Introduction
Cancer is one of the most malignant diseases
worldwide. Numerous signaling pathways have been identified
involved in the development of severe cancers. Abnormal
transcriptional programs have been reported playing important roles
in the generation and development of malignant carcinoma (1). Expression profiling using cDNA
microarray on primary samples extracted from patients, as well as
established mouse models, has revealed hundreds of transcriptional
factors that are dysregulated in various types of cancers,
including PU.1 (2).
PU.1 is a hematopoietic transcription factor, known
for its role in myeloid development (3). PU.1 depletion is lethal for animal
during embryonic development due to hematopoietic failure, with
abnormal lymphoid and myeloid cell lineages (3–5). It has
also been implicated in tumor progression, especially in leukemia
(2). Strong PU.1 expression were
shown associated with longer survival in follicular lymphoma
(6). Deletion and downregulation of
PU.1 were also found in human acute myeloid leukemia (AML) and
indolent types of lymphomas (7,8). On the
contrary, surprising results were reported in a recent study by
Zhou et al showing that PU.1 is required and works as an
essential regulator for the development of mixed lineage leukemia
(MLL) (9), one of the aggressive
forms of AMLs with poor prognosis (10,11). The
molecular mechanism for PU.1 in tumor progression has not been
fully identified. Some study indicated that PU.1 could contribute
to leukemia through transcriptional activation of leukemia fusion
proteins (9). It is also reported to
promote cytokine production as well as monocytes activation
(12), indicating PU.1 could affect
tumor severity through immune responses. Although PU.1 play
contradictory roles in different types of leukemia, whether PU.1 is
involved in other types of cancers is not known.
Glioma is a devastating brain cancer (13). Patients diagnosed with glioblastoma
multiforme (GBM) have low survival rate and high morbidity, despite
all kinds of medical treatments (14). Although tumors in the brain suffer
many restrictions for clinical therapy, such as the blood brain
barrier, similar to other solid tumors, the immune cells, i.e.,
microglia in the brain, are attracted to the GBM tissues and
support tumor progression (15–19). Thus
recently studies have been done focusing on glioma-associated
immune cells for disease intervention (20,21).
Although PU.1 is important for immune cells in the circulatory
system, its role in the central nervous system, especially in
glioma, is unknown.
Here we examined the clinical relevance of the PU.1
in patients with different kinds of gliomas, and found that the
expression of genes coding PU.1 is significantly increased in
glioma patient samples. We also aim to explore potential factors
involved in PU.1 signaling and glioma. Using the online available
tool to search for the genotype-phenotype associations between PU.1
and glioma, we found that several genes could be the targets of
PU.1 in mediating tumor severity. Our results provide initiative
evidence that PU.1 could contributes to the progress of glioma,
probably via its targeting gene signatures.
Materials and methods
GEO datasets and statistical
analysis
Publicly available gene-expression datasets of
glioma patients were downloaded from NCBI-GEO with accession
numbers GSE4290 and GSE15824 (http://www.ncbi.nlm.nih.gov/geo/). Statistical
analysis for relative gene-expression data were performed using
DESeq2 version:v1.4.5. The P-values were calculated using the
Benjamini and Hochberg method in the R DESeq2 package.
Immunostaining
Biospecimens were obtained from patients from the
Shanghai First People's Hospital, according to their surgical
pathology reports and clinical records. The study was approved by
the Ethics Committee of the Shanghai First People's Hospigal,
Shanghai Jiaotong University. All subjects agreed to donate their
tissue for this study. Tissues were fixed in formalin. Slicing and
immunohistochemistry were performed according to standard protocol
in a previous study (22). Antibodies
used are: PU.1, 9G7 Rb IgG, 1:100, Novex; CD68, FA11 Rt IgG, 1:200,
Serotec.
Genotype-phenotype association
analysis
Analysis was performed using an online system to
search for genotype-phenotype associations (http://literome.azurewebsites.net/). Key words used to
search for genotype-phenotype associations are ‘SPI1’ and
‘glioma’.
Target gene identification
Online available database were applied to identify
transcription targets of PU.1 (https://cb.utdallas.edu/cgi-bin/TRED/tred.cgi?process=searchTFGeneForm).
90 genes were identified as potential targets of PU.1.
Biological pathway analysis
Biological pathway analysis was performed using an
online available tool, the Reactome database (http://www.reactome.org/). Multiple biological
pathways in Homo sapiens were showed linked to the network
containing genes of interest.
