Introduction
Pancreatic cancer is recognized as a highly
malignant and incurable disease worldwide. In 2016, an estimated
53,070 new cases were diagnosed and 41,780 people eventually died
from pancreatic cancer, making it the fourth leading cause of
cancer deaths in the USA. Although the survival rate has increased
for most cancers, improvements have been marginal for patients with
pancreatic cancer; the 5-year relative survival is merely 8%
(1). The dismal prognosis for
pancreatic cancer is attributed to multiple factors, such as the
advanced stage of initial detection, the high risk for distant
metastasis, and a subpar response to conventional therapies.
Sequencing the genomes of pancreatic cancers indicated 12 important
signaling pathways that do not promote distant metastasis until
nearly 20 years after the initial mutation occurred. This evidence
has offered a time frame for pancreatic carcinogenesis, suggesting
that a large window of opportunity may exist for early detection,
which could improve the prognosis of this lethal disease (2,3). Thus,
further studies are required into the molecular pathology that
orchestrates the process of pancreatic tumorigenesis, in order to
improve personalized diagnostics and therapeutic regimens.
Exosomes, which are secreted by multiple cell types
and by cancer cells, contain functional biomolecules (including
proteins, nucleic acids and lipids) that can be horizontally
delivered to recipient cells. They are distinguished from apoptotic
bodies and microvesicles by their heterogeneous size, origin and
composition (4,5). Exosomes (30–150 nm in diameter) are
extracellular vesicles generated by the inward budding of the
membrane (endocytosis), which leads to the subsequent formation of
multivesicular bodies and their release by exocytosis. Thus, on
their membranes, exosomes express a variety of markers of
multivesicular bodies, such as tetraspanins. Ectosomes (0.1–2 µm in
diameter) are formed by outward blebbing from the plasma membrane
and are subsequently released by proteolytic cleavage from the cell
surface, thus expressing antigens of the origin cell. Apoptotic
bodies (1–4 µm in diameter) are generated upon apoptotic cell death
(6,7).
Exosomes are the most prominent mediators of
intercellular communications that regulate, instruct, and
re-educate their surrounding microenvironment and target organs
(8,9).
The interaction between exosomes and recipient cells is mediated by
the membrane proteins of exosomes, such as the phosphatidylserine
and proteoglycan receptors (9,10).
Additionally, the presence of particular tetraspanins or integrins
on the exosomal membrane is likely to play a vital role in
selectively acting on their target cells or organs (11,12).
Exosome-associated diagnosis and follow-up has been extensively
promoted due to the easy isolation and identification of exosomes
in body fluids, especially the peripheral blood (13). Furthermore, Li et al (14) identified >1,000 circular RNAs
(circRNAs) in human plasmatic exosomes. Their study indicated the
potential of circRNAs as a novel exosome-associated tumor biomarker
and discussed the molecular functions of exosomal circRNAs.
Components and functions of exosomes
A systematic search of ExoCarta (http://www.exocarta.org/), EVpedia (http://student4.postech.ac.kr/evpedia2_xe/xe/) and
Vesiclepedia (http://microvesicles.org/index.html#) was performed to
select the most important components of exosomes. The top five most
common identified exosomal proteins were CD9, heat shock protein
family A member 8 (HSPA8), programmed cell death 6 interacting
protein (PDCD6IP), glyceraldehyde-3-phosphate dehydrogenase
(GADPH), and actin β (ACTB); the majority of these are known to be
implicated in the biogenesis, sorting and release of exosomes. The
simple structure and components of exosomes are summarized in
Fig. 1.
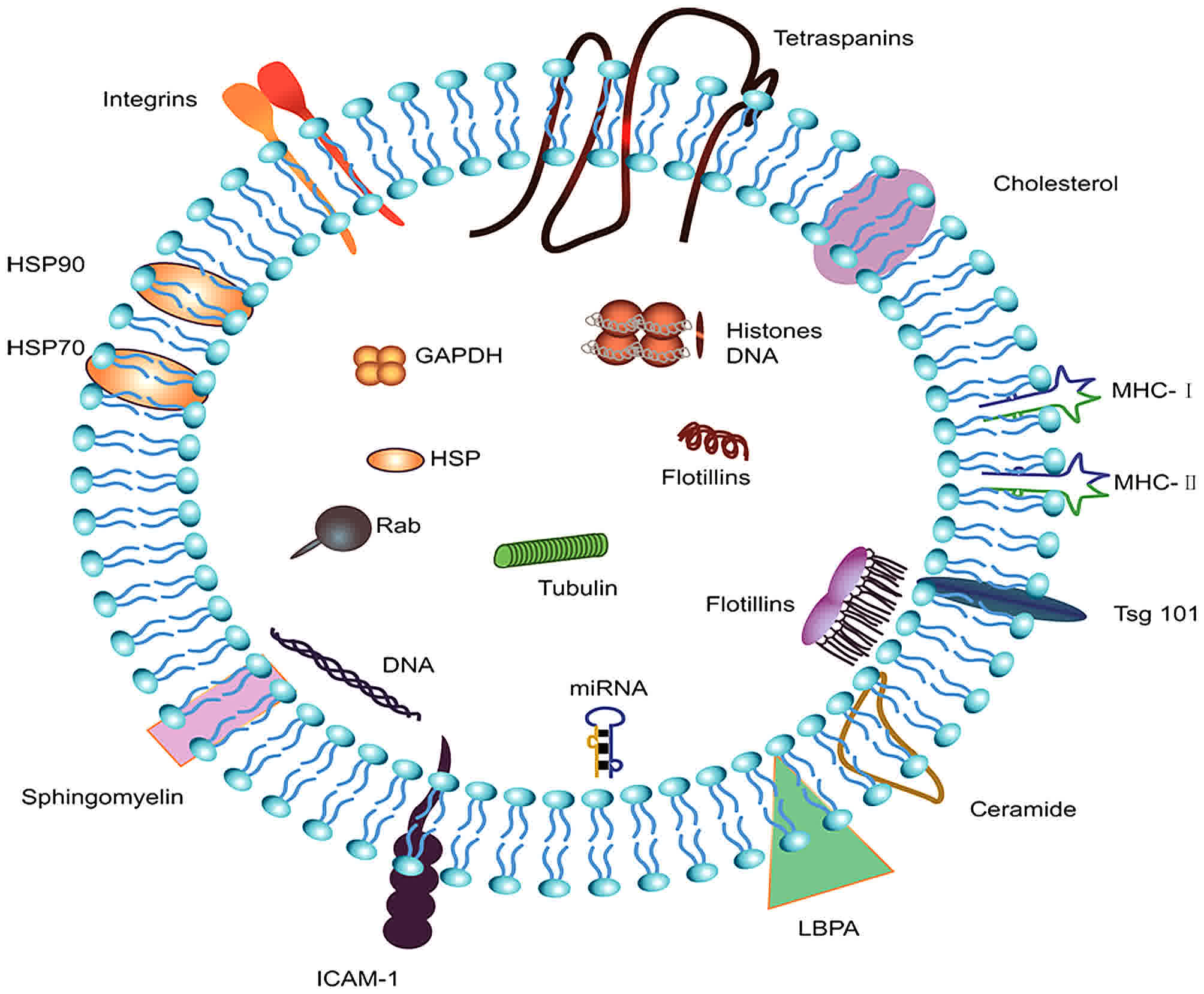 | Figure 1.Components and structure of exosomes.
