Introduction
Gastric cancer (GC) ranks fourth in incidence and
second as a cause of mortality among all types of cancer worldwide
(despite the decreased incidence in certain regions) (1). Surgical resection, chemotherapy and
radiation remain the most common therapeutic modalities. Surgical
resection is currently the only curative treatment for early-stage
GC. However, the majority of patients are diagnosed at advanced
disease stages or relapse following curative surgical treatment
(1).
Despite advances in the detection, surgical
resection and adjuvant therapy for GC, the 5-year survival rates of
these patients remain <30% (2).
The aggressive nature of human GC is associated with a variety of
intracellular events including activation of various oncogenes,
inactivation of tumor suppressor genes and abnormal expression of
growth factors and their receptors (3,4). These
perturbations result in a marked growth advantage for GC cells.
Thus, to improve the low survival outcomes and assist in earlier
diagnosis of patients with GC, identification and validation of new
prognostic and therapeutic tumor markers is urgently required.
These improved biomarkers may in turn provide new approaches for
the early detection and effective treatment of GC.
Endoplasmic reticulum (ER) stress and the
unfolded protein response (UPR)
The ER is an organelle responsible for the
synthesis, initial post-translational modification, folding, export
and secretion of proteins (5).
Disturbances in the ER environment by cellular stress conditions,
including nutrient deprivation, alterations in glycosylation
status, hypoxia, pH changes, poor vascularization, changes in
calcium homeostasis and treatment with a variety of agents, may
lead to ER stress and subsequent accumulation of unfolded or
misfolded proteins in the ER (6,7). To
overcome perturbations in ER function and ER stress to improve
survival, the ER has evolved specific signaling pathways, which are
collectively termed the UPR (8). The
UPR is initiated in concerted action through the signaling of three
prototypical ER-localized stress sensors: RNA-dependent protein
kinase-like ER kinase (PERK), activating transcription factor 6
(ATF6) and inositol-requiring enzyme 1 (IRE1) (6,9,10). Upon ER stress, ER-resident chaperones
[e.g., glucose-regulated protein (GRP) 78] bind to misfolded
proteins, activating IRE1, ATF6 and PERK. PERK is also activated by
dimerization and autophosphorylation, subsequently phosphorylates
eukaryotic initiation factor 2α (eIF2α). Phosphorylated eIF2α
inhibits protein synthesis and activates the transcription of ATF4,
inducing the transcription of its downstream genes (11–13). IRE1
assists in protein folding and degradation, and produces a spliced
form of X box-binding protein-1 due to its RNase activity. ATF6
translocates from the ER to the Golgi apparatus where it is cleaved
by protease activity, forming active nuclear ATF6, a regulator of
gene expression (14). Collectively,
ER stress is alleviated by the downstream effects of the UPR.
However, if ER stress is severe or prolonged, distinct death
signals may be transduced during the UPR, leading to cellular
apoptosis (15,16). These signals include
CCAAT/enhancer-binding protein homologous protein (CHOP)
transcription factor, p53 unregulated modulator of apoptosis
(PUMA), c-Jun N-terminal kinase (JNK), nicotinamide adenine
dinucleotide phosphate oxidase activator (NOXA), B-cell lymphoma 2
(Bcl-2)-like 11 and Bcl-2 homology (BH) 3-only proteins and
caspases (16–22).
ER stress and tumors
Tumor cells proliferate continuously and require
effective high-energy-producing systems due to their high
proliferation characteristics compared with non-tumorigenic cells.
Solid tumors typically grow faster than their blood supply is able
to nurture, creating specific growth conditions characterized by
hypoxia, glucose deprivation and lactic acidosis, which trigger ER
stress (23). The interactions
between cancerous cells and this tumor microenvironment during the
course of multistep tumorigenesis are reported to serve a critical
role in the modulation of tumor growth, metabolism and metastasis
(24–26).
ER stress has a dual effect on tumors. First, it has
an adaptive effect, enhancing tumor growth. ER stress may restore
homeostasis and make the adjacent environment hospitable for tumor
survival, growth and expansion (27).
Baird et al (28) demonstrated
that the UPR was induced and promoted the neoplastic transformation
of Helicobacter-infected gastric mucosa in the milieu of
Helicobacter-induced chronic inflammation and mucous
metaplasia. Pike et al (29)
revealed that under severely hypoxia conditions, ATF4
transcriptionally upregulated unc-51-like autophagy-activating
kinase 1 (ULK1), which is required for autophagy contributing to
human breast cancer cell survival. Hypoxia also induces breast
cancer cell migration via the PERK/ATF4/lysosome-associated
membrane protein 3 (LAMP3) signaling pathway (30). On the other hand, ER stress
contributes cytotoxic effects, inducing apoptosis. Signal
transducer and activator of transcription 6 (STAT6) silencing
elicited ER stress-mediated apoptosis in lung cancer cells through
CHOP induction, alteration of BH3 protein expression and reactive
oxygen species (ROS) production (31). Resveratrol
(3,5,4′-trihydroxy-trans-stilbene) exerted its cytotoxic role in
cancer cells exposed to palmitate, a saturated fatty acid,
triggering a lipid-mediated cell death, which is promoted by ER
stress through a CHOP-mediated apoptotic process (32). Lactacystin (LAC) treatment increases
the expression of protein disulfide-isomerase (PDI), GRP78, CHOP,
cleaved caspase-4 and cleaved caspase-3 induced by cisplatin in
HeLa cells, suggesting that LAC may enhance cisplatin cytotoxicity
by increasing ER stress-associated apoptosis (33).