Results
To examine the clinical relevance of PU.1 expression
in glioma patients, we performed analysis on SPI1 (gene that
encode PU.1) expression using publicly available RNA-sequencing
array data. Each dataset can be classified as four groups:
Non-tumor sample, astrocytoma patient sample, glioblastoma patient
sample and oligodendrioglioma patient sample. Analysis was
performed as control vs. disease conditions. Non-tumor samples were
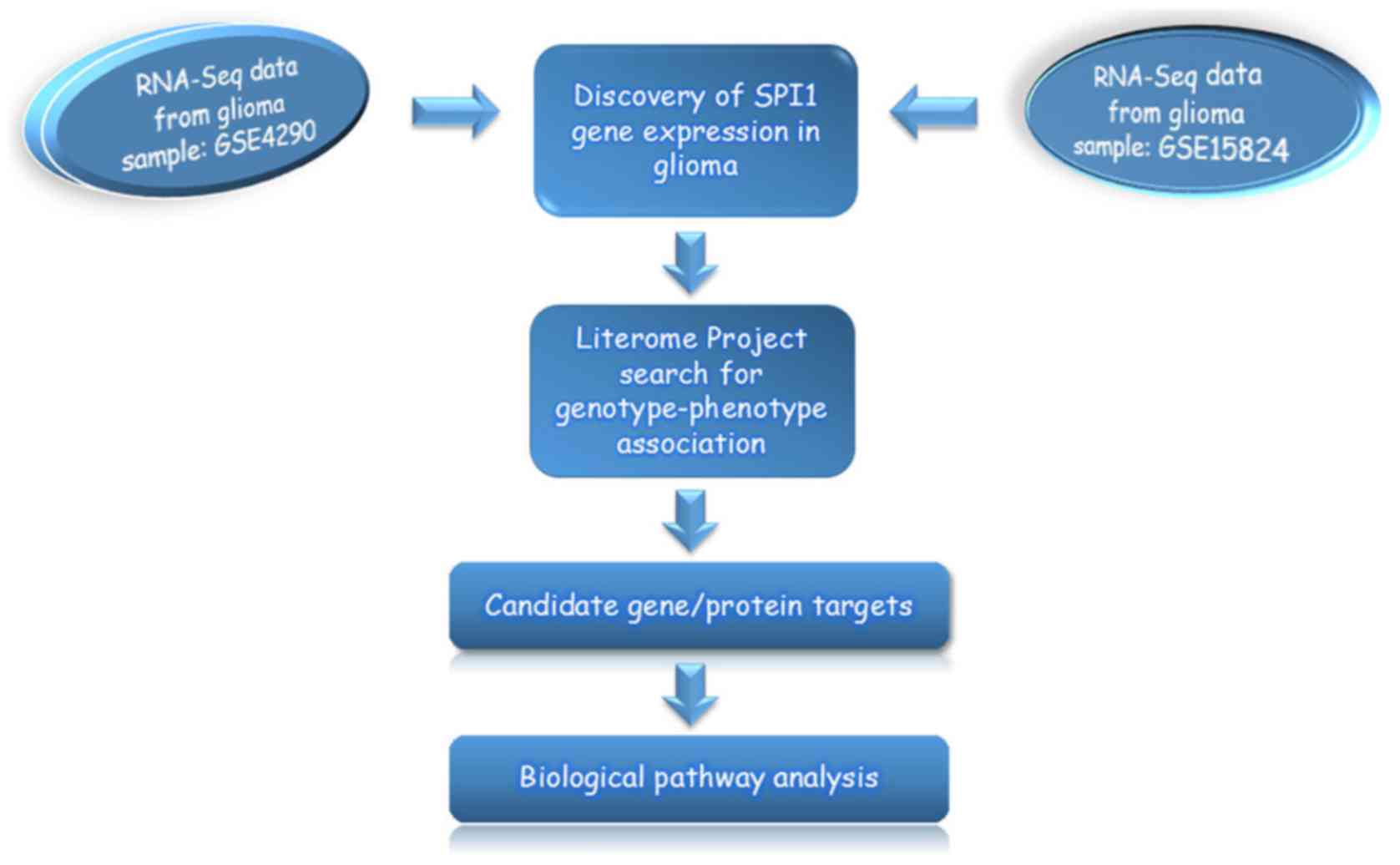
shared for comparisons in each dataset. Fig. 1 detailed the analysis strategy
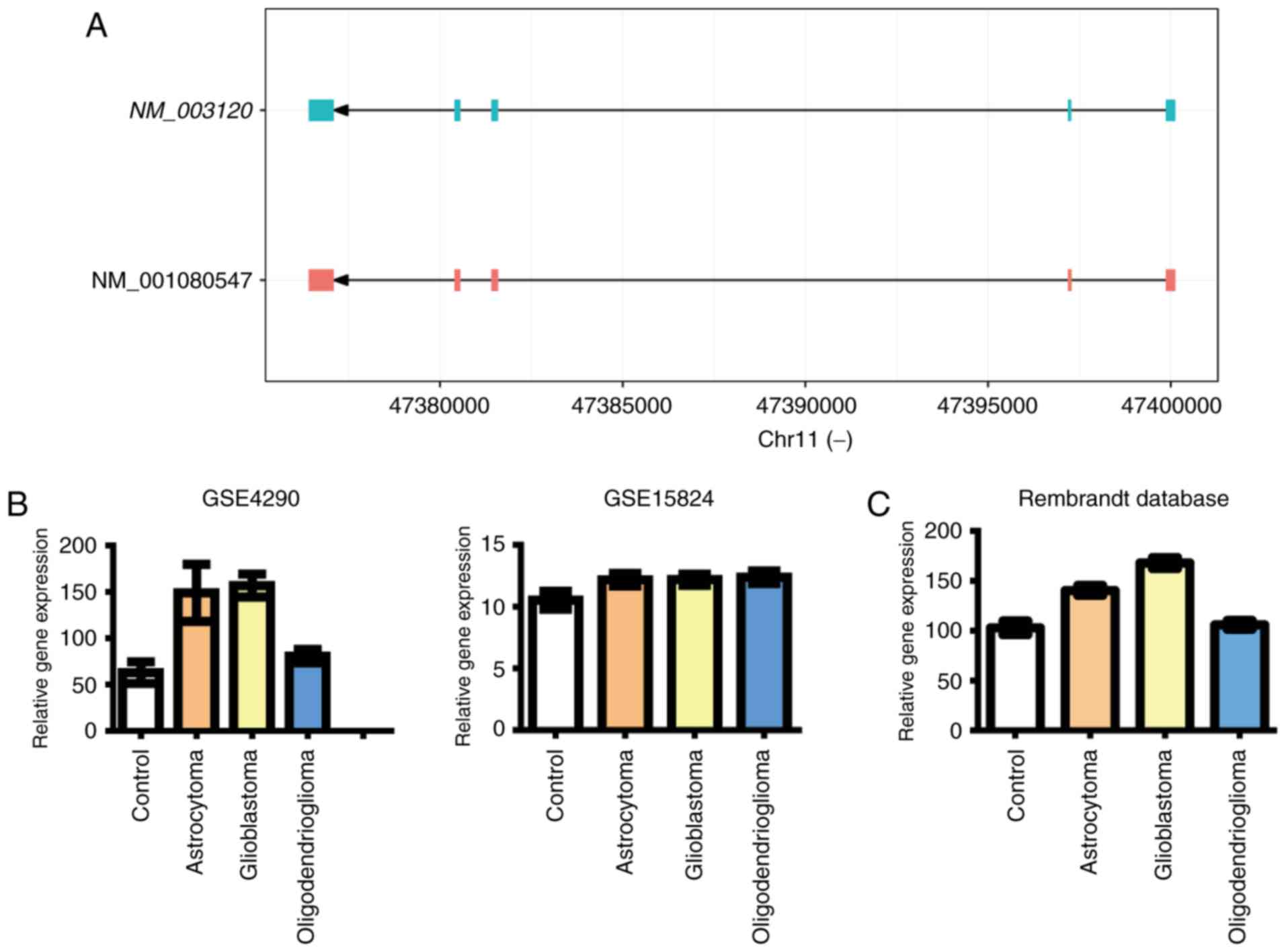
included in this study. The position of gene SPI1 in genome
of Homo sapiens was shown in Fig.