Exosomes are small vesicular structures carrying various pathogenic
miRNAs, mRNAs, DNA fragments, and proteins. The top 5 most commonly
identified exosomal proteins are CD9, HSPA8, PDCD6IP, GADPH, and
ACTB. HSPA8, heat shock protein family A member 8; PDCD6IP,
programmed cell death 6 interacting protein; GAPDH,
glyceraldehyde-3-phosphate dehydrogenase; ACTB, actin β; HSP, heat
shock protein; ICAM-1, intercellular adhesion molecule 1; MHC,
major histocompatibility complex; LBPA, lysobisphosphatidic
acid. |
Lipids
Exosomes consist of a lipid bilayer, and
transmembrane proteins have crucial roles in the development of a
mosaic model. Lipids exert a vital function in maintaining the
rigidity and stability of the exosomal membrane, as well as in the
processes of fusion and budding. Exosomal RNA is well-protected in
exosomes due to the integrity of the membrane (15). In particular, exosomal membranes have
abundant phosphatidylserine, monosialodihexosylganglioside
cholesterol and sphingomyelin (16).
Exosomal lipids can carry other lipids with biological activities,
as well as second messengers, such as phosphatidic acid, ceramides
and diglycerides, that are implicated in exosome biogenesis.
Beloribi et al (17)
demonstrated that exosomal lipids initiated apoptosis in human
pancreatic carcinoma SOJ-6 cells by inhibiting the Notch-1 pathway.
Exosomal lipids also led to drug resistance in human pancreatic
cancer MiaPaCa-2 cells via the C-X-C motif chemokine receptor 4
(CXCR4)/stromal cell derived factor (SDF)-1α signaling axis, which
implied that exosomal lipids might function in the progression and
drug resistance of pancreatic cancer (18). At present, there is only one study on
prostate cancer that indicates that exosomal lipids have the
potential to serve as cancer biomarkers (19). It is possible that progress in
lipidomics will aid in the improved understanding of the biological
mechanisms of exosomal formation, sorting and release under
physiological and pathological conditions.
Proteins
Several studies on exosomes from multiple
organs/cell types and cell lines, under physiological and
pathophysiological conditions, have demonstrated the enrichment of
proteins in exosomes (20–22). Exosomal proteins differ vastly by the
type and state of the cell the exosomes were derived from,
suggesting that they may be ideal candidates for cancer biomarkers.
To date, >7,000 proteins have been identified through
significant technical improvements in mass spectrometry. Proteomic
analysis of exosomes from multiple tissues indicate that exosomes
are comprised of some common vesicular proteins, such as
tetraspanins [CD9, CD82, CD81, CD63, CD151 and tetraspanin 8
(Tspan8)], which are constituent components of exosomes (23,24). The
tetraspanins CD151 and Tspan8 are exosome components that are
critical for the interactions between tumor-initiating cells and
the tumor microenvironment (25).
Cytoplasmic and plasma membrane proteins are more commonly released
into exosomes than mitochondrial and nuclear proteins, which
indicates that membrane proteins have great potential to be
candidates for protein markers (26,27).
Exosomes also contain cell type-specific proteins,
which are particularly useful for the identification of specific
surface biomarkers. Most of the cell type-specific surface
biomarkers have been validated as cancer and cancer stem
cell-associated markers. Among them, melanoma tumor exosomes
express the tumor-related protein melanoma antigen recognized by T
cells 1 (MART1), while epithelial cell-originated tumor exosomes
express the epithelial cell adhesion molecule (EpCAM) (27,28).
Glioblastoma tumor exosomes express epidermal growth factor
receptor (EGFR) variant III (29) and
the tumor exosomes of docetaxel-resistant prostate cancer cells
express the multidrug resistance gene 1 (MDR-1) (30).
Furthermore, it has been proposed that mutations in
>1% of the human genome are correlated with carcinogenesis;
mutant proteins, such as mutant KRAS, the oncoprotein MET and
tissue factor proteins that uniquely appear in pathological cells,
have been validated to be released via exosomes (31–33). EGFR
is secreted by five pancreatic cancer cell lines in exosomes as two
distinct EGFR forms. The identification of EGFR as a plasma
biomarker and a personalized therapeutic is promising in pancreatic
cancer (34).
Nucleic acids
Exosomes also contain microRNA (miRNAs), tRNA, rRNA
mRNA, DNA, and the recently identified circRNA (14,35–37).
Exosomes interact with their target cells mainly via membrane
fusion and transfer of exosomal contents. miRNAs could deliver
biological information that regulates the function of target
cells.
miRNAs are 19–25 nt long non-coding RNAs that
repress the stability and translation of mRNAs, controlling genes
implicated in many cellular processes. They have important roles in
the mediation of nearly all signaling pathways within a cell, and
their dysfunction is associated with the initiation, progression
and metastasis of cancer, including pancreatic cancer (38–40).
Furthermore, miRNA-155 was validated to control exosome synthesis
and to promote gemcitabine resistance in pancreatic cancer.
Targeting therapy to miR-155 or exosome secretion effectively
attenuated tumor chemoresistance, representing a novel therapeutic
target for gemcitabine treatment in pancreatic cancer (41). In pancreatic cancer, Zinc finger
E-box-binding homeobox (ZEB)1 and ZEB2 are important mediators of
the epithelial-to-mesenchymal transition and maintain stemness by
suppressing the miR-200 family, thereby promoting migration
(42). Manier et al (43) identified miRNAs as the most prominent
small RNAs present in exosomes isolated from the serum of multiple
myeloma patients and healthy controls by using small RNA sequencing
on circulating exosomes. Two of these miRNAs, namely, let-7b and
miR-18a, were significantly associated with both progression-free
survival and overall survival in a univariate analysis. These
findings support the use of circulating exosomal miRNAs to improve
the identification of patients with newly diagnosed multiple
myeloma with poor prognosis. The presence of miRNAs in numerous
body fluids, especially serum, correlates with the recovery of
solid cancers and hematologic malignancies and may be valuable for
the early diagnosis of multiple types of cancer, potentially
offering superior performance over conventional methods for cancer
diagnosis (44,45).
Representing a novel type of endogenous non-coding
RNAs, circRNAs have gained attention recently for their involvement
in multiple biological processes. CircRNAs are ubiquitously
expressed in eukaryotic cells and modulate gene expression by
acting as sponges of miRNAs or proteins, such as RNA-binding
proteins (RBPs) (46,47). Li et al (14) demonstrated that circRNA is enriched
and stable in exosomes, identified up to 1,000 serum exosomal
circRNAs, and discriminated between patients with colon cancer and
healthy controls; expression patterns of circ-KLDHC10 were
dramatically enhanced in serum from cancer patients. Their finding
that serum exosomal circRNAs could discriminate patients with
cancer from normal controls suggests exosomal circRNAs have great
translational potential as a circulating biomarker for the early
detection of cancer.
Recently, lncRNAs were demonstrated to exist in
exosomes, and they could function as potential stable biomarkers
for cancer patients. Li et al (48) have found that plasma Long intergenic
non-protein-coding RNA 152 (LINC00152) in exosomes was a potential
biomarker and highly stable in patients with gastric cancer. A
similar result revealed that the exosomal transfer of the lncRNA
mediator of reprogramming (linc-RoR) may be involved in
hepatocellular carcinoma progression and treatment failure
(49). These findings demonstrate
that the components of serum-derived exosomes can be utilized to
predict, treat and evaluate therapy resistance for many
cancers.
The role of exosomes in the pathogenesis of
pancreatic cancer
Exosomes engage in tumorigenesis and
the aggressive behavior of pancreatic cancer
Invasion is one of the primary features of
pancreatic cancer, and failures in pancreatic cancer therapy are
generally due to metastasis, because distant lesions are generally
resistant to most chemotherapeutic agents. Metastasis is a
complicated, sequential event and is a sign of malignant cancer.
