Introduction
Ewing's sarcoma (ES), the second most frequent bone
tumor, mainly affects children and adolescents (1). ES is an aggressive and metastatic tumor
type; approximately one-third of patients with ES present with
metastasis at diagnosis, resulting in poor prognosis. The 5-year
survival rate of patients with metastatic ES ranges between 20 and
45%, depending on the metastatic location, compared with 60–70% in
those with localized disease (2).
Therefore, an improved understanding of the biology of this
malignancy is crucial to the development of innovative therapeutic
strategies, the identification of novel therapeutic targets, and
ultimately the improvement of patient prognosis.
MicroRNAs (miRNAs or miRs) are a specific type of
small, non-coding RNA, which regulate gene expression by
suppressing mRNA translation. The aberrant expression of miRNAs
contributes to tumor initiation and progression, and affects
prognosis (3–5). Previously, numerous studies have
indicated that miRNAs are important in the initiation and
progression of ES (6–8). Of these cancer-associated miRNAs,
miR-30d has been shown to exert antitumor effects in a several
types of human cancer (9–11). However, it has been reported that high
tissue expression of miR-30d predicts poor prognosis in patients
with prostate cancer (12). To date,
the exact effects of miR-30d on the progression of ES, and the
underlying mechanisms, remain to be fully elucidated.
In the present study, ES SK-ES-1 cells were
transfected with an miR-30d mimic to examine the effects of miR-30d
on cell proliferation, invasion, migration and apoptosis, and to
elucidate the underlying molecular mechanisms. To the best of our
knowledge, the present study represents the first attempt to
demonstrate the effects of miR-30d on the biological progression of
ES in vitro.
Materials and methods
Materials and reagents
The SK-ES-1 human ES cell line was obtained from the
American Type Culture Collection (Manassas, VA, USA).
Scrambled-sequence mimic and miR-30d mimic were purchased from
Biotend (Shanghai, China). Lipofectamine 2000 and OPTI-MEM were
purchased from Invitrogen; Thermo Fisher Scientific, Inc. (Waltham,
MA, USA). RPMI 1640 medium, fetal bovine serum (FBS),
phosphate-buffered saline (PBS), dimethyl sulfoxide (DMSO), bovine
serum albumin (BSA) and Cell Counting Kit-8 (CCK-8) were purchased
from Beijing Transgen Biotech Co., Ltd. (Beijing, China). The
Cycletest Plus DNA Reagent kit, Annexin-V/FITC kit and Matrigel
were purchased from BD Biosciences (San Jose, CA, USA). The
Transwell invasion chambers were purchased from Costar (Cambridge,
MA, USA). Crystal violet staining solution was purchased from
Beyotime Institute of Biotechnology (Haimen, China). A Hoechst
33258 staining kit was purchased from Nanjing Keygen Biotech Co.,
Ltd. (Nanjing, China). Antibodies against MMP-2 (cat. no. 40994),
MMP-9 (cat. no. 13667), Bax (cat. no. 5023), Bcl-2 (cat. no. 4223),
caspase-3 (cat. no. 9665), PARP (cat. no. 9532), MEK1/2 (cat. no.
8727), ERK1/2 (cat. no. 4695), phosphorylated MEK1/2 (p-MEK1/2)
(cat. no. 9154), phosphorylated ERK1/2 (p-ERK1/2) (cat. no. 4370),
phosphoinositide 3-kinase (PI3K) (cat. no. 4249), Akt (cat. no.
4691), phosphorylated Akt (p-Akt) (cat. no. 4060) and β-actin (cat.
no. 4970) were purchased from Cell Signaling Technology, Inc.
(Danvers, MA, USA). Horseradish peroxidase-conjugated secondary
antibodies (cat. no. HS101-01) were purchased from Beijing Transgen
Biotech Co., Ltd. (Beijing, China).
Cell culture and transfection
The SK-ES-1 cells were grown in RPMI 1640 medium
supplemented with 10% (v/v) FBS, 100 U/ml penicillin, and 100 µg/ml
streptomycin. The cells were maintained at 37°C in an incubator
containing 5% CO2 and passaged every 2–3 days. All cells
used in the present study were subjected to <15 passages. Cells
at logarithmic phase were seeded in 6-well plates at a density of
3×105 cells/well and cultured overnight prior to
transfection. Subsequently, transfections were performed for the
scrambled-sequence mimic and miR-30d mimic (Biotend) using
Lipofectamine 2000 reagent, according to the manufacturer's
protocol. The culture medium was changed from OPTI-MEM to RPMI 1640
medium containing 10% FBS 5 h following transfection. After 24 or
48 h, the transfected cells were collected and used in subsequent
experiments.
Cell proliferation assay
For the analysis of cell proliferation, the cells
were seeded into 96-well plates at 3×103 cells/well, and
incubated in 10% CCK-8 diluted in normal culture medium at 37°C
until visual color conversion was observed. Proliferation was
examined at 24, 48 and 72 h post-transfection by measuring the
absorbance in each well at 450 nm using a universal microplate
reader (EL800; BioTek Instruments, Inc., Winooski, VT, USA). All
experiments were repeated three times.
Boyden chamber Transwell assays
The invasion capacity of the SK-ES-1 cells was
determined using Matrigel-coated Transwell cell culture chambers
(pore size, 8 µm). At 24 h post-transfection, 5×104
cells suspended in serum-free RPMI 1640 medium were added to the
upper chamber, and medium containing 20% FBS was added to the lower
chamber as chemoattractant. After 24 h, non-invading cells on the
upper membrane surface were removed using a cotton swab, whereas
invading cells on the lower membrane surface were fixed, stained
with crystal violet staining solution, and counted under a
phase-contrast microscope (Olympus Corporation, Tokyo, Japan) in
three randomly selected fields (magnification, ×100). The
experiments were performed in triplicate.
