Introduction
As the third most commonly occurring cancer type in
females, cervical cancer (CC) is the fourth leading cause of
cancer-associated mortality globally, resulting in ~300,000
mortalities each year (1–3). In previous years, diagnosis and
treatment options for CC have been markedly improved; however, CC
remains a public health concern worldwide due to the lack of
detailed data on the molecular mechanisms underlying its
pathogenesis. There is therefore an urgent requirement to uncover
novel molecular diagnostic markers and therapeutic targets for the
treatment of CC.
MicroRNAs (miRNAs/miRs) are a family of small
non-coding RNA molecules that are able to bind to the
3′-untranslated region (3′-UTR) of specific mRNA targets to inhibit
their translation. Several previous studies have demonstrated that
miRNAs are typically deregulated or induce the abnormal expression
of key genes and pathways governing cancer-associated processes
(4,5).
Previous studies have demonstrated that miR-21-5p is
upregulated in numerous types of cancer and has been associated
with cellular proliferation, migration, invasion and apoptosis,
which are the main processes underlying cancer pathogenesis
(6,7).
It has been demonstrated that the downregulation of special AT-rich
sequence-binding protein 1 and miR-21-5p overexpression in clear
cell renal cell carcinoma are associated with each other, and with
poor prognosis (8). A prior study
demonstrated the association of miR-21-5p overexpression with
tongue cancer cell chemo-resistance via cell adhesion molecule 1
targeting (9). miR-21-5p
overexpression has also been identified as a possible predictor of
tumor regression following chemoradiotherapy in patients with
rectal cancer (10), and as a
biomarker of recurrence in young patients with gastric cancer
(11) and thymic epithelial tumors
(12). It has been reported that the
competing endogenous regulation of miR-21-5p by the long non-coding
RNA maternally expressed 3 inhibits CC cell proliferation and
apoptosis (13). miR-21-5p
dysregulation has shown promise as a biomarker for the diagnosis of
CC (7). At the molecular level, it
has been demonstrated that miR-21-5p promotes CC cell proliferation
and migration by inhibiting phosphatase and tensin homolog (PTEN)
(3). However, the underlying
molecular mechanisms of miR-21-5p in these processes in CC remain
unresolved.
Typically, miRNAs execute their function by
targeting tumor suppressor genes or oncogenes. To fully appreciate
the implication of miR-21-5p in CC pathogenesis, it is important to
study its association with these molecules. Among the tumor
suppressor genes, von Hippel-Lindau tumor suppressor (VHL), which
is located on chromosome 3p25, is downregulated due to mutations or
silencing in the majority of carcinomas (14–16). This
downregulation of VHL induces hypoxia via hypoxia-inducible factors
(HIF), and subsequent angiogenesis and cell growth (14–16). The
VHL-dependent regulation of miRNAs in renal cancer has been
previously studied and results indicated that miR-21-5p may be
regulated by VHL. Thus far, the association between miR-21-5p and
VHL remains unresolved.
The objective of the present study was therefore to
verify whether miR-21-5p and VHL interact with each other and serve
a function in CC, and to uncover the molecular mechanisms
underlying these processes in CC.
Materials and methods
Cell line and cell culture
Human cervical carcinoma SiHa, HeLa, CaSki, c4-1 and
c-33a cell lines and normal cervical Ect1/E6E7 cells were provided
by the American Type Culture Collection (Manassas, VA, USA).
Dulbecco's modified Eagle's medium (DMEM; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) supplemented with 10% bovine calf serum
(Gibco; Thermo Fisher Scientific, Inc.) was used for cell culture.
All cells were cultured at 37°C in a humidified incubator with 5%
CO2.
Transfection
The miR-21-5p mimic (catalog no. HMI0371;
Mission®) (sense, 5′-UAGCUUAUCAGACUGAUGUUGAA-3′, and
antisense, 5′-UCAACAUCAGUCUGAUAAGCUAUU-3′), negative control (NC)
(sense, 5′-UUCUCCGAACGUGUCACGUTT-3′, and antisense,
5′-ACGUGACACGUUCGGAGAATT-3′), miR-21-5p inhibitor (catalog no.
HLTUD0371; Mission; 5′-CUUCAACAUCAGUCUGAUAAGCUA-3′) were purchased
from Sigma-Aldrich (Merck KGaA, Darmstadt, Germany). The human VHL
small interfering (si)RNAs (cat no. CRH5082-30) (A, sense,
5′-CCAGGUCAUCUUCUGCAAUTT-3′, and antisense,
5′-AUUGCAGAAGAUGACCUGGTT-3′; B, sense, 5′-GGGCUUCUGGUUAACCAAATT-3′,
and antisense, 5′-UUUGGUUAACCAGAAGCCCTT-3′; C, sense,
5′-GGAGCGCAUUGCACAUCAATT-3′, and antisense,
5′-UUGAUGUGCAAUGCGCUCCTT-3′) were purchased from Cohesion
Biosciences (London, UK). The siRNAs A, B and C were transfected at
an equimolar ratio. A total of 5,000 CaSki cells were seeded in
96-well plates and once cells had grown to 50–80% confluence, they
were transfected with miR-21-5p mimic (10 nM) or transduced with
the miR-21-5p inhibitor (10 nM) following the procedures described
in the manufacturer's protocol, while VHL siRNA (100 nM) was
transfected with Lipofectamine® 2000 (Invitrogen; Thermo
Fisher Scientific, Inc.) according to the manufacturer's protocol.
