Introduction
Integrins are transmembrane glycoproteins that
consist of an α subunit and a β subunit. A total of eight different
β subunits may dimerize, in limited combinations, with 18α subunits
to form ≥24 distinct integrins (1,2). Specific
integrin heterodimers preferentially bind to distinct extracellular
matrix (ECM) proteins. Integrins may bind the ligands in ECM
directly, including fibronectin and laminin, to affect the
characteristics of cells or the components of ECM. Regarding cancer
cells, integrins serve roles in numerous aspects, including
proliferation, survival, migration and invasion (1).
Integrins primarily affect cells in two ways, the
first is through binding with proteins directly, including talin,
vinculin and filamin, which may regulate the actin cytoskeleton of
cells (3). The other is by
phosphorylating the relative kinases, including focal adhesion
kinases (FAKs), proto-oncogene tyrosine-protein kinase (Src)-family
kinases (SFKs) and integrin-linked kinase (ILK), to activate or
cooperate with the other cell signaling pathways (1,4).
Additionally, integrin clustering on the cell surface and
trafficking from the endosomes may affect the ligand affinity and
quantity of the protein on cell surface (5–7).
Among the different integrins, β1 and β3 integrins
serve essential roles in the progression of different types of
cancer (1,2). Furthermore, previous studies
investigating the association between these two integrins have
demonstrated many different perspectives (8–11),
together providing a novel and more comprehensive understanding of
cancer.
Functions of β1 integrin in cancer
In tumors, the β1 subunit of integrin may combine
with different α subunits, including α4, α5 and α2, to affect the
characteristics of cancer cells, and the progression of tumors
(1). The primary function of β1
integrin is to form focal adhesion between cancer cells and ECM.
This adhesion is the basis for the survival of cancer cells and is
also associated with their migratory and metastatic capabilities
(2). There are series of proteins in
the cytoplasm, including talin, kindlin and ILK, which can affect
the ligand affinity and activation of integrin, subsequently
regulating focal adhesion and various characteristics of cancer
cells, including invasion and metastasis (1,3,4). Simultaneously, β1 integrin binding with
variant ligands in ECM, including laminin-1 and fibronectin, may
induce the secretion of certain cytokines and the progression of
tumor (12). Furthermore, a previous
study demonstrated that β1 integrin may also affect cell-cell
junctions (11). β1 integrin may also
affect the function of transforming growth factor-β (TGF-β) and
regulate microRNA-200/zinc finger E-box-binding homeobox 2 to
facilitate the expression of epithelial (E)-cadherin, which forms
cell-cell junctions (9,11).
Proliferation and survival
Regarding numerous different types of cancer cells,
the expression of β1 integrin may facilitate the growth of tumors.
β1 integrin on the surface of the cells, which does not bind with
the ECM, induces integrin-mediated death (IMD) of cells (1). However, when combined with their
ligands, this integrin promotes the survival of cancer cells by
activating different cell signaling proteins, including
phosphoinositide 3-kinase/RAC-α serine/threonine protein kinase
(AKT), FAKs and SFKs (1,13). While in certain tumors, β1 integrin
may also induce anoikis resistance of cancer cells in suspension by
phosphorylating FAKs and AKT (14,15).
Additionally, β1 integrin may promote the proliferation of cells by
phosphorylating FAKs and regulating SRC/mitogen-activated protein
kinase (MAPK) to facilitate the expression of v-myc avian
myelocytomatosis viral oncogene homolog (c-Myc) and cyclin D1 in
cancer cells (16,17). Additionally, although the
characteristics of the association between β1 integrin and cancer
stem cells are unclear, a previous study has identified that the
expression of β1 integrin in cancer stem cells is upregulated
(18).
Metastasis
Regarding metastasis, the effect of β1 integrin is
controversial. A previous study suggested that mutant cellular
tumor antigen p53 promoted β1 integrin-dependent cell motility and
invasion by reusing α5β1 integrin, and epidermal growth factor
receptor (EGFR) through recycling endosomes to the tumor cell
surface, facilitating the metastasis of cancer cells (19,20).