Metastasis-initiating cells that disseminate from invasive
carcinomas disperse via the blood or lymphatic vessels. Circulating
tumor cells that survive in the bloodstream subsequently migrate to
target tissues. Infiltrated cells in the new microenvironment can
survive chemoradiotherapy and initiate cancer regrowth, leading to
cancer recurrence and metastasis (50). These steps are supported by the
complex interaction of cancer cells with the tumor stroma that
involves multiple intercellular communications pathways (51,52).
Recently, there has been an increase in the number
of studies demonstrating the important function exerted by exosomes
in cancer initiation, progression and distant metastasis. For
instance, exosomes have been verified to have the ability to
transform normal cells into cancer cells in breast and prostate
cancer (53,54). Exosomes promote cancer cell
proliferation by delivering carcinogenic proteins,
cell-cycle-associated mRNAs, specific miRNAs, cytokines and
adhesion molecules to cancer cells (55–57). They
can deliver tumor suppressor miRNA, such as miR-200, to confer
metastatic properties and promote breast cancer cell metastasis
(58). Cancer cell-derived exosomes
can also activate the phosphoinositide 3-kinase (PI3K)/AKT
serine/threonine kinase 1 (Akt) and mitogen-activated protein
kinase (MAPK) pathways to facilitate the proliferation of gastric
cancer cell lines (59). However, a
different result was obtained in pancreatic cancer cells, where
exosome lipids led to cancer cell apoptosis via the repression of
the Notch-1 survival signaling pathway (17). Another study (18) in pancreatic cancer cell line,
MiaPaCa-2, revealed that exosomal lipids could induce the
activation of nuclear factor (NF)-kB, which facilitated the
expression and release of SDF-1α. The binding of SDF-1α and its
receptor CXCR4 activated the Akt signaling pathway, inhibiting cell
death (18). This finding suggested
that exosomal lipids may be involved in pancreatic cancer
development and drug resistance. Exosomes derived from pancreatic
stellate cells contained abundant miR-21-5p and miR-451a.
Exosome-mediated delivery of these components enhanced
proliferation, migration and the specific gene expression of
pancreatic cancer cells (60).
Costa-Silva et al (8) demonstrated that pancreatic
cancer-derived exosomes could lead to liver pre-metastatic niche
initiation in mice and could gradually increase the cancer niche
burden. Primary pancreatic cancer-derived exosomes alter the
pre-metastatic liver microenvironment by initiating an inflammatory
response, which results in the preferential implantation of
pancreatic cancer cells in the liver. Kupffer cells selectively
absorb the exosomes released by pancreatic cancer and can increase
the secretion of transforming growth factor β (TGFβ). TGFβ
activates hepatic stellate cells and then upregulates fibronectin
production. The fibrotic microenvironment induces the accumulation
of bone marrow-derived cells, such as neutrophils and macrophages.
These cells bind primarily to fibronectin-associated hepatic sites,
consequently facilitating the initiation of the liver
pre-metastatic niche. Further study revealed that the macrophage
migration inhibitory factor (MIF) was highly abundant in pancreatic
cancer-derived exosomes, which orchestrated the liver metastasis
(8). Compared with subjects who had a
bulky primary tumor, exosomal MIF was significantly upregulated in
early stage pancreatic cancer patients who later developed distant
liver metastasis. The identification of MIF pancreatic
cancer-derived exosomes has opened novel avenues for the biological
mechanisms of metastasis and might be a potential prognostic marker
for the progression of pancreatic cancer metastasis.
An et al (61)
and colleagues used proteomic analysis to quantify the protein
level of serum exosomes from patients with locally advanced
pancreatic cancer undergoing chemoradiotherapy. They isolated
exosomes from the serum of 10 patients with locally advanced
pancreatic cancer at serial time points over the course of therapy,
and quantitative analysis was performed using the iTRAQ method.
They detected approximately 700–800 exosomal proteins per sample,
several of which have been implicated in metastasis and treatment
resistance. They compared the exosomal proteome of patients at
different time points during treatment to healthy controls and
identified eight proteins that exhibited global treatment-specific
changes. They then tested the effect of patient-derived exosomes on
the migration of tumor cells and demonstrated that patient-derived
exosomes, but not exosomes from healthy controls, induced cell
migration, supporting their role in metastasis. Their data
demonstrated that exosomes can be reliably extracted from patient
serum and analyzed for protein content. The differential loading of
exosomes during the course of therapy suggests that exosomes may
provide novel insights into the development of treatment resistance
and metastasis.
Exosome-mediated immunosuppression in
pancreatic cancer
Emerging evidence has demonstrated that
communications between tumor and immune cells through exosomes have
a dynamic and complex role in regulating tumor immunity, which is
determined by specific proteins and genetic components (62). During pancreatic cancer progression
and distant metastasis, most cancer cells are suppressed by the
immune response. Most patients with pancreatic cancer are in a
condition of immune disorder or immunosuppression. Increasing
studies point to the well-documented role of pancreatic
cancer-derived exosomes in immunosuppression.
Representative literature suggests that pancreatic
cancer-derived exosomes modulate the expression pattern of
toll-like receptor 4 (TLR4) in dendritic cells (DCs) through
miRNA-203. When absorbed by dendritic cells, these exosomes
decreased TLR4 expression and the downstream associated cytokines,
such as interleukin (IL)-12 and tumor necrosis factor (TNF)-α.
Thus, exosomes attenuate DC-mediated tumor suppressive responses
initiated by TLR4 (63). Similarly,
Ding et al (64) have
demonstrated that pancreatic cancer-associated miRNAs could be
delivered to DCs through exosomes and suppress target mRNA
expression. Furthermore, pancreatic cancer-generated exosomes
restrain regulatory factor X-associated protein expression by means
of miR-212-3p, which could decrease the expression level of major
histocompatibility complex (MHC) II and result in the immune
tolerance of DCs. However, pancreatic cancer-derived exosomal
miRNAs can inhibit the tumor suppressive function of
DC/cytokine-induced killer cells (CIKs), and miRNA-depleted
exosomes may increase the tumor suppressive activity of DC/CIKs. In
summary, pancreatic cancer-derived exosomes regulate host
immunosuppression via regulating the function, maturation and
differentiation of DCs.
Macrophages exhibit dramatic plasticity and alter
their physiological state based on environmental conditions,
especially the cancer microenvironment (65). A previous study has revealed that the
content of pancreatic cancer-derived exosomes was altered by
transfection with miR-155 and miR-125b2, inducing differential
communication and remodeling the macrophage polarization to the M1
phenotype (66). This alteration can
cause an inhibitory effect on pancreatic cancer invasion and
metastasis. Another study provides evidence that salivary exosomes
from mice with pancreatic ductal adenocarcinoma exhibit a
suppressive effect that results in reduced tumor-killing capacity
by NK cells (67). Salivary exosomes
from mice with pancreatic ductal adenocarcinoma where pancreatic
tumors were engineered to suppress exosome biogenesis failed to
suppress NK cell cytotoxicity against tumor cells, as opposed to
salivary exosomes from mice with pancreatic ductal adenocarcinoma
with normal tumor exosome biogenesis (67). According to these findings,
immunotherapy aimed towards alterative manipulation of cancer
cell-derived exosome components, such as miRNAs, may have
therapeutic potential in treating pancreatic cancer.
Clinicopathological characteristics of
pancreatic cancer associated with exosomes
Weight loss and new-onset diabetes are
characteristics of the paraneoplastic effect of pancreatic cancer,
and these phenomena precede the diagnosis of the disease (68). A previous study suggested that
exosomal adrenomedullin serves a crucial role in the initiation of
diabetes and lipolysis in adipocytes (69). The biological mechanism of pancreatic
cancer-associated diabetes is complicated. A potential mechanism
has been revealed where pancreatic cancer-derived exosomal
adrenomedullin, triggers endoplasmic reticulum stress and
dysfunction of the unfolded protein response, resulting in β-cell
disorder and apoptosis. Further experiments demonstrated that
exosome-mediated transfer of adrenomedullin into β-cells was
regulated through micropinocytosis and caveolin-dependent
endocytosis (70).
