Cancers result from abnormal cell growth, which is
caused by abnormalities in the genome and the epigenome (1). Alterations of epigenetic modifications
are always reversible and can be influenced by external factors,
including diet or environmental exposure (2). Given that epigenetic dysregulation
occurs early during tumorigenesis, epigenetic modifications have
been confirmed as potential targets for cancer prevention or cancer
therapy (3). Nutri-epigenetics, which
revolves around the impact of micronutritions and macronutritions
on epigenetic mechanisms, has renewed the study of traditional
methods in the epigenetic field. Epigenetic modifications in
response to functional foods consist of DNA methylation, histone
modifications and the effect of non-coding RNAs. However, it has
been confirmed that a few functional foods not only play a role in
health promotion, but also promote anticancer activity for several
tumors. Epigallocatechin-3-gallate (EGCG) extracted from green tea
inhibits DNA methylation and increases histone acetylation to
recover silenced tumor suppressor genes in cancer cells (4,5).
Isothiocyanates extracted from cruciferous vegetables suppress the
metastasis potential in lung cancer cells by inducing apoptosis and
cell cycle arrest, and inhibiting deubiquitinating enzymes
associated with tumorigenesis to exert anticancer effects (6,7).
The occurrence of gastric adenocarcinoma is a
stepwise process that may follow several years or decades of
gastritis, intestinal metaplasia and dysplasia to malignancy with
epigenetic alterations (3). Gradual
epigenetic dysregulations are strongly associated with the
tumorigenesis of certain inflammation-associated cancer types,
particularly gastric cancer (GC). Hence, more and more natural
anticancer components are being studied. The search for effective
and safe natural anticancer factors, and mechanisms to reverse or
counteract cancer-associated epigenetic alterations for GC
prevention and interventions are required.
To date, nutraceuticals for cancer chemoprevention
have primarily been derived from plants, for example, curcumin from
ginger plants (8), allicin from
garlic (9), resveratrol from grapes
and EGCG from green tea (10,11). However, the influence of
animal-derived macronutrition, including proteins or peptides, on
cancer epigenetic mechanisms remains uninvestigated. Despite the
fact that the consumption of red meat is undeniably associated with
cancer risk (12–14), molecular mechanisms of animal peptides
on epigenetic regulation have not been fully elucidated. The
soft-shelled turtle is commonly consumed in China. According to
excavations from the Hemudu site (Zhejiang, China), the history of
eating soft-shelled turtles in Asia dates back to the Hemudu
culture ~6,000 years ago in the neolithic period. The soft-shelled
turtle is also used in traditional Chinese medicine where it is
believed to strengthen immunity among other benefits (15). Recently, with the development of
turtle aquaculture, soft-shelled turtles have been processed into
various types of health products, including products in capsule and
liquid forms, and are used to improve the prognosis of cancer
patients undergoing radiotherapy and chemotherapy, according to the
beliefs of traditional Chinese medicine (16). To the best of our knowledge, for the
first time, the present study investigates the impact of
soft-shelled turtle peptides on epigenetic mechanisms in GC cells.
A microarray was used to detect the expression profile of microRNA
(miRNA) in the human GC AGS cell line treated with soft-shelled
turtle peptide. The target genes of the soft-shelled turtle
peptide-specific miRNAs and corresponding pathways were further
analyzed to determine the potential anticancer properties of the
soft-shelled turtle peptide.
The peptides extracted from soft-shelled turtles
were offered by Zhejiang Agricultural Group Co., Ltd. (Hangzhou,
China) and stored in the form of a dry powder in a brown glass
bottle at room temperature. Every unit of the soft-shelled turtle
peptide powder consisted of a polypeptide (40.68%), acid soluble
protein (47.97%), free amino acids (7.29%) and hydroxyproline
(4.06%). The polypeptide was the major functional component and the
treatment concentration was dependent on the actual polypeptide
content. The peptide powder was dissolved in cell culture medium at
the required concentration immediately prior to use.
The human GC AGS cell line was purchased from
American Type Culture Collection (ATCC; Manassas, VA, USA) and
cultured in F12 medium supplemented with 10% fetal bovine serum
(Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA) in T-75
flasks (Corning Inc., Corning, NY, USA) at 37°C in a humidified
atmosphere supplied with 5% CO2. The AGS cells were
treated during the exponential phase of growth. The experimental
group of cells were incubated with the soft-shelled turtle peptide
dissolved in F12 medium (100 mg/l), while the control group of
cells were incubated with F12 medium only. The experimental group
and the control group were incubated for 72 h, and the medium was
renewed every 24 h. After 72 h, the cells were collected by
trypsinization.
The total RNA of AGS cells with or without
soft-shelled turtle peptide treatment was extracted and purified
using the mirVana™ miRNA Isolation kit without phenol (Ambion;
Thermo Fisher Scientific, Inc.), following the manufacturers
instructions, and checked for an RNA integrity number to inspect
RNA integration using an Agilent Bioanalyzer 2100 (Agilent
Technologies, Santa Clara, CA, USA). The genome-wide miRNA
expression profiling was detected by the Affymetrix platform with
Genechip miRNA 3.0 array (Affymetrix; Thermo Fisher Scientific,
Inc.) based on the Sanger miRBase version 17. This chip contained
19,913 probe sets, including 1,789 mature miRNAs, 1,693 precursor
miRNAs and 2,336 small nucleolar RNAs and small Cajal body-specific
RNAs. After total RNA quality inspection, total RNAs of each sample
were tailed with poly A and labeled with biotin by the FlashTag™
Biotin HSR RNA Labeling kit (Affymetrix; Thermo Fisher Scientific,
Inc.) according to the manufacturers instructions. Hybridization of
bio-labeled RNA samples was performed according to the
manufacturers instructions for the Affymetrix GeneChip miRNA 3.0
Array with GeneChip Hybridization Wash and Stain kit, and GeneChip
Hybridization Oven 645 (all from Affymetrix; Thermo Fisher
Scientific, Inc.). After washing and staining with GeneChip
Fluidics Station 450, the arrays were scanned by the GeneChip
Scanner 3000 (both from Affymetrix; Thermo Fisher Scientific,
Inc.).
Differentially expressed miRNAs were evaluated by
the ratio of fluorescence between the control and the soft-shelled
turtle peptide treated sample. Fold-change of ≥2 and ≤0.5, used for
threshold values, were regarded as upregulation and downregulation,
respectively. Hierarchical clustering analysis was performed to
display the discrepant miRNA expression profile in two samples
through red and green color blocks via Gene Cluster (version 3.0)
and Java TreeView software program (bonsai.hgc.jp/~mdehoon/software/cluster/software.htm).
