Introduction
In recent years, colorectal cancer remains a primary
cause of morbidity and it is the fourth leading cause of
cancer-associated mortality worldwide (1). In the USA (2) and Europe (3), colorectal cancer is the second leading
cause of cancer-associated mortality. In China, it has been noted
that the morbidity and mortality rates of colorectal cancer were
increased compared with previous years (4). In Taiwan, colon cancer is the fourth
most common type of cancer, accounting for 23.9 mortalities per
100,000 individuals, based on the 2014 report from the Department
of Health, Executive Yuan, Taiwan (5). For patients with superficial cancer
(Duke's staging of colorectal cancer), the 5-year survival rate was
up to 90%; however, for patients with distant metastasis, the
survival rate was ~9% (6,7). Therefore, it is well known that
investigating the mechanisms underlying metastatic disease is
critical for the treatment and development of metastatic prevention
strategies of patients with cancer.
It is well known that tumor metastasis involves
epithelial cancer cell adhesion, migration, invasion and
angiogenesis for the development of cancer in other sites of the
body (8,9). Furthermore, numerous factors are
associated with tumor metastasis, including matrix
metalloproteinases (MMPs) and urokinase plasminogen activator
(uPA), which serve critical roles in degrading the extracellular
matrix and basement membrane collagen for cancer cells to invade
into new sites (10–12). Epithelial mesenchymal-transition (EMT)
is an important process for epithelial cancer cell loss of polarity
and cell to cell contact (13), and
EMT is one of the initial and primary events in tumor progression
(14). The fibroblast growth factor
family has been revealed to be associated with tumor metastasis in
EMT (15,16). Other factors, including secreted
factors, cytokines, chemokines and growth factors have been
revealed to be associated with the distinct modes of metastasis and
subsequent mortality in tumors (17).
A previous study demonstrated that activation of the
phosphatidylinositol 3 kinase (PI3K)/protein kinase B (Akt)
signaling pathway is involved in cancer cell metastasis (18). Therefore, numerous studies have aimed
to investigate the use of novel compounds extracted from natural
products as treatments for colon cancer cell metastasis (19–21).
Tetrandrine (TET), a bisbenzylisoquinoline alkaloid
isolated from the root of Stephania tetrandra S. Moore, has
been revealed to have biological activity, including cytotoxic
effects, cell cycle arrest and induction of cell apoptosis in a
number of human cancer cell lines (22–26). It
was reported that TET suppresses proliferation, induces apoptosis
and inhibits migration and invasion in human prostate cancer cells
(27). It was also reported that TET
regulates metastatic- and angiogenic-associated proteins, including
vascular endothelial growth factor, hypoxia-inducible factor-1,
integrin β5, endothelial cell specific molecule-1 and intercellular
adhesion molecule-1 (28).
Previously, it was demonstrated that TET targets epidermal growth
factor receptor signaling and its downstream molecules contribute
to the inhibition of epidermal growth factor (EGF)-induced HT29
cell metastasis in vitro (29). Furthermore, it was also reported that
TET-loaded PVP-b-PCL nanoparticles more efficiently inhibit cell
migration and invasion compared with free TET in A549 human lung
cancer cells (30). Although it was
reported that TET inhibits cell migration and invasion in human
colon cancer HT29 cells via inhibition of EGF, whether nuclear
factor (NF)-κB is involved in TET suppression of SW620 human colon
cancer cell metastasis remains unclear. The present study revealed
that TET inhibited cell migration and invasion of SW620 cells via
the PI3K, NF-κB and mitogen-activated protein kinase signaling
pathways.
Materials and methods
Chemicals and reagents
TET, dimethyl sulfoxide (DMSO) and propidium iodide
were obtained from Sigma-Aldrich (Merck KGaA, Darmstadt, Germany).
Leibovitz's L-15 medium, fetal bovine serum (FBS), L-glutamine and
antibiotics (penicillin-streptomycin) were purchased from Gibco
(Thermo Fisher Scientific, Inc., Waltham, MA, USA). Primary and
secondary antibodies were obtained from Cell Signaling Technology,
Inc. (Danvers, MA, USA). Polyvinylidene difluoride (PVDF) membrane
was obtained from EMD Millipore (Billerica, CA, USA).
Cell culture
The SW620 human colon cancer cell line was purchased
from the Food Industry Research and Development Institute (Hsinchu,
Taiwan). Cells were cultured in Leibovitz's L-15 medium
supplemented with 10% FBS, 100 units/ml penicillin and 100 µg/ml
streptomycin in a 75 cm2 tissue culture flask at 37°C in
a humidified atmosphere containing 5% CO2 (31,32).
Cell viability assays
SW620 cells were seeded in a 96-well plate at a
density of 1.5×104 cells/well and treated with TET at
the final concentrations of 0, 0.2, 0.39, 0.78, 1.56, 3.12, 6.25,
12.5, 25 and 50 µM or 0.5% DMSO as the vehicle control. Following
exposure to the drug for 24 or 48 h, 100 µl MTT (0.5 mg/ml;
Sigma-Aldrich; Merck KGaA) was added to each well and the plates
were incubated for an additional 4 h at 37°C. MTT solution in the
medium was aspirated off. To achieve solubilization of the formazan
crystals formed in viable cells, 200 µl DMSO was added to each well
prior to evaluation of absorbance at a wavelength of 570 nm
(33).
Adhesion assay
SW620 cells (1×106 cells/well) were
cultured with 0, 1, 5 and 10 µM TET for 48 h at 37°C in 12-well
plates, which were pre-coated with type I collagen (10 µg/ml)
(Merck KGaA, Darmastadt, Germany) for 60 min at room temperature.
Unattached cells were removed and attached cells were mixed in 1%
glutaraldehyde (Sigma-Aldrich; Merck KGaA) supplemented with PBS
for 20 min, and stained with 0.02% crystal violet solution for 5
min at room temperature. Ethanol (70%) was used to dissolve crystal
violet in the stained cells. Optical density (O.D.) was evaluated
at 570 nm using a microplate reader with a reference of 405 nm. The
adhesion ability (percentage of adhesive cells, %) was determined
by measuring the treated cells compared with the control cells
(34).
Adhesion ability(%)=O.D.TET
treatmentO.D.Control×100%
Wound healing assay
SW620 cells (5×105 cells/well) were
cultured in 6-well plate until cell growth reached 100% confluence.
A sterile yellow micropipette tip was used to scrape the cell
monolayers in the well and cells were washed with PBS three times.
Cells were then cultured in medium containing 0, 1, 5 and 10 µM TET
for 24 and 48 h at 37°C. Cells were examined and imaged using an
inverted microscope (×100 magnification) (32,34).
Invasion and migration assays
Evaluation of SW620 cell invasion was performed
using Matrigel-coated Transwell cell culture chambers (8 µm pore
size). Cells (8×104 cells/well) were seed in the upper
chamber and incubated with Leibovitz's L-15 medium supplemented
with 0% FBS, and 0 or 10 µM TET for 48 h at 37°C. Leibovitz's L-15
medium supplemented with 10% FBS was placed in the lower chamber.