Adipose tissue lipolysis is partly responsible for
weight loss, which is one of the alarming symptoms associated with
pancreatic cancer. Exosomal adrenomedullin, a candidate mediator,
promotes lipolysis in adipocytes by selectively targeting its
receptor. This interaction induces the activation of the p38 and
extracellular signal-regulated kinase (ERK)1/2 MAP kinase signaling
pathways that can promote the phosphorylation of hormone-sensitive
lipase, therefore facilitating lipolysis (69,71).
Exosomes regulate therapy resistance in
pancreatic cancer
Up to 74% of patients with pancreatic cancer who
receive adjuvant therapy with gemcitabine ultimately present with
rapid tumor regression and treatment failure (72). The elucidation of the detailed
mechanisms that lead to chemotherapy resistance is of great
importance for improving pancreatic cancer survival. Exosomes
induce the progression of chemotherapy resistance in cancer cells
via various mechanisms. Cancer-derived exosomes could deliver
multiple drug resistance-related proteins and miRNAs to specific
cells (30,73). Additionally, chemotherapy agents could
also be encapsulated and secreted from the extracellular matrix by
exosomes, and this mechanism partly accounts for chemotherapy
resistance (51).
Much of the focus on cancer therapy concentrates on
inhibiting the abnormal proliferation of epithelial cells. However,
this approach has been proven ineffective, with resistance emerging
in many types of cancer following adjuvant chemotherapy. Stromal
interaction with cancer cells could affect the response to
chemotherapy. A detailed hypothesis on the mechanism of resistance
has focused on heterotypic interactions, where exosomes are
delivered from cancer-associated fibroblasts (CAFs) to pancreatic
cancer cells (74). CAFs are
inherently insensitive to gemcitabine. When exposed to gemcitabine,
pancreatic fibroblasts dramatically increase the release of Snail
and miR-146a via exosomes. Utilizing paracrine or potentially an
unknown signaling mechanism, these exosomes can be taken up by
recipient epithelial cells. During the process, the amount of
miR-146a and Snail mRNA in epithelial cells is enhanced. As a
result, CAF-derived exosomes facilitate proliferation and
chemotherapy resistance. Notably, CAF exosome secretion inhibition
could remarkably decrease cell survival, proliferation and drug
resistance.
Inhibitors of apoptosis (IAPs) could facilitate cell
death, but their expression pattern is decreased in various
cancers. In pancreatic cancer, it was confirmed that IAPs are
constitutively enhanced by NF-κB in tissues and cell lines
(75). This ectopic overexpression of
IAPs is associated with cancer progression and chemotherapy
resistance. It has been reported that exosomes contain protein and
mRNA IAPs (76). When exposed to
chemotherapy, the amount of IAP protein or mRNA in the cytoplasm
either remained unchanged or was moderately upregulated. Similarly,
the expression pattern of IAPs in exosomes exhibited no significant
alteration. This stability may partly account for why pancreatic
cancer is resistant to chemotherapy (75).
Exosomal miRNAs and proteins for pancreatic
cancer diagnosis
Strong efforts have focused on developing sensitive
diagnostics tools improving early detection of pancreatic cancer
via identifying pancreatic cancer-associated exosomal markers. The
development of next generation sequencing, novel mass spectrometry
techniques and new biochemical analyses have rendered the
characterization of exosomes more convenient, which means that all
exosomal contents have the potential to function as diagnostic
biomarkers. Based on a comprehensive literature search, exosomal
membrane proteins and miRNAs have provided promising insights into
pancreatic cancer progression and tumor burden. Potential exosomal
biomarkers for the early detection of pancreatic cancer are
summarized in Table I.
 | Table I.Exosomal biomarkers and early
detection of pancreatic cancer. |
Table I.
Exosomal biomarkers and early
detection of pancreatic cancer.
| Disease | Study | Exosome
isolation | Exosomal
biomarkers | Sensitivity | Specificity |
|---|
| Primary
pancreatic | Que et al
(2013) |
Ultracentrifugation | miR-21 | 95.5% | 81.5% |
| cancer | (77) |
| miR-17-5P | 72.7% | 92.6% |
|
| Madhavan et
al |
Ultracentrifugation | miR-1246 | 100% | 80% |
|
| (2015) (80) | and
immunoaffinity | miR-4644 |
|
|
|
|
|
| miR-3976 |
|
|
|
|
|
| miR-4306 |
|
|
|
| Joshi et al
(2015) | LSPR-based
assay | miR-10b | Statistically | Statistically |
|
| (79) |
|
| significant | significant |
|
| Melo et al
(2015) (81) |
Ultracentrifugation | GPC1 protein | 100% | 100% |
| Pancreatic
cancer | Costa-Silva et
al |
Ultracentrifugation | MIF | Statistically | Statistically |
| liver
pre-metastasis | (2015) (8) |
|
| significant | significant |
|
| Hoshino et
al |
Ultracentrifugation | Integrins αvβ5 | Statistically | Statistically |
|
| (2015) (11) |
|
| significant | significant |
A recent study evaluated the expression patterns of
four pancreatic cancer-associated miRNAs in circulating exosomes
(77). Compared with multiple
controls, the expression profile of exosomal miR-21 and miR-17-5p
was remarkably enhanced in pancreatic cancer patients, and this
distinction could be utilized to discriminate between pancreatic
cancer and non-malignant chronic pancreatitis (CP) patients. With
the application of this innovative technique, label-free,
nanoplasmonic-based short non-coding RNA sensing, a distinctly
enhanced amount of exosomal miR-10b was observed in patients with
pancreatic cancer compared with CP or normal controls (78,79).
Madhavan et al (80) published
a study based on the analysis of five pancreatic cancer-initiating
cell markers (CD104, MET, EpCAM, Tspan8 and CD44v6) and four miRNAs
(miR-4306, miR-3976, miR-4644 and miR-1246) in serum exosomes, and
demonstrated that evaluation of pancreatic cancer-initiating cell
markers and miRNA serum-exosome marker panels dramatically enhanced
sensitivity (1.00, CI: 0.95–1) with a specificity of 0.80 (CI:
0.67–0.90) for pancreatic cancer vs. all others groups and of 0.93
(CI: 0.81–0.98) excluding non-pancreatic cancer. Concomitant
evaluation of these factors distinguished patients with pancreatic
cancer from normal controls, CP subjects and patients with benign
pancreatic lesions, with 93% specificity, eliminating
non-pancreatic neoplasms (80).
Encouraging advances have been achieved in the
search for specific and sensitive biomarkers that could facilitate
the early diagnosis of pancreatic cancer. Melo et al
(81) reported that glypican-1 (GPC1)
expression patterns in exosomes secreted by pancreatic cancer could
be utilized to identify subjects with pancreatic cancer early and
offer considerable insights into the disease progress and tumor
load. A comparison of exosomes from pancreatic cancer and control
cell lines indicated that the exosomes from cancer exhibited
enhanced levels of GPC1, which has been demonstrated to be
upregulated in breast cancer and pancreatic cancer. In serum
specimens from subjects with pancreatic cancer (n=190), a
significantly larger amount of GPC1+ circulating
exosomes was present compared with normal controls (n=100).
Furthermore, GPC1+ exosomes were also confirmed to
contain identical KRAS mutations, which frequently are present in
pancreatic cancer and precancerous lesions and have been considered
a fundamental mutation.