Wound healing assays
The migration of SK-ES-1 cells was measured using
wound healing assays. The cells were grown in 6-well plates and
transfected with the scrambled-sequence mimic and miR-30d mimic. At
24 h post-transfection, linear scratch wounds were created on the
confluent cell monolayers using 100 µl pipette tips. To prevent
cells from entering the cell cycle prior to wounding, they were
cultured in serum-free medium. Scratch wounds in the three groups
were observed using a phase-contrast microscope and images were
captured at 0 and 24 h (magnification, ×40). Experiments were
repeated independently three times.
Hoechst 33258 staining of SK-ES-1
cells
At 48 h post-transfection, cells of the three groups
in 6-well plates were fixed with 4% paraformaldehyde for 30 min at
25°C, washed three times with ice-cold PBS, and stained with 10
mg/l Hoechst 33258 for 10 min in the dark at 25°C. Subsequently,
the stained nuclei were observed under a fluorescence microscope
(Olympus Corporation) with excitation at 350 nm and emission at 460
nm (magnification, ×200). The experiments were performed in
triplicate.
Cell cycle analysis
Cell cycle was analyzed via flow cytometry 48 h
post-transfection. The transfected cells of the three groups were
collected and stained with propidium iodide (PI) using a Cycletest
Plus DNA Reagent kit, according the manufacturer's protocol. Cell
cycle distribution was analyzed using a FACSVerse flow cytometer
(BD Biosciences). The percentages of cells in each phase of the
cell cycle were compared. The experiments were performed in
triplicate.
Cell apoptosis analysis
The cell apoptosis assays were performed using an
Annexin-V/FITC kit. Cell apoptosis was detected by flow cytometry
48 h post-transfection. The transfected cells were collected and
washed in ice-cold PBS prior to being stained with Annexin-V/FITC
and PI solution for 15 min in the dark. The ratios of apoptotic
cells were determined using a FACSVerse flow cytometer. All
experiments were repeated three times.
Western blot analysis
Following transfection, the SK-ES-1 cells were grown
in 6-well plates and incubated in RPMI 1640 medium with 10% FBS for
48 h. Subsequently, the cells of the three groups were collected
and lysed in RIPA buffer containing PMSF and phosphatase inhibitor
cocktail. Each sample was centrifuged at 17,105.6 g for 10 min at
4°C to remove cell debris using UNIVERSAL 320R (Hettich Corp.,
Tuttlingen, Germany), and the supernatant was collected for
immunoblotting. The protein concentrations were calculated using
BSA as the standard. Proteins (10 µl) were loaded and separated by
electrophoresis on 10% SDS-polyacrylamide gels at 110 V for 2 h
under reducing conditions. Following electrophoresis, the proteins
were transferred onto PVDF membranes in a Tris-glycine transfer
buffer, and incubated with antibodies against MMP-2 (1:1,000),