The non-invaded cells were removed using a cotton swab on the upper
surface of the membrane and the invaded cells on the lower surface
of the membrane were fixed with 4% cold formaldehyde, stained with
0.1% crystal violet for 15 min at room temperature and then imaged
using an inverted light microscope (×200 magnification). The
invaded cells in the chamber were counted. For the determination of
cell migration, the same invasion assay was performed with the
membrane coated without Matrigel, as previously described (34). Cell migration was quantified by ImageJ
(version 1.49o software, National Institutes of Health, Bethesda,
MD, USA) based on the change in the area of the cell-free gap
before and after TET stimulation:
24h Inhibitory ability of Migration(%of
control)=(wouldareaTET24h/wouldareaTET0hwouldareaControl24h/wouldareaControl0h)×100%48h
Inhibitory ability of Migration(%of
control)=(wouldareaTET48h/wouldareaTET0hwouldareaControl48h/wouldareaControl0h)×100%
Western blot analysis
SW620 cells (6×106) were plated in 10-cm
dishes and incubated with 0, 1, 5, 10, 20 and 30 µM TET for 48 h at
37°C, subsequently the cells were collected and lysed in a lysis
buffer [40 mM Tris-HCl (pH 7.4), 10 mM EDTA, 120 mM NaCl, 1 mM
dithiothreitol, 0.1% Nonide P-40]. The total protein concentration
from each treatment was evaluated as previously described (34). A total of 30 µg protein was separated
by SDS-PAGE (5% stacking gel and 10–12% separation gel) for western
blot analysis. The gel was transferred to a PVDF membrane and the
membrane was blocked in 5% fat-free dry milk solution in PBS
containing 0.1% Tween-20 for 1 h at room temperature, and then
incubated with primary antibodies overnight at 4°C. The phospho-Jun
N-terminal kinase (p-JNK) 1/2 (sc-6254), p-38 (sc-136210),
phospho-p-38 (sc-166182), ras homolog family member A (Rho A;
sc-418), growth factor receptor bound protein 2 (GRB2; sc-503) and
14-3-3 protein σ (sc-100638) antibodies were supplied by Santa-Cruz
Biotechnology, Inc. (Dallas, TX, USA, dilution 1:1,000). The
anti-matrix metalloproteinase (MMP)-1 (MAB13439) and tissue
inhibitor of metalloproteinase (TIMP)-1 (AB6007) antibodies were
supplies by Merck Millipore Corp. (Billerica, MA, USA; dilution,
1:1,000). The Son of sevenless homolog (SOS)-1 (610095, dilution,
1:250), phosphoinositide 3-kinase (PI3K) (610046, dilution,
1:2,500), signal transducer and activator of transcription 1
(STAT1) (610115, dilution, 1:1,000), cyclooxygenase-2 (Cox-2)
(610204, dilution, 1:500) and -nuclear factor kappa B (NF-κB p65)
(610868, dilution, 1:500) antibodies were obtained from BD
Biosciences (Bedford, MA, USA). The anti-MMP-2 (ab7032, dilution,
1:1,000) antibody was obtained from Abcam (Cambridge, MA, USA), and
the MMP-9 (GTX32122, dilution, 1:1,000) antibody was supplied by
GeneTex, Inc. (Irvine, CA, USA) for the β-Catenin (C2206, dilution,
1:4,000) and β-actin (A5316, dilution, 1:10,000) antibodies were
supplied by Sigma-Aldrich (St. Louis, MO, USA). Subsequently, the
membranes were incubated with secondary antibodies [horseradish
peroxidase (HRP)-conjugated mouse immunoglobulin G (IgG; GTX213112)
and rabbit HRP-conjugated IgG secondary antibodies (GTX213110),
dilution, 1:5,000; GeneTex, Irvine, CA, USA] for 1 h at room
temperature. Proteins were visualized using enhanced