Notably, exosomes from patients with intraductal
papillary mucinous neoplasm (IPMN; a precancerous lesion of
pancreatic cancer) expressed dramatically higher levels of GPC1
than subjects with pancreatitis or cystic adenomas (81). The identification of GPC1+
circulating exosomes had 100% specificity in differentiating IPMN
from healthy individuals or benign pancreatic disease. When the
efficiency of GPC1+ circulating exosomes was compared to
carbohydrate antigen 19-9 (CA19-9) in differentiating benign from
precancerous lesion and healthy subjects, GPC1+
circulating exosomes was validated to be prominently superior.
Tumor burden was associated positively with levels of
GPC1+ circulating exosomes. In most subjects, the
exosome levels reduced following the removal of the solid tumor. A
decrease in levels of GPC1+ circulating exosomes was
associated with the benefit of overall and disease-specific
survival in these patients. Additionally, using a genetically
engineered mouse that progressively developed into pancreatic
cancer, the identification of GPC1+ exosomes exhibited positive
results prior to pancreatic lesions being detectable. Another
recent paper (11) has reported that
integrin αvβ5-expressing pancreatic cancer exosomes can determine
liver metastasis. Targeting integrin αvβ5 reduced exosome
absorption by resident cells, and inhibited liver metastasis. The
levels of integrin αvβ5 were dramatically enhanced in exosomes
isolated from pancreatic cancer subjects with live metastasis
compared with subjects with no distant metastasis. Further
examination of the molecular mechanisms revealed that integrin αvβ5
uptake by targeted cells could lead to activating Src
phosphorylation and upregulation of pro-inflammatory S100 gene.
Collectively, these findings have indicated that GPC1+
and integrin αvβ5-expressing circulating exosomes have great
potential as indicators of pancreatic cancer progression and
organ-specific metastasis in patients.
A schematic of the roles that exosomes may have in
pancreatic cancer metastasis and their potential for early
detection is summarized in Fig.
2.
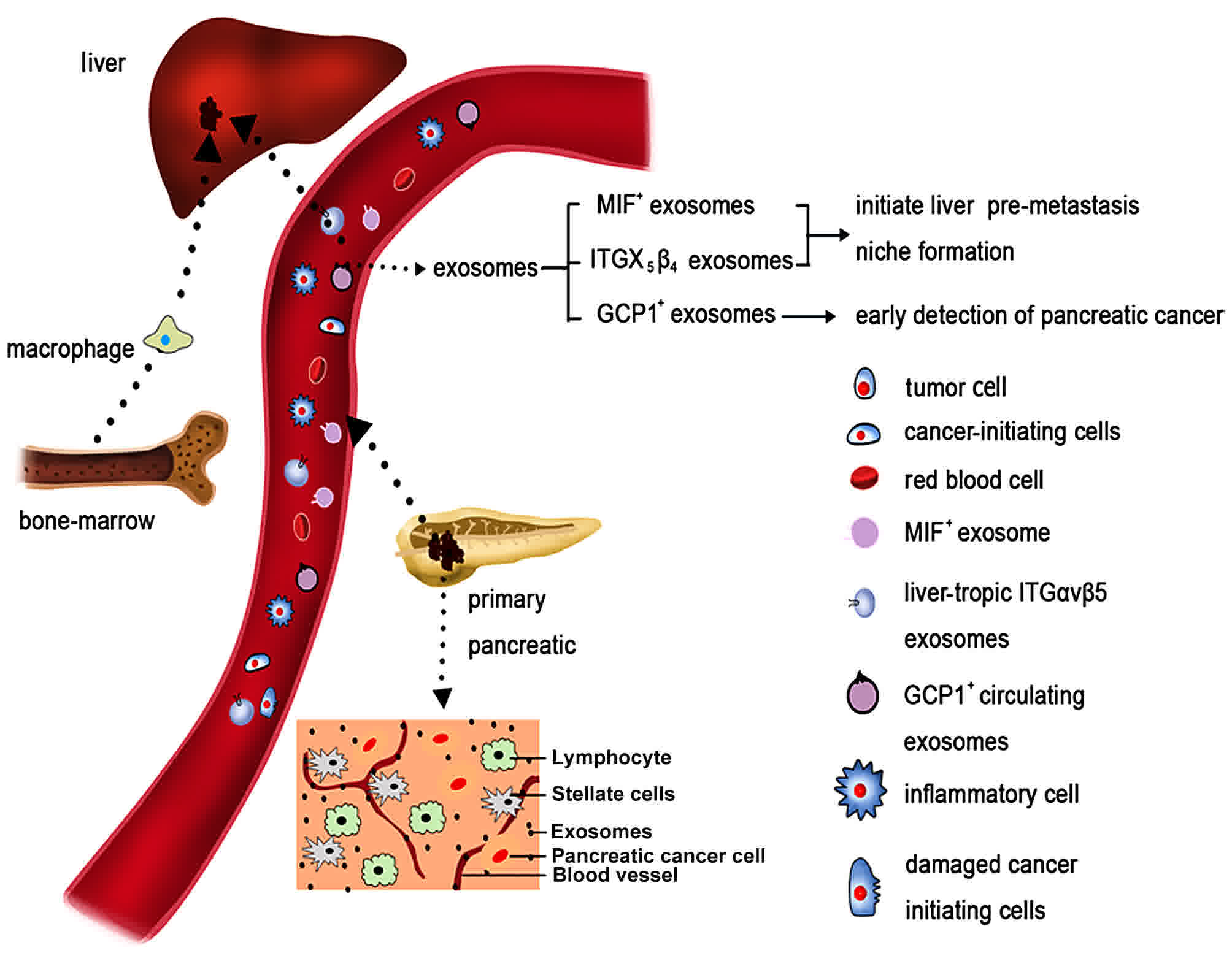 | Figure 2.Pancreatic cancer, its
microenvironment and its metastatic route. Pancreatic cancer is
characterized by a dense stromal reaction, consisting of a
desmoplastic reaction with the extracellular matrix, CAFs, tumor
cells and immune cells. Both pancreatic cancer cells and CAFs can
release exosomes. Exosomes released by pancreatic cancer cells
enter the vessels. Due to specific integrin complex expression,
exosomes migrate preferentially to the liver. There, they are taken
up by Kupffer cells (also known as stellate macrophages), which
react by increasing TGF-β signaling. Subsequently, macrophage MIF
is released from the exosomes, which results in the initiation of
an immune-evasive response. This model supports the creation of a
metastatic niche for pancreatic cancer cells mediated by exosomes,
which is then followed by the establishment of liver metastases.
CAFs, cancer-associated fibroblasts; TGF, transforming growth
factor; MIF, migration inhibitory factor; ITG, integrin; GCP1,
glypican-1. |
Conclusion
Circulating exosomes have demonstrated potential as
reliable candidates for the early diagnosis of pancreatic cancer,
for use in screening high-risk individuals without clinical
presentation of cancer, and for monitoring the course of the
disease. Accumulating evidence has revealed that exosomes can
indicate the clinical management of pancreatic cancer, improve
overall survival and prevent organ-specific metastasis.
Nevertheless, the clinical application of exosomes in the
differential diagnosis of pancreatic cancer requires further
research. More studies are needed to better elucidate the dynamic
alteration of serous exosomes in the progression of pancreatic
cancer, to evaluate the possibility of plasmatic exosomal biomarker
panels for high-risk individuals in a large-scale cohort, and
understand the cellular and molecular functions of exosomes in
pancreatic cancer.