The potential target genes of differentially expressed miRNAs in
the treated cells were predicted using five databases, including
TARGETMINER (www.isical.ac.in/~bioinfo_miu/targetminer20.htm),
miRDB (mirdb.org), microRNA (www.mirbase.org/), TarBase (diana.imis.athena-innovation.gr/DianaTools/index.php?r=tarbase/index)
and RNA22 (cm.jefferson.edu/rna22/Interactive). Gene Ontology
(GO) analysis and Kyoto Encyclopedia of Genes and Genomes (KEGG)
pathway analysis (https://david.ncifcrf.gov/; DAVID Bioinformatics
Resources 6.8) were performed to demonstrate corresponding
biological processes and the regulatory network in which specific
miRNA-genes may have participated (17,18).
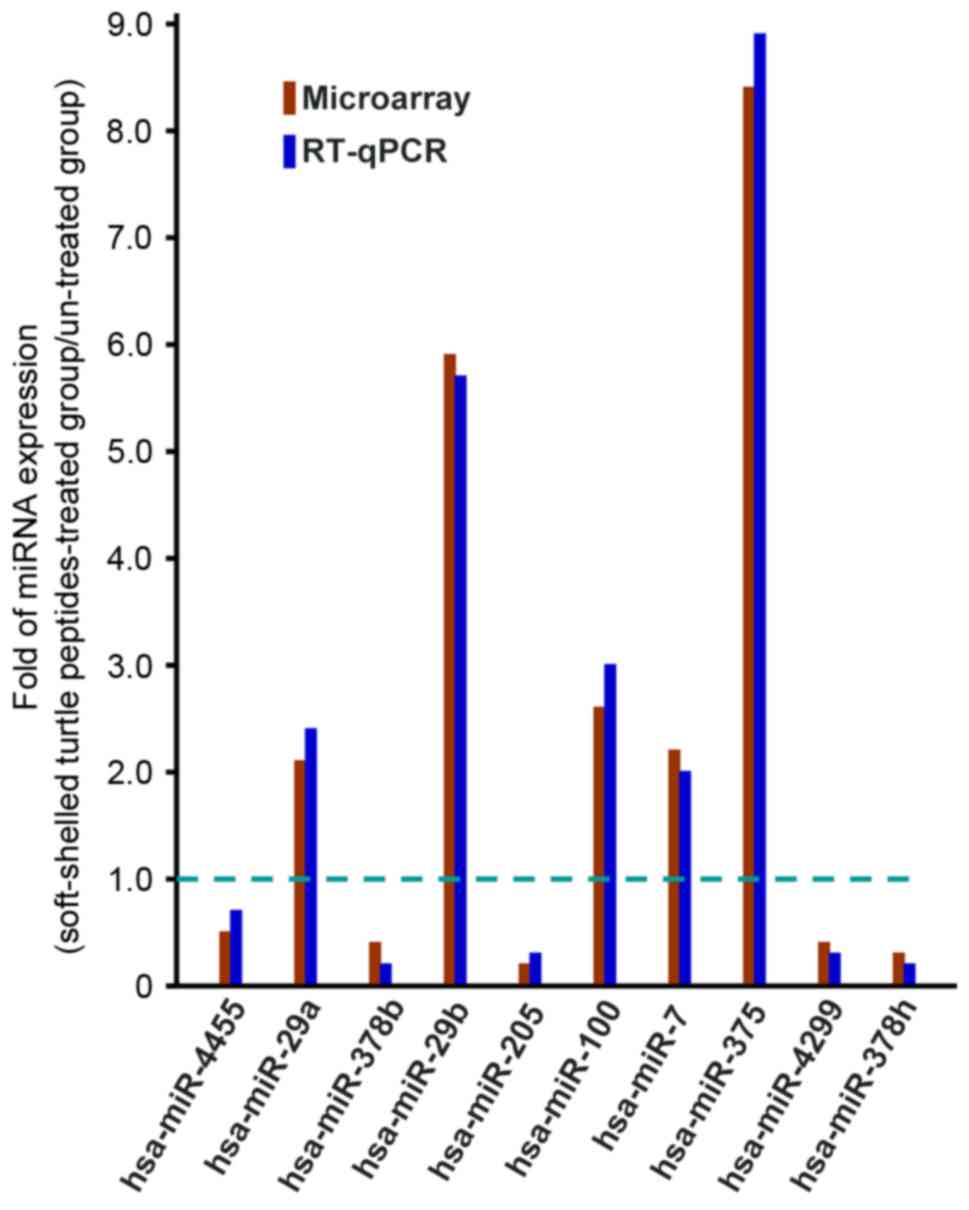
Differentially expressed miRNAs were selected and
validated according to RT-qPCR analysis with Platinum Quantitative
PCR SuperMix-UDG w/ROX (Invitrogen; Thermo Fisher Scientific,
Inc.). Total RNA were extracted from AGS cells with or without
soft-shelled turtle peptide treatment. PCR was performed using a
Applied Biosystems 7500 (Foster City, CA, USA). Amplification was
performed at 95°C for 30 sec, followed by 40 cycles of 95°C for 5
sec and 60°C for 34 sec. U6 was selected as an internal reference
to normalize the miRNA expression levels, and each sample was
validated in triplicate. The specific primer sequences of U6 is as
follow: 5′-CGCAAGGATGACACGCAAATTC-3. The reverse transcription of
poly-A tailed miRNAs using anchored Oligo dT primers for the
candidate miRNAs are presented in Table
I. The relative expression of differentially expressed miRNAs
was evaluated according to the 2−∆∆Cq method (19).