chemiluminescencereagents (GE Healthcare, Chicago, IL, USA) to
stain, as previously described (34).
Statistical analysis
All data are expressed as the mean ± standard
deviation. Differences between groups were analyzed by one-way
analysis of variance. Statistical comparisons were made using
Tukey's test (SigmaPlot for Windows v12.0; Systat Software, Inc.,
San Jose, CA), and P<0.05 was considered to indicate a
statistically significant difference. Differences between two
groups were determined using the unpaired Student's t-test
(SigmaPlot for Windows version 10.0; Systat Software, Inc., San
Jose, CA), and P<0.01 was considered to indicate a statistically
significant difference.
Results
TET decreases the cell viability of
SW620 cells
SW620 cells were treated with TET (0, 0.2, 0.39,
0.78, 1.56, 3.12, 6.25, 12.5, 25 and 50 µM) for 24 and 48 h prior
to collection of the cells to determine the percentage of total
viable cell number (Fig. 1). The data
indicated a significant dose-dependent reduction of living SW620
cells treated with TET at 0.2–50 µM concentrations for 24 and 48 h
(P<0.001). Thus the present study selected 0, 1, 5 and 10 µM for
cell migration and invasion experiments.
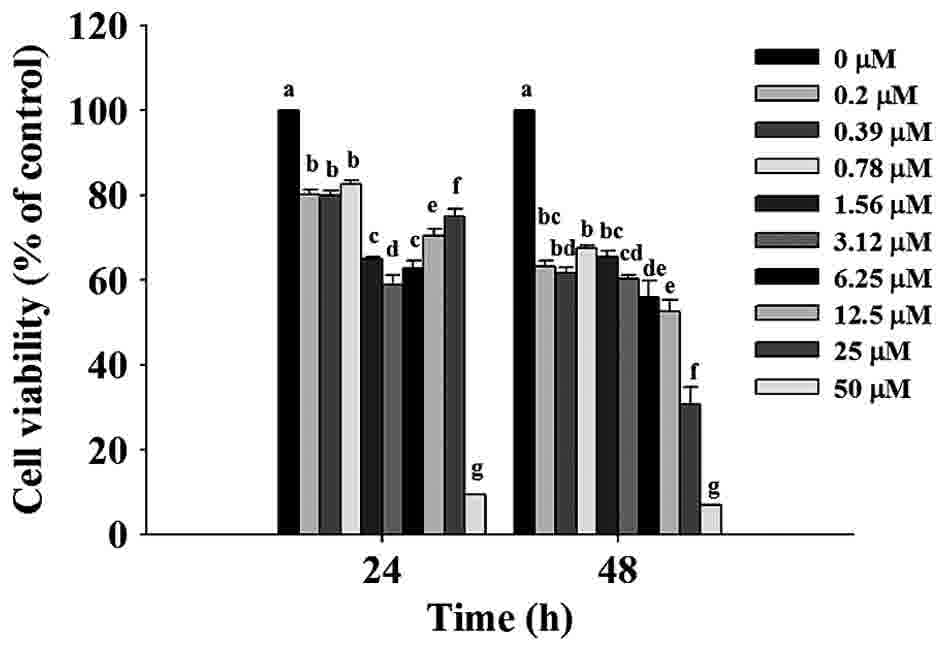 | Figure 1.TET decreases the percentage of
viable SW620 cells. Cells (1.5×104 cells/well) were
treated with 0, 0.2, 0.39, 0.78, 1.56, 3.12, 6.25, 12.5, 25 and 50
µM TET or 0.5% dimthylsulfoxide as a vehicle control for 24 and 48
h. Cell growth inhibition was assessed by MTT assay. The values
with different letters were significantly different from each
other, P<0.05 (Tukey's test). TET, tetrandrine. |
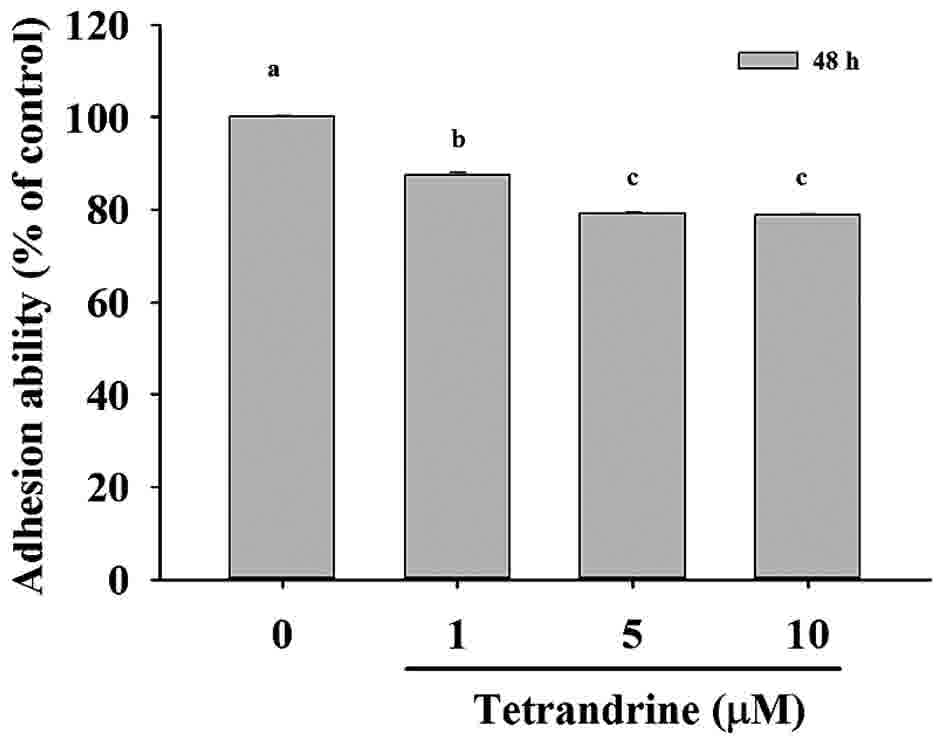
TET decreases the cell adhesion of
SW620 cells
SW620 cells were cultured with 0, 1, 5 and 10 µM TET
for 48 h and the total percentage of adhesion was determined and
presented in Fig. 2, [1 µM
(87.36±0.71%, P<0.05); 5 µM (79.22±0.18%, P<0.05); 10 µM
(78.72±0.18%, P<0.05) compared to untreated control cells
(100.00±0.18%)]. Based on these results, it was indicated that TET
at 1–10 µM for 48 h treatment significantly reduced cell adhesion
in SW620 cells in vitro.
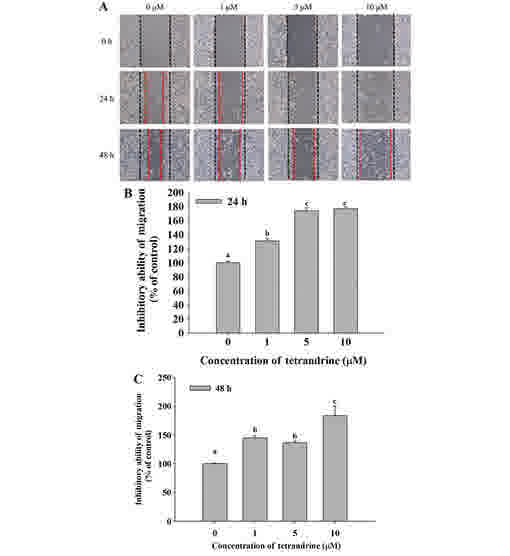
TET decreases cell mobility of SW620
cells
Cell mobility was evaluated using a wound healing
assay. SW620 cells were cultured in 6-well plates and the cell
monolayers were scraped and then cultured in medium containing 0,
1, 5 and 10 µM TET for 24 and 48 h (Fig.
3). Fig. 3A demonstrated that
closure of the scraped area at the highest dose of TET was
decreased compared with the control. TET significantly reduced cell
mobility, and increased the ability to inhibit migration at 24 and
48 h up to 176.74 and 183.45% in the 10 µM TET treated cells,
respectively, compared with control cells (Fig. 3B and C).
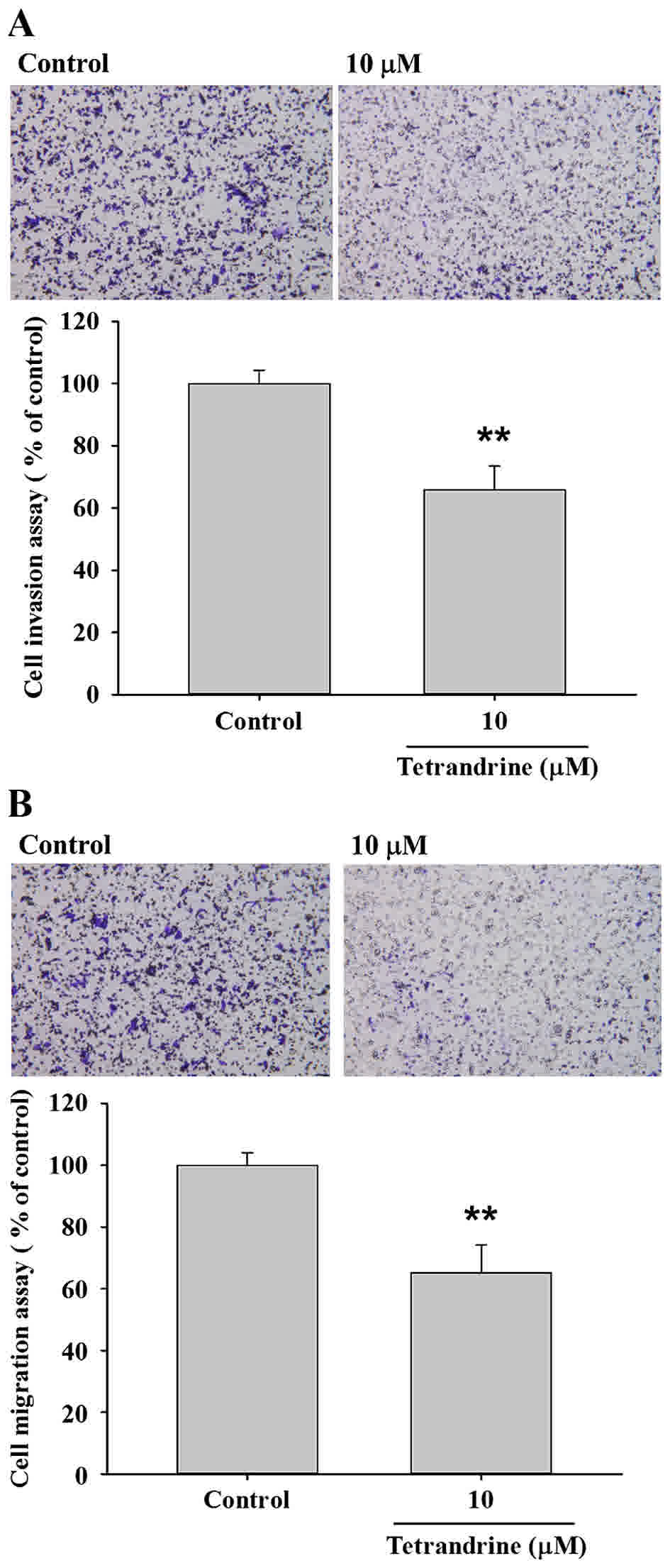
TET inhibits the migration and
invasion of SW620 cells
Transwell migration and invasion assays were
performed to investigate the inhibitory role of TET on SW620 cell
migration and invasion, the results are presented in Fig. 4. The results indicated that TET
significantly (P<0.05) inhibited cell invasion by 35% for 10 µM
TET treated cells for 48 h (P<0.01; Fig. 4A), and inhibited cell migration by 35%
for 10 µM TET treated cells for 48 h compared with the control
cells (P<0.01; Fig. 4B).