Future perspective
The prognosis of patients with pancreatic cancer has
improved slowly during the past decade, and the low survival rate
is due to multiple factors, but, perhaps the predominant
determinant of survival is the advanced stage at which most
subjects are diagnosed. The development of biomarkers to improve
detection at early stages could apply to high-risk individuals,
such as those with weight loss and new-onset diabetes. New-onset
diabetes and concurrent weight loss have been demonstrated to occur
a few months prior to the clinical manifestation of pancreatic
cancer (68). Multiple studies have
demonstrated that new-onset diabetes of <3 years enhances the
risk of pancreatic cancer diagnosis 5–7-fold, which is
significantly higher than the effect of long-term diabetes, which
only accounts for a 1.5-fold increase in risk within a course of
>5 years (82,83). The discovery of novel
exosome-associated biomarkers for paraneoplastic effects, such as
weight loss and new-onset diabetes, will aid in the early diagnosis
of pancreatic cancer and improve the quality of life of the
patients with the disease.
However, the exact sensitivity and specificity of
these discovered biomarkers for disease prescreening in diagnosed
individuals remains uncertain. For instance, the amount of
GCP1+ circulating exosomes detectable in blood that may
be needed to provide enough information for the diagnosis and
prognosis of pancreatic cancer remains unknown. The paper published
by Costa-Silva et al (8)
elaborated on organotropic metastasis mechanisms and emphasized the
vital role of exosomes in facilitating cancer metastasis.
Furthermore, another recent study demonstrated that pancreatic
cancer-derived exosomes could result in increasing liver metastatic
load by delivering MIF+ exosomes to Kupffer cells and
attracting inflammatory cells to elicit the liver pre-metastatic
niche formation (84). More studies
that further evaluate the association between levels of
MIF+ exosomes and the progression and diagnosis of the
disease are needed. Finally, compared with subjects whose
pancreatic cancer has not progressed, exosomal integrin αvβ5 was
markedly upregulated in exosomes from patients who eventually
developed distant liver metastasis. These findings indicated that
exosomal integrin αvβ5 and MIF orchestrated organ-specific
metastasis and might be reliable biomarkers for the diagnosis of
pancreatic cancer and metastatic stage. Particularly, strategies
targeting these molecular components could be utilized to prevent
organotropic metastasis.
In conclusion, a better understanding of the
molecular mechanisms that initiate pre-metastatic niche formation
and an evaluation of their features and functions in cancer
metastasis will provide novel insights for the early diagnosis and
may lead to therapeutic regimens for the treatment of pancreatic
cancer.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
Not applicable.
Author's contributions
YY performed the literature search and was a major
contributor towards writing the manuscript. GF contributed towards
writing and critically revising the manuscript and for important
intellectual content. LM made contributions to the conception and
design of the present review. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
MIF
|
migration inhibitory factor
|
|
TLR4
|
toll-like receptor 4
|
|
DCs
|
dendritic cells
|
|
CIKs
|
cytokine-induced killer cells
|
|
CAFs
|
cancer-associated fibroblast
|
|
IAP
|
inhibitors of apoptosis
|
|
CP
|
chronic pancreatitis
|
|
GPC1
|
glypican-1
|
|
IPMN
|
intraductal papillary mucinous
neoplasm
|
|
CA19-9
|
carbohydrate antigen 19-9
|
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Iacobuzio-Donahue CA: Genetic evolution of
pancreatic cancer: Lessons learnt from the pancreatic cancer genome
sequencing project. GUT. 61:1085–1094. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yachida S, Jones S, Bozic I, Antal T,
Leary R, Fu B, Kamiyama M, Hruban RH, Eshleman JR, Nowak MA, et al:
Distant metastasis occurs late during the genetic evolution of
pancreatic cancer. Nature. 467:1114–1117. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
György B, Szabó TG, Pásztói M, Pál Z,
Misják P, Aradi B, László V, Pállinger E, Pap E, Kittel A, et al:
Membrane vesicles, current state-of-the-art: Emerging role of
extracellular vesicles. Cell Mol Life Sci. 68:2667–2688. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Simons M and Raposo G: Exosomes-vesicular
carriers for intercellular communication. Curr Opin Cell Biol.
21:575–581. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cocucci E and Meldolesi J: Ectosomes and
exosomes: Shedding the confusion between extracellular vesicles.
Trends Cell Biol. 25:364–372. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cocucci E and Meldolesi J: Ectosomes. Curr
Biol. 21:R940–R941. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Costa-Silva B, Aiello NM, Ocean AJ, Singh
S, Zhang H, Thakur BK, Becker A, Hoshino A, Mark MT, Molina H, et
al: Pancreatic cancer exosomes initiate pre-metastatic niche
formation in the liver. Nat Cell Biol. 17:816–826. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Christianson HC, Svensson KJ, van
Kuppevelt TH, Li JP and Belting M: Cancer cell exosomes depend on
cell-surface heparan sulfate proteoglycans for their
internalization and functional activity. Proc Natl Acad Sci USA.
110:17380–17385. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Miyanishi M, Tada K, Koike M, Uchiyama Y,
Kitamura T and Nagata S: Identification of Tim4 as a
phosphatidylserine receptor. Nature. 450:435–439. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hoshino A, Costa-Silva B, Shen TL,
Rodrigues G, Hashimoto A, Mark Tesic M, Molina H, Kohsaka S, Di
Giannatale A, Ceder S, et al: Tumour exosome integrins determine
organotropic metastasis. Nature. 527:329–335. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Nazarenko I, Rana S, Baumann A, McAlear J,
Hellwig A, Trendelenburg M, Lochnit G, Preissner KT and Zöller M:
Cell surface tetraspanin Tspan8 contributes to molecular pathways
of exosome-induced endothelial cell activation. Cancer Res.
70:1668–1678. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zöller M: Exosomes in cancer disease.
Methods Mol Biol. 1381:111–149. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li Y, Zheng Q, Bao C, Li S, Guo W, Zhao J,
Chen D, Gu J, He X and Huang S: Circular RNA is enriched and stable
in exosomes: A promising biomarker for cancer diagnosis. Cell Res.
25:981–984. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ge Q, Zhou Y, Lu J, Bai Y, Xie X and Lu Z:
miRNA in plasma exosome is stable under different storage
conditions. Molecules. 19:1568–1575. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Subra C, Grand D, Laulagnier K, Stella A,
Lambeau G, Paillasse M, De Medina P, Monsarrat B, Perret B,
Silvente-Poirot S, et al: Exosomes account for vesicle-mediated
transcellular transport of activatable phospholipases and
prostaglandins. J Lipid Res. 51:2105–2120. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Beloribi S, Ristorcelli E, Breuzard G,
Silvy F, Bertrand-Michel J, Beraud E, Verine A and Lombardo D:
Exosomal lipids impact notch signaling and induce death of human
pancreatic tumoral SOJ-6 cells. PLoS One. 7:e474802012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Beloribi-Djefaflia S, Siret C and Lombardo
D: Exosomal lipids induce human pancreatic tumoral MiaPaCa-2 cells
resistance through the CXCR4-SDF-1α signaling axis. Oncoscience.