MMP-9 (1:1,000), Bax (1:1,000), Bcl-2 (1:1,000), caspase-3
(1:1,000), PARP (1:1,000), MEK1/2 (1:1,000), ERK1/2 (1:1,000),
p-MEK1/2 (1:1,000), p-ERK1/2 (1:1,000), PI3K (1:1,000), Akt
(1:1,000), p-Akt (1:1,000) and β-actin (1:1,000) overnight at 4°C.
The PVDF membranes were washed in TBST for 10 min three times,
following which secondary HRP-conjugated antibodies (1:2,000) were
added at 1:5,000 dilution and incubated for 2 h at 25°C. The PVDF
membranes were subsequently washed three times in TBST.
Immunoreactive proteins were detected by enhanced chemiluminescence
(ECL kit; Beijing Transgen Biotech Co., Ltd.) followed by exposure
to X-ray films. The western blot signals were semi-quantified by
densitometry using ImageQuant TL software 7.0 (GE Healthcare Life
Sciences, Chalfont, UK). All western blot experiments were repeated
at least three times.
Statistical analysis
SPSS 19.0 software (IBM SPSS, Armonk, NY, USA) was
used for statistical analysis of data. Quantitative data are
expressed as the mean ± standard deviation. Independent two-sample
t-tests were used to compare differences between the two groups.
One-way analysis of variance with a multiple comparison test was
used to analyze differences between three or more groups. P<0.05
was considered to indicate a statistically significant
difference.
Results
Overexpression of miR-30d inhibits the
proliferation of SK-ES-1 cells
The effect of the overexpression of miR-30d on
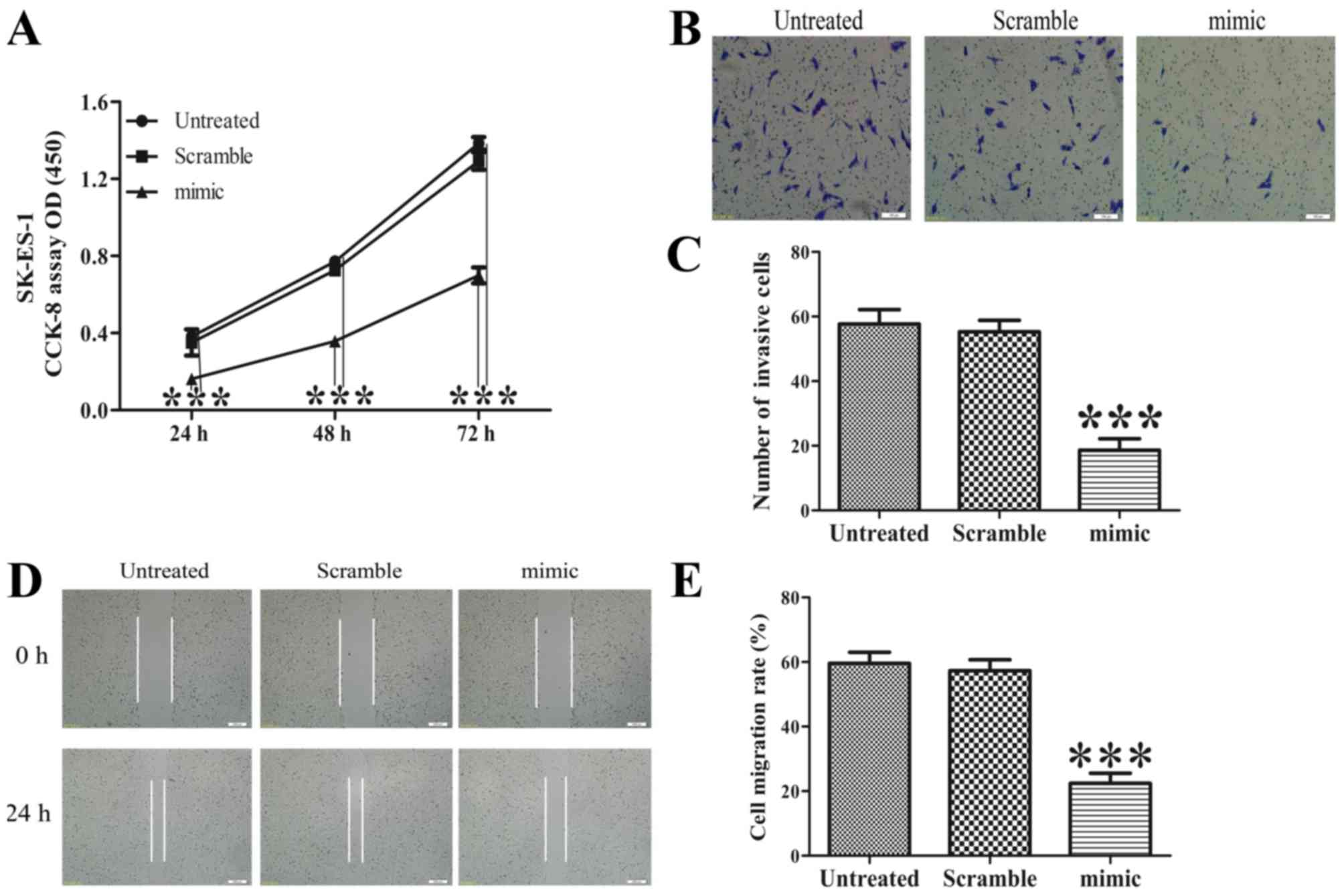
SK-ES-1 cell growth was determined using a CCK-8 assay. As shown in
Fig. 1A, the increased expression of
miR-30d significantly suppressed cell growth of the SK-ES-1 cells
(P<0.001).
Increased expression of miR-30d
represses the invasion and migration of SK-ES-1 cells
Boyden chamber Transwell and wound healing assays
were performed to determine the effect of ectopic expression of
miR-30d on the invasion and migration of SK-ES-1 cells.
Representative images of the Transwell filters are shown in
Fig. 1B. The invasive cell count,
shown in Fig. 1C, demonstrated that
the invasive potential was significantly reduced in the miR-30d
mimic group, compared with that in the untreated group P<0.001).
In addition, the overexpression of miR-30d led to a decrease in
migration capability, as shown in Fig. 1D
and E (P<0.001).
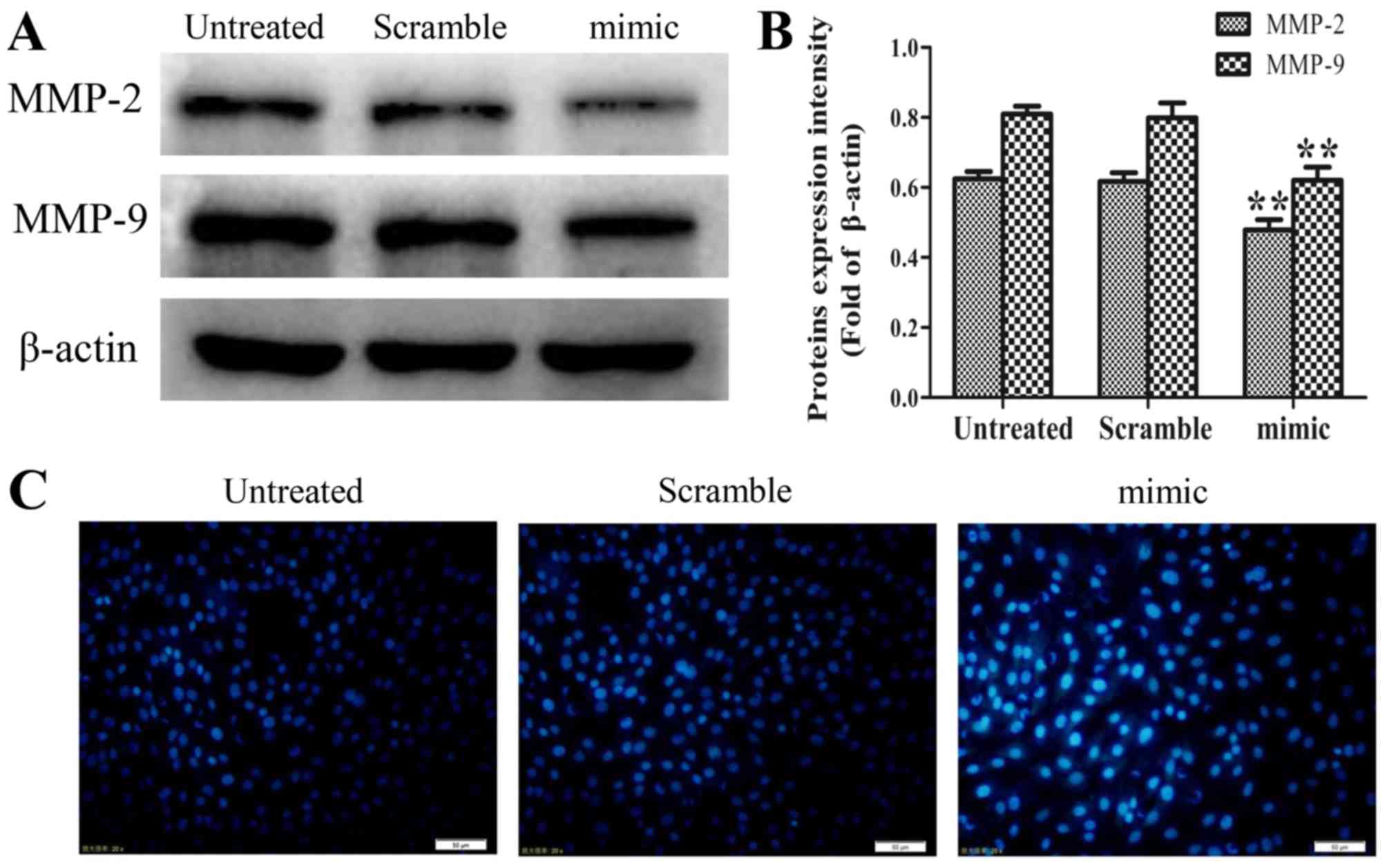
Western blot assays were performed to examine the
effect of increased levels of miR-30d on the expression of MMP-2
and MMP-9, which are closely linked with tumor invasion and
metastasis (13–15). As shown in Fig. 2A and B, the expression levels of MMP-2
and MMP-9 were significantly reduced in the miR-30d mimic group
compared with those in the untreated group (P<0.01). These
findings suggested that the overexpression of miR-30d inhibits the
invasion and migration of SK-ES-1 cells by suppressing the
expression of MMP-2 and MMP-9.