2A. As shown in Fig. 2B, we found
significant increases of SPI1 mRNA expression in several
kinds of glioma samples, while there is trend of upregulated
SPI1 expression in all glioma samples, including
astrocytoma, glioblastoma and oligodendrioglioma.
We next explored such correlation in another public
database, the REMBRANDT database (http://www.betastasis.com/glioma/rembrandt/). We found
that SPI1 expression levels were significantly increased in
patients with glioblastoma and astrocytoma (Fig. 2C). These results suggest clear
positive correlation between SPI1 expression and glioma, especially
glioblastoma and astrocytoma.
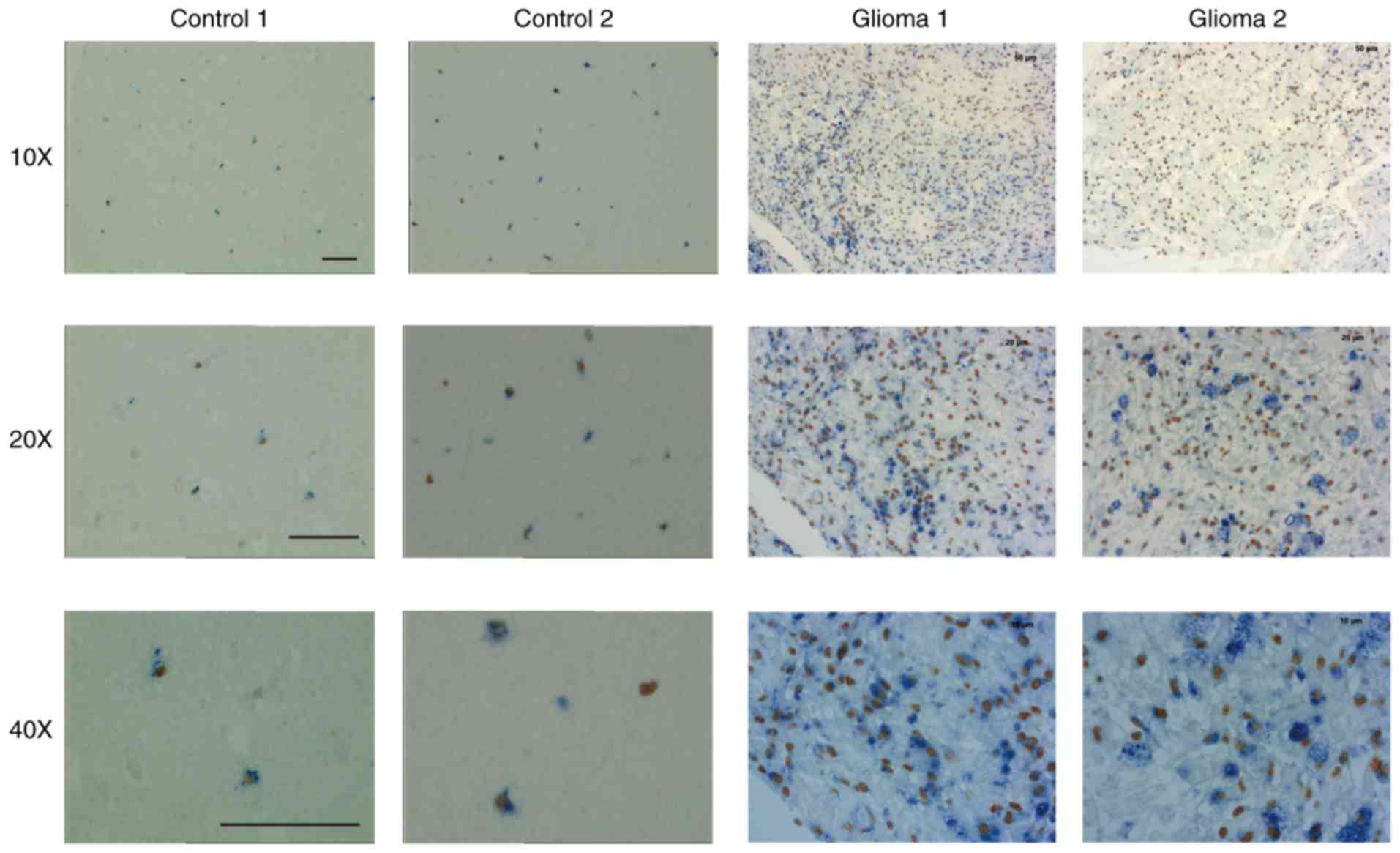
To further demonstrate the relevance between PU.1
expression and glioma malignancy, we tested PU.1 protein expression
in patient brain tissues. We found that PU.1 was highly expressed
in glioma patients, but not in non-glioma patients (Fig. 3). CD68 was stained as marker of
macrophage. Some of the PU.1 positive cells are colocalized with
the CD68+ cells.
We also searched TCGA database for further evidence.