In order to evaluate the soft-shelled turtle
peptide-mediated anticancer capabilities in GC cells, the AGS cells
were treated with 100 mg/l soft-shelled turtle peptide for 72 h and
the medium was renewed every 24 h. A total of 1,744 human miRNAs
were screened to analyze the regulation of soft-shelled turtle
peptide on miRNA expression in the AGS cells. Compared with the
untreated cells, the miRNA expression of AGS cells with
soft-shelled turtle peptide treatment varied markedly. According to
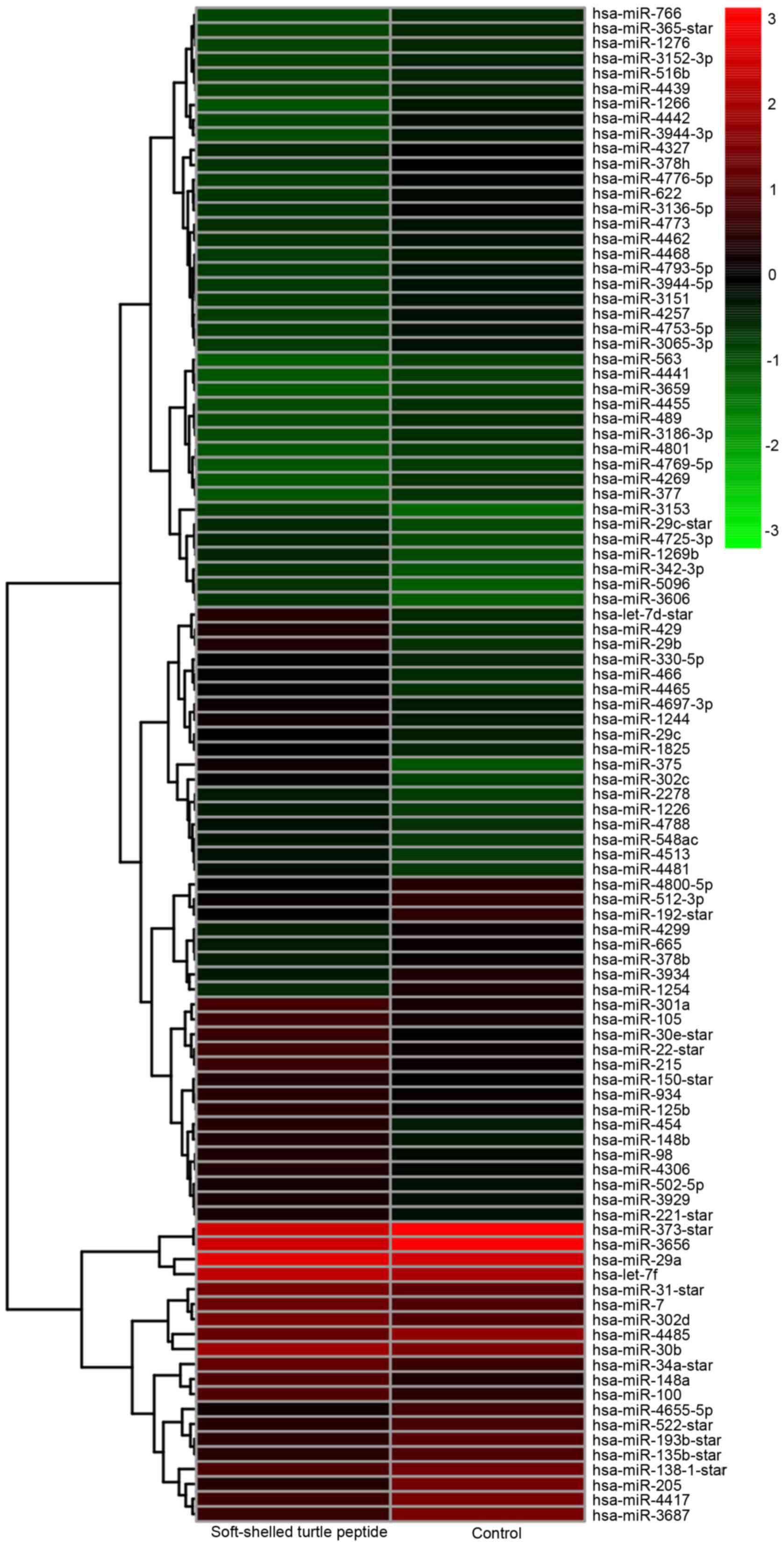
the hierarchical cluster analysis, there were 101 differentially
expressed miRNAs in the AGS cells treated with the soft-shelled
turtle peptide (Fig. 1). Among them,
49 miRNAs were upregulated and 52 miRNAs were downregulated after
treatment with the soft-shelled turtle peptide (Table II). Particularly, the expression of
miRNA-375 was increased intensely by 8.32-fold and other
upregulated miRNAs, including let-7d, miRNA-29b, miRNA-429,
miRNA-454, miRNA-148a, miRNA-22, miRNA-30e, miRNA-302 and
miRNA-148b, were altered markedly by >3-fold. Additionally, the
expression of miRNA 205, miRNA-1254, miRNA-3687 and miRNA-1266 was
significantly downregulated by <0.25-fold.
With the identification of differentially expressed
miRNAs in AGS cells following treatment of the soft-shelled turtle
peptide, the target genes of these miRNA were further investigated
according to 5 online software programs, including TargetMiner,
miRDB, microRNA, TarBase, and RNA-22 (Table IV). As presented in Table V, potential target genes were
identified.
To further evaluate the role of soft-shelled turtle
peptide influenced miRNAs in physiological functions and
pathological processes in GC, interactions between miRNAs and
associated target genes were predicted according to GO analysis and
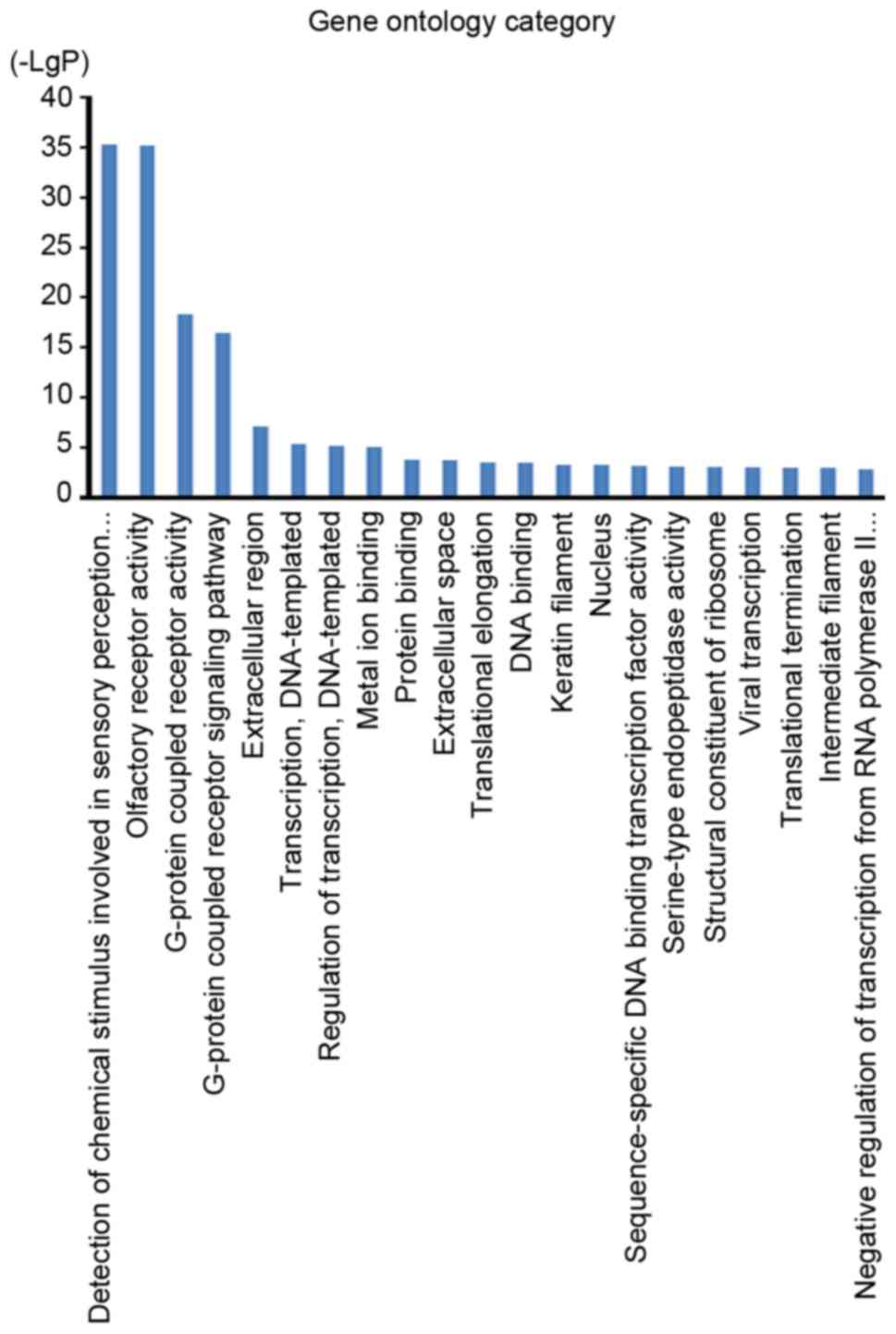
pathway analysis. The GO cellular component analysis demonstrated
that target genes were mainly clustered into the extracellular
region, extracellular space, keratin filament and nucleus