However, this metastasis induced by β1 integrin depends on the
expression of EGFR and the phosphorylation of AKT, which is a
downstream signaling protein of EGFR (20). Furthermore, in certain types of
cancer, the data demonstrated that β1 integrin may activate Src or
phosphorylate p38 and AKT to affect urokinase-type plasminogen
activator (uPA), and matrix metalloprotease (MMP)-2, promoting the
metastasis of cancer cells (21,22).
Additionally, due to the lack of vasculature, cancer cells enhance
β1 integrin activity to induce vessel cooption by adhering to the
vascular basement membrane, thus providing immediate vasculature
structures for newly metastatic or locally invasive of cancer cells
(23).
In contrast, β1 integrin may cooperate with TGF-β to
enhance the expression of E-cadherin and inhibit metastasis in
breast cancer (11). Additionally,
downregulation of β1 integrin induces not only
epithelial-to-mesenchymal transition (EMT) and anokisis resistance
in cancer cells, but also the expression of MMP-9, and vascular
endothelial growth factor (VEGF), promoting migration and invasion
(10).
Prognosis and clinical features
Similar to the controversy concerning metastasis,
the associations between β1 integrin and the clinical features of
patients are unclear. In certain types of cancer, β1 integrin is
associated with poor prognosis or metastasis, including in
prostatic cancer (15,24), melanoma (25), gastric carcinoma (26) and hypopharyngeal squamous cell
carcinoma (27). However, in other
types of cancer, including breast cancer, studies have reported
different or contradictory conclusions (28–30). This
suggests that additional clinical observations and studies are
required to understand this association.
Functions of β3 integrin in cancer
Regarding β3 integrin in tumors, the subtype that
exhibits the widest range of functions is αvβ3 integrin (1). It is associated with the growth,
survival, invasion and metastasis of different cancer cells
(10,31–35).
Furthermore, in certain types of cancer, it is an indicator of
increased lymph node or bone metastasis and decreased patient
survival (36–38).
Similar to β1 integrin, β3 integrin may also bind
with the components of ECM to form focal adhesions between cancer
cells and the ECM. Concomitantly, in suspension, β3 integrin
prevents cancer cells from IMD by activating a non-canonical
FAK-independent signaling pathway (31). Additionally, cancer cells as well as
somatic cells, including endothelial cells, have been demonstrated
to affect the growth of tumors by regulating angiogenesis (33).
Tumor growth and initiation
Regarding tumor growth, β3 integrin serves roles in
cancer cell survival, tumor initiation and tumor stemness,
primarily by regulating cytokines (31,34).
A previous study demonstrated that β3 integrin was
associated with cancer stem cells (39). An additional study identified that,
mechanistically, αvβ3 integrin in the unbound state recruits GTPase
KRas and Ras-related protein Ral-B to the tumor cell plasma
membrane, leading to the activation of TANK binding kinase 1 and
nuclear factor-κB (34). These two
proteins are necessary and sufficient for tumor initiation,
anchorage independence, and self-renewal (34). Additionally, in the β3 signaling
pathway, the receptor tyrosine kinase (RTK) is unnecessary for the
survival of cancer cells; therefore, cancer cells may survive using
β3 integrin, without RTK, which will induce resistance to RTK
inhibition therapy (34).
Furthermore, in a suspension of pancreatic cancer cells, β3
integrin was revealed to activate Src directly without
phosphorylating FAKs, to facilitate the survival of tumor cells
(31). Simultaneously, β3 integrin in
endothelial cells can affect the growth of tumors by regulating
angiogenesis (33). A study reported
that β3 integrin decreases the expression of VEGF receptor 2
(VEGFR2), thus inhibiting VEGF/VEGFR-induced angiogenesis and tumor
growth (33). These results suggest
that β3 integrin exhibits the ability to regulate RTK; however, the
features of the association remain unclear (40).
EMT and metastasis
EMT affects the metastasis of tumors in different
ways, including through migration, invasion and adhesion. β3
integrin has been demonstrated to be associated with EMT in cancer
cells. The integrin cooperates with TGF-β to form β3 integrin-TGF-β
receptor (TβR) type II complexes, which may activate TβR-II through
the β3 integrin/SRC signaling pathway and induce EMT by activating
MAPKs (32). Furthermore, β3 integrin
elevates the expression of MMP-9 and VEGF in cancer cells,
contributing to autocrine TGF-β signaling, and activation of EMT
processes (10). Additionally, αvβ3
integrin facilitates FAKs in regulating actin cytoskeleton
remodeling and the induction of EMT (41). Concomitantly, β3 integrin has the
ability to activate canonical FAKs-dependent cytokines to affect
cell migration and invasion (35). In
addition, β3 integrin is able to contribute to anchorage
independence, thus facilitating cancer cell survival and increasing
tumor malignancy, including lymph node metastasis (31). However, due to the co-expression of αv
and β3 subunits in cancer cells, future studies are warranted to
identify which of these factors are essential for specific
characteristics of cancer cells.