Cells were harvested after 72 h for subsequent analysis.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
For the analysis of VHL mRNA expression, total RNA
was extracted from CC cell lines and normal cervical cells using
TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.) and
reverse transcribed using the Invitrogen SuperScript™ III Reverse
Transcriptase kit (Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. For miR-21-5p expression analysis, the
mirVana miRNA Isolation kit (Thermo Fisher Scientific, Inc.) was
used according to the manufacturer's protocol and cDNA was
generated with miRNA Reverse Transcription kit (Qiagen China Co.,
Ltd., Shanghai, China). qPCR was performed using StepOnePlus
Real-Time PCR system (Thermo Fisher Scientific, Inc.) with The
PrimePCR™ Template for SYBR® Green assay (VHL, Human)
primers (sense, 5′-GAGATGCAGGGACACACGAT-3′, and antisense,
5′-ATCCGTTGATGTGCAATGCG-3′; internal reference GAPDH, sense,
5′-GCCAGTAGAGGCAGGGATGATGTTC-3′, and antisense,
5′-CCATGTTCGTCATGGGTGTGAACCA-3′) (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA) for VHL PCR reaction and MystiCq®
microRNA qPCR Assay Primer hsa-miR-21-5p (Sigma Aldrich; Thermo
Fisher Scientific, Inc.) for the miR-21-5p PCR reaction. GAPDH and
U6 small nuclear RNA were used for the normalization of VHL and
miR-21-5p levels, respectively. Relative expression was estimated
based on the 2−ΔΔCq method (17).
Cell viability assay
Proliferation of CaSki cells was measured using MTT
assay. Cells were transfected for 24 h then seeded in 96-well
microtiter plates for 72 h (1×104 cells/well). MTT was
added for another 4-h incubation at 37°C. The medium was aspirated
and 150 µl DMSO was added for 15 min until the crystals were
dissolved. The absorbance was determined using spectrophotometry at
490 nm using a microplate reader. The experiments were repeated in
triplicate.
Cell migration and invasion
assays
To determine their invasion and migration abilities
CaSki cells, which expressed a higher level of miR-21-5p than the
other examined cell lines, were cultured in Matrigel-coated
(invasion) or uncoated (migration) Transwell inserts. Cells
transfected with miR-21-5p mimic/inhibitor or VHL inhibitor or
controls (~2×104 cells) were suspended in 200 ml
serum-free DMEM and seeded in the upper chamber. In the lower
chamber, the DMEM was supplemented with bovine calf serum (Gibco;
Thermo Fisher Scientific, Inc.) and used as a chemo-attractant.
After 48 h of incubation, the cells that had migrated through the
Matrigel-coated or uncoated membrane were fixed in 100% methanol
for 10 min, stained with 0.1% crystal violet for 10 min at room
temperature, and counted under a white light microscope (Olympus,
Tokyo, Japan).
Western blot analysis
CaSki cells were lysed with radioimmunoprecipitation
assay lysis buffer (Beyotime Institute of Biotechnology, Haimen,
China). Protein concentrations were determined by a BCA assay.
Protein samples (20 µg per lane) were separated using 12% SDS-PAGE
and transferred to a PVDF membrane (Bio-Rad Laboratories, Inc.).
The membrane was blocked with skimmed milk solution (10%) in TBST
at room temperature and incubated overnight at 4°C with VHL
antibody at dilution of 1:3,000 (catalog no. FL-181; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA), caspase-3 at dilution of
1:500 and cleaved caspase-3 antibodies at dilution of 1:2,000
(catalog no. Asp175; Cell Signaling Technology, Inc., Danvers, MA,
USA). Following washing with TBST buffer (three ×15 min), the
appropriate horseradish peroxidase-conjugated goat anti-rabbit
secondary antibody at a dilution of 1:4,000 (Invitrogen; Thermo
Fisher Scientific, Inc.; catalog no. 65-6120), was added for 1 h at
room temperature. β-actin antibody at a dilution of 1:2,000 was
used as an internal reference (Cell Signaling Technology, Inc.;
catalog no. 4970). The immunoreactive proteins were visualized
using an enhanced chemiluminescence system (GE Healthcare Life
Sciences, Little Chalfont, UK) and grayscale value analysis by
ImageJ software was used for quantification.
Caspase-Glo 3/7 assay
CaSki cells were transfected for 24 h then seeded in
96-well microliter plates for 48 h (1×104 cells/well).
The 96-well plates containing transfected cells were removed from
the incubator and were allowed to equilibrate to room temperature.
A total of 100 µl Caspase-Glo® 3/7 reagent (Promega
Corporation, Madison, WI, USA) was then added to each well of a
white-walled 96-well plate containing 100 µl of culture medium
containing untransfected cells, transfection control cells or
transfected cells in culture medium. The contents of the wells were
mixed using a plate shaker at 300 rpm for 30 sec, and incubated at
room temperature for 1 h. The luminescence of each sample was
measured using a plate-reading luminometer.