2:15–30. 2014.PubMed/NCBI
|
|
19
|
Llorente A, Skotland T, Sylvänne T,
Kauhanen D, Róg T, Orłowski A, Vattulainen I, Ekroos K and Sandvig
K: Molecular lipidomics of exosomes released by PC-3 prostate
cancer cells. Biochim Biophys Acta. 1831:1302–1309. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Minciacchi VR, You S, Spinelli C, Morley
S, Zandian M, Aspuria PJ, Cavallini L, Ciardiello C, Sobreiro Reis
M, Morello M, et al: Large oncosomes contain distinct protein cargo
and represent a separate functional class of tumor-derived
extracellular vesicles. Oncotarget. 6:11327–11341. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Keerthikumar S, Gangoda L, Liem M, Fonseka
P, Atukorala I, Ozcitti C, Mechler A, Adda CG, Ang CS and
Mathivanan S: Proteogenomic analysis reveals exosomes are more
oncogenic than ectosomes. Oncotarget. 6:15375–15396. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lötvall J, Hill AF, Hochberg F, Buzás EI,
Di Vizio D, Gardiner C, Gho YS, Kurochkin IV, Mathivanan S,
Quesenberry P, et al: Minimal experimental requirements for
definition of extracellular vesicles and their functions: A
position statement from the International Society for Extracellular
Vesicles. J Extracell Vesicles. 3:269132014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zöller M: Tetraspanins: Push and pull in
suppressing and promoting metastasis. Nat Rev Cancer. 9:40–55.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Pols MS and Klumperman J: Trafficking and
function of the tetraspanin CD63. Exp Cell Res. 315:1584–1592.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yue S, Mu W, Erb U and Zöller M: The
tetraspanins CD151 and Tspan8 are essential exosome components for
the crosstalk between cancer initiating cells and their
surrounding. Oncotarget. 6:2366–2384. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Henderson MC and Azorsa DO: The genomic
and proteomic content of cancer cell-derived exosomes. Front Oncol.
2:382012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Raimondo F, Morosi L, Chinello C, Magni F
and Pitto M: Advances in membranous vesicle and exosome proteomics
improving biological understanding and biomarker discovery.
Proteomics. 11:709–720. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Runz S, Keller S, Rupp C, Stoeck A, Issa
Y, Koensgen D, Mustea A, Sehouli J, Kristiansen G and Altevogt P:
Malignant ascites-derived exosomes of ovarian carcinoma patients
contain CD24 and EpCAM. Gynecol Oncol. 107:563–571. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Al-Nedawi K, Meehan B, Micallef J, Lhotak
V, May L, Guha A and Rak J: Intercellular transfer of the oncogenic
receptor EGFRvIII by microvesicles derived from tumour cells. Nat
Cell Biol. 10:619–624. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Corcoran C, Rani S, O'Brien K, O'Neill A,
Prencipe M, Sheikh R, Webb G, McDermott R, Watson W, Crown J and
O'Driscoll L: Docetaxel-resistance in prostate cancer: Evaluating
associated phenotypic changes and potential for resistance transfer
via exosomes. PLoS One. 7:e509992012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Beckler Demory M, Higginbotham JN,
Franklin JL, Ham AJ, Halvey PJ, Imasuen IE, Whitwell C, Li M,
Liebler DC and Coffey RJ: Proteomic analysis of exosomes from
mutant KRAS colon cancer cells identifies intercellular transfer of
mutant KRAS. Mol Cell Proteomics. 12:343–355. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ji H, Greening DW, Barnes TW, Lim JW,
Tauro BJ, Rai A, Xu R, Adda C, Mathivanan S, Zhao W, et al:
Proteome profiling of exosomes derived from human primary and
metastatic colorectal cancer cells reveal differential expression
of key metastatic factors and signal transduction components.
Proteomics. 13:1672–1686. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Park JA, Sharif AS, Tschumperlin DJ, Lau
L, Limbrey R, Howarth P and Drazen JM: Tissue factor-bearing
exosome secretion from human mechanically stimulated bronchial
epithelial cells in vitro and in vivo. J Allergy Clin Immunol.
130:1375–1383. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Adamczyk KA, Klein-Scory S, Tehrani MM,
Warnken U, Schmiegel W, Schnölzer M and Schwarte-Waldhoff I:
Characterization of soluble and exosomal forms of the EGFR released
from pancreatic cancer cells. Life Sci. 89:304–312. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Salido-Guadarrama I, Romero-Cordoba S,
Peralta-Zaragoza O, Hidalgo-Miranda A and Rodríguez-Dorantes M:
MicroRNAs transported by exosomes in body fluids as mediators of
intercellular communication in cancer. Onco Targets Ther.
7:1327–1338. 2014.PubMed/NCBI
|
|
36
|
Balaj L, Lessard R, Dai L, Cho YJ, Pomeroy
SL, Breakefield XO and Skog J: Tumour microvesicles contain
retrotransposon elements and amplified oncogene sequences. Nat
Commun. 2:1802011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Gibbings DJ, Ciaudo C, Erhardt M and
Voinnet O: Multivesicular bodies associate with components of miRNA
effector complexes and modulate miRNA activity. Nat Cell Biol.
11:1143–1149. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Medina PP, Nolde M and Slack FJ: OncomiR
addiction in an in vivo model of microRNA-21-induced pre-B-cell
lymphoma. Nature. 467:86–90. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Garzon R, Calin GA and Croce CM: MicroRNAs
in cancer. Annu Rev Med. 60:167–179. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Costinean S, Zanesi N, Pekarsky Y, Tili E,
Volinia S, Heerema N and Croce CM: Pre-B cell proliferation and
lymphoblastic leukemia/high-grade lymphoma in E(mu)-miR155
transgenic mice. Proc Natl Acad Sci USA. 103:7024–7029. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Mikamori M, Yamada D, Eguchi H, Hasegawa
S, Kishimoto T, Tomimaru Y, Asaoka T, Noda T, Wada H, Kawamoto K,
et al: MicroRNA-155 controls exosome synthesis and promotes
gemcitabine resistance in pancreatic ductal adenocarcinoma. Sci
Rep. 7:423392017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Park SM, Gaur AB, Lengyel E and Peter ME:
The miR-200 family determines the epithelial phenotype of cancer
cells by targeting the E-cadherin repressors ZEB1 and ZEB2. Genes
Dev. 22:894–907. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Manier S, Liu CJ, Avet-Loiseau H, Park J,
Shi J, Campigotto F, Salem KZ, Huynh D, Glavey SV, Rivotto B, et
al: Prognostic role of circulating exosomal miRNAs in multiple
myeloma. Blood. 129:2429–2436. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Cortez MA, Bueso-Ramos C, Ferdin J,
Lopez-Berestein G, Sood AK and Calin GA: MicroRNAs in body
fluids-the mix of hormones and biomarkers. Nat Rev Clin Oncol.
8:467–477. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Boeri M, Verri C, Conte D, Roz L, Modena
P, Facchinetti F, Calabrò E, Croce CM, Pastorino U and Sozzi G:
MicroRNA signatures in tissues and plasma predict development and
prognosis of computed tomography detected lung cancer. Proc Natl
Acad Sci USA. 108:3713–3718. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Hansen TB, Jensen TI, Clausen BH, Bramsen
JB, Finsen B, Damgaard CK and Kjems J: Natural RNA circles function
as efficient microRNA sponges. Nature. 495:384–388. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Memczak S, Jens M, Elefsinioti A, Torti F,
Krueger J, Rybak A, Maier L, Mackowiak SD, Gregersen LH, Munschauer
M, et al: Circular RNAs are a large class of animal RNAs with
regulatory potency. Nature. 495:333–338. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Li Q, Shao Y, Zhang X, Zheng T, Miao M,
Qin L, Wang B, Ye G, Xiao B and Guo J: Plasma long noncoding RNA
protected by exosomes as a potential stable biomarker for gastric
cancer. Tumour Biol. 36:2007–2012. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Takahashi K, Yan IK, Kogure T, Haga H and
Patel T: Extracellular vesicle-mediated transfer of long non-coding
RNA ROR modulates chemosensitivity in human hepatocellular cancer.
FEBS Open Bio. 4:458–467. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Mohme M, Riethdorf S and Pantel K:
Circulating and disseminated tumour cells-mechanisms of immune
surveillance and escape. Nat Rev Clin Oncol. 14:155–167. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Yu S, Cao H, Shen B and Feng J:
Tumor-derived exosomes in cancer progression and treatment failure.