Ratio between β1 and β3 integrins in cancer
cells
Regarding cancer cells, the expression levels of β1
integrin and β3 integrin are associated; and changes to either
integrin exhibit significant effects on cancer cells (9–11)
(Fig. 1). In breast cancer, the
inactivation of β1 integrin elicits the robust compensatory
expression of β3 integrin (10,11).
However, the inhibition of β1 integrin cannot induce the
compensatory β3 integrin expression in normal mammary epithelial
cells (10). Furthermore, this
compensatory β3 integrin expression is essential for the growth and
metastasis of tumors (10). A
previous study demonstrated that when downregulating the expression
of β1 and β3 integrins simultaneously, cancer cell survival was
reduced (11). This suggests that,
perhaps, when β1 integrin is inactivated, the overexpression of β3
integrin is necessary and important to maintain the survival, and
characteristics of cancer cells.
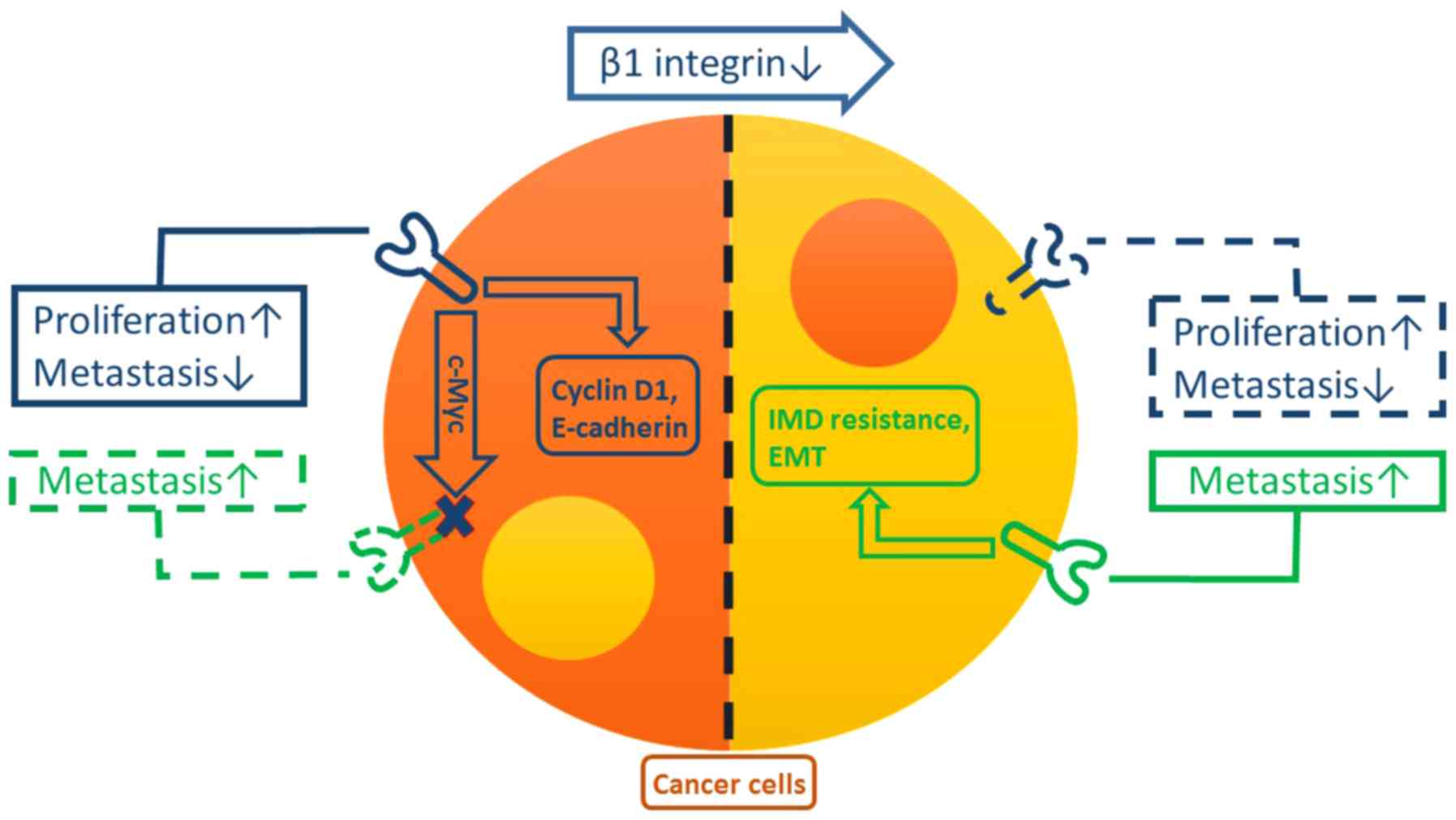 | Figure 1.Functions of, and the association
between, β1 and β3 integrins in cancer cells. β1 integrin (blue)
may facilitate cyclin D1 and E-cadherin in promoting proliferation
and reducing the rate of metastasis in cancer cells, respectively.
Concurrently, β1 integrin inhibits the expression of β3 integrin
(green) in cancer cells, probably by the regulation of c-Myc.
However, reductions in levels of β1 integrin induce the
compensatory expression of β3 integrin, which promotes EMT and IMD
resistance in cancer cells, resulting in metastasis. EMT,
epithelial-mesenchymal transition; IMD, integrin-mediated death;
E-cadherin, epithelial cadherin; c-Myc, v-myc avian
myelocytomatosis viral oncogene homolog. |
In addition, this ratio between β1 and β3 integrin
activity is not only involved for maintaining the functions of
cancer cells, but also for changing the survival and metastasis
rates of tumors. In pancreatic cancer, β1 integrin is able
facilitate the growth of tumors, and its inhibition reduces the
proliferation of cancer cells (11).
Additionally, the inhibition of β1 integrin may affect the function
of TGF-β, therefore attenuating the expression of E-cadherin to