TET alters expression levels of
proteins associated with migration and invasion of SW620 cells
The present study further investigated the role of
upstream regulated proteins associated with SW620 cell migration
and invasion following exposure to TET (Fig. 5). TET significantly reduced protein
expression levels of MMP-9, MMP-2, MMP-1, SOS Ras/Rac guanine
nucleotide exchange factor 1 (SOS-1), PI3K, phosphorylated (p)-c
Jun N-terminal kinase (JNK)1/2, growth factor receptor bound
protein 2 (GRB2) and TIMP metallopeptidase inhibitor 1 (TIMP1;
Fig. 5A), p-p38, p38, 14-3-3, Rho A,
signal transducer and activator of transcription-1 (STAT-1) and
cyclooxygenase-2 (Cox-2; Fig. 5B),
β-catenin and NF-κB (Fig. 5C). The
protein expression levels were decreased in TET-treated cells
compared with untreated-cells. TET inhibited the p38, JNK and Rho A
signaling pathways by reducing PI3K, Cox-2 and NF-κB p65 expression
levels, which induced MMP-2/-9 downregulation (Fig. 6).
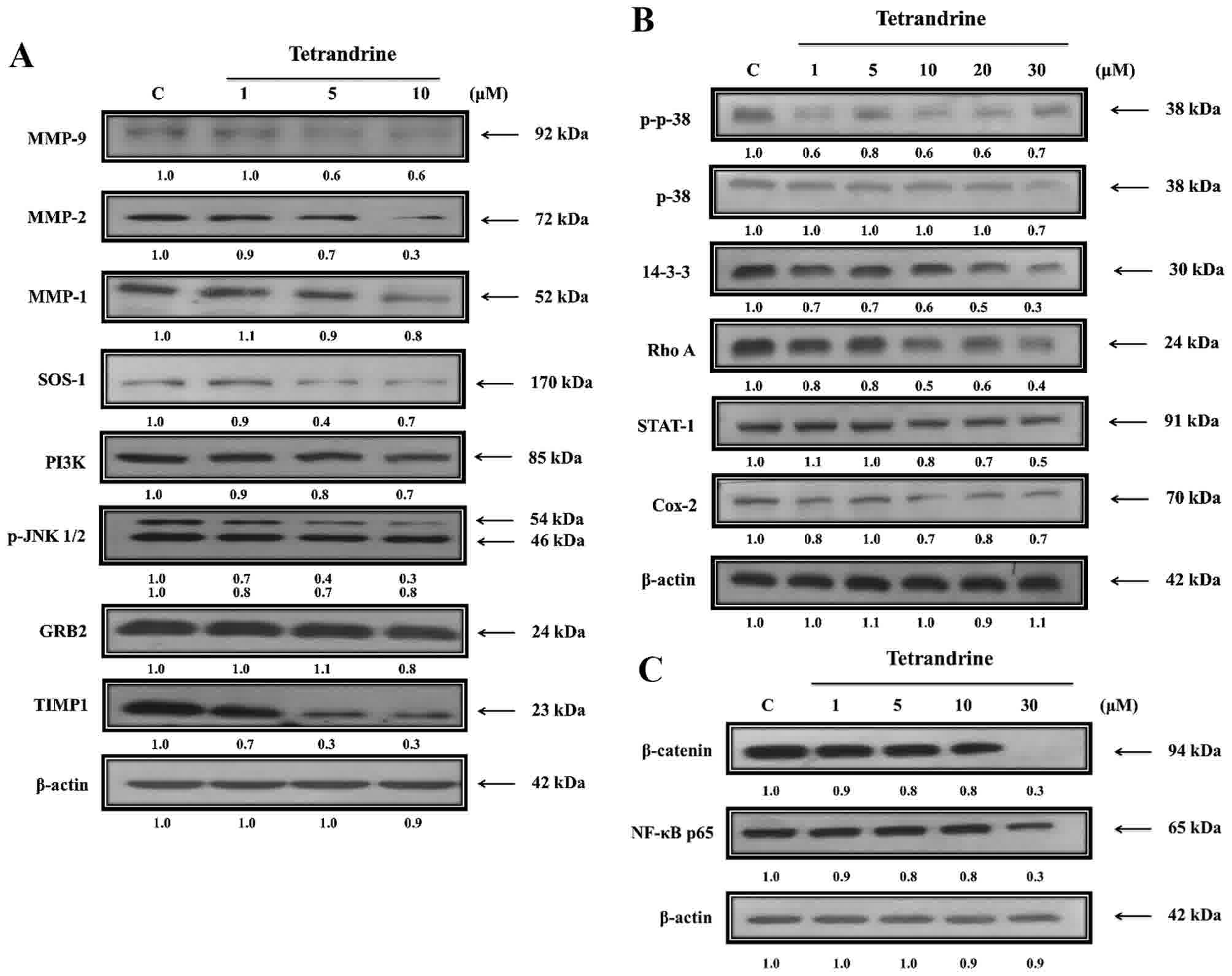 | Figure 5.TET alters the expression levels of
proteins associated with migration and invasion of SW620 cells.