Upregulation of miR-30d induces
morphological changes in SK-ES-1 cells
Staining with the fluorescent DNA-binding dye
Hoechst 33258 revealed that the SK-ES-1 cells transfected with the
miR30d mimic possessed condensed and fragmented nuclei, which are
typical morphological features of apoptotic cells. By contrast, no
morphological signs of apoptosis were observed in the cells of the
untreated group and scrambled-sequence mimic group. These results
indicated that the ectopic expression of miR-30d induced cell death
via apoptosis (Fig. 2C).
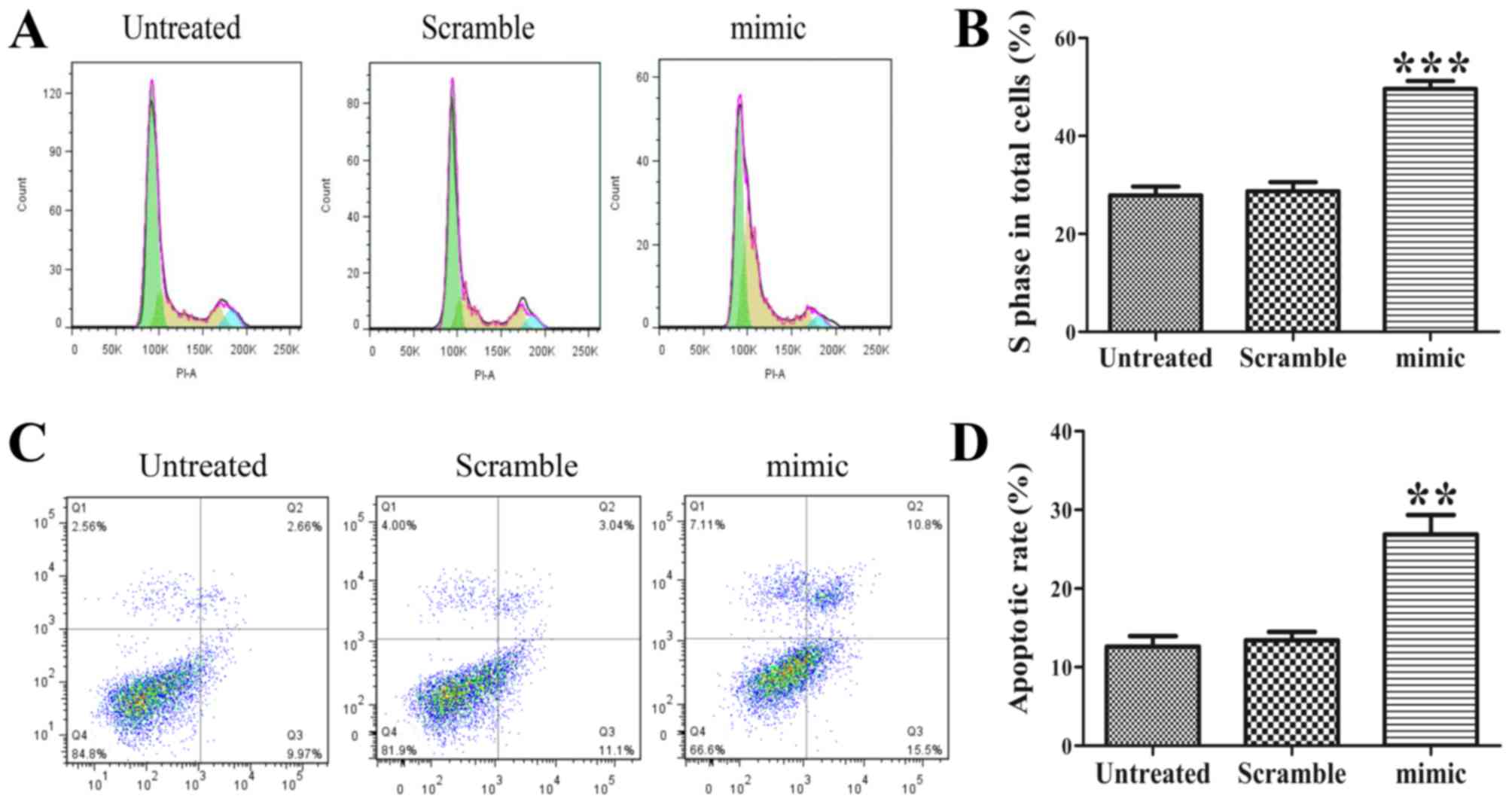
Ectopic expression of miR-30d arrests
the cell cycle at the S-phase
As proliferation is directly linked to cell cycle
distribution, the effect of the miR-30d mimic on cell cycle
progression was analyzed. The results showed that abnormal miR-30d
led to an accumulation of SK-ES-1 cells in the S-phase (Fig. 3A). The statistical analysis revealed a
significant difference in the proportion of S-phase cells between
the miR-30d mimic group and the untreated group (Fig. 3B, P<0.001).