categories (Fig. 3). The GO molecular
function analysis indicated that the bulk of the target genes
participated in olfactory receptor activity and G-protein coupled
receptor activity (Fig. 3). In
addition, the GO biological process analysis demonstrated that
these target genes were closely associated with the detection of
chemical stimuli involved in the sensory perception of smell and
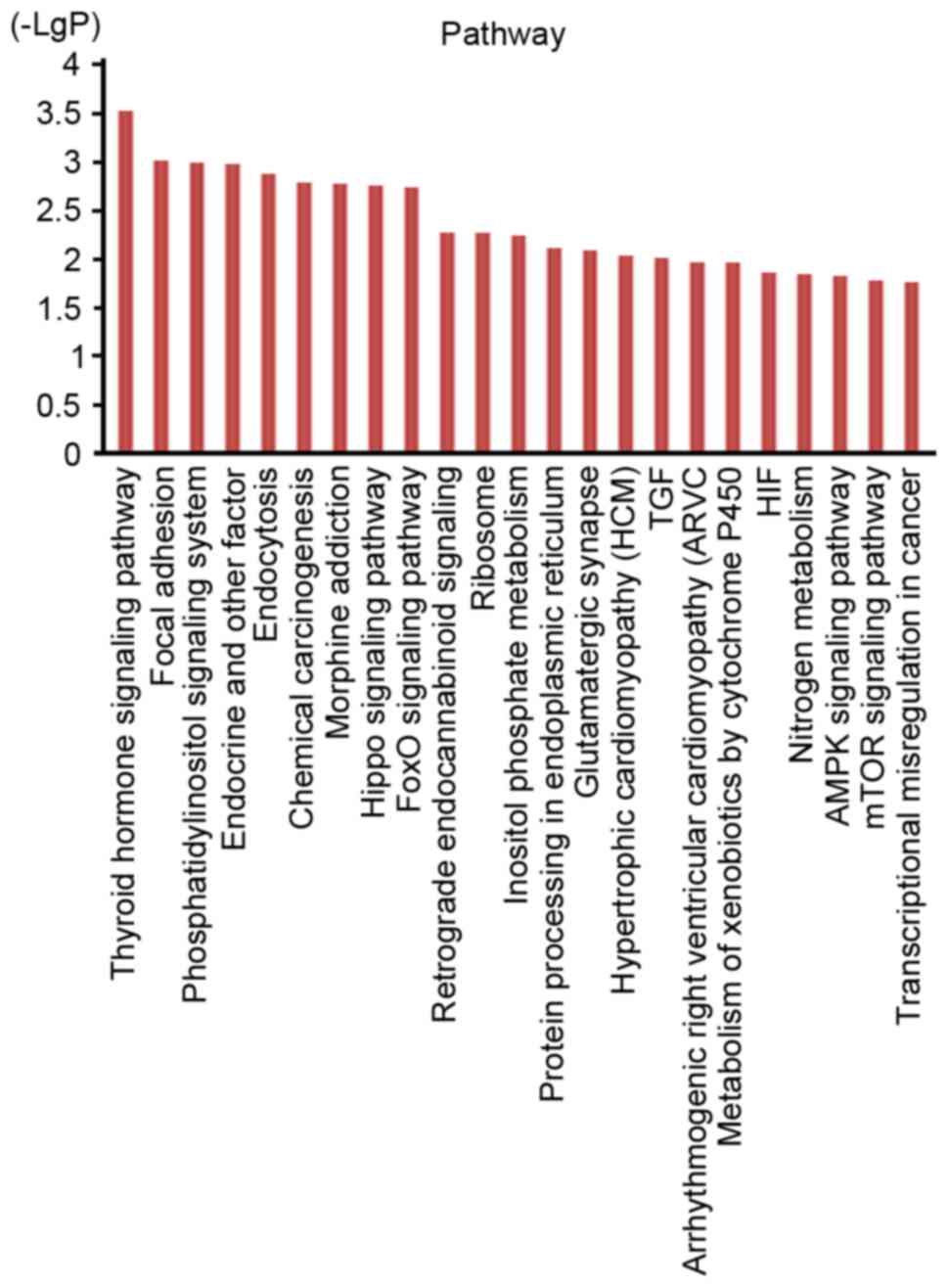
the G-protein coupled receptor signaling pathway (Fig. 3). However, the pathway analysis
indicated that the target genes were associated with the thyroid
hormone signaling pathway, the Hippo signaling pathway, the
forkhead box O (FoxO) signaling pathway, the AMP-activated protein
kinase signaling pathway, the mechanistic target of rapamycin
signaling pathway and transcriptional misregulation in cancer,
among others (Fig. 4).
Dietary components function as cancer
chemoprevention agents principally through improving the
nutritional supplement, enhancing immunity or directly blocking
cancer cells (49). Soft-shelled
turtle derived proteins can alleviate the side effects of cancer
patients undergoing chemotherapy and radiotherapy, including
myelosuppression (16). In the
present study, the primary focus was upon the direct influence of
soft-shelled turtle extract on the miRNA expression profile in
human GC AGS cells.
From the results of the miRNA array, a total of 101
differentially expressed miRNAs, including 49 upregulated miRNAs
and 52 downregulated miRNAs, were identified. Among the 49 miRNAs
with increased expression, the expression of miRNA-375 was altered
markedly, with 8.32-fold upregulation. The expression of miRNA-375
has been identified to be frequently downregulated in various types
of cancer, including GC, head and neck squamous cell carcinoma,
esophageal cancer, pancreatic ductal adenocarcinoma and
hepatocellular carcinoma (21,50–53).
In GC, miRNA-375 may function as a tumor suppressor. Overexpression
of miRNA-375 could inhibit the proliferation of GC cells by
targeting Janus kinase 2 (21).