reduce the activity of the cell-cell junctions and enhancing
motility and migration (11).
Nonetheless, whether β3 integrin is involved in these processes is
controversial (10,11). In contrast, β3 integrin can induce EMT
(9–11)
and activate a non-canonical FAKs-independent signaling pathway,
thus preventing cancer cells from undergoing IMD (31), and promoting the metastasis of cancer
cells. Therefore, according to these studies, the functions of β1
integrin are the promotion of proliferation and inhibition of
metastasis, and the functions of β3 integrin are opposing. This
indicates that proliferation and metastasis of cancer cells are not
always parallel, and may be considered independently maintained.
Furthermore, the association between clinical features, including
prognosis, and these two integrins, is complicated and
paradoxical.
Targeting therapies
The expression of β1 and β3 integrins in cancer is
involved in tumor progression, and various other pathways, which
suggests that they are potential therapeutic targets. In
preclinical studies, the antagonists of β1 or β3 integrin
effectively inhibited tumor growth by affecting tumor cells and
tumor-associated host cells (33,40,42–45).
These antagonists include monoclonal antibodies and
arginylglycylaspartic acid (RGD) peptide mimetics, which mimic the
structure of the RGD sequence in the ligands, and inhibit the
binding of integrins with their ligands. Furthermore, certain
antagonists have been demonstrated to be effective in clinical
trials (1).
Cilengitide, a RGD peptide mimetic, inhibits the
function of αvβ3 integrin and lengthens the survival time of
patients with certain types of cancer with minimal side effects in
clinical trials (46,47). Nevertheless, in vivo, specific
studies identified that the continuous infusion of low doses of RGD
peptides stimulates tumor growth and angiogenesis by increasing
VEGFR2 recycling to the endothelial cells membrane, and promoting
VEGF-induced migration (33,40). The angiogenesis of tumors may increase
the delivery of chemotherapeutic agents to the target areas, which
may explain why the combination of cilengitide and chemotherapy is
more effective compared with chemotherapy alone (1).