Luciferase reporter assay
The miR-21-5p binding sites from the 3′-UTR of VHL
or mutant 3′-UTR were cloned into the pGL3 reporter luciferase
vector (Shanghai GeneChem Co., Ltd., Shanghai, China). CaSki cells
were cultured in 96-well plates. Aliquots of 10 nM miR-21-5p mimic
or control miRNA were cotransfected with 0.1 mg of the pGL3-3′ UTR
wild-type or mutant plasmid DNA into CaSki cells using
Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific, Inc.).
Following transfection for 48 h, luciferase activity was measured
using the Dual-Luciferase Reporter Assay system according to the
manufacturer's protocol (Promega Corporation). The relative
luciferase activity was normalized against Renilla luciferase
activity. The experiments were conducted in triplicate.
Bioinformatics prediction
TargetScan Human 7.0 (http://www.targetscan.org/) was used to identify
potential targets of miR-21-5p.
Statistical analysis
All statistical analyses were conducted using
GraphPad Prism software (version 6; GraphPad Software, Inc., La
Jolla, CA, USA). The data are expressed as the mean ± standard
deviation. Differences between groups were assessed using a one-way
analysis of variance (ANOVA) followed by the Bonferroni multiple
comparison post-hoc test or using a two-way ANOVA with the
Bonferroni multiple comparison post-hoc test. P<0.05 was
considered to indicate a statistically significant difference.
Results
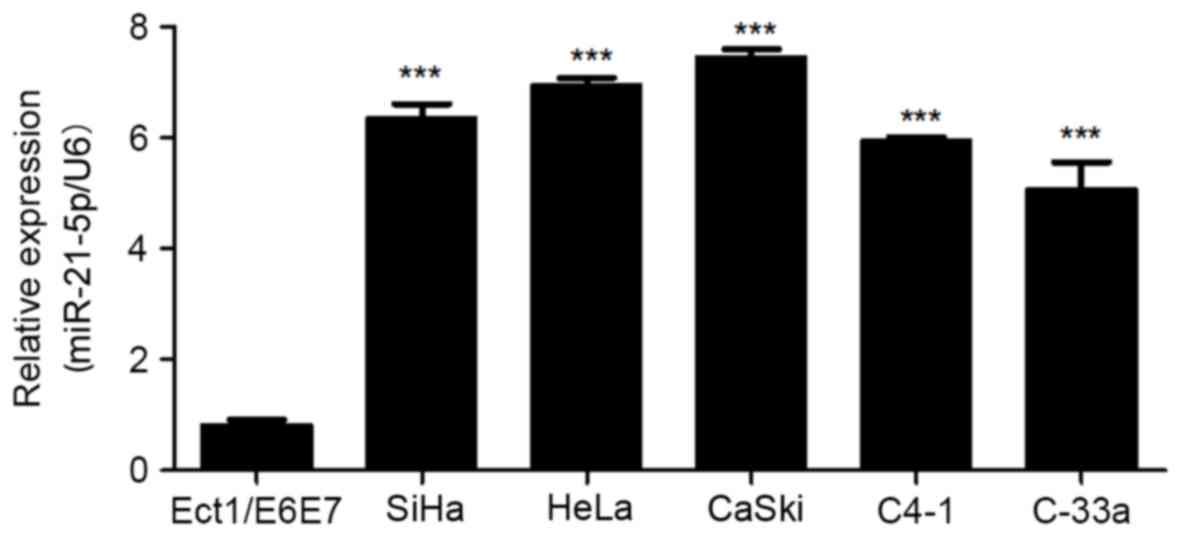
miR-21-5p is upregulated in CC cell
lines
In order to analyze the effect of miR-21-5p on the
progression of CC, the expression level of miR-21-5p was measured
in CC cell lines. As presented in Fig.
1, miR-21-5p expression was significantly upregulated in CC
SiHa, HeLa, CaSki, C4-1 and C-33a cell lines compared with that in
the normal cervical cells. This suggests that miR-21-5p may be
involved in the development of CC.
