cell Sci.
115:2461–2464. 2002.PubMed/NCBI
|
|
17
|
Huang Y, Sramkoski RM and Jacobberger JW:
The kinetics of G2 and M transitions regulated by B cyclins. PLoS
One. 8:e808612013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kosugi S, Hasebe M, Tomita M and Yanagawa
H: Systematic identification of cell cycle-dependent yeast
nucleocytoplasmic shuttling proteins by prediction of composite
motifs. Proc Natl Acad Sci USA. 106:pp. 10171–10176. 2009;
View Article : Google Scholar : PubMed/NCBI
|