Three genomic mutations have been identified in two glioma patients
at the SPI1 locus (23). These
mutations include: Chr11:g.47378343G>A, chr11:g.47359851A>T,
chr11:g.47375730C>A. Interestingly, we also noticed a negative
correlation between SPI1 expression and patient survival rate
(Kaplan-Meier estimator, P=0.0035) in the REMBRANDT database,
further demonstrating the involvement of PU.1 in glioma
malignance.
Next we are interested in what are the potential
mechanisms that PU.1 is involved in glioma. We applied analysis
using an online system to search for genotype-phenotype
associations (http://literome.azurewebsites.net/) (24,25). The
basic principle is to extract genomic and phenotypic knowledge from
PubMed articles thus providing possible signaling pathways for one
gene's function. Using this online available tool, we found
associations between SPI1 and glioma via 8 different genes,
including BTK, ERVK-6, FLI1, FURIN, GRAP2, IL1B, MYLIP and
SERPINB1. We further examined the viability of these associations
through literature studying, and identified that several genes
could be the potential targets of PU.1 in mediating glioma
severity. BTK is the Bruton's agammaglobulinemia tyrosine kinase,
known for its role in X-linked agammaglobulinemia. It is reported
that BTX gene contains PU.1 binding sites at its promoter region.
In PU.1−/− mice, BTK expression was significantly
reduced (26). Another important
potential regulator is FLI1, a transcription factor in the ETS
family. FLI1 transcription was regulated under SPI1 transfection
(27). PU.1 was also reported binding
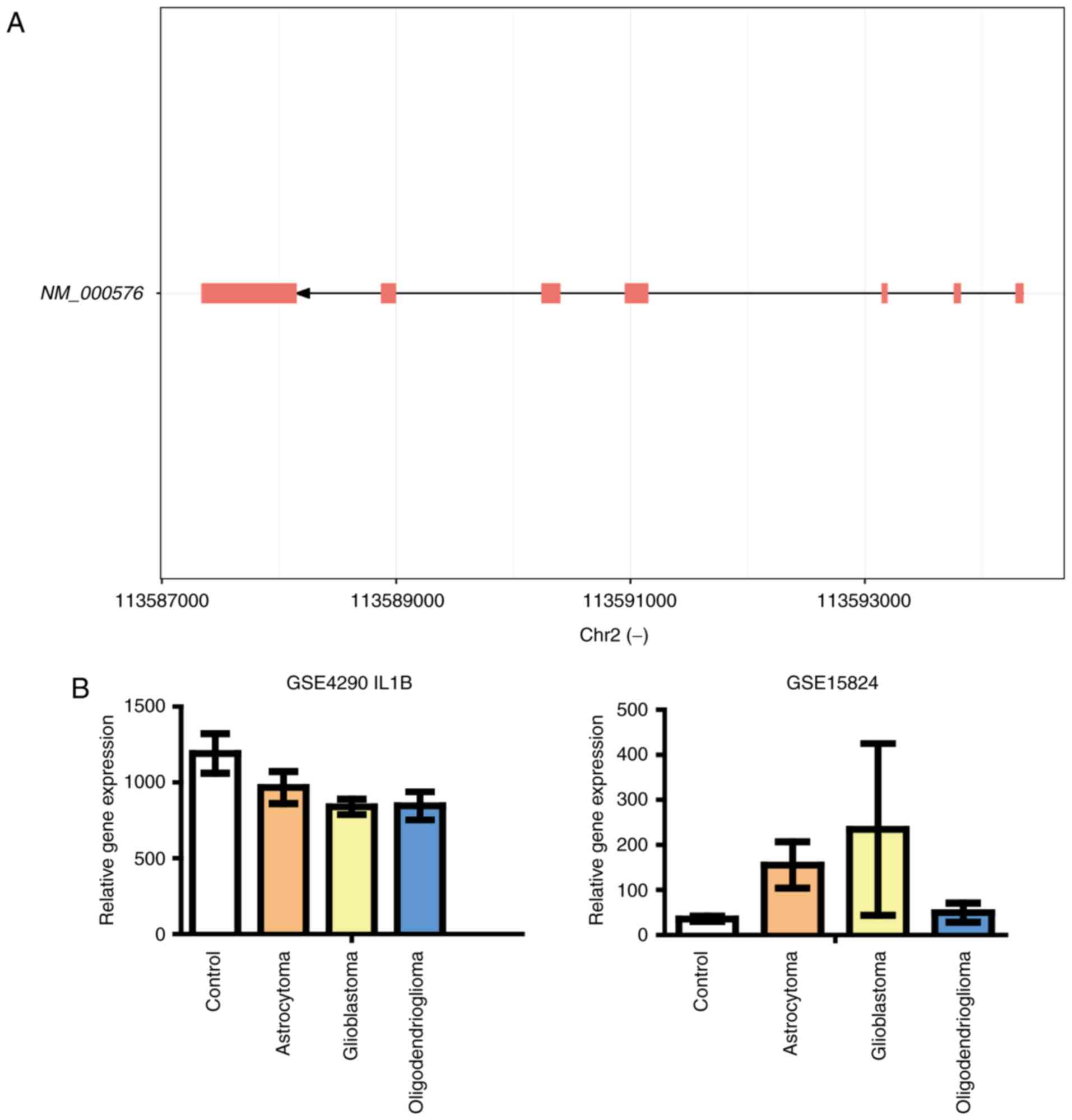
to the promoter of IL1B to activate its expression (12). These three candidates were all found
involved in the progress of glioma (28–30).