GRP78 and its signaling pathway
GRP78, also known as the immunoglobulin heavy
chain-binding protein, and other GRPs are ER chaperones which
belong to the heat-shock protein family (34,35). In
the late 1970s, upon rapid depletion of glucose from the culture
medium of chick embryo fibroblasts, the amount of an unknown
protein with a molecular mass of 78 kDa was identified to be
significantly increased and was subsequently termed GRP78 (36). GRP78 was also revealed to be present
in the plasma membrane, cytoplasm, mitochondria, nucleus and the
cellular secretions of tumor cells (37). The GRP78 promoter region contains a
highly conserved region (consisting of CCAAT-like sequences flanked
by GC-rich motifs) termed the cis-acting endoplasmic
reticulum stress-response element (ERSE), reportedly required for
transcriptional activation in response to ER stress (38). The promoter also contains other
important motifs including a cyclic adenosine 3′,5′-monophosphate
response element (CRE) and 12-O-tetradecanoylphorbol-13-acetate
(TPA) DNA-response element (TRE) motif (39,40). As an
ER stress-associated protein, GRP78 is involved in protein folding
and assembly, proteasomal degradation of misfolded proteins, ER
Ca2+ binding and cell survival during damaging
conditions (41).
Under non-stress conditions, GRP78 binds to three
UPR sensors (PERK, IRE1 and ATF6) rendering them inactive. In
response to ER stress, GRP78 preferentially associates with the
unfolded proteins instead of the typical sensors (42), following which the UPR becomes
activated. The UPR sensors are important as they elicit damage
control pathways synergistically, partially due to the activation
by ATFs (ATF4 and ATF6). Nuclear form ATFs act on the ERSE,
increasing the expression of GRP78. Overexpression of GRP78 is
hypothesized to be redistributed to the cell surface by means of
vesicle transport (43). It
recognizes extracellular ligands including
α2-macroglobulin, kringle 5, prostate apoptosis
response-4 (Par-4), major histocompatibility complex I (MHC-I) and
T-cadherin, transducing corresponding signals. Ligands binding to
the N-terminal domain of GRP78 induce a proproliferative and
antiapoptotic response, whereas binding to the C-terminal domain
inhibits cell proliferation and triggers apoptosis (44). This biological effect is associated
with the activation of the phosphoinositide 3-kinase/protein kinase
B (PI3K/Akt) and mitogen-activated protein kinase (MAPK) signaling
pathways (45,46). The activation of the MAPK signaling
pathways leads to a rapid induction of GRP78 (47), presenting GRP78-associated signal
transduction in the form of a feedback loop (Fig. 1) (48).
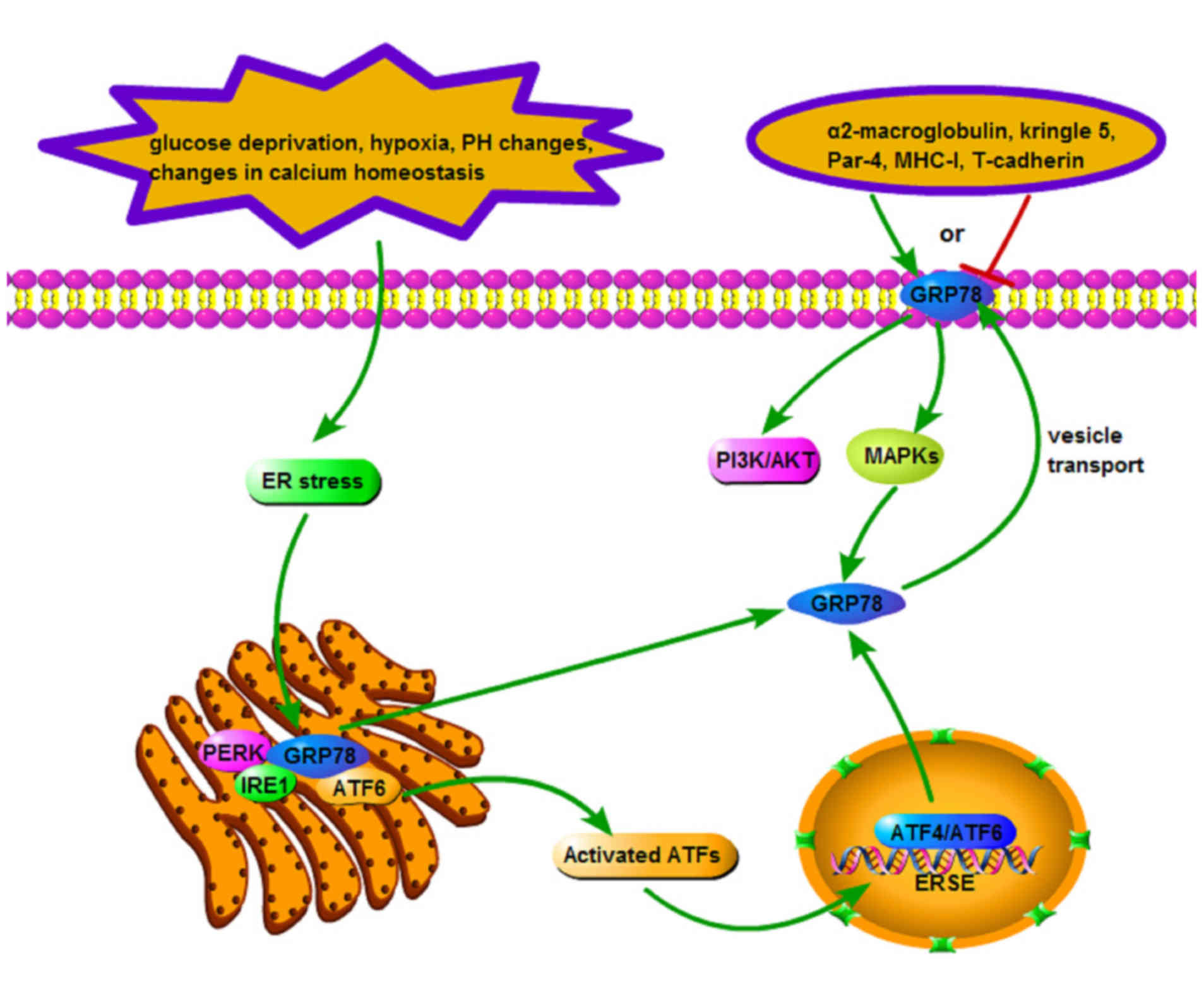 | Figure 1.Signaling pathway of GRP78. Upon ER
stress, GRP78 separates from UPR sensors (PERK, IRE1 and ATF6) and
preferentially associates with unfolded proteins, upon which UPR is
activated. These sensors elicit damage control pathways
synergistically, activated partially by ATFs (ATF4 and ATF6). The
nuclear form ATFs subsequently act on ERSE, increasing the
expression of GRP78. Overexpressed GRP78 may be redistributed to
the cell surface by means of vesicle transport, recognizing
extracellular ligands and activating PI3K/AKT and MAPK signaling
pathways. The activation of PI3K/AKT and MAPK signaling pathways
may also lead to the rapid induction of GRP78. GRP,
glucose-regulated protein; ER, endoplasmic reticulum; UPR, unfolded
protein response; PERK, protein kinase-like ER kinase; IRE1,
inositol-requiring enzyme 1; ATF, activating transcription factor;
ERSE, endoplasmic reticulum stress-response element; PI3 K,
phosphoinositide 3-kinase; AKT, protein kinase B; MAPK,
mitogen-activated protein kinase; MHC, major histocompatibility
complex; Par-4, prostate apoptosis response-4. |
GRP78 and gastric cancer
Numerous studies on GRP78 have been focused on tumor