Regarding monoclonal antibodies, etaracizumab, a
function-blocking monoclonal antibody of αvβ3 integrin, has
demonstrated anti-angiogenic activity, direct inhibition of the
tumor cell growth and a reduction in bone metastasis rates in
preclinical studies (43,45). In clinical trials, etaracizumab also
demonstrated antiangiogenic activity, low toxicity and disease
stabilization in patients with certain types of cancer (48,49). In
addition, in preclinical and clinical trials, volociximab, a
function-blocking monoclonal antibody against integrin α5β1, has
exhibited efficacy in inhibiting angiogenesis and tumor growth
(42,50).
In specific previous studies, variant peptide
antagonists have been developed. For example, ATN-161 is a
non-RGD-based peptide inhibitor of integrin α5β1 that inhibits
cancer growth and metastasis in vivo (44). It also prolongs disease stabilization
in patients with advanced solid tumors (51).
Future perspectives
Certain previous studies have demonstrated that the
ratio between β1 and β3 integrins activity is associated with
specific important cytokines in cancer cells (8,10,17). A previous study has also suggested
that bound β1 integrin may activate Src and extracellular
signal-related kinase 1/2 MAPK in mammary epithelial cells, which
induces the overexpression of c-Myc (17). An additional previous study revealed
that MYC repressed transcription of the two subunits of αvβ3
integrin, thus suppressing cancer metastasis in breast cancer cells
(8). However, the lack of definitive
data makes identifying the association between MYC, and the ratio
between β1 and β3 integrins in cancer cells difficult. The details
of the association between these two integrins require additional
study.
Regarding clinical features, due to the association
between these two integrins, relying on one of them to evaluate the
status of patient is not comprehensive. The combination of β1 and
β3 integrins for prognosis and treatment is necessary. In addition,
with reference to previous studies, targeted therapy also requires
the combination of these two targets, or a focus on the common
downstream cytokines, including FAKs and SFKs, to increase
effectiveness.
Conclusion
β1 and β3 integrins are essential focal adhesion
proteins in various cancer cell types, which may affect the
initiation, proliferation, survival and metastasis of tumors.
Previous studies have demonstrated a ratio between these two
integrins in cancer cells, with contradictory functions. This
indicates that perhaps the proliferation and metastasis of cancer
cells are not always parallel; therefore, may be considered
independently maintained. Furthermore, the association between
clinical features and these integrins is more complicated than
previously expected. Therefore, there is a requirement for
additional clinical and experimental studies to elucidate the role
of these notable proteins.
Acknowledgements
The present study was supported by the National
High-tech R&D Program (863 Program; grant no.
2014AA020609).
References
|
1
|
Desgrosellier JS and Cheresh DA: Integrins
in cancer: Biological implications and therapeutic opportunities.
Nat Rev Cancer. 10:9–22. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Howe GA and Addison CL: β1 integrin: An
emerging player in the modulation of tumorigenesis and response to
therapy. Cell Adh Migr. 6:71–77. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Calderwood DA, Campbell ID and Critchley
DR: Talins and kindlins: Partners in integrin-mediated adhesion.
Nat Rev Mol Cell Biol. 14:503–517. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Legate KR, Montañez E, Kudlacek O and
Fässler R: ILK, PINCH and parvin: The tIPP of integrin signalling.
Nat Rev Mol Cell Biol. 7:20–31. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ewald AJ and Egeblad M: Cancer:
Sugar-coated cell signalling. Nature. 511:298–299. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Paszek MJ, DuFort CC, Rossier O, Bainer R,
Mouw JK, Godula K, Hudak JE, Lakins JN, Wijekoon AC, Cassereau L,
et al: The cancer glycocalyx mechanically primes integrin-mediated
growth and survival. Nature. 511:319–325. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Reverter M, Rentero C, Garcia-Melero A,
Hoque M, Vilà de Muga S, Alvarez-Guaita A, Conway JR, Wood P,
Cairns R, Lykopoulou L, et al: Cholesterol regulates Syntaxin 6
trafficking at trans-Golgi network endosomal boundaries. Cell Rep.
7:883–897. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu H, Radisky DC, Yang D, Xu R, Radisky
ES, Bissell MJ and Bishop JM: MYC suppresses cancer metastasis by
direct transcriptional silencing of αv and β3 integrin subunits.