Overexpression of miR-30d promotes the
apoptosis of SK-ES-1 cells
Flow cytometric assays were performed to determine
whether the increased expression of miR-30d induced apoptosis of
the SK-ES-1 cells. The flow cytometric analysis showed that the
proportions of early and late apoptotic cells were significantly
elevated in the miR-30d mimic group compared with those in the
untreated group (Fig. 3C and D,
P<0.01). Therefore, the ectopic expression of miR-30d induced
apoptosis of the SK-ES-1 cells.
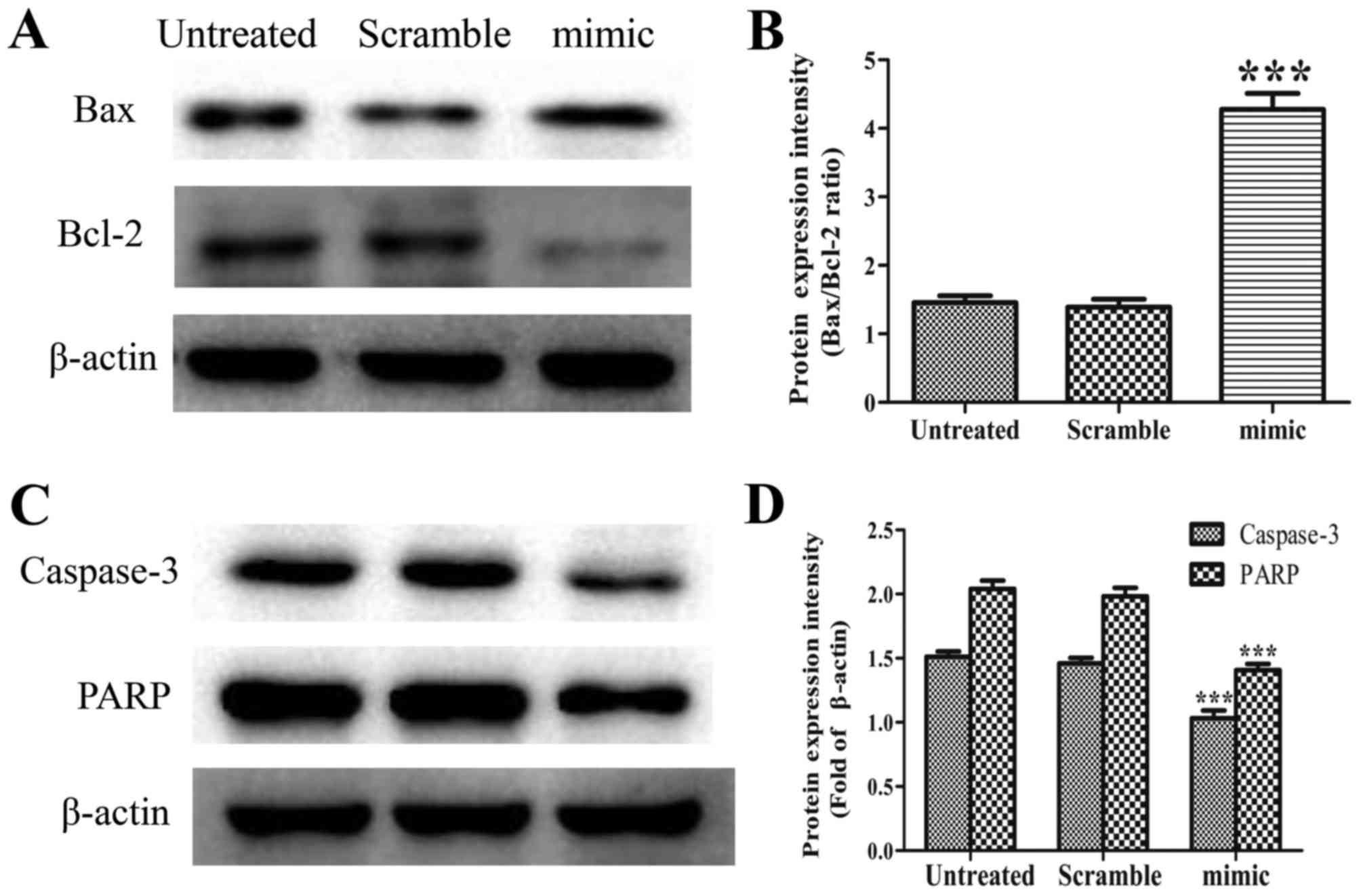
Among the 11 known members of the caspase family,
which are responsible for key aspects of apoptosis-induced cell
death, caspase-3 is considered to be an executioner in caspase
cascades and to have a primary role in apoptosis (16). Following caspase-3 activation,
specific substrates for caspase-3, including PARP, are cleaved,
which are pivotal for the occurrence of apoptosis (17,18). In
addition, changes in the ratio of Bcl-2 family proteins are closely
associated with an imbalance in mitochondrial homeostasis, which
results in apoptosis (19,20). Elevated levels of pro-apoptotic Bax
and/or reduced levels of anti-apoptotic Bcl-2 lead to loss of
mitochondrial membrane potential, which is a key process in the
initiation of apoptosis (21).
To elucidate the mechanism underlying apoptosis,
Bax, Bcl-2, caspase-3 and PARP were detected by western blot
analysis. As shown in Fig. 4A, the
abnormal expression of miR-30d elevated the expression of Bax, but
caused a marked reduction in the level of Bcl-2. The ratio of
Bax/Bcl-2 in the miR-30d mimic group was accordingly increased
(Fig. 4B, P<0.001). The
overexpression of miR-30d led to a marked increase in the cleavage
of caspase-3 and PARP compared with that in the untreated group
(Fig. 4C and D, P<0.001).