development and progression. Overexpression of GRP78 has been
identified in a variety of tumors including digestive, urinary,
cerebral, mammary and respiratory system tumors. In general, GRP78
expression is positively associated with tumor malignancy. For
example, GRP78 expression increased with the progression from early
to advanced colorectal cancer stages (49). Furthermore, a significant association
was identified between GRP78 expression and response to
chemotherapy (49). Guan et al
(50) reported that GRP78 and
melanoma differentiation-associated gene-9 (MDA-9) were expressed
in lymph node metastases at increased levels. Furthermore, exosomes
from serum samples of patients with metastatic melanoma contained
increased levels of MDA-9 and GRP78 compared with patients without
metastases, indicating the potential of MDA-9 and GRP78 as
biomarkers for the early detection of metastasis. Caspases cause
poly (ADP-ribose) polymerase (PARP) cleavage and inactivation
during apoptosis. Jiang et al (51) demonstrated that knockdown of GRP78 by
small interfering (si)RNA enhanced PARP cleavage in human
pancreatic cancer cell lines. Conversely, induction of GRP78 on the
cell surface by doxorubicin and tunicamycin has been previously
associated with CHOP/growth arrest- and DNA damage-inducible gene
153 (GADD153) upregulation and increased apoptosis in
triple-negative breast cancer tumor cells (52). Overexpression of GRP78 is associated
with early clinical stage and improved survival in patients with
neuroblastoma (53). The following
subsections systematically and comprehensively summarize the
studies which have elucidated the role of GRP78 in GC.
GRP78 expression and its clinical
characteristics in GC
Numerous studies have identified that GRP78 is
detected in the sera of patients with GC along with its
autoantibody (54), and it is
significantly upregulated in GC cells as well as in surgical
specimens of gastric tumors (55–58). Other
studies have demonstrated that GRP78 is positively associated with
tumor size, depth of invasion, poor differentiation,
tumor-node-metastasis stage, lymphatic and venous invasion, lymph
node metastasis, short time to recurrence and chemoresistance,
although it is not associated with sex or age (55–58). In
the light of this evidence, GRP78 is considered an objective and
effective marker for predicting the aggressive behavior and poor
prognosis of patients with GC.
GRP78, cell proliferation and
apoptosis in GC
In a gastric tumor, cancer cells are able to adapt
to a variety of ER stressors by inducing GRP78, which promotes
cancer cell proliferation and inhibits apoptosis. For example, in
flow cytometry analysis, it had previously been revealed that the
downregulation of GRP78 markedly inhibited the proliferation of GC
cells at the G1 phase, whereas GRP78 overexpression promoted cell
cycle progression (55). These
results suggest that GRP78 promotes GC cell proliferation. Under
hypoxia stress, a protein kinase
Cε/Raf-1/MAPK-extracellular-signal-regulated kinase (ERK) kinase
(MEK)/ERK/activator protein 1 (AP1) signaling cascade induced GRP78
expression in human GC cells by acting on a TPA-response
element-like element of the GRP78 promoter (47). In the presence of the MEK inhibitor
U0126, activation of caspase-3 and cleavage of its substrate PARP
by the ER stress inducer tunicamycin or thapsigargin was enhanced
in GC cells, although overexpression of Bcl-2 inhibited this
apoptosis (59). Therefore, Zhang
et al (59) concluded that the
inhibition of MEK blocked the ER stress-mediated upregulation of
GRP78 and enhanced ER stress-induced apoptosis through a caspase-
and mitochondria-mediated mechanism. This activation of the MEK/ERK
signaling pathway by ER stress is hypothesized to be necessary for
the induction of GRP78, which protects GC cells against apoptosis
(59).