Cells were treated with various concentrations of TET for 48 h and
then total proteins were quantified and apoptosis associated
proteins were examined by western blotting. (A) MMP-9, MMP-2 and
MMP-1, SOS-1, PI3K, p-JNK1/2, GRB2, TIMP1. (B) p-p38, p38, 14-3-3,
Rho A, STAT-1 and Cox-2. (C) β-catenin and NF-κB p65. TET,
tetrandrine. MMP, matrix metalloproteinase; SOS-1, SOS Ras/Rac
guanine nucleotide exchange factor 1; PI3K, phosphatidylinositol 3
kinase; p, phosphorylated; JNK1/2, c Jun N-terminal kinase; GRB2,
growth factor receptor bound protein 2; TIMP1, TIMP
metallopeptidase inhibitor 1; STAT-1, signal transducer and
activator of transcription-1; Cox-2, cyclooxygenase-2; NF-κB,
nuclear factor-κB. |
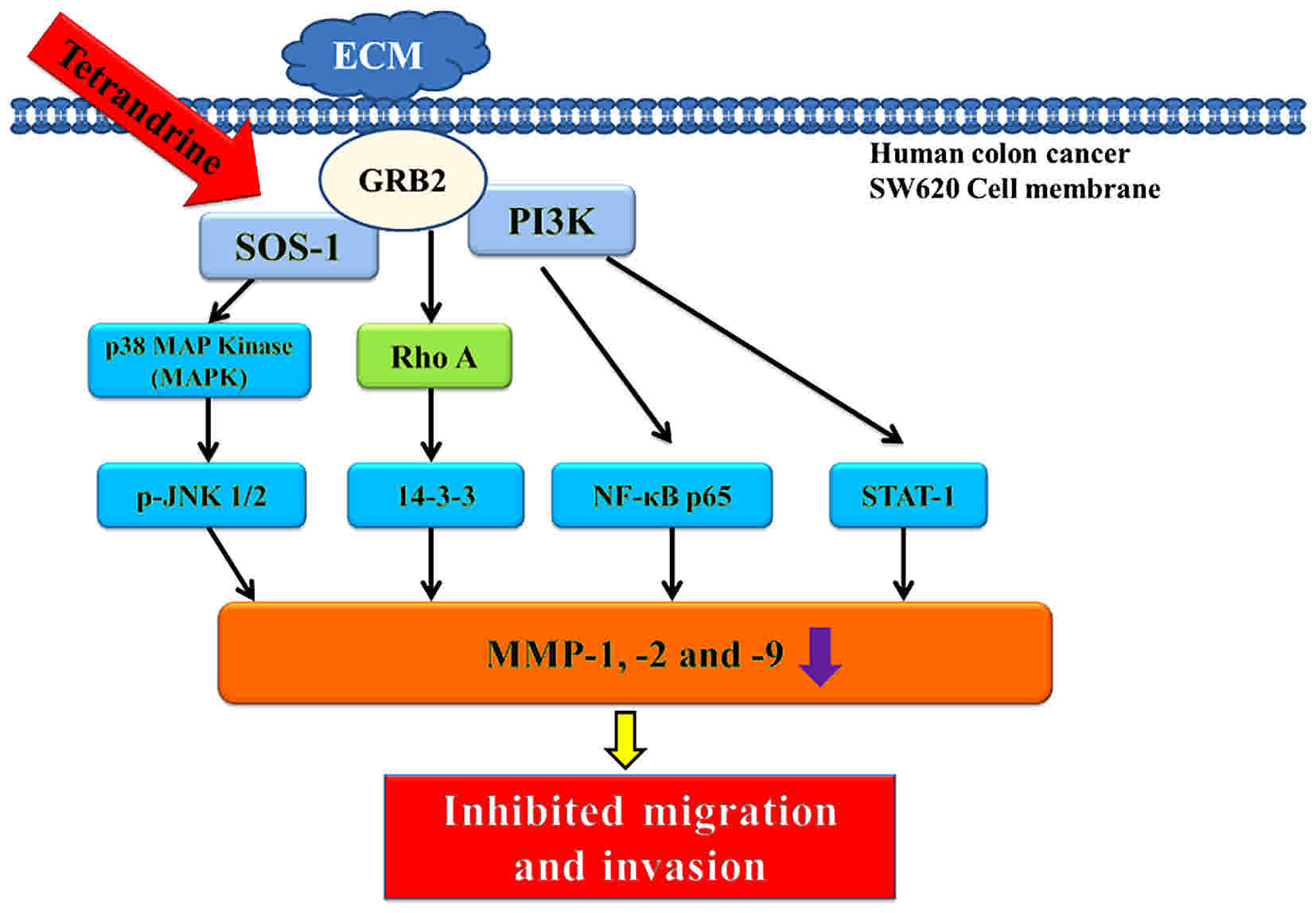 | Figure 6.The possible signaling pathways for
TET inhibited cell mobility, adhesion, migration and invasion in
SW620 cells in vitro. TET, tetrandrine; ECM, extracellular
matrix; GRB2, growth factor receptor bound protein 2; SOS-1, SOS
Ras/Rac guanine nucleotide exchange factor 1; PI3K,
phosphatidylinositol 3 kinase; p, phosphorylated; JNK1/2, c Jun
N-terminal kinase; MAP, mitogen-activated protein; NF-κB, nuclear
factor-κB; STAT-1, signal transducer and activator of
transcription-1; MMP, matrix metalloproteinase. |
Discussion
Previous studies have demonstrated that cancer cells
exhibit extensive invasive and migratory abilities, which are
factors that may block the effectiveness of clinical treatments
against cancer, including chemotherapy (35,36).
Cancer cell metastasis involves a complex multistep process, which
includes cell movement and cell adhesion accompanied with
migration, invasion and angiogenesis to develop new tumors in other
sites of body (37,38). Therefore, investigators focus on the
inhibition of cancer cell migration and invasion, as an anticancer
strategy. It has previously been reported that TET induces cancer
cell death via cell cycle arrest and induction of apoptosis in
numerous human cancer cell lines; however, there is no available
information to demonstrate TET inhibiting migration and invasion in
human colon cancer SW620 cells. The present study investigated the
effects of TET on adhesion, migration and invasion of SW620 cells
in vitro.
Firstly, the present study examined the cytotoxic
effects of TET on SW620 cells in vitro and the results
indicated that TET induced cell death in a dose-dependent manner.
Therefore, 1, 5 and 10 µM TET treatments were selected for further
experiments. The present study also investigated cell adhesion of
SW620 cells following exposure to 0, 1, 5 and 10 µM TET for 48 h
and the results indicated that TET inhibited cell adhesion in a
concentration-dependent manner. It is well documented that wound
healing is one of the methods for examining cancer cell mobility
(39,40); thus, the results from the wound
healing assay indicated that TET inhibited cell mobility in SW620
cells in a dose-dependent manner. The Transwell assay has been
recognized to be effective in the analysis of cell migration and
invasion (41,42). The present study performed Transwell
assays to investigate cell migration and invasion of SW620 cells
following exposure to TET in vitro. The findings indicated
that TET significantly inhibited cell migration and invasion when
compared with the control groups. Based on these observations, the
present study suggested that TET suppressed cell migration and
invasion via the inhibition of cell attachment (adhesion) to the
basement membrane.
MMPs, a family of zinc-dependent proteases, serve
essential roles in defining how cells interact with their
surrounding microenvironment (43).
It was reported that increased expression levels of MMPs are
associated with increased levels of cancer cell angiogenesis,
migration and invasion (44); thus,
MMPs have previously been used as drug targets (45). Therefore, the present study first
examined the protein expression levels of MMP-2 and MMP-9 in SW620
cells following exposure to various concentrations of TET, and the
results indicated that TET decreased the protein expression levels
of MMP-2, MMP-9, MMP-1 and TIMP1 in a concentration-dependent
manner, which was revealed by western blotting. MMP-2 and MMP-9
serve important roles in cancer invasion and metastasis (46,47).