Moreover, Snail, a metastasis-associated transcription factor,
directly binds to the putative promoter of miR-375 (54). Chang et al (55) also identified that the expression of
miRNA-375 was dependent on Helicobacter pylori infection in
GC; the miRNA-375 expression level in H. pylori-positive GC
tissues was lower than that in the H. pylori-negative GC
tissues. In the present study, the expression of miRNA-375 was
upregulated to the greatest extent following treatment with the
soft-shelled turtle peptide. This indicated that the soft-shelled
turtle peptide may have the capacity to block the proliferation and
metastasis of GC cells by increasing the expression of miRNA-375.
Other significantly upregulated miRNAs, including let-7d,
miRNA-29b, miRNA-429, miRNA-22, miRNA-148a/148b and miRNA-34a,
played an anticancer role in GC. miRNA-429 acts as a tumor
suppressor and has been identified to be downregulated in certain
cancer types, including GC (26),
clear cell renal cell carcinoma (56)
and glioblastoma multiforme (57). It
has been demonstrated that miRNA-429 induces the apoptosis of
glioblastoma cells via B-cell lymphoma 2 (Bcl-2), while the
suppression of miRNA-429 promotes Bcl-2-mediated cancer cell
survival (57). Other studies
indicated that c-Myc was a critical target gene of miRNA-429, and
that miRNA-429 significantly downregulated the expression of c-myc
to control the growth of GC cells (26). Additionally, miRNA-429 was
demonstrated to not only inhibit GC cell growth, but to regulate GC
cell invasiveness through zinc finger E-box-binding homeobox
protein (58). miRNA-148a and
miRNA-148b play important roles, functioning as tumor suppressors
in GC. However, downregulation of miRNA-148a contributes to GC
lymph node-metastasis and progression, while upregulation of
miRNA-148b suppresses GC cell growth. These two miRNAs are
therefore potential therapeutic target candidates (37,59).
However, in the study, it was revealed that after treatment with
soft-shelled turtle peptide, these suppressor function-associated
miRNAs whose expression was restrained in GC cells were
significantly restored to an upregulated state. This indicated that
the soft-shelled turtle peptide not only improves immunity to
contend against cancer cells indirectly in vivo, but that it
also has the capacity to block GC cell growth directly through
regulation of associated miRNAs and target genes. Furthermore,
certain miRNAs, including miRNA-375, could be detected in plasma
and may become potential biomarkers for the dynamic monitoring and
evaluation of cancer prognosis (20,60).
Olfactory receptor neurons express olfactory
receptor gene to detect odor molecules and transport the action
potential to the olfactory bulb of the brain. Olfactory receptors
are G protein-coupled receptors that are mainly expressed in the
olfactory epithelium to detect odorants (61). However, besides olfactory tissues,
non-olfactory tissues have also recently been demonstrated to
exhibit the expression of olfactory signaling components (olfactory
receptor, olfactory G-protein, adenylate cyclase III and olfactory
marker protein) (62,63). The role of olfactory signaling
components in non-olfactory tissues has not been clarified.
Nevertheless, it has been demonstrated that olfactory receptors are
not only involved in monitoring extracellular chemical cues, but
that they are also frequently overexpressed in various types of
cancer cells (64,65). In the present study, the GO analysis
results indicated that target genes of soft-shelled turtle
peptide-specific miRNAs were involved in a number of essential
biological processes. Moreover, two biological processes, namely
detection of chemical stimuli involved in the sensory perception of
smell and the G-protein coupled receptor signaling pathway, were
closely associated with the target genes. The role of olfactory
receptor-associated events in GC has not been reported. In
pancreatic cancer cells, the involvement of PI3 kinase γ-dependent
signaling pathway in the promotion of tumor cell invasiveness could
be triggered by olfactory receptor stimulation (64). Hence, the tight association between
olfactory receptors and soft-shelled turtle peptide-specific
miRNA-gene indicated that soft-shelled turtle peptide may influence
the abilities of growth, metastasis and invasion in cancer cells
through the regulation of olfactory receptor-associated events.
In the present study, KEGG analysis revealed that
the target genes were associated with several cancer-related
pathways. Particularly, the Hippo signaling pathway and the FoxO
signaling pathway, which function as tumor suppressors, may be
regulated by soft-shelled turtle peptide. The major functions of
the Hippo signaling pathway have been defined as restricting cell
growth and modulating cell proliferation and differentiation in
developing organs (66,67). Transcriptional coactivator
YAP1/telomere length regulator Taz1 (YAP1/TAZ) are the most
important effectors and are negatively regulated by the Hippo
pathway through phosphorylation-dependent and -independent
mechanisms. The upregulation of YAP1 and TAZ induces
epithelial-mesenchymal transition and increases drug resistance in
cancer cells (68–70). Deregulation of the Hippo signaling
pathway is involved in the initiation, progression and metastasis
of a number of cancer types (71–73),
including GC (74). Hence, the Hippo
pathway has been speculated to be a drug target inhibitor of YAP1
and TAZ in cancer therapy (75,76).
Additionally, miRNA-375 has been identified to target the Hippo
pathway effector YAP to inhibit the proliferation and invasion of
liver cancer cells (77). Therefore,
we hypothesize that soft-shelled turtle peptide may exert
anticancer functions through the miRNA-375-Hippo pathway. FoxO
factors not only play an anticancer role in various types of tumors
(78,79), but also contribute to extreme
longevity and life span (80).
miRNA-22 forms a regulatory loop to fine-tune the dynamics of the
phosphatase and tensin homolog/AKT/FoxO1 pathway (81). miRNA-30d induces apoptosis and is
regulated by the AKT/FoxO pathway in renal cell carcinoma, and is
formed as the AKT/FoxO/miR-30d/metastasis adhesion protein
signaling transduction pathway (82).
In conclusion, the microarray results of the present
study indicated that the soft-shelled turtle peptide has the
potential function to influence cancer-related miRNAs and pathways.
However, the anticancer properties of soft-shelled turtle peptide
in GC require further validation in vitro and in
vivo.
This study was supported in part by Special Grants
from the Zhejiang Agricultural Group Co., Ltd., Scientific Research
Grants in Traditional Chinese Medicine, Zhejiang Province of China
(no. 2012ZB018), Research Grants from the National Natural Science
Foundation of China (nos. 81372332 and 81572822) and Key Projects
from the Zhejiang Provincial Natural Science Foundation of China
(no. LZ13H160002 and LZ18H160002), and was partly sponsored by
Zhejiang Provincial Program for the Cultivation of High-level
Innovative Health talents (no. Zjwjw2014-108) and the Major
Training Personnel from Zhejiang Provincial Program for the
Training and Development Project for 151 talents (no.