Conversely, if the stress exceeds the threshold that
GC cells can afford, apoptosis may be initiated. For instance,
following treatment with 10 µg/ml tunicamycin for 24 h, GRP78 and
CHOP were upregulated whereas Bcl-2 was downregulated in the GC
cell line BGC823, ultimately leading to apoptosis (60). Vitamin E succinate (RRR-α-tocopheryl
succinate; VES) causes cytological changes typical of apoptosis by
increasing ER dilation and cytosolic Ca2+ concentration.
Upon treatment with VES at a concentration of 20 µg/ml, GRP78 was
demonstrated to be transcriptionally and translationally induced in
a time-dependent manner while the induction of CHOP, caspase-4 and
JNK were observed (61). Huang et
al (62) additionally revealed
that, in response to α-tocopheryl succinate (α-TOS), induction of
GRP78 and CHOP and activation of caspase-4 were also observed which
are cytological changes typical of apoptosis.
GRP78 variants and promoter
polymorphisms in GC
Rauschert et al (63) isolated a human monoclonal IgM
antibody, SAM-6, from a patient with GC. The antibody was revealed
to bind a previously unknown variant of GRP78 with a molecular mass
of 82 kDa, a variant eventually known as GRP78SAM-6. The
epitope is an O-linked carbohydrate moiety which is only expressed
on malignant cell membranes. This variant qualifies as a target for
immune surveillance and antibody responses, making it an ideal
target for novel therapeutic approaches of patients with GC. Winder
et al (64) have additionally
reported that patients with GC with the combined GRP78 rs391957 C/T
and T/T genotype exhibit an increased risk of tumor recurrence and
mortality than those with C/C. These data suggest that GRP78
rs391957 polymorphism may be capable of predicting clinical
outcomes in patients with localized GC (64).
GRP78 and GC cell invasion and
metastasis
Previous studies have identified that the expression
of GRP78 has a clear association with the invasion and metastasis
of GC cells (55,65). Zhang et al (65) revealed that overexpression of GRP78,
induced by the transcription factor specificity protein 1 (Sp1),
increased lymph node metastasis in patients with GC. Knocking down
GRP78 expression inhibited GC cell invasion in vitro and
cellular proliferation and metastasis in a xenograft nude mouse
model. Yang et al (55)
similarly reported that GRP78 expression was increased in tumors
from GC patients with deep tumor infiltration and lymph node
metastasis compared with tumors from patients without these
features. These results suggest that GRP78 may promote invasion and
metastasis of GC cells and that the dysregulated expression of
GRP78 may contribute to the development and progression of GC.
GRP78 and chemoresistance of GC
cells
Over the last decades, standard multimodal treatment
strategies have failed to cure a large proportion of patients with
GC, particularly those with advanced and metastatic disease,
ultimately contributing to poor survival rates (1). Certain investigators have suggested that
this is possibly due to a chemoresistance phenomenon occurring
during treatment (66). For example,
an adenosine 5′-triphosphate tumor chemosensitivity assay has
demonstrated that increased GRP78 expression is associated with the
chemoresistance of GC cells to chemotherapeutic agents, whereas
negative GRP78 expression was associated with increased sensitivity
to drugs and regimens (56). However,
the underlying molecular mechanisms of this observed outcome remain
to be further clarified and are urgently required for more
effective clinical intervention and improved patient
management.
Celecoxib, a non-steroidal anti-inflammatory drug,
induces apoptosis in cancer cells. In human GC cells,
overexpression of GRP78 induced by celecoxib partially suppresses
the induction of CHOP and protects cancer cells from
celecoxib-induced apoptosis (67).
Additionally, suppression of GRP78 expression by siRNA markedly
stimulates the expression of CHOP and cellular apoptosis in the
presence of celecoxib. These results suggest that upregulation of
ER chaperones by celecoxib decreases the potential antitumor
activity of celecoxib.
GRP78, diagnosis and targeted therapy
of GC
Since no specific symptoms have been characterized
in patients with early GC, there is a lack of convenient means of
census screening, consequently contributing to the currently low
detection rate of patients with early GC. Radiation or chemotherapy
of patients with GC is often accompanied by enormous
biocytotoxicity and side effects, often resulting in poor patient
outcomes. Molecular targeting therapy has been proposed to
eradicate tumors through the targeting of specific tumor markers.
Uncovering reliable biomarkers of GC associated with tumorigenesis
and progression is crucial for effective diagnosis and successful
treatment, leading to improved therapeutic outcomes and patient
quality of life (68,69). Interestingly, the radioactive
intensity measured in animals with GC xenografts administered with
GRP78-binding peptide-guided 111In-labeled micelles is
statistically increased compared with animals administered with
111In-labeled micelles alone (70). These results suggest that GRP78 is an
effective probing target in the application of nuclear imaging for
GC diagnosis. Kang et al (71)
identified that GC multidrug resistance (MDR) cell-specific binding
peptide GMBP1 may specifically bind to GRP78 on the surface of GC
MDR cells, resensitizing GC MDR cells to a variety of
chemotherapeutic agents by downregulating GRP78 expression and
inhibiting MDR1 expression. These results provide new insight into
the management of MDR in GC cells.