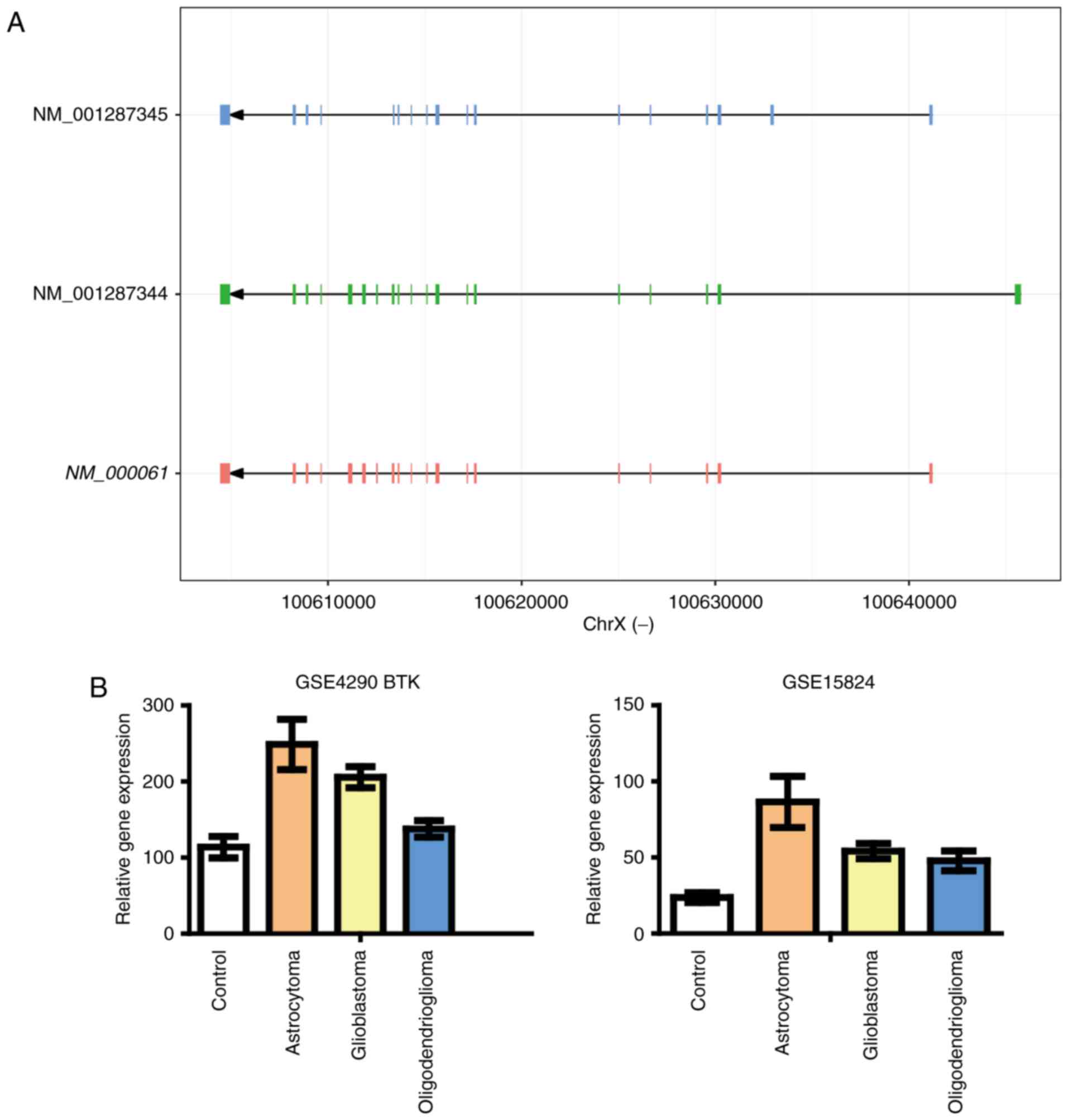
We analyzed the expression changes of the three
predicted genes in glioma RNA array data used for analysis of
SPI1 expression. As shown in Fig.
4, we found significant increases of BTK mRNA expression
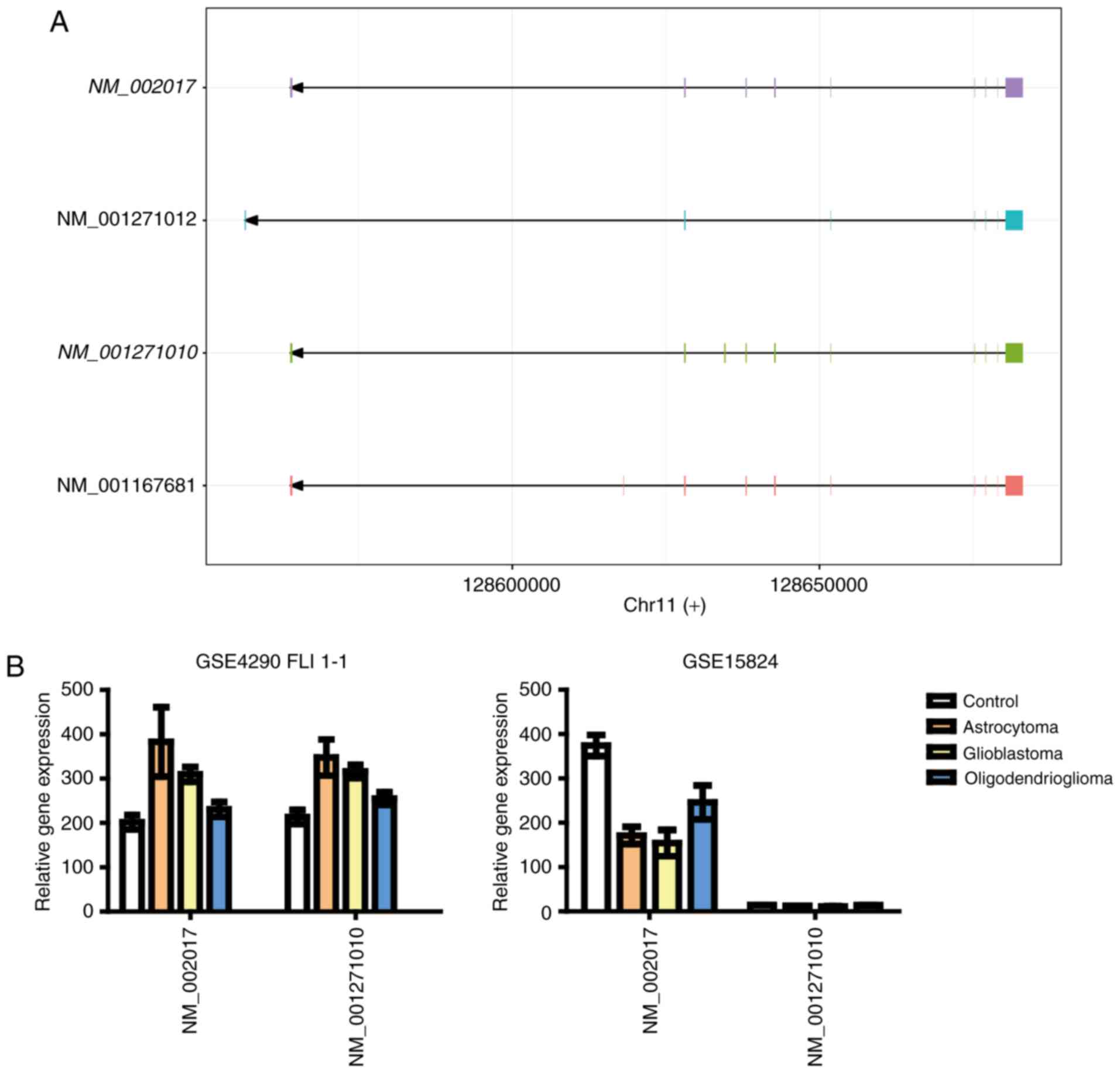
in both glioma datasets. On the other hand, FLI1 mRNA level
was increased in GSE4290 dataset, while decreased in the GSE15824
dataset (Fig. 5B). In terms of the
expression level of IL1B, it was decreased in the GSE4290
dataset, while increased in the GSE15824 dataset (Fig. 6). We plotted the fold changes of these
genes as bar graphs, as shown in Figs.
4–6, the expression pattern of
BTK is more comparable to that of the gene SPI1.
These results suggest that BTK is more probably an important factor
involved in the role of PU.1 in glioma progression.
Although only three genes were predicted involved in
the regulation of glioma by PU.1, PU.1 could target a number of
genes via transcriptional regulation. Taking advantage of an online
available tool, we identified 90 potential transcription targets of
PU.1 (https://cb.utdallas.edu/cgi-bin/TRED/tred.cgi?process=searchTFGeneForm).
We analyzed the expression levels of these genes in glioma samples
(data not shown). The results showed that genes, such as LTF, FSF1,
CD163 and MMP2, were consistently upregulated in glioma tissues.
Although there is no direct link for these factors and brain tumor,
indirect mechanisms/signaling pathways could be involved in the
role of PU.1 in glioma. In summary, our results indicate that PU.1
could contribute to the progress of glioma, probably via its
targeting gene signatures.
We next searched for biological pathways linked to
the PU.1-associated genes in glioma using Reactome database
(31,32). Three nodes were identified
representing PU.1-targeting pathways in glioma, including immune
system, signal transduction, and disease. Meanwhile, multiple
biological pathways were found associated with thus process,
including the immune system, signal transduction, disease, gene
expression, and metabolism of proteins.
In conclusion, our study identified significant
increase of PU.1 expression in human glioma patients. Through
genotype-phenotype association test and biological interaction
network building, our results indicate that PU.1 could affect the
development of glioma by targeting various genes in different
signaling pathways.
Discussion
Glioma is a kind of severe cancer starts in the
brain or spinal cord with low survival rate and high morbidity
(13,14). Although previous studies have
implicated a lot of signaling pathways involved in the progress of
glioma, until now there is no cure for the severe disease. Thus, it
is urgent to identify more changes in glioma condition for medical