Nat Cell Biol. 14:567–574. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Madamanchi A, Zijlstra A and Zutter MM:
Flipping the switch: Integrin switching provides metastatic
competence. Sci Signal. 7:pe92014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Parvani JG, Galliher-Beckley AJ, Schiemann
BJ and Schiemann WP: Targeted inactivation of β1 integrin induces
β3 integrin switching, which drives breast cancer metastasis by
TGF-β. Mol Biol Cell. 24:3449–3459. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Truong HH, Xiong J, Ghotra VP, Nirmala E,
Haazen L, Le Dévédec SE, Balcioğlu HE, He S, Snaar-Jagalska BE,
Vreugdenhil E, et al: β1 integrin inhibition elicits a
prometastatic switch through the TGFβ-miR-200-ZEB network in
E-cadherin-positive triple-negative breast cancer. Sci Signal.
7:ra152014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Grzesiak JJ, Smith KC, Burton DW, Deftos
LJ and Bouvet M: Integrin-mediated laminin-1 adhesion upregulates
CXCR4 and IL-8 expression in pancreatic cancer cells. Surgery.
141:804–814. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Aoudjit F and Vuori K: Integrin signaling
inhibits paclitaxel-induced apoptosis in breast cancer cells.
Oncogene. 20:4995–5004. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jin JK, Tien PC, Cheng CJ, Song JH, Huang
C, Lin SH and Gallick GE: Talin1 phosphorylation activates β1
integrins: A novel mechanism to promote prostate cancer bone
metastasis. Oncogene. 34:1811–1821. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lee YC, Jin JK, Cheng CJ, Huang CF, Song
JH, Huang M, Brown WS, Zhang S, Yu-Lee LY, Yeh ET, et al: Targeting
constitutively activated β1 integrins inhibits prostate cancer
metastasis. Mol Cancer Res. 11:405–417. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bartolome RA, Barderas R, Torres S,
Fernandez-Aceñero MJ, Mendes M, García-Foncillas J, Lopez-Lucendo M
and Casal JI: Cadherin-17 interacts with α2β1 integrin to regulate
cell proliferation and adhesion in colorectal cancer cells causing
liver metastasis. Oncogene. 33:1658–1669. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Benaud CM and Dickson RB: Regulation of
the expression of c-Myc by beta1 integrins in epithelial cells.
Oncogene. 20:759–768. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhu J, He J, Liu Y, Simeone DM and Lubman
DM: Identification of glycoprotein markers for pancreatic cancer
CD24+CD44+ stem-like cells using nano-LC-MS/MS and tissue
microarray. J Proteome Res. 11:2272–2281. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Muller PA, Caswell PT, Doyle B, Iwanicki
MP, Tan EH, Karim S, Lukashchuk N, Gillespie DA, Ludwig RL,
Gosselin P, et al: Mutant p53 drives invasion by promoting integrin
recycling. Cell. 139:1327–1341. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Selivanova G and Ivaska J: Integrins and
mutant p53 on the road to metastasis. Cell. 139:1220–1222. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ashour AA, Gurbuz N, Alpay SN, Abdel-Aziz
AA, Mansour AM, Huo L and Ozpolat B: Elongation factor-2 kinase
regulates TG2/β1 integrin/Src/uPAR pathway and
epithelial-mesenchymal transition mediating pancreatic cancer cells
invasion. J Cell Mol Med. 18:2235–2251. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Huang WS, Chin CC, Chen CN, Kuo YH, Chen
TC, Yu HR, Tung SY, Shen CH, Hsieh YY, Guo SE, et al: Stromal
cell-derived factor-1/CXC receptor 4 and β1 integrin interaction
regulates urokinase-type plasminogen activator expression in human
colorectal cancer cells. J Cell Physiol. 227:1114–1122. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jahangiri A, Aghi MK and Carbonell WS: β1
integrin: Critical path to antiangiogenic therapy resistance and
beyond. Cancer Res. 74:3–7. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Pontes-Junior J, Reis ST, Bernardes FS,
Oliveira LC, Barros ÉA, Dall'Oglio MF, Timosczuk LM, Ribeiro-Filho
LA, Srougi M and Leite KR: Correlation between β1 integrin
expression and prognosis in clinically localized prostate cancer.