Zjhrss2014-150).
|
1
|
Toyota M, Suzuki H, Yamamoto E, Yamano H,
Imai K and Shinomura Y: Integrated analysis of genetic and
epigenetic alterations in cancer. Epigenomics. 1:291–299. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gerhauser C: Cancer chemoprevention and
nutriepigenetics: State of the art and future challenges. Top Curr
Chem. 329:73–132. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lu XX, Yu JL, Ying LS, Han J, Wang S, Yu
QM, Wang XB, Fang XH and Ling ZQ: Stepwise cumulation of RUNX3
methylation mediated by Helicobacter pylori infection contributes
to gastric carcinoma progression. Cancer. 118:5507–5517. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Fang MZ, Wang Y, Ai N, Hou Z, Sun Y, Lu H,
Welsh W and Yang CS: Tea polyphenol (−)-epigallocatechin-3-gallate
inhibits DNA methyltransferase and reactivates methylation-silenced
genes in cancer cell lines. Cancer Res. 63:7563–7570.
2003.PubMed/NCBI
|
|
5
|
Nandakumar V, Vaid M and Katiyar SK:
(−)-Epigallocatechin-3-gallate reactivates silenced tumor
suppressor genes, Cip1/p21 and p16INK4a, by reducing DNA
methylation and increasing histones acetylation in human skin
cancer cells. Carcinogenesis. 32:537–544. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yan H, Zhu Y, Liu B, Wu H, Li Y, Wu X,
Zhou Q and Xu K: Mitogen-activated protein kinase mediates the
apoptosis of highly metastatic human non-small cell lung cancer
cells induced by isothiocyanates. Br J Nutr. 106:1779–1791. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lawson AP, Long MJ, Coffey RT, Qian Y,
Weerapana E, El Oualid F and Hedstrom L: Naturally occurring
isothiocyanates exert anticancer effects by inhibiting
deubiquitinating enzymes. Cancer Res. 75:5130–5142. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Guan F, Ding Y, Zhang Y, Zhou Y, Li M and
Wang C: Curcumin suppresses proliferation and migration of
MDA-MB-231 breast cancer cells through autophagy-dependent Akt
degradation. PLoS One. 11:e01465532016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chhabria SV, Akbarsha MA, Li AP, Kharkar
PS and Desai KB: In situ allicin generation using targeted
alliinase delivery for inhibition of MIA PaCa-2 cells via
epigenetic changes, oxidative stress and cyclin-dependent kinase
inhibitor (CDKI) expression. Apoptosis. 20:1388–1409. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sinha D, Sarkar N, Biswas J and Bishayee
A: Resveratrol for breast cancer prevention and therapy:
Preclinical evidence and molecular mechanisms. Semin Cancer Biol.
40-41:1–232. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Park JS, Khoi PN, Joo YE, Lee YH, Lang SA,
Stoeltzing O and Jung YD: EGCG inhibits recepteur dorigine nantais
expression by suppressing Egr-1 in gastric cancer cells. Int J
Oncol. 42:1120–1126. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang H, Iwasaki M, Haiman CA, Kono S,
Wilkens LR, Keku TO, Berndt SI, Tsugane S and Le Marchand L:
Interaction between red meat intake and NAT2 genotype in increasing
the risk of colorectal cancer in Japanese and African Americans.
PLoS One. 10:e01449552015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Taunk P, Hecht E and Stolzenberg-Solomon
R: Are meat and heme iron intake associated with pancreatic cancer?
Results from the NIH-AARP Diet and Health Cohort. Int J Cancer.
138:2172–2189. 2015. View Article : Google Scholar
|
|
14
|
Lippi G, Mattiuzzi C and Cervellin G: Meat
consumption and cancer risk: A critical review of published
meta-analyses. Crit Rev Oncol Hematol. 97:1–14. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhao CG and Shen YM: Anti-cancer effect
and edible methods of soft-shelled turtle. China Agricultural
Publishing House; Beijing: 2008
|
|
16
|
Fu JJ, Tan SL, Li YG, Lv H, Zhu WF and Liu
HN: Adjuvant effects of snapping turtle co-peptide (STCP) on
radiotherapy for cancer. J Jiangxi Uni Trad Chin Med. 27:68–71.
2015.(In Chinese).
|
|
17
|
Huang da W, Sherman BT and Lempicki RA:
Systematic and integrative analysis of large gene lists using DAVID
bioinformatics resources. Nat Protoc. 4:44–57. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Huang da W, Sherman BT and Lempicki RA:
Bioinformatics enrichment tools: Paths toward the comprehensive
functional analysis of large gene lists. Nucleic Acids Res.
37:1–13. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Juzėnas S, Saltenienė V, Kupcinskas J,
Link A, Kiudelis G, Jonaitis L, Jarmalaite S, Kupcinskas L,
Malfertheiner P and Skieceviciene J: Analysis of deregulated
microRNAs and their target genes in gastric cancer. PLoS One.
10:e01323272015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ding L, Xu Y, Zhang W, Deng Y, Si M, Du Y,
Yao H, Liu X, Ke Y, Si J and Zhou T: miR-375 frequently
downregulated in gastric cancer inhibits cell proliferation by
targeting JAK2. Cell Res. 20:784–793. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu Y, Xing R, Zhang X, Dong W, Zhang J,
Yan Z, Li W, Cui J and Lu Y: miR-375 targets the p53 gene to
regulate cellular response to ionizing radiation and etoposide in
gastric cancer cells. DNA Repair (Amst). 12:741–750. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shiotani A, Murao T, Kimura Y, Matsumoto
H, Kamada T, Kusunoki H, Inoue K, Uedo N, Iishi H and Haruma K:
Identification of serum miRNAs as novel non-invasive biomarkers for
detection of high risk for early gastric cancer. Br J Cancer.
109:2323–2330. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cui H, Wang L, Gong P, Zhao C, Zhang S,
Zhang K, Zhou R, Zhao Z and Fan H: Deregulation between miR-29b/c
and DNMT3A is associated with epigenetic silencing of the CDH1
gene, affecting cell migration and invasion in gastric cancer. PLoS
One. 10:e01239262015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lang N, Liu M, Tang QL, Chen X, Liu Z and
Bi F: Effects of microRNA-29 family members on proliferation and
invasion of gastric cancer cell lines. Chin J Cancer. 29:603–610.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sun T, Wang C, Xing J and Wu D: miR-429
modulates the expression of c-myc in human gastric carcinoma cells.
Eur J Cancer. 47:2552–2559. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liu D, Xia P, Diao D, Cheng Y, Zhang H,
Yuan D, Huang C and Dang C: miRNA-429 suppresses the growth of
gastric cancer cells in vitro. J Biomed Res. 26:389–393. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ning X, Shi Z, Liu X, Zhang A, Han L,
Jiang K, Kang C and Zhang Q: DNMT1 and EZH2 mediated methylation
silences the microRNA-200b/a/429 gene and promotes tumor
progression. Cancer Lett. 359:198–205. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhu P, Zhang J, Zhu J, Shi J, Zhu Q and
Gao Y: MiR-429 induces gastric carcinoma cell apoptosis through
Bcl-2. Cell Physiol Biochem. 37:1572–1580. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sakamoto N, Naito Y, Oue N, Sentani K,
Uraoka N, Zarni Oo H, Yanagihara K, Aoyagi K, Sasaki H and Yasui W:
MicroRNA-148a is downregulated in gastric cancer, targets MMP7, and
indicates tumor invasiveness and poor prognosis. Cancer Sci.