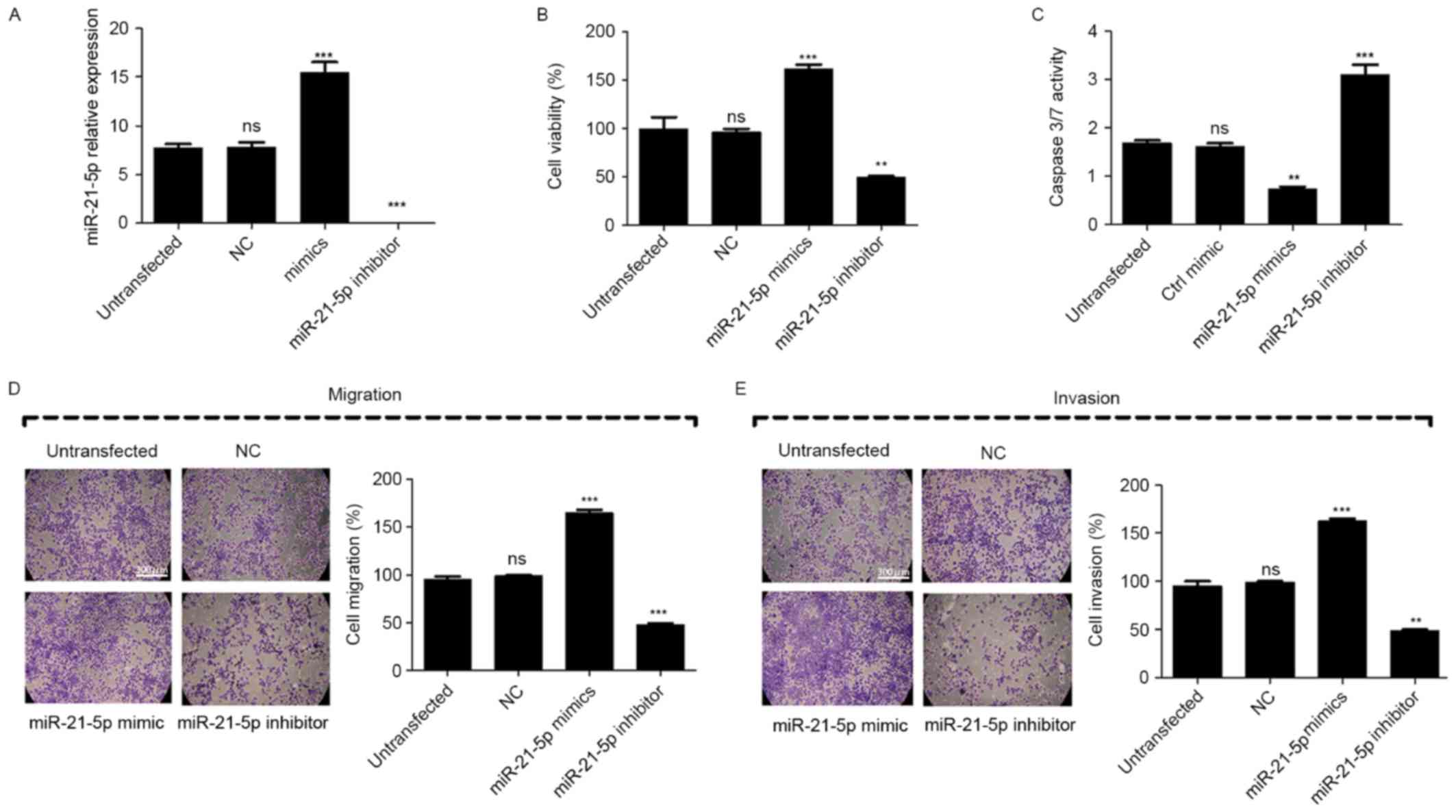
Overexpression of miR-21-5p suppresses
cell proliferation, migration and invasion, and induces apoptosis
in vitro
To further analyze the effect of miR-21-5p on the
proliferation, migration and invasion of CC cells, miR-21-5p mimic
or inhibitor was transfected into CaSki cells. Compared with
untransfected cells, the transfection of miR-21-5p mimic led to
significant upregulation of miR-21-5p in CaSki cells, while
miR-21-5p inhibitor led to significant repression of miR-21-5p
(Fig. 2A).
The result of the MTT assay revealed that inhibition
of miR-21-5p significantly suppressed the proliferation of the
CaSki cells compared with untransfected cells, while the
overexpression of miR-21-5p increased the proliferation of the
CaSki cells (Fig. 2B). To evaluate
the effect of miR-21-5p on the apoptosis of the CaSki cells, a
caspase3/7 activity assay was performed and the results of the
assay revealed that when cells were transfected with miR-21-5p
inhibitor, the combined activity of Caspase 3 and Caspase 7 was
significantly increased compared with that of untransfected cells,
while the overexpression of miR-21-5p significantly decreased this
activity (Fig. 2C). These results
suggest that the inhibition of miR-21-5p was followed by induction
of CaSki cell apoptosis. Furthermore, Transwell assays were used to
measure the effect of miR-21-5p on cell migration and invasion.
Compared with the untransfected control, the results revealed that
the migration and invasion of CaSki cells were significantly
suppressed when cells were transfected with miR-21-5p inhibitor,
but increased when cells were transfected with miR-21-5p mimic,
compared with the untransfected control (Fig. 2D and E). These results demonstrated
that the inhibition of miR-21-5p expression suppressed CC cell
proliferation, migration and invasion, and induced apoptosis in
vitro.
miR-21-5p directly targets the VHL
gene
VHL was predicted as the target gene of miR-21-5p
using TargetScan Human 7.0 software (Fig.
3A). The expression of VHL in CC cells and normal cervical
cells was measured. The results revealed that VHL expression was
significantly decreased in all CC cell lines compared with the
controls (Fig. 3B-D). To further
confirm whether VHL is a target of miR-21-5p, a dual luciferase
assay was performed. The results revealed that the miR-21-5p mimic
significantly increased luciferase activity in the mutant cells
compared with that in the wild-type cells. Treatment with an
miR-21-5p inhibitor produced no significant difference in
luciferase activity between wild-type and mutant cells (Fig. 3E). The results of qPCR identified that
the expression of VHL mRNA was not significantly altered following
overexpression of miR-21-5p in the CaSki cells (Fig. 3F). Furthermore, overexpression of
miR-21-5p resulted in downregulation of VHL protein expression
(P<0.05; Fig. 3G). These results
suggest that VHL is a direct target gene of miR-21-5p.