Furthermore, versipelostatin (VST), a novel
macrocyclic compound, may inhibit transcription from the GRP78
promoter. VST alone and in combination with cisplatin significantly
inhibits tumor growth of GC cell MKN74 xenografts compared with
untreated controls (72). Cheng et
al (73) designed a GRP78-binding
peptide which may selectively recognize and bind to GC MKN45 cells
in vitro. Additionally, the overexpression of GRP78 has been
used as a targeted protein to guide drugs to GC cells, leading to a
more effective treatment for GC xenografts (73). This study demonstrated that a
GRP78-mediated drug targeting system may deliver chemotherapeutic
drugs with increased targeting precision to GC cells, leading to
minimized side effects in patients during chemotherapy (73).
Finally, the thymidine kinase gene of herpes simplex
virus (HSV-tk) is a suicide gene when administrated with the
prodrug ganciclovir (GCV). HSV-tk may phosphorylate GCV to become
GCV triphosphate which is incorporated into cellular DNA, resulting
in termination of DNA synthesis and cell death (74). In an experimental system, the HSV-tk
gene may be controlled by the GRP78 promoter or the long terminal
repeat (LTR) promoter. Under stress conditions in a fast-growing
solid tumor, the LTR promoter was suppressed and thus unable to
sustain the foreign gene expression (75); however, the stress-inducible protein
GRP78 was markedly induced (76).
This constitutes an ideal gene therapy system to selectively kill
tumor cells without affecting normal tissues. Azatian et al
(77) have demonstrated that,
compared with LTR-tk/GCV, GRP78-tk/GCV treatment resulted in
complete tumor elimination in GC cells with no p53 mutations in
vitro and in vivo (Table
I).
 | Table I.Anti-GRP78 drugs and their effects
and molecular mechanisms in GC. |
Table I.
Anti-GRP78 drugs and their effects
and molecular mechanisms in GC.
| Anti-GRP78
drug | Effect | Mechanism |
|---|
| GMBP1 | Resensitizes GC MDR
cells to chemotherapeutic agents | Downregulates GRP78
and MDR1 expression |
|
Versipelostatin | Inhibits tumor
growth of GC cell | Inhibits
transcription from the promoter of GRP78 |
| GRP78BP | A targeted protein
to guide drugs to GC cells | Selectively
recognizes and binds to GC cells |
| GRP78-tk/GCV | A suicide gene when
administrated with the prodrug GCV | The HSV-tk gene may
be controlled by GRP78 promoter |
Conclusions and perspective
The ER stress-associated protein GRP78 is
overexpressed in GC and promotes proliferation and inhibition of
apoptosis of GC cells. GRP78 may serve a critical role in GC cell
invasion and metastasis as well as the development of
chemotherapeutic resistance. Clinically, GRP78 expression has a
clear association with the prognosis of patients with GC. As a
biomarker of GC, GRP78 may improve the efficiency of early
diagnosis for patients with GC. As a therapeutic target,
GRP78-targeting therapy may improve therapeutic outcomes and
quality of life in patients with GC. However, the causal role of
GRP78 in GC pathogenesis and eventual translation into the clinic
warrants further study, particularly the aspects of the underlying
molecular mechanisms of action. Other functions of GRP78 in GC,
including angiogenesis, should also be explored in depth. Further
understanding of the roles of GRP78 in GC may provide physicians
broader prospects for the effective treatment of GC patients.
Acknowledgements
The preparation of the present review was supported
by the National Natural Science Foundation of China (grant no.
81401025) and the Youth Science and Research Foundation of Nantong
Municipal Planning Commission (grant no. WQ2016058).
References
|
1
|
Piazuelo MB and Correa P: Gastric cáncer:
Overview. Colomb Med (Cali). 44:192–201. 2013.PubMed/NCBI
|
|
2
|
Nagini S: Carcinoma of the stomach: A
review of epidemiology, pathogenesis, molecular genetics and
chemoprevention. World J Gastrointest Oncol. 4:156–169. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ma Y and Hendershot LM: The role of the
unfolded protein response in tumour development: Friend or foe? Nat
Rev Cancer. 4:966–977. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lee AS: The glucose-regulated proteins:
Stress induction and clinical applications. Trends Biochem Sci.
26:504–510. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Schmitz A and Herzog V: Endoplasmic
reticulum-associated degradation: Exceptions to the rule. Eur J
Cell Biol. 83:501–509. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Harding HP, Calfon M, Urano F, Novoa I and
Ron D: Transcriptional and translational control in the Mammalian
unfolded protein response. Annu Rev Cell Dev Biol. 18:575–599.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Schröder M and Kaufman RJ: The mammalian
unfolded protein response. Annu Rev Biochem. 74:739–789. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kaufman RJ: Orchestrating the unfolded
protein response in health and disease. J Clin Invest.
110:1389–1398. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ma Y and Hendershot LM: The unfolding tale
of the unfolded protein response. Cell. 107:827–830. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kaufman RJ, Scheuner D, Schröder M, Shen
X, Lee K, Liu CY and Arnold SM: The unfolded protein response in
nutrient sensing and differentiation. Nat Rev Mol Cell Biol.
3:411–421. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shi Y, Vattem KM, Sood R, An J, Liang J,
Stramm L and Wek RC: Identification and characterization of
pancreatic eukaryotic initiation factor 2 alpha-subunit kinase,
PEK, involved in translational control. Mol Cell Biol.