105:236–243. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zuo J, Xia J, Ju F, Yan J, Zhu A, Jin S,
Shan T and Zhou H: MicroRNA-148a can regulate runt-related
transcription factor 3 gene expression via modulation of DNA
methyltransferase 1 in gastric cancer. Mol Cells. 35:313–319. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kim SY, Jeon TY, Choi CI, Kim DH, Kim DH,
Kim GH, Ryu DY, Lee BE and Kim HH: Validation of circulating miRNA
biomarkers for predicting lymph node metastasis in gastric cancer.
J Mol Diagn. 15:661–669. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zuo QF, Cao LY, Yu T, Gong L, Wang LN,
Zhao YL, Xiao B and Zou QM: MicroRNA-22 inhibits tumor growth and
metastasis in gastric cancer by directly targeting MMP14 and Snail.
Cell Death Dis. 6:e20002015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tang Y, Liu X, Su B, Zhang Z, Zeng X, Lei
Y, Shan J, Wu Y, Tang H and Su Q: microRNA-22 acts as a metastasis
suppressor by targeting metadherin in gastric cancer. Mol Med Rep.
11:454–460. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wang X, Yu H, Lu X, Zhang P, Wang M and Hu
Y: miR-22 suppresses the proliferation and invasion of gastric
cancer cells by inhibiting CD151. Biochem Biophys Res Commun.
445:175–179. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Sugihara H, Ishimoto T, Watanabe M,
Sawayama H, Iwatsuki M, Baba Y, Komohara Y, Takeya M and Baba H:
Identification of miR-30e* regulation of Bmi1 expression mediated
by tumor-associated macrophages in gastrointestinal cancer. PLoS
One. 8:e818392013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Song YX, Yue ZY, Wang ZN, Xu YY, Luo Y, Xu
HM, Zhang X, Jiang L, Xing CZ and Zhang Y: MicroRNA-148b is
frequently down-regulated in gastric cancer and acts as a tumor
suppressor by inhibiting cell proliferation. Mol Cancer. 10:12011.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yang B, Huang J, Liu H, Guo W and Li G:
miR-335 directly, while miR-34a indirectly modulate survivin
expression and regulate growth, apoptosis, and invasion of gastric
cancer cells. Tumour Biol. 37:1771–1779. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Peng Y, Guo JJ, Liu YM and Wu XL:
MicroRNA-34A inhibits the growth, invasion and metastasis of
gastric cancer by targeting PDGFR and MET expression. Biosci Rep.
34:e001122014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Cao W, Fan R, Wang L, Cheng S, Li H, Jiang
J, Geng M, Jin Y and Wu Y: Expression and regulatory function of
miRNA-34a in targeting survivin in gastric cancer cells. Tumour
Biol. 34:963–971. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
He M, Gao L, Zhang S, Tao L, Wang J, Yang
J and Zhu M: Prognostic significance of miR-34a and its target
proteins of FOXP1, p53, and BCL2 in gastric MALT lymphoma and
DLBCL. Gastric Cancer. 17:431–441. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Yin WZ, Li F, Zhang L, Ren XP, Zhang N and
Wen JF: Down-regulation of microRNA-205 promotes gastric cancer
cell proliferation. Eur Rev Med Pharmacol Sci. 18:1027–1032.
2014.PubMed/NCBI
|
|
43
|
Chen L, Lü MH, Zhang D, Hao NB, Fan YH, Wu
YY, Wang SM, Xie R, Fang DC, Zhang H, et al: miR-1207-5p and
miR-1266 suppress gastric cancer growth and invasion by targeting
telomerase reverse transcriptase. Cell Death Dis. 5:e10342014.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Xu YJ and Fan Y: miR-215/192 participates
in gastric cancer progression. Clin Transl Oncol. 17:34–40. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Jin Z, Selaru FM, Cheng Y, Kan T, Agarwal
R, Mori Y, Olaru AV, Yang J, David S, Hamilton JP, et al:
MicroRNA-192 and −215 are upregulated in human gastric cancer in
vivo and suppress ALCAM expression in vitro. Oncogene.
30:1577–1585. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Chen Q, Ge X, Zhang Y, Xia H, Yuan D, Tang
Q, Chen L, Pang X, Leng W and Bi F: Plasma miR-122 and miR-192 as
potential novel biomarkers for the early detection of distant
metastasis of gastric cancer. Oncol Rep. 31:1863–1870. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Wen X, Wu JQ, Peng W, Feng JF and Tang JH:
MicroRNA-377 predicts poor clinical outcome of gastric cancer and
induces tumorigenesis by targeting multiple tumor-suppressor genes.
Oncol Rep. 34:203–210. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Chen J, Sun D, Chu H, Gong Z, Zhang C,
Gong B, Li Y, Li N and Jiang L: Screening of differential microRNA
expression in gastric signet ring cell carcinoma and gastric
adenocarcinoma and target gene prediction. Oncol Rep. 33:2963–2971.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Link A, Balaguer F and Goel A: Cancer
chemoprevention by dietary polyphenols: Promising role for
epigenetics. Biochem Pharmacol. 80:1771–1792. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Avissar M, Christensen BC, Kelsey KT and
Marsit CJ: MicroRNA expression ratio is predictive of head and neck
squamous cell carcinoma. Clin Cancer Res. 15:2850–2855. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Mathé EA, Nguyen GH, Bowman ED, Zhao Y,
Budhu A, Schetter AJ, Braun R, Reimers M, Kumamoto K, Hughes D, et
al: MicroRNA expression in squamous cell carcinoma and
adenocarcinoma of the esophagus: Associations with survival. Clin
Cancer Res. 15:6192–6200. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Bhatti I, Lee A, James V, Hall RI, Lund
JN, Tufarelli C, Lobo DN and Larvin M: Knockdown of microRNA-21
inhibits proliferation and increases cell death by targeting
programmed cell death 4 (PDCD4) in pancreatic ductal
adenocarcinoma. J Gastrointest Surg. 15:199–208. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Ladeiro Y, Couchy G, Balabaud C,
Bioulac-Sage P, Pelletier L, Rebouissou S and Zucman-Rossi J:
MicroRNA profiling in hepatocellular tumors is associated with
clinical features and oncogene/tumor suppressor gene mutations.
Hepatology. 47:1955–1963. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Xu Y, Jin J, Liu Y, Huang Z, Deng Y, You
T, Zhou T, Si J and Zhuo W: Snail-regulated MiR-375 inhibits
migration and invasion of gastric cancer cells by targeting JAK2.