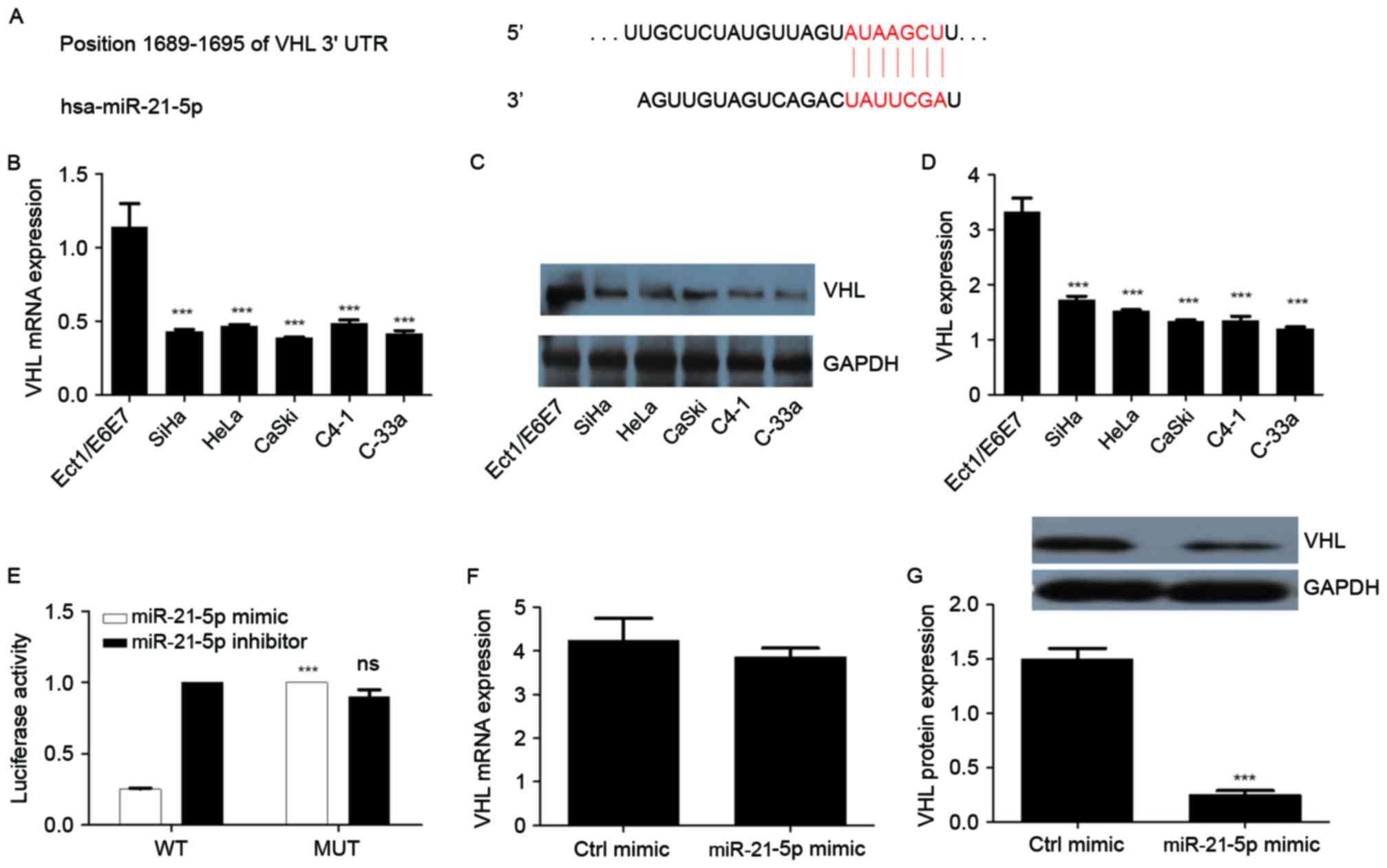 | Figure 3.VHL, downregulated in CC, is a direct
target of miR-21-5p. (A) Targetscan prediction of miR-21-5p binding
site on the 3′-UTR region of VHL. (B) mRNA expression of VHL in CC
cells. (C) Western blot analysis of VHL in different CC cell lines.
(D) Quantitative representation of VHL in different CC cell lines.
GAPDH was used as endogenous control. (E) Luciferase reporter assay
for VHL. (F) Effect of miR-21-5p expression on the mRNA expression
of VHL in CaSki cells. (G) Effect of miR-21-5p expression on the
protein expression of VHL in CaSki cells. Statistical analysis was
performed using one-way ANOVA or two-way ANOVA (for luciferase
activity assay). Error bars represent the standard deviation.