18:7499–7509. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Harding HP, Novoa I, Zhang Y, Zeng H, Wek
R, Schapira M and Ron D: Regulated translation initiation controls
stress-induced gene expression in mammalian cells. Mol Cell.
6:1099–1108. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Scheuner D, Song B, McEwen E, Liu C,
Laybutt R, Gillespie P, Saunders T, Bonner-Weir S and Kaufman RJ:
Translational control is required for the unfolded protein response
and in vivo glucose homeostasis. Mol Cell. 7:1165–1176. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Schindler AJ and Schekman R: In vitro
reconstitution of ER-stress induced ATF6 transport in COPII
vesicles. Proc Natl Acad Sci USA. 106:pp. 17775–17780. 2009;
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Harding HP, Zhang Y, Bertolotti A, Zeng H
and Ron D: Perk is essential for translational regulation and cell
survival during the unfolded protein response. Mol Cell. 5:897–904.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
McCullough KD, Martindale JL, Klotz LO, Aw
TY and Holbrook NJ: Gadd153 sensitizes cells to endoplasmic
reticulum stress by down-regulating Bcl2 and perturbing the
cellular redox state. Mol Cell Biol. 21:1249–1259. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yamaguchi H and Wang HG: CHOP is involved
in endoplasmic reticulum stress-induced apoptosis by enhancing DR5
expression in human carcinoma cells. J Biol Chem. 279:45495–45502.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Boyce M and Yuan J: Cellular response to
endoplasmic reticulum stress: A matter of life or death. Cell Death
Differ. 13:363–373. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ferri KF and Kroemer G: Organelle-specific
initiation of cell death pathways. Nat Cell Biol. 3:E255–E263.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Rutkowski DT and Kaufman RJ: Atripto the
ER: Coping with stress. Trends Cell Biol. 14:20–28. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mori K, Ma W, Gething MJ and Sambrook J: A
transmembrane protein with a cdc2+/CDC28-related kinase activity is
required for signaling from the ER to the nucleus. Cell.
74:743–756. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Morishima N, Nakanishi K, Takenouchi H,
Shibata T and Yasuhiko Y: An endoplasmic reticulum stress-specific
caspase cascade in apoptosis. Cytochrome c-independent activation
ofcaspase-9 by caspase-12. J Biol Chem. 277:34287–34294. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Vaupel P, Kallinowski F and Okunieff P:
Blood flow, oxygen and nutrient supply, and metabolic
microenvironment of human tumors: A review. Cancer Res.
49:6449–6465. 1989.PubMed/NCBI
|
|
24
|
Karnoub AE, Dash AB, Vo AP, Sullivan A,
Brooks MW, Bell GW, Richardson AL, Polyak K, Tubo R and Weinberg
RA: Mesenchymal stem cells within tumour stroma promote breast
cancer metastasis. Nature. 449:557–563. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Vermeulen L, De Sousa E, Melo F, van der
Heijden M, Cameron K, de Jong JH, Borovski T, Tuynman JB, Todaro M,
Merz C, Rodermond H, et al: Wnt activity defines colon cancer stem
cells and is regulated by the microenvironment. Nat Cell Biol.
12:468–476. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yauch RL, Gould SE, Scales SJ, Tang T,
Tian H, Ahn CP, Marshall D, Fu L, Januario T, Kallop D, et al: A
paracrine requirement for hedgehog signalling in cancer. Nature.
455:406–410. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Martinon F: Targeting endoplasmic
reticulum signaling pathways in cancer. Acta Oncol. 51:822–830.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Baird M, Woon Ang P, Clark I, Bishop D,
Oshima M, Cook MC, Hemmings C, Takeishi S, Worthley D, Boussioutas
A, et al: The unfolded protein response is activated in
Helicobacter-induced gastric carcinogenesis in a non-cell
autonomous manner. Lab Invest. 93:112–122. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Pike LR, Singleton DC, Buffa F, Abramczyk
O, Phadwal K, Li JL, Simon AK, Murray JT and Harris AL:
Transcriptional up-regulation of ULK1 by ATF4 contributes to cancer
cell survival. Biochem J. 449:389–400. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Nagelkerke A, Bussink J, Mujcic H, Wouters
BG, Lehmann S, Sweep FC and Span PN: Hypoxia stimulates migration
of breast cancer cells via the PERK/ATF4/LAMP3-arm of the unfolded
protein response. Breast Cancer Res. 15:R22013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Dubey R and Saini N: STAT6 silencing
up-regulates cholesterol synthesis via miR-197/FOXJ2 axis and
induces ER stress-mediated apoptosis in lung cancer cells. Biochim
Biophys Acta. 1849:32–43. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Rojas C, Pan-Castillo B, Valls C, Pujadas
G, Garcia-Vallve S, Arola L and Mulero M: Resveratrol enhances
palmitate-induced ER stress and apoptosis in cancer cells. PLoS
One. 9:e1139292014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Xu Y, Li D, Zeng L, Wang C, Zhang L, Wang
Y, Yu Y, Liu S and Li Z: Proteasome inhibitor lactacystin enhances
cisplatin cytotoxicity by increasing endoplasmic reticulum
stress-associated apoptosis in HeLa cells. Mol Med Rep. 11:189–195.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Little E, Ramakrishnan M, Roy B, Gazit G
and Lee AS: The glucose-regulated proteins (GRP78 and GRP94):
Functions, gene regulation, and applications. Crit Rev Eukaryot
Gene Expr. 4:1–18. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bertolotti A, Zhang Y, Hendershot LM,
Harding HP and Ron D: Dynamic interaction of BiP and ER stress
transducers in the unfolded-protein response. Nat Cell Biol.