PLoS One. 9:e995162014. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Chang H, Kim N, Park JH, Nam RH, Choi YJ,
Lee HS, Yoon H, Shin CM, Park YS, Kim JM and Lee DH: Different
microRNA expression levels in gastric cancer depending on
Helicobacter pylori infection. Gut Liver. 9:188–196. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Chen D, Li Y, Li Y, Jin L, Su Z, Yu Z,
Yang S, Mao X and Lai Y: Tumor suppressive microRNA429 regulates
cellular function by targeting VEGF in clear cell renal cell
carcinoma. Mol Med Rep. 13:1361–1366. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Zhang Z, Zhou Q, Miao Y, Tian H, Li Y,
Feng X and Song X: MiR-429 induces apoptosis of glioblastoma cell
through Bcl-2. Tumour Biol. Oct 28–2015.(Epub ahead of print).
|
|
58
|
Liu W, An J, Li K and Hou H: miR-429
regulates gastric cancer cell invasiveness through ZEB proteins.
Tumour Biol. Oct 15–2015.(Epub ahead of print).
|
|
59
|
Zheng B, Liang L, Wang C, Huang S, Cao X,
Zha R, Liu L, Jia D, Tian Q, Wu J, et al: MicroRNA-148a suppresses
tumor cell invasion and metastasis by downregulating ROCK1 in
gastric cancer. Clin Cancer Res. 17:7574–7583. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Patel M, Verma A, Aslam I, Pringle H and
Singh B: Novel plasma microRNA biomarkers for the identification of
colitis-associated carcinoma. Lancet. 385 Suppl 1:S782015.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Touhara K, Sengoku S, Inaki K, Tsuboi A,
Hirono J, Sato T, Sakano H and Haga T: Functional identification
and reconstitution of an odorant receptor in single olfactory
neurons. Proc Natl Acad Sci USA. 96:pp. 4040–4045. 1999; View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Feldmesser E, Olender T, Khen M, Yanai I,
Ophir R and Lancet D: Widespread ectopic expression of olfactory
receptor genes. BMC Genomics. 7:1212006. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Kang N, Kim H, Jae Y, Lee N, Ku CR,
Margolis F, Lee EJ, Bahk YY, Kim MS and Koo J: Olfactory marker
protein expression is an indicator of olfactory receptor-associated
events in non-olfactory tissues. PLoS One. 10:e01160972015.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Sanz G, Leray I, Dewaele A, Sobilo J,
Lerondel S, Bouet S, Grébert D, Monnerie R, Pajot-Augy E and Mir
LM: Promotion of cancer cell invasiveness and metastasis emergence
caused by olfactory receptor stimulation. PLoS One. 9:e851102014.
View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Gelis L, Jovancevic N, Bechara FG, Neuhaus
EM and Hatt H: Functional expression of olfactory receptors in
human primary melanoma and melanoma metastasis. Exp Dermatol.
26:569–576. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Zeng Q and Hong W: The emerging role of
the Hippo pathway in cell contact inhibition, organ size control,
and cancer development in mammals. Cancer Cell. 13:188–192. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Meng Z, Moroishi T and Guan KL: Mechanisms
of Hippo pathway regulation. Genes Dev. 30:1–17. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Zhao Y and Yang X: The Hippo pathway in
chemotherapeutic drug resistance. Int J Cancer. 137:2767–2773.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Xie D, Cui J, Xia T, Jia Z, Wang L, Wei W,
Zhu A, Gao Y, Xie K and Quan M: Hippo transducer TAZ promotes
epithelial mesenchymal transition and supports pancreatic cancer
progression. Oncotarget. 6:35949–35963. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Li Z, Wang Y, Zhu Y, Yuan C, Wang D, Zhang
W, Qi B, Qiu J, Song X, Ye J, et al: The Hippo transducer TAZ
promotes epithelial to mesenchymal transition and cancer stem cell
maintenance in oral cancer. Mol Oncol. 9:1091–1105. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Yagi H, Asanoma K, Ohgami T, Ichinoe A,
Sonoda K and Kato K: GEP oncogene promotes cell proliferation
through YAP activation in ovarian cancer. Oncogene. 35:4471–4480.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Lamar JM, Stern P, Liu H, Schindler JW,
Jiang ZG and Hynes RO: The Hippo pathway target, YAP, promotes
metastasis through its TEAD-interaction domain. Proc Natl Acad Sci
USA. 109:pp. E2441–E2450. 2012; View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Wang G, Lu X, Dey P, Deng P, Wu CC, Jiang
S, Fang Z, Zhao K, Konaparthi R, Hua S, et al: Targeting
YAP-dependent MDSC infiltration impairs tumor progression. Cancer
Dis. 6:80–95. 2016. View Article : Google Scholar
|
|
74
|
Zhou GX, Li XY, Zhang Q, Zhao K, Zhang CP,
Xue CH, Yang K and Tian ZB: Effects of the hippo signaling pathway
in human gastric cancer. Asian Pac J Cancer Prev. 14:5199–5205.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Perra A, Kowalik MA, Ghiso E,
Ledda-Columbano GM, Di Tommaso L, Angioni MM, Raschioni C, Testore
E, Roncalli M, Giordano S and Columbano A: YAP activation is an
early event and a potential therapeutic target in liver cancer
development. J Hepatol. 61:1088–1096. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Santucci M, Vignudelli T, Ferrari S, Mor
M, Scalvini L, Bolognesi ML, Uliassi E and Costi MP: The Hippo
pathway and YAP/TAZ-TEAD protein-protein interaction as targets for
regenerative medicine and cancer treatment. J Med Chem.
58:4857–4873. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Liu AM, Poon RT and Luk JM: MicroRNA-375
targets Hippo-signaling effector YAP in liver cancer and inhibits
tumor properties. Biochem Biophys Res Commun. 394:623–627. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Guan H, Tan P, Xie L, Mi B, Fang Z, Li J,
Yue J, Liao H and Li F: FOXO1 inhibits osteosarcoma oncogenesis via
Wnt/β-catenin pathway suppression. Oncogenesis. 4:e1662015.
View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Bullock M: FOXO factors and breast cancer:
Outfoxing endocrine resistance. Endocr Relat Cancer. 23:R113–R130.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Martins R, Lithgow GJ and Link W: Long
live FOXO: Unraveling the role of FOXO proteins in aging and
longevity. Aging Cell. 15:196–207. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Bar N and Dikstein R: miR-22 forms a
regulatory loop in PTEN/AKT pathway and modulates signaling
kinetics. PloS One. 5:e108592010. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Wu C, Jin B, Chen L, Zhuo D, Zhang Z, Gong
K and Mao Z: MiR-30d induces apoptosis and is regulated by the
Akt/FOXO pathway in renal cell carcinoma. Cell Signal.
25:1212–1221. 2013. View Article : Google Scholar : PubMed/NCBI
|