Int Braz J Urol. 39:335–343. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Danen EH, Ten Berge PJ, Van Muijen GN,
Van't Hof-Grootenboer AE, Bröcker EB and Ruiter DJ: Emergence of
alpha 5 beta 1 fibronectin- and alpha v beta 3 vitronectin-receptor
expression in melanocytic tumour progression. Histopathology.
24:249–256. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Matsuoka T, Yashiro M, Nishimura S, Inoue
T, Fujihara T, Sawada T, Kato Y, Seki S and Hirakawa-Ys Chung K:
Increased expression of alpha2beta1-integrin in the peritoneal
dissemination of human gastric carcinoma. Int J Mol Med. 5:21–25.
2000.PubMed/NCBI
|
|
27
|
Hong YM, Gan WG and Xu ZH: Significance of
the expression of integrin β1, VEGF and MVD in hypopharyngeal
squamous cell carcinoma. Genet Mol Res. 13:6455–6465. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
dos Santos PB, Zanetti JS, Ribeiro-Silva A
and Beltrão EI: Beta 1 integrin predicts survival in breast cancer:
A clinicopathological and immunohistochemical study. Diagn Pathol.
7:1042012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Gonzalez MA, Pinder SE, Wencyk PM, Bell
JA, Elston CW, Nicholson RI, Robertson JF, Blamey RW and Ellis IO:
An immunohistochemical examination of the expression of E-cadherin,
alpha- and beta/gamma-catenins, and alpha2- and beta1-integrins in
invasive breast cancer. J Pathol. 187:523–529. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Restucci B, De Vico G and Maiolino P:
Expression of beta 1 integrin in normal, dysplastic and neoplastic
canine mammary gland. J Comp Pathol. 113:165–173. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Desgrosellier JS, Barnes LA, Shields DJ,
Huang M, Lau SK, Prévost N, Tarin D, Shattil SJ and Cheresh DA: An
integrin alpha(v)beta(3)-c-Src oncogenic unit promotes
anchorage-independence and tumor progression. Nat Med.
15:1163–1169. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
32
|
Galliher AJ and Schiemann WP: Beta3
integrin and Src facilitate transforming growth factor-beta
mediated induction of epithelial-mesenchymal transition in mammary
epithelial cells. Breast Cancer Res. 8:R422006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Reynolds LE, Wyder L, Lively JC, Taverna
D, Robinson SD, Huang X, Sheppard D, Hynes RO and Hodivala-Dilke
KM: Enhanced pathological angiogenesis in mice lacking beta3
integrin or beta3 and beta5 integrins. Nat Med. 8:27–34. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Seguin L, Kato S, Franovic A, Camargo MF,
Lesperance J, Elliott KC, Yebra M, Mielgo A, Lowy AM, Husain H, et
al: An integrin β3 KRAS-RalB complex drives tumour stemness and
resistance to EGFR inhibition. Nat Cell Biol. 16:457–468. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zheng DQ, Woodard AS, Fornaro M, Tallini G
and Languino LR: Prostatic carcinoma cell migration via
alpha(v)beta3 integrin is modulated by a focal adhesion kinase
pathway. Cancer Res. 59:1655–1664. 1999.PubMed/NCBI
|
|
36
|
Hosotani R, Kawaguchi M, Masui T, Koshiba
T, Ida J, Fujimoto K, Wada M, Doi R and Imamura M: Expression of
integrin alphaVbeta3 in pancreatic carcinoma: Relation to MMP-2
activation and lymph node metastasis. Pancreas. 25:e30–e35. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
McCabe NP, De S, Vasanji A, Brainard J and
Byzova TV: Prostate cancer specific integrin alphavbeta3 modulates
bone metastatic growth and tissue remodeling. Oncogene.
26:6238–6243. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Sloan EK, Pouliot N, Stanley KL, Chia J,
Moseley JM, Hards DK and Anderson RL: Tumor-specific expression of
alphavbeta3 integrin promotes spontaneous metastasis of breast
cancer to bone. Breast Cancer Res. 8:R202006. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Vaillant F, Asselin-Labat ML, Shackleton
M, Forrest NC, Lindeman GJ and Visvader JE: The mammary progenitor
marker CD61/beta3 integrin identifies cancer stem cells in mouse
models of mammary tumorigenesis. Cancer Res. 68:7711–7717. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Robinson SD and Hodivala-Dilke KM: The
role of β3-integrins in tumor angiogenesis: Context is everything.