treatment. However, it is still unknown whether PU.1, a
transcription factor known for its role in myeloid differentiation
and immune responses, is involved in the progress of glioma.
In this present study, we found significant increase
of SPI1, the gene coding transcription factor PU.1, in
glioma patient samples through analysis of RNA array dataset. We
hypothesized that PU.1 could function through its transcription
targets. 90 candidate genes were analyzed in this circumstance. We
provided several potential candidates that could mediate the role
of PU.1 in glioma through genotype-phenotype association analysis,
including BTX, FLI1 and IL1B. These genes were
found upregulated in glioma samples in different cases. Among the
potential targets of PU.1, FLI1 has been directly implicated
with glioma cell proliferation, migration and invasion (30). Fli1 and PU.1 are two members of the
ETS family of transcription factors. Previous study has reported
that FLI1 expression is upregulated under transfection of SPI1
vector via binding of its promoter region (27). In glioma cells, expression of the
dominant-negative form of Ets1 significantly inhibited cell
proliferation and migration, it is also the case for Fli1 (30). These results implicated that other ETS
family members maybe also capable of promoting glioma cell
proliferation, such as PU.1.
IL1B is a cytokine that belongs to the interleukin 1
family (33). Its precursor is
produced by immune cells and be cleaved by caspase-1 to form mature
IL1B (34). The cytokine is an
important factor mediating immune responses, and be involved in
various cellular activities, including cell proliferation,
differentiation and apoptosis (35).
Although the clear role of IL1B in glioma development has not been
identified, previous studies have shown that immune cells in the
brain can produce functional IL1B. Also, IL1B expression was
altered in cells treated with conditional medium from glioma cells
(36), indicating that IL1B could
contribute to the progress of brain tumors. In the present study,
we analyzed its gene expression level in two transcriptome
databases. However, changes in IL1B expression showed opposite
trends in these two datasets. It worth noting that there is a large
error bar exhibited by glioblastoma group in the right panel of
Fig. 6B. Nine subjects were involved
in this group (the glioblastoma group in database GSE15824). Among
these subjects, one showed extremely high expression of IL1B (7
fold of the average level), leading to the large error bar in this
dataset. If the outlier was removed, the average expression of IL1B
in glioblastoma group is 22% higher than the control group. In any
case, more evidences are needed to further demonstrate whether PU.1
affect glioma through regulation of IL1B.