***P<0.001 compared with Ect1/E6E7 or MUT groups. ns,
non-significant; CC, cervical cancer; miR, microRNA; VHL, von
Hippel-Lindau suppressor; ANOVA, analysis of variance; WT,
wild-type; MUT, mutant; ctrl, control; UTR, untranslated
region. |
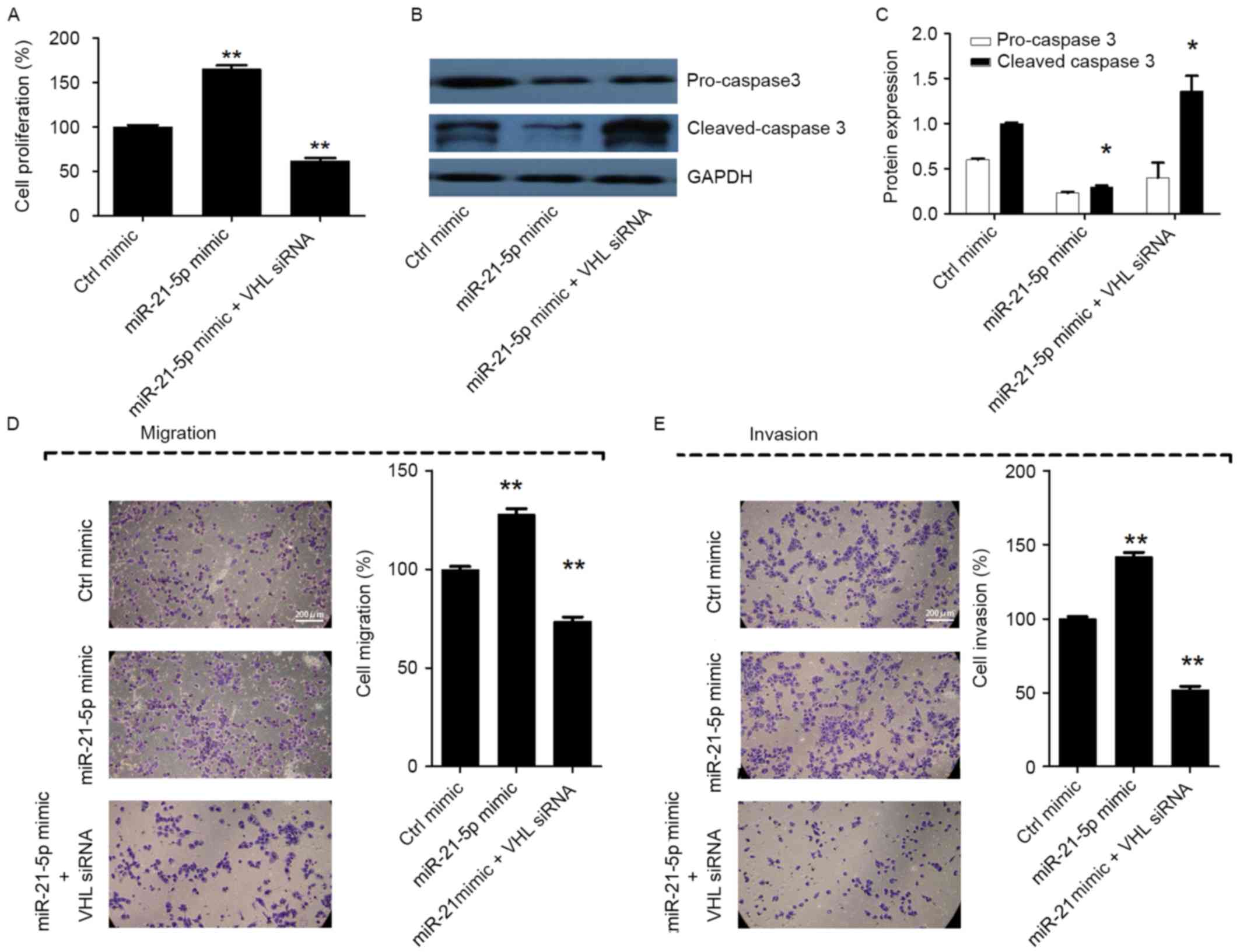
Silencing of miR-21-5p suppresses cell
proliferation, migration and invasion, and induces apoptosis by
targeting VHL in CC
In order to analyze whether miR-21-5p promotes cell
proliferation, migration and invasion, and inhibits apoptosis by
targeting VHL, miR-21-5p mimic and VHL siRNA were cotransfected
into CaSki cells. The results revealed that when cells were
co-transfected with miR-21-5p mimic and VHL siRNA, the cell
proliferation, migration and invasion were markedly decreased, and
cell apoptosis was increased, compared with the group transfected
with miR-21-5p mimic only (P<0.05; Fig. 4A-E). The results indicated that VHL
siRNA counteracted the effect of miR-21-5p on CaSki cells. Taken
together, these results suggest that silencing of miR-21-5p
suppresses cell proliferation, migration and invasion, and induces
apoptosis by upregulating VHL in CC cells.
Discussion
Current therapies for managing CC, one of the major
mortality factors in the female population, are typically
unsuccessful due to the invasive and metastatic phenotype of CC
cells. Previous studies have demonstrated that miR-21-5p is
upregulated in the pathogenesis of CC and other tumors (3,7,10,11,13).
Parallel studies have equally demonstrated loss or downregulation
of VHL in several types of cancer, particularly during oxidative
stress, one of the factors initiating and controlling cancer
progression and metastasis (14–16,18–22).