Curr Opin Cell Biol. 23:630–637. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Shah PP, Fong MY and Kakar SS: PTTG
induces EMT through integrin αVβ3-focal adhesion kinase signaling
in lung cancer cells. Oncogene. 31:3124–3135. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Besse B, Tsao LC, Chao DT, Fang Y, Soria
JC, Almokadem S and Belani CP: Phase Ib safety and pharmacokinetic
study of volociximab, an anti-α5β1 integrin antibody, in
combination with carboplatin and paclitaxel in advanced
non-small-cell lung cancer. Ann Oncol. 24:90–96. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Gramoun A, Shorey S, Bashutski JD, Dixon
SJ, Sims SM, Heersche JN and Manolson MF: Effects of Vitaxin, a
novel therapeutic in trial for metastatic bone tumors, on
osteoclast functions in vitro. J Cell Biochem. 102:341–352. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Khalili P, Arakelian A, Chen G, Plunkett
ML, Beck I, Parry GC, Doñate F, Shaw DE, Mazar AP and Rabbani SA: A
non-RGD-based integrin binding peptide (ATN-161) blocks breast
cancer growth and metastasis in vivo. Mol Cancer Ther. 5:2271–2280.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Mulgrew K, Kinneer K, Yao XT, Ward BK,
Damschroder MM, Walsh B, Mao SY, Gao C, Kiener PA, Coats S, et al:
Direct targeting of alphavbeta3 integrin on tumor cells with a
monoclonal antibody, Abegrin. Mol Cancer Ther. 5:3122–3129. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Reardon DA, Fink KL, Mikkelsen T,
Cloughesy TF, O'Neill A, Plotkin S, Glantz M, Ravin P, Raizer JJ,
Rich KM, et al: Randomized phase II study of cilengitide, an
integrin-targeting arginine-glycine-aspartic acid peptide, in
recurrent glioblastoma multiforme. J Clin Oncol. 26:5610–5617.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Vansteenkiste J, Barlesi F, Waller CF,
Bennouna J, Gridelli C, Goekkurt E, Verhoeven D, Szczesna A, Feurer
M, Milanowski J, et al: Cilengitide combined with cetuximab and
platinum-based chemotherapy as first-line treatment in advanced
non-small-cell lung cancer (NSCLC) patients: Results of an
open-label, randomized, controlled phase II study (CERTO). Ann
Oncol. 26:1734–1740. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Delbaldo C, Raymond E, Vera K,
Hammershaimb L, Kaucic K, Lozahic S, Marty M and Faivre S: Phase I
and pharmacokinetic study of etaracizumab (Abegrin), a humanized
monoclonal antibody against alphavbeta3 integrin receptor, in
patients with advanced solid tumors. Invest new Drugs. 26:35–43.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Gutheil JC, Campbell TN, Pierce PR,
Watkins JD, Huse WD, Bodkin DJ and Cheresh DA: Targeted
antiangiogenic therapy for cancer using Vitaxin: A humanized
monoclonal antibody to the integrin alphavbeta3. Clin Cancer Res.
6:3056–3061. 2000.PubMed/NCBI
|
|
50
|
Bhaskar V, Zhang D, Fox M, Seto P, Wong
MH, Wales PE, Powers D, Chao DT, Dubridge RB and Ramakrishnan V: A
function blocking anti-mouse integrin alpha5beta1 antibody inhibits
angiogenesis and impedes tumor growth in vivo. J Transl Med.
5:612007. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Cianfrocca ME, Kimmel KA, Gallo J, Cardoso
T, Brown MM, Hudes G, Lewis N, Weiner L, Lam GN, Brown SC, et al:
Phase 1 trial of the antiangiogenic peptide ATN-161
(Ac-PHSCN-NH(2)), a beta integrin antagonist, in patients with
solid tumours. Br J Cancer. 94:1621–1626. 2006. View Article : Google Scholar : PubMed/NCBI
|