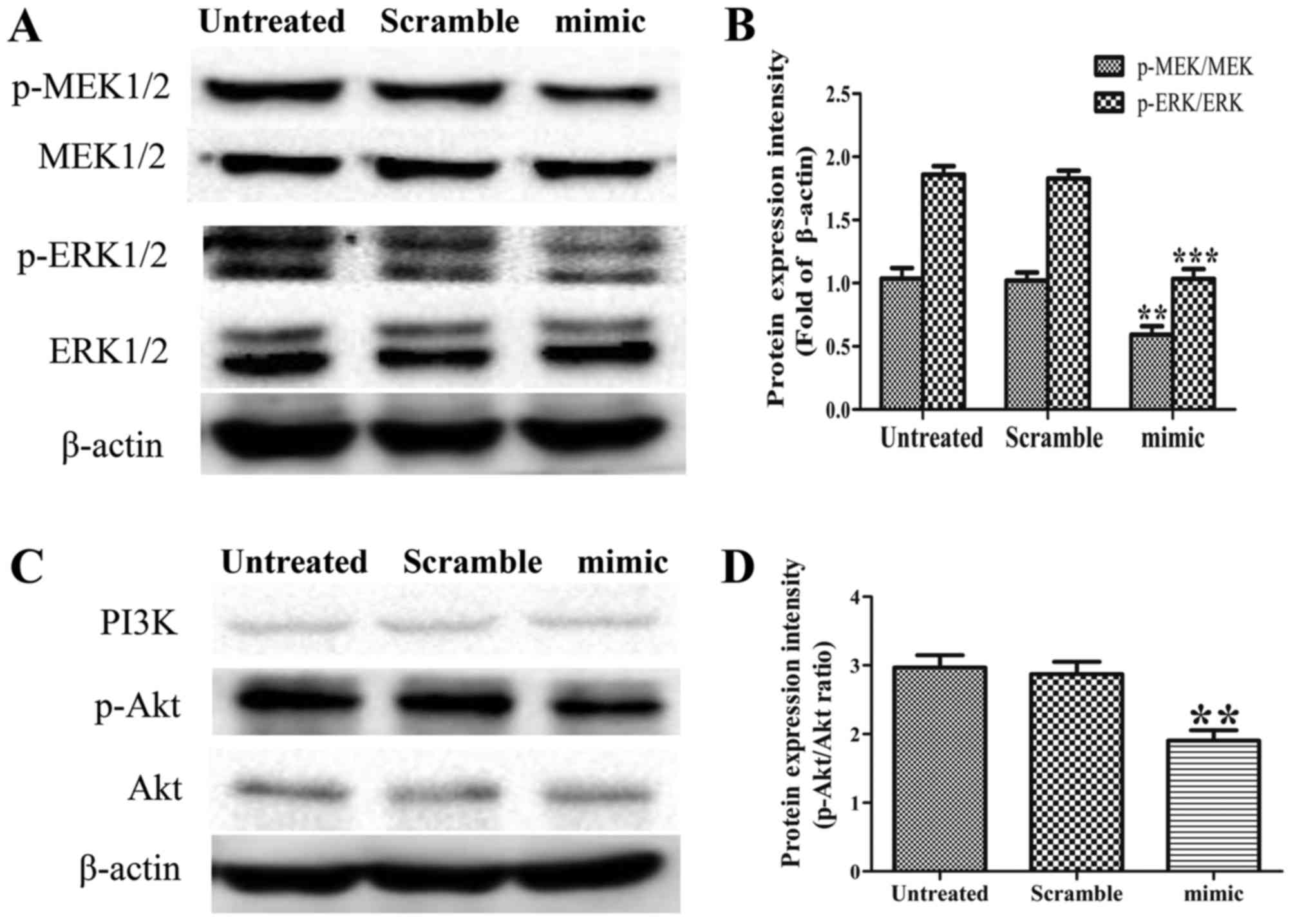
Elevated miR-30d suppresses the
MEK/ERK and PI3K/Akt signaling pathways in SK-ES-1 cells
MEK/ERK, a signaling pathway constitutively
activated in ES, is involved in several cellular processes,
including survival, proliferation, cell cycle regulation,
migration, differentiation, metabolism and protein expression
(22,23). The PI3K/AKT pathway mediates survival
signaling in ES cells (24,25). In the present study, the protein
expression levels of p-MEK1/2, MEK1/2, p-ERK1/2, ERK1/2, PI3K,
p-Akt and Akt were examined in the three groups by western blot
analysis. The results revealed that the overexpression of miR-30d
reduced the expression levels of p-MEK1/2, p-ERK1/2 and p-Akt, but
caused no changes in the levels of MEK1/2, ERK1/2, PI3K or Akt. The
ratios of p-MEK/MEK, p-ERK/ERK and p-Akt/Akt in the miR-30d mimic
group were accordingly decreased, compared with those in the
untreated group (Fig. 5A-D, P<0.01
and P<0.001). These findings demonstrated that miR-30d acts as
an upstream mediator in the MEK/ERK and PI3K/Akt pathways in
SK-ES-1 cells.
Discussion
ES, an aggressive bone and soft tissue malignant
tumor principally affecting children and young adults, is
characterized by small round cells (26,27). Owing
to the development of multi-agent systemic chemotherapy and
aggressive local control methods, the overall survival rate of
patients with localized disease has risen over the last few
decades. Despite these advances, the high rates of relapse and
metastasis, and poor prognosis of patients with ES necessitates the
development of novel treatments to improve patient outcomes
(26,28). A more detailed understanding of the
biology of this malignancy is essential for the identification of
novel therapeutic targets.
miRNAs are small, non-coding RNA molecules with a
length of ~18-25 nucleotides. miRNAs comprise a large family of
non-coding, single-stranded RNAs, which are highly conserved in the
majority of eukaryotic organisms (29,30).