In the present study, the expression patterns of
miR-21-5p and VHL were assessed in CC cell lines. The results
demonstrated that miR-21-5p was upregulated in CC cell lines
compared with normal control cells. Additionally, as cell viability
and the occurrence of invasion and distant metastasis are important
factors that affect the prognosis and treatment of CC, it was
hypothesized that the upregulation of miR-21-5p may be involved in
the proliferation and metastasis of CC cells. Therefore, the effect
of miR-21-5p on the cell proliferation, migration, invasion and
apoptosis of CaSki cells was investigated. The results revealed
that inhibition of miR-21-5p expression significantly suppressed
CaSki cell proliferation, migration and invasion, and induced cell
apoptosis. This suggests that miR-21-5p may be associated with the
progression of CC malignancy. The results were corroborated by
previous studies, which produced similar results (3,9–12). Notably, it was identified that
miR-21-5p stimulates the proliferation and migration of CC cells
through inhibition of the PTEN pathway (3). However, the underlying molecular
mechanism of miR-21-5p in cancer, particularly CC, remains
unresolved.
Therefore, the objective of the present study was to
explore the association between miR-21-5p and VHL. The expression
level of VHL in CC cells was measured and it was revealed that
unlike miR-21-5p, the expression of VHL was downregulated in the CC
cell lines relative to the normal cells. Using bioinformatics a
putative binding site for miR-21-5p was identified in the 3′-UTR
region of VHL, which was validated as a direct target for miR-21-5p
by luciferase assay. Overexpression of miR-21-5p significantly
decreased the expression of VHL and the activity of the VHL 3′-UTR.
Furthermore, miR-21-5p mimics significantly promoted CaSki cell
proliferation, migration and invasion, and inhibited cell
apoptosis, whereas silencing of VHL counteracted these effects.
Taken together, these results suggest that miR-21-5p may act as an
oncogene by suppressing VHL expression in CC. To the best of our
knowledge, the present study is the first to disclose this
mechanistic and functional role of miR-21-5p in CC. A prior study
suggested that tuberous sclerosis complex protein 1 expression is
affected by VHL gene alterations and HIF-1α expression in renal
cell carcinoma (23). It was also
demonstrated that miR-566 regulates vascular endothelial growth
factor (VEGF) by targeting VHL in human glioblastoma in
vitro and in vivo (24).
Another study suggested that hsa-miR-331-3p downregulates VHL
expression in hepatocellular carcinoma cell lines by targeting its
3′-UTR (25), and that miR-101
targets VHL to promote HIF-1α-mediated apoptosis and cell cycle
arrest (26). Similarly, the
inhibitory effects of miR-185 on cell proliferation and its
activation of cell apoptosis by targeting VEGFA have been
attributed to VHL inactivation in clear cell renal cell carcinoma
(27). All these findings support the
results of the present study and indicate the role of VHL,
particularly the miR-21-5p/VHL axis, in tumorigenesis and cancer
progression.
In summary, the present study provides novel
evidence that the silencing of miR-21-5p inhibits the metastatic
phenotype of CC cells by downregulating VHL. The results also
suggest that miR-21-5p may be a novel therapeutic target for
CC.
References
|
1
|
Sherris J, Herdman C and Elias C: Cervical
cancer in the developing world. West J Med. 175:231–233. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Huang P, Xi J and Liu S: MiR-139-3p
induces cell apoptosis and inhibits metastasis of cervical cancer
by targeting NOB1. Biomed Pharmacother. 83:850–856. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Xu J, Zhang W, Lv Q and Zhu D:
Overexpression of miR-21 promotes the proliferation and migration
of cervical cancer cells via the inhibition of PTEN. Oncol Rep.
33:3108–3116. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Babashah S, Bakhshinejad B, Birgani MT,
Pakravan K and Cho WC: Regulation of microRNAs by phytochemicals: A
promising strategy for cancer chemoprevention. Curr Cancer Drug
Targets. Jun 23–2017.(Epub ahead of print). PubMed/NCBI
|
|
5
|
Zhang H, Li T, Zheng L and Huang X:
Biomarker MicroRNAs for diagnosis of oral squamous cell carcinoma
identified based on gene expression data and MicroRNA-mRNA network
analysis. Comput Math Methods Med. 2017:98030182017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Makiguchi T, Yamada M, Yoshioka Y, Sugiura
H, Koarai A, Chiba S, Fujino N, Tojo Y, Ota C, Kubo H, et al: Serum
extracellular vesicular miR-21-5p is a predictor of the prognosis
in idiopathic pulmonary fibrosis. Respir Res. 17:1102016.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Han Y, Xu GX, Lu H, Yu DH, Ren Y, Wang L,
Huang XH, Hou WJ, Wei ZH, Chen YP, et al: Dysregulation of miRNA-21
and their potential as biomarkers for the diagnosis of cervical
cancer. Int J Clin Exp Pathol. 8:7131–7139. 2015.PubMed/NCBI
|
|
8
|
Kowalczyk AE, Krazinski BE, Godlewski J,
Grzegrzolka J, Kiewisz J, Kwiatkowski P, Sliwinska-Jewsiewicka A,
Dziegiel P and Kmiec Z: SATB1 is down-regulated in clear cell renal
cell carcinoma and correlates with miR-21-5p overexpression and
poor prognosis. Cancer Genomics Proteomics. 13:209–217.