2:326–332. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Pouysségur J, Shiu RP and Pastan I:
Induction of two transformation-sensitive membrane polypeptides in
normal fibroblasts by a block in glycoprotein synthesis or glucose
deprivation. Cell. 11:941–947. 1977. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Suzuki CK, Bonifacino JS, Lin AY, Davis MM
and Klausner RD: Regulating the retention of T-cell receptor alpha
chain variants within the endoplasmic reticulum: Ca(2+)-dependent
association with BiP. J Cell Biol. 114:189–205. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yoshida H, Haze K, Yanagi H, Yura T and
Mori K: Identification of the cis-acting endoplasmic reticulum
stress response element responsible for transcriptional induction
of mammalian glucose-regulated proteins. Involvement of basic
leucine zipper transcription factors. J Biol Chem. 273:33741–33749.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lee AS: Mammalian stress response:
Induction of the glucose-regulated protein family. Curr Opin Cell
Biol. 4:267–273. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kaufman RJ: Stress signaling from the
lumen of the endoplasmic reticulum: coordination of gene
transcriptional and translational controls. Genes Dev.
13:1211–1233. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Li X, Zhang K and Li Z: Unfolded protein
response in cancer: The physician's perspective. J Hematol Oncol.
4:82011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lai E, Teodoro T and Volchuk A:
Endoplasmic reticulum stress: Signaling the unfolded protein
response. Physiology (Bethesda). 22:193–201. 2007.PubMed/NCBI
|
|
43
|
Misra UK, Gonzalez-Gronow M, Gawdi G and
Pizzo SV: The role of MTJ-1 in cell surface translocation of GRP78,
a receptor for alpha 2-macroglobulin-dependent signaling. J
Immunol. 174:2092–2097. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Misra UK and Pizzo SV: Modulation of the
unfolded protein response in prostate cancer cells by
antibody-directed against the carboxyl-terminal domain of GRP78.
Apoptosis. 15:173–182. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Fu R, Yang P, Wu HL, Li ZW and Li ZY:
GRP78 secreted by colon cancer cells facilitates cell proliferation
via PI3K/Akt signaling. Asian Pac J Cancer Prev. 15:7245–7249.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Lu MC, Lai NS, Yin WY, Yu HC, Huang HB,
Tung CH, Huang KY and Yu CL: Anti-citrullinated protein antibodies
activated ERK1/2 and JNK mitogen-activated protein kinases via
binding to surface-expressed citrullinated GRP78 on mononuclear
cells. J Clin Immunol. 33:558–566. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Song MS, Park YK, Lee JH and Park K:
Induction of glucose-regulated protein 78 by chronic hypoxia in
human gastric tumor cells through a protein kinase
C-epsilon/ERK/AP-1 signaling cascade. Cancer Res. 61:8322–8330.
2001.PubMed/NCBI
|
|
48
|
Zhang LH and Zhang X: Roles of GRP78 in
physiology and cancer. J Cell Biochem. 110:1299–1305. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Mhaidat NM, Alzoubi KH, Almomani N and
Khabour OF: Expression of glucose regulated protein 78 (GRP78)
determines colorectal cancer response to chemotherapy. Cancer
Biomark. 15:197–203. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Guan M, Chen X, Ma Y, Tang L, Guan L, Ren
X, Yu B, Zhang W and Su B: MDA-9 and GRP78 as potential diagnostic
biomarkers for early detection of melanoma metastasis. Tumour Biol.
36:2973–2982. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Jiang X, Kanda T, Nakamoto S, Haga Y,
Sasaki R, Nakamura M, Wu S, Mikata R and Yokosuka O: Knockdown of
glucose-regulated protein 78 enhances poly (ADP-ribose) polymerase
cleavage in human pancreatic cancer cells exposed to endoplasmic
reticulum stress. Oncol Rep. 32:2343–1248. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Raiter A, Yerushalmi R and Hardy B:
Pharmacological induction of cell surface GRP78 contributes to
apoptosis in triple negative breast cancer cells. Oncotarget.
5:11452–11463. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Weinreb I, Goldstein D, Irish J and
Perez-Ordonez B: Expression patterns of Trk-A, Trk-B, GRP78, and
p75NRT in olfactory neuroblastoma. Hum Pathol. 40:1330–1335. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Tsunemi S, Nakanishi T, Fujita Y, Bouras
G, Miyamoto Y, Miyamoto A, Nomura E, Takubo T and Tanigawa N:
Proteomics-based identification of a tumor-associated antigen and
its corresponding autoantibody in gastric cancer. Oncol Rep.
23:949–956. 2010.PubMed/NCBI
|
|
55
|
Yang L, Yang S, Liu J, Wang X, Ji J, Cao
Y, Lu K, Wang J and Gao Y: Expression of GRP78 predicts
taxane-based therapeutic resistance and recurrence of human gastric
cancer. Exp Mol Pathol. 96:235–241. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Yang L, Yang SY, Ji JM, Cao YF, Ji CF, Ji
JF, Xu WW and Wang JH: GRP78 expression in gastric cancer and its
clinical significance. Zhonghua Zhong Liu Za Zhi. 35:837–842.