Through complementary binding to 3′-untranslated regions of target
mRNAs, miRNAs regulate gene expression either by suppressing
translation or by directly inducing the degradation of the target
mRNA (31). Increasing evidence shows
that miRNAs are involved in multiple physiological and pathological
processes, including human malignancies (32,33).
Functionally, miRNAs act as either oncogenes or as tumor
suppressors in various types of malignancy (34).
miR-30d, a member of the newly identified miR-30
family, is implicated in tumor development and progression,
however, its precise function in this context remains
controversial. miR-30d is reported to act as a tumor suppressor in
renal carcinoma and anaplastic thyroid carcinoma (10,35), and
as an oncogene in melanoma and hepatocellular carcinoma (36,37). To
date, there have been no reports of the exact role of miR-30d in
the proliferation, malignant phenotype and apoptosis of ES cells,
or the underlying molecular mechanisms.
In the present study, it was demonstrated that
increased expression of miR-30d inhibited the proliferation,
migration and invasion of ES cells, and promoted morphological
changes, S-phase arrest and apoptosis in this cell type.
Additionally, it was observed that aberrant expression of miR-30d
suppressed the expression of MMP-2 and MMP-9, and inhibited the
activity of the MEK/ERK and PI3K/Akt pathways. The overexpression
of miR-30d also increased the ratio of Bax/Bcl-2 and promoted the
cleavage of caspase-3 and PARP. These findings confirmed that
ectopic miR-30d induced cell apoptosis via the Bax/Bcl-2 and
caspase-3 cascade, and that miR-30d is likely to act as an upstream
mediator in the MEK/ERK and PI3K/Akt pathways in SK-ES-1 cells.
There were certain limitations to the present study.
First, knockdown of the expression of miR-30d was not performed,
which may provide confirmation of the effect of miR-30d on SK-ES-1
cells. Second, the present study did not exclude the possibility
that the apoptotic effect may result in the suppression of
proliferation, invasion and migration. Additionally, the present
study was more observational than an in-depth investigation of the
mechanism. There was no detailed examination of the molecular
mechanism concerning the findings observed. For example, no
experiments were performed to determine whether the inhibition of
ERK or AKT caused S-phase arrest. In addition, only S473 was
examined for AKT phosphorylation, which usually consists of S473
and T307. To confirm the target gene of miR-30d, a dual luciferase
reporter assay is also required. Therefore, further investigations
are urgently required to examine the underlying molecular
mechanisms in detail.
In conclusion, the findings observed in SK-ES-1
cells suggested that miR-30d inhibited the biological progression
of ES in vitro by suppressing the MEK/ERK and PI3K/Akt
pathways. Therefore, miR-30d is a promising novel therapeutic
target for treating ES. However, further investigations are
urgently required to examine the underlying molecular mechanisms of
the effects of miR-30d on ES for a comprehensive understanding of
the tumorigenesis and progression of this type of cancer.
Acknowledgements
The present study was supported by the Foundation of
the Health Department of Jiangxi Province on Traditional Chinese
Medicine (grant no. 2016A073) and Gan-Po Talents Project 555 of
Jiangxi Province.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gaspar N, Hawkins DS, Dirksen U, Lewis IJ,
Ferrari S, Le Deley MC, Kovar H, Grimer R, Whelan J, Claude L, et
al: Ewing sarcoma: Current management and future approaches through
collaboration. J Clin Oncol. 33:3036–3046. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Medina-Villaamil V, Martínez-Breijo S,
Portela-Pereira P, Quindós-Varela M, Santamarina-Caínzos I,
Antón-Aparicio LM and Gómez-Veiga F: Circulating microRNAs in blood
of patients with prostate cancer. Actas Urol Esp. 38:633–639. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yang M, Liu R, Sheng J, Liao J, Wang Y,
Pan E, Guo W, Pu Y and Yin L: Differential expression profles of
microRNAs as potential biomarkers for the early diagnosis of
esophageal squamous cell carcinoma. Oncol Rep. 29:169–176. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Leite KR, Tomiyama A, Reis ST,
Sousa-Canavez JM, Sañudo A, Camara-Lopes LH and Srougi M: MicroRNA
expression profles in the progression of prostate cancer-from
high-grade prostate intraepithelial neoplasia to metastasis. Urol
Oncol. 31:796–801. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Li Y, Shao G, Zhang M, Zhu F, Zhao B, He C
and Zhang Z: miR-124 represses the mesenchymal features and
suppresses metastasis in Ewing sarcoma. Oncotarget. 8:10274–10286.
2017.PubMed/NCBI
|
|
7
|
Schwentner R, Herrero-Martin D, Kauer MO,
Mutz CN, Katschnig AM, Sienski G, Alonso J, Aryee DN and Kovar H:
The role of miR-17-92 in the miRegulatory landscape of Ewing
sarcoma. Oncotarget. 8:10980–10993. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kawano M, Tanaka K, Itonaga I, Iwasaki T
and Tsumura H: MicroRNA-301a promotes cell proliferation via PTEN
targeting in Ewing's sarcoma cells. Int J Oncol. 48:1531–1540.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Esposito F, Tornincasa M, Pallante P,
Federico A, Borbone E, Pierantoni GM and Fusco A: Down-regulation
of the miR-25 and miR-30d contributes to the development of
anaplastic thyroid carcinoma targeting the polycomb protein EZH2. J
Clin Endocrinol Metab. 97:E710–E718. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yu H, Lin X, Wang F, Zhang B, Wang W, Shi
H, Zou B and Zhao J: Proliferation inhibition and the underlying
molecular mechanisms of microRNA-30d in renal carcinoma cells.