Furthermore, results indicated that TET suppressed the protein
expression levels of SOS-1, PI3K, GRB2 and p-JNK1/2 in SW620 cells.
SOS-1 and GRB2 have been observed in HT 29 colon cancer cells
(48). To the best of the author's
knowledge, the present study is the first demonstrate that TET
inhibited the protein expression levels of SOS-1 and GRB2.
GRB2-associated binding protein 2 serves a critical role in the
proliferation and migration of various types of cancer (49). Therefore, further investigations are
required to understand the role of SOS-1 and GRB2 in cancer cell
metastasis. The results of the present study also revealed that TET
inhibited the protein expression levels of PI3K in SW620 cells.
PI3K/Akt and extracellular signal regulated kinase pathways are
involved in growth factor-mediated colon cancer proliferation
(50). It was reported that
17β-estradiol treatment inhibited prostaglandin E2-induced uPA,
MMP-9 and cellular motility by suppressing activation of JNK1/2 in
LoVo human colon cancer cells (51).
The results of the present study demonstrated that
TET inhibited the protein expression levels of p-p38, p38, 14-3-3
and Rho A in SW620 cells. p-p38 and p38 were significantly reduced
in TET-treated SW620 cells compared with untreated cells. It was
previously reported that in SW620 human colon cancer-derived
metastatic cells, nicotine stimulates the invasion and metastasis
of colon cancer cells in vitro via activation of the p38
MAPK downstream signaling pathway (52). The present study revealed that TET
significantly reduced the protein expression levels of 14-3-3 in
SW620 cells in a dose-dependent manner. It was previously
demonstrated that 14-3-3 protein overexpression promotes lung
cancer progression when combined with HSP27 overexpression
(53). A previous study revealed that
in patient colorectal cancer samples, Rho A is associated with the
invasion of lymph nodes and blood vessels, thus, Rho A may be a
promising target for cancer treatment (54).
The results of the present study additionally
indicated that TET significantly suppressed the protein expression
levels of β-catenin and NF-κB p65 in SW620 cells. β-catenin is a
92-kDa cellular protein and a member of the Wnt signaling pathway
that has been revealed to serve an important role in colorectal
cancer tumorigenesis (55,56), and is associated with E-cadherin in
maintaining cellular adhesion (57).
The aberrant activation of β-catenin increases its translocation to
the nucleus in colorectal cancer (58). Therefore, targeting the Wnt/β-catenin
signaling pathway to develop novel chemotherapeutic agents against
colon cancer may be a promising strategy. NF-κB is a transcription
factor closely associated with cell survival, proliferation and
metastasis (59). It is well
documented that agents blocking the NF-κB signaling pathway may act
as therapeutic agents to treat inflammation and cancer (60). The results of the present study
indicated that TET inhibited cell migration and invasion of SW620
cells via inhibition of NF-κB. It was also revealed that TET
suppressed the protein expression levels of STAT1 and Cox-2 in
SW620 cells. Constitutive overexpression of STAT1 in tumor cells is
correlated with protection of tumor cells to genotoxic stress
following doxorubicin (61) or
cisplatin (62) treatment. Cox-2 has
tumor promoting properties and is expressed in approximately 40–50%
of colonic adenomas and in 80–90% of colorectal carcinomas
(63,64). Cox-2 is also associated with cancer
cell invasion (65), serves an
important role in carcinogenesis and therefore has the potential to
be used as a novel anticancer therapeutic target (66,67).
In conclusion, the present study revealed that TET
suppressed cell mobility, adhesion, invasion and migration in SW620
cells via the inhibition of metastasis-associated proteins such as
MMP-2/-9.
Therefore, the results of the present study
suggested that TET may be a potential candidate for developing
preventive agents against human colon cancer metastasis.
Acknowledgements
The present study was supported by the China Medical
University Beigang Hospital, Yunlin, Taiwan (grant no. CMUBH
R103-011).
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel R, Desantis C and Jemal A:
Colorectal cancer statistics, 2014. CA Cancer J Clin. 64:104–117.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Labianca R, Beretta GD, Kildani B, Milesi
L, Merlin F, Mosconi S, Pessi MA, Prochilo T, Quadri A, Gatta G, et
al: Colon cancer. Crit Rev Oncol Hematol. 74:106–133. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ferlay J, Parkin DM and Steliarova-Foucher
E: Estimates of cancer incidence and mortality in Europe in 2008.
Eur J Cancer. 46:765–781. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Li M and Gu J: Changing patterns of
colorectal cancer in China over a period of 20 years. World J
Gastroenterol. 11:4685–4688. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ministry of Health and Welfare: The cancer
mortality report of the Department of Health. Taiwan: 2014
|
|
6
|
Camp ER and Ellis LM: CCR 20th Anniversary
Commentary: RAS as a Biomarker for EGFR-targeted therapy for
colorectal cancer-from concept to practice. Clin Cancer Res.
21:3578–3580. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hazan RB, Qiao R, Keren R, Badano I and
Suyama K: Cadherin switch in tumor progression. Ann N Y Acad Sci.
1014:155–163. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Makrilia N, Kollias A, Manolopoulos L and
Syrigos K: Cell adhesion molecules: Role and clinical significance
in cancer. Cancer Invest. 27:1023–1037. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Babykutty S, Suboj P, Srinivas P, Nair AS,
Chandramohan K and Gopala S: Insidious role of nitric oxide in
migration/invasion of colon cancer cells by upregulating MMP-2/9
via activation of cGMP-PKG-ERK signaling pathways. Clin Exp
Metastasis. 29:471–492. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Dung TD, Feng CC, Kuo WW, Pai P, Chung LC,
Chang SH, Hsu HH, Tsai FJ, Lin YM and Huang CY: Suppression of
plasminogen activators and the MMP-2/-9 pathway by a Zanthoxylum
avicennae extract to inhibit the HA22T human hepatocellular
carcinoma cell migration and invasion effects in vitro and in vivo
via phosphatase 2A activation. Biosci Biotechnol Biochem.
77:1814–1821. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liotta LA, Tryggvason K, Garbisa S, Hart
I, Foltz CM and Shafie S: Metastatic potential correlates with
enzymatic degradation of basement membrane collagen. Nature.
284:67–68. 1980. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lamouille S, Xu J and Derynck R: Molecular
mechanisms of epithelial-mesenchymal transition. Nat Rev Mol Cell
Biol. 15:178–196. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kalluri R: EMT: When epithelial cells
decide to become mesenchymal-like cells. J Clin. Invest.