BTK, a gene encoding the Bruton Tyrosine
Kinase, was found to be consistently upregulated in both glioma
datasets. It is one of the cytoplasmic protein tyrosine kinases
that expressed by immune cells. BTK can be activated by the B cell
receptor pathways or the FcRγ pathways (27). It has been reported that BTK
activation was involved in several kinds of tumors. Although the
exact role of BTK in glioma is unknown, it could be one of the
important factors in such process (26).
It worth noting that the expression of these
factors, such as FLI1 and IL1B, could be higher in some glioma
tissues, while lower in the others. It is possible there is
overexpression of an abbreviated product, through alternative
splicing or post-transcriptional protein modification. Changes in
the expression pattern of splicing regulator proteins were also
noted at different stages of cancer progression (37). In any case, these factors could
mediate the role of PU.1 in glioma. Further experimental
examination is necessary to explore their function.
To explore the association between gene expression
and patient prognosis, we searched TCGA database the REMBRANDT
database for clinical evidence. Genomic mutations at the SPI1 locus
were found in glioma patients (23).
Moreover, we also noticed that patients with higher SPI1 or FLI1
(REMBRANDT database, P=0.0465) expression level showed lower
survival rate. These results suggest that the expression levels of
some key genes could be considered as prognostic indicators of
glioma malignancy.
Reactome is an online database of biological
pathways (31,32,38). Here
we analyzed potential targets participating PU.1-associated glioma
in Reactome to form a network of biological interactions in this
process. Multiple pathways were identified including immune system,
metabolism, gene expression and signaling transduction, indicating
that PU.1 might influence the progress of glioma via different
biological pathways.
In conclusion, our study provides evidence that PU.1
could play a role in the progress in glioma through its
transcriptional targets. Therefore, PU.1 is involved in the
regulation of glioma and probably in other types of malignant
cancers through different mechanisms. These results suggest a
diverse role for PU.1 in addition to its function in mediating
myeloid linage differentiation and regulation of leukemia probably
via its transcription targets in multiple biological pathways.
Acknowledgements
This study was supported by Cross Research Fund of
Medicine and Engineering of Shanghai Jiaotong University (no.
YG2016QN32) and Natural Science Foundation of Ningbo (no.
2016A610119).
References
|
1
|
Darnell JE Jr: Transcription factors as
targets for cancer therapy. Nat Rev Cancer. 2:740–749. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Verbiest T, Bouffler S, Nutt SL and Badie
C: PU.1 downregulation in murine radiation-induced acute myeloid
leukaemia (AML): From molecular mechanism to human AML.
Carcinogenesis. 36:413–419. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Scott EW, Simon MC, Anastasi J and Singh
H: Requirement of transcription factor PU.1 in the development of
multiple hematopoietic lineages. Science. 265:1573–1577. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
McKercher SR, Torbett BE, Anderson KL,
Henkel GW, Vestal DJ, Baribault H, Klemsz M, Feeney AJ, Wu GE,
Paige CJ and Maki RA: Targeted disruption of the PU.1 gene results
in multiple hematopoietic abnormalities. EMBO J. 15:5647–5658.
1996.PubMed/NCBI
|
|
5
|
Iwasaki H, Somoza C, Shigematsu H, Duprez
EA, Iwasaki-Arai J, Mizuno S, Arinobu Y, Geary K, Zhang P, Dayaram
T, et al: Distinctive and indispensable roles of PU.1 in
maintenance of hematopoietic stem cells and their differentiation.
Blood. 106:1590–1600. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Torlakovic EE, Bilalovic N, Golouh R,
Zidar A and Angel S: Prognostic significance of PU.1 in follicular
lymphoma. J Pathol. 209:352–359. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bonadies N, Pabst T and Mueller BU:
Heterozygous deletion of the PU.1 locus in human AML. Blood.
115:331–334. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Huskova H, Korecka K, Karban J, Vargova J,
Vargova K, Dusilkova N, Trneny M and Stopka T: Oncogenic
microRNA-155 and its target PU.1: An integrative gene expression
study in six of the most prevalent lymphomas. Int J Hematol.
102:441–450. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhou J, Wu J, Li B, Liu D, Yu J, Yan X,
Zheng S, Wang J, Zhang L, Zhang L, et al: PU.1 is essential for MLL
leukemia partially via crosstalk with the MEIS/HOX pathway.
Leukemia. 28:1436–1448. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
de Boer J, Walf-Vorderwülbecke V and
Williams O: In focus: MLL-rearranged leukemia. Leukemia.
27:1224–1228. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Krivtsov AV and Armstrong SA: MLL
translocations, histone modifications and leukaemia stem-cell
development. Nat Rev Cancer. 7:823–833. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Toda Y, Tsukada J, Misago M, Kominato Y,
Auron PE and Tanaka Y: Autocrine induction of the human
pro-IL-1beta gene promoter by IL-1beta in monocytes. J Immunol.
168:1984–1991. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Dolecek TA, Propp JM, Stroup NE and
Kruchko C: CBTRUS statistical report: Primary brain and central
nervous system tumors diagnosed in the United States in 2005–2009.
Neuro Oncol. 14 Suppl 5:v1–v49. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wen PY and Kesari S: Malignant gliomas in
adults. N Engl J Med. 359:492–507. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Phillips HS, Kharbanda S, Chen R, Forrest
WF, Soriano RH, Wu TD, Misra A, Nigro JM, Colman H, Soroceanu L, et
al: Molecular subclasses of high-grade glioma predict prognosis,
delineate a pattern of disease progression, and resemble stages in
neurogenesis. Cancer Cell. 9:157–173. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sottoriva A, Spiteri I, Piccirillo SG,
Touloumis A, Collins VP, Marioni JC, Curtis C, Watts C and Tavaré
S: Intratumor heterogeneity in human glioblastoma reflects cancer
evolutionary dynamics. Proc Natl Acad Sci USA. 110:pp. 4009–4014.