Oncol Lett. 7:799–804. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chen D, Guo W, Qiu Z, Wang Q, Li Y, Liang
L, Liu L, Huang S, Zhao Y and He X: MicroRNA-30d-5p inhibits tumour
cell proliferation and motility by directly targeting CCNE2 in
non-small cell lung cancer. Cancer Lett. 362:208–217. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kobayashi N, Uemura H, Nagahama K, Okudela
K, Furuya M, Ino Y, Ito Y, Hirano H, Inayama Y, Aoki I, et al:
Identification of miR-30d as a novel prognostic maker of prostate
cancer. Oncotarget. 3:1455–1471. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li H, Zhang K, Liu LH, Ouyang Y, Bu J, Guo
HB and Xiao T: A systematic review of matrix metalloproteinase 9 as
a biomarker of survival in patients with osteosarcoma. Tumour Biol.
35:5487–5491. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang J, Shi Q, Yuan TX, Song QL, Zhang Y,
Wei Q, Zhou L, Luo J, Zuo G, Tang M, et al: Matrix
metalloproteinase 9 (MMP-9) in osteosarcoma: Review and
meta-analysis. Clin Chim Acta. 433:225–231. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Shang HS, Chang JB, Lin JH, Lin JP, Hsu
SC, Liu CM, Liu JY, Wu PP, Lu HF, Au MK and Chung JG: Deguelin
inhibits the migration and invasion of U-2 OS human osteosarcoma
cells via the inhibition of matrix metalloproteinase-2/-9 in vitro.
Molecules. 19:16588–16608. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Vinatier D, Dufour P and Subtil D:
Caspases: Pharmacological manipulation of cell death. Eur J Obstet
Gynecol Reprod Biol. 67:85–102. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Norbury CJ and Zhivotovsky B: DNA
damage-induced apoptosis. Oncogene. 23:2797–2808. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Soldani C and Scovassi AI:
Poly(ADP-ribose) polymerase-1 cleavage during apoptosis: An update.
Apoptosis. 7:321–328. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Orrenius S: Mitochondrial regulation of
apoptotic cell death. Toxicol Lett. 149:19–23. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Scorrano L and Korsmeyer SJ: Mechanisms of
cytochrome c release by proapoptotic Bcl-2 family members. Biochem
Biophys Res Commun. 304:437–444. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Meeran SM and Katiyar SK: Grape seed
proanthocyanidins promote apoptosis in human epidermoid carcinoma
A431 cells through alterations in Cdki-Cdk-cyclin cascade, and
caspase-3 activation via loss of mitochondrial membrane potential.
Exp Dermatol. 16:405–415. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zenali MJ, Zhang PL, Bendel AE and Brown
RE: Morphoproteomic confrmation of constitutively activated mTOR,
ERK, and NF-kappaB pathways in Ewing family of tumors. Ann Clin Lab
Sci. 39:160–166. 2009.PubMed/NCBI
|
|
23
|
Roskoski R Jr: ERK1/2 MAP kinases:
Structure, function and regulation. Pharmacol Res. 66:105–143.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Datta SR, Brunet A and Greenberg ME:
Cellular survival: A play in three Akts. Genes Dev. 13:2905–2927.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Toretsky JA, Thakar M, Eskenazi AE and
Frantz CN: Phosphoinositide 3-hydroxide kinase blockade enhances
apoptosis in the Ewing's sarcoma family of tumors. Cancer Res.
59:5745–5750. 1999.PubMed/NCBI
|
|
26
|
Balamuth NJ and Womer RB: Ewing's sarcoma.
Lancet Oncol. 11:184–192. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Iwamoto Y: Diagnosis and treatment of
Ewing's sarcoma. Jpn J Clin Oncol. 37:79–89. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gorlick R, Janeway K, Lessnick S, Randall
RL and Marina N; COG bone tumor committee, : Children's oncology
group's 2013 blueprint for research: Bone tumors. Pediatr Blood
Cancer. 60:1009–1015. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lee RC, Feinbaum RL and Ambros V: The C.
elegans heterochronic gene lin-4 encodes small RNAs with antisense
complementarity to lin-14. Cell. 75:843–854. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Šulc M, Marín RM, Robins HS and Vaníček J:
PACCMIT/PACCMIT-CDS: Identifying microRNA targets in 3′ UTRs and
coding sequences. Nucleic Acids Res. 43:W474–W479. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Fang Z, Tang J, Bai Y, Lin H, You H, Jin
H, Lin L, You P, Li J, Dai Z, et al: Plasma levels of microRNA-24,
microRNA-320a, and microRNA-423-5p are potential biomarkers for
colorectal carcinoma. J Exp Clin Cancer Res. 34:862015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Nishikawa R, Goto Y, Kurozumi A,
Matsushita R, Enokida H, Kojima S, Naya Y, Nakagawa M, Ichikawa T
and Seki N: MicroRNA-205 inhibits cancer cell migration and
invasion via modulation of centromere protein F regulating pathways
in prostate cancer. Int J Urol. 22:867–877. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Volinia S, Calin GA, Liu CG, Ambs S,
Cimmino A, Petrocca F, Visone R, Iorio M, Roldo C, Ferracin M, et
al: A microRNA expression signature of human solid tumors defines
cancer gene targets. Proc Natl Acad Sci USA. 103:pp. 2257–2261.
2006; View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhang Y, Yang WQ, Zhu H, Qian YY, Zhou L,
Ren YJ, Ren XC, Zhang L, Liu XP, Liu CG, et al: Regulation of
autophagy by miR-30d impacts sensitivity of anaplastic thyroid
carcinoma to cisplatin. Biochem Pharmacol. 87:562–570. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Gaziel-Sovran A, Segura MF, Di Micco R,
Collins MK, Hanniford D, Vega-Saenz de Miera E, Rakus JF, Dankert
JF, Shang S, Kerbel RS, et al: miR-30b/30d regulation of GalNAc
transferases enhances invasion and immunosuppression during
metastasis. Cancer Cell. 20:104–118. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yao J, Liang L, Huang S, Ding J, Tan N,
Zhao Y, Yan M, Ge C, Zhang Z, Chen T, et al: MicroRNA-30d promotes
tumor invasion and metastasis by targeting Galphai2 in
hepatocellular carcinoma. Hepatology. 51:846–856. 2010.PubMed/NCBI
|