119:1417–1419. 2009. View
Article : Google Scholar
|
|
15
|
Acloque H, Adams MS, Fishwick K,
Bronner-Fraser M and Nieto MA: Epithelial-mesenchymal transitions:
The importance of changing cell state in development and disease. J
Clin Invest. 119:1438–1449. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kortylewski M, Xin H, Kujawski M, Lee H,
Liu Y, Harris T, Drake C, Pardoll D and Yu H: Regulation of the
IL-23 and IL-12 balance by Stat3 signaling in the tumor
microenvironment. Cancer Cell. 15:114–123. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Rieger-Christ KM, Lee P, Zagha R,
Kosakowski M, Moinzadeh A, Stoffel J, Ben-Ze'ev A, Libertino JA and
Summerhayes IC: Novel expression of N-cadherin elicits in vitro
bladder cell invasion via the Akt signaling pathway. Oncogene.
23:4745–4753. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tong W, Wang Q, Sun D and Suo J: Curcumin
suppresses colon cancer cell invasion via AMPK-induced inhibition
of NF-κB, uPA activator and MMP9. Oncol Lett. 12:4139–4146. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Han M, Song Y and Zhang X: Quercetin
suppresses the migration and invasion in human colon cancer Caco-2
cells through regulating toll-like receptor 4/nuclear factor-kappa
B Pathway. Pharmacogn Mag. 12 Suppl 2:S237–S244. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yun JH, Kim KA, Yoo G, Kim SY, Shin JM,
Kim JH, Jung SH, Kim J and Nho CW: Phenethyl isothiocyanate
suppresses cancer stem cell properties in vitro and in a xenograft
model. Phytomedicine. 30:42–49. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kuo PL and Lin CC: Tetrandrine-induced
cell cycle arrest and apoptosis in Hep G2 cells. Life Sci.
73:243–252. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lee JH, Kang GH, Kim KC, Kim KM, Park DI,
Choi BT, Kang HS, Lee YT and Choi YH: Tetrandrine-induced cell
cycle arrest and apoptosis in A549 human lung carcinoma cells. Int
J Oncol. 21:1239–1244. 2002.PubMed/NCBI
|
|
24
|
Meng LH, Zhang H, Hayward L, Takemura H,
Shao RG and Pommier Y: Tetrandrine induces early G1 arrest in human
colon carcinoma cells by down-regulating the activity and inducing
the degradation of G1-S-specific cyclin-dependent kinases and by
inducing p53 and p21Cip1. Cancer Res. 64:9086–9092. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen XL, Ren KH, He HW and Shao RG:
Involvement of PI3K/AKT/GSK3beta pathway in tetrandrine-induced G1
arrest and apoptosis. Cancer Biol Ther. 7:1073–1078. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
McCubrey JA, Basecke J, Cervello M,
Martelli AM and Franklin RA: GSK-3beta is a critical mediator of
tetrandrine induced cell cycle arrest and cytotoxicity. Cancer Biol
Ther. 7:10792008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liu W, Kou B, Ma ZK, Tang XS, Lv C, Ye M,
Chen JQ, Li L, Wang XY and He DL: Tetrandrine suppresses
proliferation, induces apoptosis, and inhibits migration and
invasion in human prostate cancer cells. Asian J Androl.
17:850–853. 2015.PubMed/NCBI
|
|
28
|
Gao JL, Ji X, He TC, Zhang Q, He K, Zhao
Y, Chen SH and Lv GY: Tetrandrine suppresses cancer angiogenesis
and metastasis in 4T1 tumor bearing mice. Evid Based Complement.
Alternat Med. 2013:2650612013.
|
|
29
|
Horng CT, Yang JS, Chiang JH, Lu CC, Lee
CF, Chiang NN and Chen FA: Inhibitory effects of tetrandrine on
epidermal growth factor-induced invasion and migration in HT29
human colorectal adenocarcinoma cells. Mol Med Rep. 13:1003–1009.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Xu H, Hou Z, Zhang H, Kong H, Li X, Wang H
and Xie W: An efficient Trojan delivery of tetrandrine by
poly(N-vinylpyrrolidone)-block-poly(ε-caprolactone) (PVP-b-PCL)
nanoparticles shows enhanced apoptotic induction of lung cancer
cells and inhibition of its migration and invasion. Int J
Nanomedicine. 9:231–242. 2014.PubMed/NCBI
|
|
31
|
Zhang JS, Li DM, Ma Y, He N, Gu Q, Wang
FS, Jiang SQ, Chen BQ and Liu JR: γ-Tocotrienol induces
paraptosis-like cell death in human colon carcinoma SW620 cells.
PLoS One. 8:e577792013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chang YM, Velmurugan BK, Kuo WW, Chen YS,
HO TJ, Tsai CT, Ye CX, Tsai CH, Tsai FJ and Huang CY: Inhibitory
effect of alpinate Oxyphyllae fructus extracts on Ang II-induced
cardiac pathological remodeling-related pathways in H9c2
cardiomyoblast cells. Biomedicine. 3:148–152. 2013. View Article : Google Scholar
|
|
33
|
Park WH, Seol JG, Kim ES, Hyun JM, Jung
CW, Lee CC, Kim BK and Lee YY: Arsenic trioxide-mediated growth
inhibition in MC/CAR myeloma cells via cell cycle arrest in
association with induction of cyclin-dependent kinase inhibitor,
p21, and apoptosis. Cancer Res. 60:3065–3071. 2000.PubMed/NCBI
|
|
34
|
Lai KC, Hsu SC, Yang JS, Yu CC, Lein JC
and Chung JG: Diallyl trisulfide inhibits migration, invasion and
angiogenesis of human colon cancer HT-29 cells and umbilical vein
endothelial cells, and suppresses murine xenograft tumour growth. J
Cell Mol Med. 19:474–484. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Verhoeff JJ, van Tellingen O, Claes A,
Stalpers LJ, van Linde ME, Richel DJ, Leenders WP and van Furth WR:
Concerns about anti-angiogenic treatment in patients with
glioblastoma multiforme. BMC Cancer. 9:4442009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Cheng SH, Jian JJ, Tsai SY, Chan KY, Yen
LK, Chu NM, Tan TD, Tsou MH and Huang AT: Prognostic features and
treatment outcome in locoregionally advanced nasopharyngeal
carcinoma following concurrent chemotherapy and radiotherapy. Int J
Radiat Oncol Biol Phys. 41:755–762. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Gupta GP and Massague J: Cancer
metastasis: Building a framework. Cell. 127:679–695. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Huang YL, Chu YL, Ho CT, Chung JG, Lai CI,
Su YC, Kuo YH and Sheen LY: Antcin K, an active triterpenoid from
the fruiting bodies of basswood-cultivated antrodia cinnamomea,
inhibits metastasis via suppression of integrin-mediated adhesion,
migration, and invasion in human hepatoma cells. J Agric Food Chem.
63:4561–4569. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Park SJ, Kong HK, Kim YS, Lee YS and Park
JH: Inhibition of S-adenosylhomocysteine hydrolase decreases cell
mobility and cell proliferation through cell cycle arrest. Am J
Cancer Res. 5:2127–2138. 2015.PubMed/NCBI
|
|
40
|
Wu ZY, Lien JC, Huang YP, Liao CL, Lin JJ,
Fan MJ, Ko YC, Hsiao YP, Lu HF and Chung JG: Casticin inhibits
A375.S2 human melanoma cell migration/invasion through
downregulating NF-κB and matrix metalloproteinase-2 and −1.