2013; View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Charles NA, Holland EC, Gilbertson R,
Glass R and Kettenmann H: The brain tumor microenvironment. Glia.
59:1169–1180. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li W and Graeber MB: The molecular profile
of microglia under the influence of glioma. Neuro Oncol.
14:958–978. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hu F, Ku MC, Markovic Da Dzaye OD,
Lehnardt S, Synowitz M, Wolf SA and Kettenmann H: Glioma-associated
microglial MMP9 expression is upregulated by TLR2 signaling and
sensitive to minocycline. Int J Cancer. 135:2569–2578. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Pyonteck SM, Akkari L, Schuhmacher AJ,
Bowman RL, Sevenich L, Quail DF, Olson OC, Quick ML, Huse JT,
Teijeiro V, et al: CSF-1R inhibition alters macrophage polarization
and blocks glioma progression. Nat Med. 19:1264–1272. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Markovic DS, Vinnakota K, van Rooijen N,
Kiwit J, Synowitz M, Glass R and Kettenmann H: Minocycline reduces
glioma expansion and invasion by attenuating microglial MT1-MMP
expression. Brain Behav Immun. 25:624–628. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yuan JX and Munson JM: Quantitative
immunohistochemistry of the cellular microenvironment in patient
glioblastoma resections. J Vis Exp. 2017. View Article : Google Scholar
|
|
23
|
Cancer Genome Atlas Research Network, .
Comprehensive genomic characterization defines human glioblastoma
genes and core pathways. Nature. 455:1061–1068. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Poon H, Quirk C, DeZiel C and Heckerman D:
Literome: PubMed-scale genomic knowledge base in the cloud.
Bioinformatics. 30:2840–2842. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Poon H, Toutanova K and Quirk C: Distant
supervision for cancer pathway extraction from text. Pac Symp
Biocomput. 1–131. 2015.PubMed/NCBI
|
|
26
|
Müller S, Sideras P, Smith CI and
Xanthopoulos KG: Cell specific expression of human Bruton's
agammaglobulinemia tyrosine kinase gene (Btk) is regulated by Sp1-
and Spi-1/PU.1-family members. Oncogene. 13:1955–1964.
1996.PubMed/NCBI
|
|
27
|
Starck J, Doubeikovski A, Sarrazin S,
Gonnet C, Rao G, Skoultchi A, Godet J, Dusanter-Fourt I and Morle
F: Spi-1/PU.1 is a positive regulator of the Fli-1 gene involved in
inhibition of erythroid differentiation in friend erythroleukemic
cell lines. Mol Cell Biol. 19:121–135. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Cinque S, Willems J, Depraetere S,
Vermeire L and Joniau M: ‘In vitro’ effect of interleukin-1 beta on
human glioma cell lines: Regulation of cell proliferation and IL-6
production. Immunol Lett. 34:267–271. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kaplitt MG, Tjuvajev JG, Leib DA, Berk J,
Pettigrew KD, Posner JB, Pfaff DW, Rabkin SD and Blasberg RG:
Mutant herpes simplex virus induced regression of tumors growing in
immunocompetent rats. J Neurooncol. 19:137–147. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sahin A, Vercamer C, Kaminski A, Fuchs T,
Florin A, Hahne JC, Mattot V, Pourtier-Manzanedo A, Pietsch T,
Fafeur V and Wernert N: Dominant-negative inhibition of Ets 1
suppresses tumor growth, invasion and migration in rat C6 glioma
cells and reveals differentially expressed Ets 1 target genes. Int
J Oncol. 34:377–389. 2009.PubMed/NCBI
|
|
31
|
Croft D, Mundo AF, Haw R, Milacic M,
Weiser J, Wu G, Caudy M, Garapati P, Gillespie M, Kamdar MR, et al:
The reactome pathway knowledgebase. Nucleic Acids Res. 42(Database
issue): D472–D477. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Fabregat A, Sidiropoulos K, Garapati P,
Gillespie M, Hausmann K, Haw R, Jassal B, Jupe S, Korninger F,
McKay S, et al: The reactome pathway knowledgebase. Nucleic Acids
Res. 44:D481–D487. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Auron PE, Webb AC, Rosenwasser LJ, Mucci
SF, Rich A, Wolff SM and Dinarello CA: Nucleotide sequence of human
monocyte interleukin 1 precursor cDNA. Proc Natl Acad Sci USA.
81:pp. 7907–7911. 1984; View Article : Google Scholar : PubMed/NCBI
|
|
34
|
March CJ, Mosley B, Larsen A, Cerretti DP,
Braedt G, Price V, Gillis S, Henney CS, Kronheim SR, Grabstein K,
et al: Cloning, sequence and expression of two distinct human
interleukin-1 complementary DNAs. Nature. 315:641–647. 1985.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Masters SL, Simon A, Aksentijevich I and
Kastner DL: Horror autoinflammaticus: The molecular pathophysiology
of autoinflammatory disease (*). Annu Rev Immunol. 27:621–668.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhang L, Alizadeh D, Van Handel M,
Kortylewski M, Yu H and Badie B: Stat3 inhibition activates tumor
macrophages and abrogates glioma growth in mice. Glia.
57:1458–1467. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Munkley J, Livermore K, Rajan P and
Elliott DJ: RNA splicing and splicing regulator changes in prostate
cancer pathology. Hum Genet. 136:1143–1154. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Croft D, O'Kelly G, Wu G, Haw R, Gillespie
M, Matthews L, Caudy M, Garapati P, Gopinath G, Jassal B, et al:
Reactome: A database of reactions, pathways and biological
processes. Nucleic Acids Res. 39(Database issue): D691–D697. 2011.
View Article : Google Scholar : PubMed/NCBI
|