2016.PubMed/NCBI
|
|
9
|
Zheng G, Li N, Jia X, Peng C, Luo L, Deng
Y, Yin J, Song Y, Liu H, Lu M, et al: MYCN-mediated miR-21
overexpression enhances chemo-resistance via targeting CADM1 in
tongue cancer. J Mol Med (Berl). 94:1129–1141. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lopes-Ramos CM, Habr-Gama A, Quevedo Bde
S, Felício NM, Bettoni F, Koyama FC, Asprino PF, Galante PA,
Gama-Rodrigues J, Camargo AA, et al: Overexpression of miR-21-5p as
a predictive marker for complete tumor regression to neoadjuvant
chemoradiotherapy in rectal cancer patients. BMC Med Genomics.
7:682014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Park SK, Park YS, Ahn JY, Do EJ, Kim D,
Kim JE, Jung K, Byeon JS, Ye BD, Yang DH, et al: MiR 21–5p as a
predictor of recurrence in young gastric cancer patients. J
Gastroenterol Hepatol. 31:1429–1435. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bellissimo T, Russo E, Ganci F, Vico C,
Sacconi A, Longo F, Vitolo D, Anile M, Disio D, Marino M, et al:
Circulating miR-21-5p and miR-148a-3p as emerging non-invasive
biomarkers in thymic epithelial tumors. Cancer Biol Ther. 17:79–82.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang J, Yao T, Wang Y, Yu J, Liu Y and
Lin Z: Long noncoding RNA MEG3 is downregulated in cervical cancer
and affects cell proliferation and apoptosis by regulating miR-21.
Cancer Biol Ther. 17:104–113. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kim WY and Kaelin WG: Role of VHL gene
mutation in human cancer. J Clin Oncol. 22:4991–5004. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kaelin WG Jr: The von Hippel-Lindau tumor
suppressor gene and kidney cancer. Clin Cancer Res. 10:6290s–6295s.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kaelin WG: The von Hippel-Lindau tumor
suppressor protein: Roles in cancer and oxygen sensing. Cold Spring
Harb Symp Quant Biol. 70:159–166. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Neal CS, Michael MZ, Rawlings LH, Van der
Hoek MB and Gleadle JM: The VHL-dependent regulation of microRNAs
in renal cancer. BMC Med. 8:642010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu X, Cai X, Hu B, Mei Z, Zhang D, Ouyang
G, Wang J, Zhang W and Xiao W: Forkhead transcription factor 3a
(FOXO3a) modulates hypoxia signaling via up-regulation of von
Hippel-Lindau gene (VHL). J Biol Chem. 291:25692–25705. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Welford SM, Dorie MJ, Li X, Haase VH and
Giaccia AJ: Renal oxygenation suppresses VHL loss-induced
senescence that is caused by increased sensitivity to oxidative
stress. Mol Cell Biol. 30:4595–4603. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chetram MA, Bethea DA, Odero-Marah VA,
Don-Salu-Hewage AS, Jones KJ and Hinton CV: ROS-mediated activation
of AKT induces apoptosis via pVHL in prostate cancer cells. Mol
Cell Biochem. 376:63–71. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
LaGory EL, Wu C, Taniguchi CM, Ding CC,
Chi JT, von Eyben R, Scott DA, Richardson AD and Giaccia AJ:
Suppression of PGC-1α is critical for reprogramming oxidative
metabolism in renal cell carcinoma. Cell Rep. 12:116–127. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Damjanovic SS, Ilic BB, Beleslin Cokic BB,
Antic JA, Bankovic JZ, Milicevic IT, Rodic GS, Ilic DS, Todorovic
VN, Puskas N and Tulic CD: Tuberous sclerosis complex protein 1
expression is affected by VHL Gene alterations and HIF-1α
production in sporadic clear-cell renal cell carcinoma. Exp Mol
Pathol. 101:323–331. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xiao B, Zhou X, Ye M, Lv S, Wu M, Liao C,
Han L, Kang C and Zhu X: MicroRNA566 modulates vascular endothelial
growth factor by targeting Von HippelLandau in human glioblastoma
in vitro and in vivo. Mol Med Rep. 13:379–385. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cao Y, Zhang J, Xiong D, Wang D, Wu T,
Huang A and Tang H: Hsa-miR-331-3p inhibits VHL expression by
directly targeting its mRNA 3′-UTR in HCC cell lines. Acta Biochim
Pol. 62:77–82. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu N, Xia WY, Liu SS, Chen HY, Sun L, Liu
MY, Li LF, Lu HM, Fu YJ, Wang P, et al: MicroRNA-101 targets von
Hippel-Lindau tumor suppressor (VHL) to induce HIF1α mediated
apoptosis and cell cycle arrest in normoxia condition. Sci Rep.
6:204892016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ma X, Shen D, Li H, Zhang Y, Lv X, Huang
Q, Gao Y, Li X, Gu L, Xiu S, et al: MicroRNA-185 inhibits cell
proliferation and induces cell apoptosis by targeting VEGFA
directly in von Hippel-Lindau-inactivated clear cell renal cell
carcinoma. Urol Oncol. 33:169.e1–11. 2015. View Article : Google Scholar
|