Molecules. 21:3842016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ji BC, Hsiao YP, Tsai CH, Chang SJ, Hsu
SC, Liu HC, Huang YP, Lien JC and Chung JG: Cantharidin impairs
cell migration and invasion of A375.S2 human melanoma cells by
suppressing MMP-2 and −9 through PI3K/NF-κB signaling pathways.
Anticancer Res. 35:729–738. 2015.PubMed/NCBI
|
|
42
|
Liao CL, Lai KC, Huang AC, Yang JS, Lin
JJ, Wu SH, Wood Gibson W, Lin JG and Chung JG: Gallic acid inhibits
migration and invasion in human osteosarcoma U-2 OS cells through
suppressing the matrix metalloproteinase-2/-9, protein kinase B
(PKB) and PKC signaling pathways. Food Chem Toxicol. 50:1734–1740.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Kessenbrock K, Plaks V and Werb Z: Matrix
metalloproteinases: Regulators of the tumor microenvironment. Cell.
141:52–67. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Shia CS, Suresh G, Hou YC, Lin YC, Chao PD
and Juang SH: Suppression on metastasis by rhubarb through
modulation on MMP-2 and uPA in human A549 lung adenocarcinoma: An
ex vivo approach. J Ethnopharmacol. 133:426–433. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Bauvois B: New facets of matrix
metalloproteinases MMP-2 and MMP-9 as cell surface transducers:
Outside-in signaling and relationship to tumor progression. Biochim
Biophys Acta. 1825:29–36. 2012.PubMed/NCBI
|
|
46
|
Birkedal-Hansen H, Moore WG, Bodden MK,
Windsor LJ, Birkedal-Hansen B, DeCarlo A and Engler JA: Matrix
metalloproteinases: A review. Crit Rev Oral Biol Med. 4:197–250.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Dutta A, Li J, Lu H, Akech J, Pratap J,
Wang T, Zerlanko BJ, FitzGerald TJ, Jiang Z, Birbe R, et al:
Integrin αvβ6 promotes an osteolytic program in cancer cells by
upregulating MMP2. Cancer Res. 74:1598–1608. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Lai KC, Hsu SC, Kuo CL, Ip SW, Yang JS,
Hsu YM, Huang HY, Wu SH and Chung JG: Phenethyl isothiocyanate
inhibited tumor migration and invasion via suppressing multiple
signal transduction pathways in human colon cancer HT29 cells. J
Agric Food Chem. 58:11148–11155. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Matsumura T, Sugimachi K, Takahashi Y,
Uchi R, Sawada G, Ueda M, Hirata H, Sakimura S, Ueo H, Takano Y, et
al: Clinical significance of GAB2, a scaffolding/docking protein
acting downstream of EGFR in human colorectal cancer. Ann Surg
Oncol. 21 Suppl 4:S743–S749. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Waseem T, Duxbury M, Ashley SW and
Robinson MK: Ghrelin promotes intestinal epithelial cell
proliferation through PI3K/Akt pathway and EGFR trans-activation
both converging to ERK 1/2 phosphorylation. Peptides. 52:113–121.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Hsu HH, Hu WS, Lin YM, Kuo WW, Chen LM,
Chen WK, Hwang JM, Tsai FJ, Liu CJ and Huang CY: JNK suppression is
essential for 17β-estradiol inhibits prostaglandin E2-induced uPA
and MMP-9 expressions and cell migration in human LoVo colon cancer
cells. J Biomed Sci. 18:612011. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Xiang T, Fei R, Wang Z, Shen Z, Qian J and
Chen W: Nicotine enhances invasion and metastasis of human
colorectal cancer cells through the nicotinic acetylcholine
receptor downstream p38 MAPK signaling pathway. Oncol Rep.
35:205–210. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Zhao GY, Ding JY, Lu CL, Lin ZW and Guo J:
The overexpression of 14-3-3ζ and Hsp27 promotes non-small cell
lung cancer progression. Cancer. 120:652–663. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Jeong D, Park S, Kim H, Kim CJ, Ahn TS,
Bae SB, Kim HJ, Kim TH, Im J and Lee MS: RhoA is associated with
invasion and poor prognosis in colorectal cancer. Int J Oncol.
48:714–722. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Clevers H: Wnt/β-catenin signaling in
development and disease. Cell. 127:469–480. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
White BD, Chien AJ and Dawson DW:
Dysregulation of Wnt/β-catenin signaling in gastrointestinal
cancers. Gastroenterology. 142:219–232. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Norwood MG, Bailey N, Nanji M, Gillies RS,
Nicholson A, Ubhi S, Darnton JJ, Steyn RS, Womack C, Hughes A, et
al: Cytoplasmic beta-catenin accumulation is a good prognostic
marker in upper and lower gastrointestinal adenocarcinomas.
Histopathology. 57:101–111. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Kobayashi M, Honma T, Matsuda Y, Suzuki Y,
Narisawa R, Ajioka Y and Asakura H: Nuclear translocation of
beta-catenin in colorectal cancer. Br J Cancer. 82:1689–1693.
2000.PubMed/NCBI
|
|
59
|
Guttridge DC, Albanese C, Reuther JY,
Pestell RG and Baldwin AS Jr: NF-kappaB controls cell growth and
differentiation through transcriptional regulation of cyclin D1.
Mol Cell Biol. 19:5785–5799. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Yamamoto Y and Gaynor RB: Therapeutic
potential of inhibition of the NF-kappaB pathway in the treatment
of inflammation and cancer. J Clin Invest. 107:135–142. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Fryknas M, Dhar S, Oberg F, Rickardson L,
Rydåker M, Göransson H, Gustafsson M, Pettersson U, Nygren P,
Larsson R and Isaksson A: STAT1 signaling is associated with
acquired crossresistance to doxorubicin and radiation in myeloma
cell lines. Int J Cancer. 120:189–195. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Roberts D, Schick J, Conway S, Biade S,
Laub PB, Stevenson JP, Hamilton TC, O'Dwyer PJ and Johnson SW:
Identification of genes associated with platinum drug sensitivity
and resistance in human ovarian cancer cells. Br J Cancer.
92:1149–1158. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Hasegawa K, Ichikawa W, Fujita T, Ohno R,
Okusa T, Yoshinaga K and Sugihara K: Expression of cyclooxygenase-2
(COX-2) mRNA in human colorectal adenomas. Eur J Cancer.
37:1469–1474. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Peek RM Jr: Prevention of colorectal
cancer through the use of COX-2 selective inhibitors. Cancer
Chemother Pharmacol. 54 Suppl 1:S50–S56. 2004.PubMed/NCBI
|
|
65
|
Dempke W, Rie C, Grothey A and Schmoll HJ:
Cyclooxygenase-2: A novel target for cancer chemotherapy? J Cancer
Res Clin Oncol. 127:411–417. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Giercksky KE: COX-2 inhibition and
prevention of cancer. Best Pract Res Clin Gastroenterol.
15:821–833. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Zhang H and Sun XF: Overexpression of
cyclooxygenase-2 correlates with advanced stages of colorectal
cancer. Am J Gastroenterol. 97:1037–1041. 2002. View Article : Google Scholar : PubMed/NCBI
|