2013.(In Chinese). PubMed/NCBI
|
|
57
|
Wu JY, Cheng CC, Wang JY, Wu DC, Hsieh JS,
Lee SC and Wang WM: Discovery of tumor markers for gastric cancer
by proteomics. PLoS One. 9:e841582014. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Zheng HC, Takahashi H, Li XH, Hara T,
Masuda S, Guan YF and Takano Y: Overexpression of GRP78 and GRP94
are markers for aggressive behavior and poor prognosis in gastric
carcinomas. Hum Pathol. 39:1042–1049. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Zhang LJ, Chen S, Wu P, Hu CS, Thorne RF,
Luo CM, Hersey P and Zhang XD: Inhibition of MEK blocks GRP78
up-regulation and enhances apoptosis induced by ER stress in
gastric cancer cells. Cancer Lett. 274:40–46. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Xu YY, You YW, Ren XH, Ding Y, Cao J, Zang
WD, Feng R and Zhang QX: Endoplasmic reticulum stress-mediated
signaling pathway of gastric cancer apoptosis.
Hepatogastroenterology. 59:2377–2384. 2012.PubMed/NCBI
|
|
61
|
Huang X, Zhang Z, Jia L, Zhao Y, Zhang X
and Wu K: Endoplasmic reticulum stress contributes to vitamin E
succinate-induced apoptosis in human gastric cancer SGC-7901 cells.
Cancer Lett. 296:123–131. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Huang X, Li L, Zhang L, Zhang Z, Wang X,
Zhang X, Hou L and Wu K: Crosstalk between endoplasmic reticulum
stress and oxidative stress in apoptosis induced by α-tocopheryl
succinate in human gastric carcinoma cells. Br J Nutr. 109:727–735.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Rauschert N, Brändlein S, Holzinger E,
Hensel F, Müller-Hermelink HK and Vollmers HP: A new tumor-specific
variant of GRP78 as target for antibody-based therapy. Lab Invest.
88:375–386. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Winder T, Bohanes P, Zhang W, Yang D,
Power DG, Ning Y, Gerger A, Wilson PM, Tang LH, Shah M, et al:
GRP78 promoter polymorphism rs391957 as potential predictor for
clinical outcome in gastric and colorectal cancer patients. Ann
Oncol. 22:2431–2439. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Zhang J, Jiang Y, Jia Z, Li Q, Gong W,
Wang L, Wei D, Yao J, Fang S and Xie K: Association of elevated
GRP78 expression with increased lymph node metastasis and poor
prognosis in patients with gastric cancer. Clin Exp Metastasis.
23:401–410. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Zhang D and Fan D: Multidrug resistance in
gastric cancer: Recent research advances and ongoing therapeutic
challenges. Expert Rev Anticancer Ther. 7:1369–1378. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Tsutsumi S, Namba T, Tanaka KI, Arai Y,
Ishihara T, Aburaya M, Mima S, Hoshino T and Mizushima T: Celecoxib
upregulates endoplasmic reticulum chaperones that inhibit
celecoxib-induced apoptosis in human gastric cells. Oncogene.
25:1018–1029. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Leja M, You W, Camargo MC and Saito H:
Implementation of gastric cancer screening-the global experience.
Best Pract Res Clin Gastroenterol. 28:1093–1106. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Smyth EC and Cunningham D: Targeted
therapy for gastric cancer. Curr Treat Options Oncol. 13:377–389.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Cheng CC, Huang CF, Ho AS, Peng CL, Chang
CC, Mai FD, Chen LY, Luo TY and Chang J: Novel targeted nuclear
imaging agent for gastric cancer diagnosis: Glucose-regulated
protein 78 binding peptide-guided 111In-labeled polymeric micelles.
Int J Nanomedicine. 8:1385–1391. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Kang J, Zhao G, Lin T, Tang S, Xu G, Hu S,
Bi Q, Guo C, Sun L, Han S, et al: A peptide derived from phage
display library exhibits anti-tumor activity by targeting GRP78 in
gastric cancer multidrug resistance cells. Cancer Lett.
339:247–259. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Park HR, Tomida A, Sato S, Tsukumo Y, Yun
J, Yamori T, Hayakawa Y, Tsuruo T and Shin-ya K: Effect on tumor
cells of blocking survival response to glucose deprivation. J Natl
Cancer Inst. 96:1300–1310. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Cheng CC, Lu N, Peng CL, Chang CC, Mai FD,
Chen LY, Liao MH, Wang WM and Chang J: Targeting to overexpressed
glucose-regulated protein 78 in gastric cancer discovered by 2D
DIGE improves the diagnostic and therapeutic efficacy of
micelles-mediated system. Proteomics. 12:2584–2597. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Culver KW, Ram Z, Wallbridge S, Ishii H,
Oldfield EH and Blaese RM: In vivo gene transfer with retroviral
vector-producer cells for treatment of experimental brain tumors.
Science. 256:1550–1552. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Dong D, Dubeau L, Bading J, Nguyen K, Luna
M, Yu H, Gazit-Bornstein G, Gordon EM, Gomer C, Hall FL, et al:
Spontaneous and controllable activation of suicide gene expression
driven by the stress-inducible grp78 promote r resulting in
eradication of sizable human tumors. Hum Gene Ther. 15:553–561.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Lee AS: GRP78 induction in cancer:
Therapeutic and prognostic implications. Cancer Res. 67:3496–3499.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Azatian A, Yu H, Dai W, Schneiders FI,
Botelho NK and Lord RV: Effectiveness of HSV-tk suicide gene
therapy driven by the Grp78 stress-inducible promoter in
esophagogastric junction and gastric adenocarcinomas. J
Gastrointest Surg. 13:1044–1051. 2009. View Article : Google Scholar : PubMed/NCBI
|