Oncotarget. 6:37151–37168. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Nguyen DX, Bos PD and Massagué J:
Metastasis: From dissemination to organ-specific colonization. Nat
Rev Cancer. 9:274–284. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Melo SA, Sugimoto H, O'Connell JT, Kato N,
Villanueva A, Vidal A, Qiu L, Vitkin E, Perelman LT, Melo CA, et
al: Cancer exosomes perform cell-independent microRNA biogenesis
and promote tumorigenesis. Cancer Cell. 26:707–721. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Abd Elmageed ZY, Yang Y, Thomas R, Ranjan
M, Mondal D, Moroz K, Fang Z, Rezk BM, Moparty K, Sikka SC, et al:
Neoplastic reprogramming of patient-derived adipose stem cells by
prostate cancer cell-associated exosomes. Stem Cells. 32:983–997.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Falasca M, Kim M and Casari I: Pancreatic
cancer: Current research and future directions. Biochim Biophys
Acta. 1865:123–132. 2016.PubMed/NCBI
|
|
56
|
Zhang X, Yuan X, Shi H, Wu L, Qian H and
Xu W: Exosomes in cancer: Small particle, big player. J Hematol
Oncol. 8:832015. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Hong BS, Cho JH, Kim H, Choi EJ, Rho S,
Kim J, Kim JH, Choi DS, Kim YK, Hwang D and Gho YS: Colorectal
cancer cell-derived microvesicles are enriched in cell
cycle-related mRNAs that promote proliferation of endothelial
cells. BMC Genomics. 10:5562009. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Le MT, Hamar P, Guo C, Basar E,
Perdigão-Henriques R, Balaj L and Lieberman J: miR-200-containing
extracellular vesicles promote breast cancer cell metastasis. J
Clin Invest. 124:5109–5128. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Qu JL, Qu XJ, Zhao MF, Teng YE, Zhang Y,
Hou KZ, Jiang YH, Yang XH and Liu YP: Gastric cancer exosomes
promote tumour cell proliferation through PI3K/Akt and MAPK/ERK
activation. Dig Liver Dis. 41:875–880. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Takikawa T, Masamune A, Yoshida N, Hamada
S, Kogure T and Shimosegawa T: Exosomes derived from pancreatic
stellate cells: MicroRNA signature and effects on pancreatic cancer
cells. Pancreas. 46:19–27. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
An M, Lohse I, Tan Z, Zhu J, Wu J,
Kurapati H, Morgan MA, Lawrence TS, Cuneo KC and Lubman DM:
Quantitative proteomic analysis of serum exosomes from patients
with locally advanced pancreatic cancer undergoing
chemoradiotherapy. J Proteome Res. 16:1763–1772. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Liu Y, Gu Y and Cao X: The exosomes in
tumor immunity. Oncoimmunology. 4:e10274722015. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Zhou M, Chen J, Zhou L, Chen W, Ding G and
Cao L: Pancreatic cancer derived exosomes regulate the expression
of TLR4 in dendritic cells via miR-203. Cell Immunol. 292:65–69.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Ding G, Zhou L, Qian Y, Fu M, Chen J, Chen
J, Xiang J, Wu Z, Jiang G and Cao L: Pancreatic cancer-derived
exosomes transfer miRNAs to dendritic cells and inhibit RFXAP
expression via miR-212-3p. Oncotarget. 6:29877–29888. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Wynn TA, Chawla A and Pollard JW:
Macrophage biology in development, homeostasis and disease. Nature.
496:445–455. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Su MJ, Aldawsari H and Amiji M: Pancreatic
cancer cell exosome-mediated macrophage reprogramming and the role
of MicroRNAs 155 and 125b2 transfection using nanoparticle delivery
systems. Sci Rep. 6:301102016. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Katsiougiannis S, Chia D, Kim Y, Singh RP
and Wong DT: Saliva exosomes from pancreatic tumor-bearing mice
modulate NK cell phenotype and antitumor cytotoxicity. FASEB J.
31:998–1010. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Kamisawa T, Wood LD, Itoi T and Takaori K:
Pancreatic cancer. Lancet. 388:73–85. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Sagar G, Sah RP, Javeed N, Dutta SK, Smyrk
TC, Lau JS, Giorgadze N, Tchkonia T, Kirkland JL, Chari ST and
Mukhopadhyay D: Pathogenesis of pancreatic cancer exosome-induced
lipolysis in adipose tissue. Gut. 65:1165–1174. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Javeed N, Sagar G, Dutta SK, Smyrk TC, Lau
JS, Bhattacharya S, Truty M, Petersen GM, Kaufman RJ, Chari ST and
Mukhopadhyay D: Pancreatic cancer-derived exosomes cause
paraneoplastic β-cell dysfunction. Clin Cancer Res. 21:1722–1733.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Patel GK, Patton MC, Singh S, Khushman M
and Singh AP: Pancreatic cancer exosomes: Shedding off for a
meaningful journey. Pancreat Disord Ther. 6:e1482016. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Oettle H, Post S, Neuhaus P, Gellert K,
Langrehr J, Ridwelski K, Schramm H, Fahlke J, Zuelke C, Burkart C,
et al: Adjuvant chemotherapy with gemcitabine vs observation in
patients undergoing curative-intent resection of pancreatic cancer:
A randomized controlled trial. JAMA. 297:267–277. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Wei Y, Lai X, Yu S, Chen S, Ma Y, Zhang Y,
Li H, Zhu X, Yao L and Zhang J: Exosomal miR-221/222 enhances
tamoxifen resistance in recipient ER-positive breast cancer cells.
Breast Cancer Res Treat. 147:423–431. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Richards KE, Zeleniak AE, Fishel ML, Wu J,
Littlepage LE and Hill R: Cancer-associated fibroblast exosomes
regulate survival and proliferation of pancreatic cancer cells.
Oncogene. 36:1770–1778. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Valenzuela Asuncion MM, Castro I, Gonda A,
Osterman Diaz CJ, Jutzy JM, Aspe JR, Khan S, Neidigh JW and Wall
NR: Cell death in response to antimetabolites directed at
ribonucleotide reductase and thymidylate synthase. Onco Targets
Ther. 8:495–507. 2015.PubMed/NCBI
|
|
76
|
Valenzuela MM, Bennit Ferguson HR, Gonda
A, Osterman Diaz CJ, Hibma A, Khan S and Wall NR: Exosomes secreted
from human cancer cell lines contain Inhibitors of Apoptosis (IAP).
Cancer Microenviron. 8:65–73. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Que R, Ding G, Chen J and Cao L: Analysis
of serum exosomal microRNAs and clinicopathologic features of
patients with pancreatic adenocarcinoma. World J Surg Oncol.
11:2192013. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Lu L and Risch HA: Exosomes: Potential for
early detection in pancreatic cancer. Future Oncol. 12:1081–1090.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Joshi GK, Deitz-McElyea S, Liyanage T,
Lawrence K, Mali S, Sardar R and Korc M: Label-Free
nanoplasmonic-based short noncoding RNA sensing at attomolar
concentrations allows for quantitative and highly specific assay of
MicroRNA-10b in biological fluids and circulating exosomes. ACS
Nano. 9:11075–11089. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Madhavan B, Yue S, Galli U, Rana S, Gross
W, Müller M, Giese NA, Kalthoff H, Becker T, Büchler MW and Zöller
M: Combined evaluation of a panel of protein and miRNA
serum-exosome biomarkers for pancreatic cancer diagnosis increases
sensitivity and specificity. Int J Cancer. 136:2616–2627. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Melo SA, Luecke LB, Kahlert C, Fernandez
AF, Gammon ST, Kaye J, LeBleu VS, Mittendorf EA, Weitz J, Rahbari
N, et al: Glypican-1 identifies cancer exosomes and detects early
pancreatic cancer. Nature. 523:177–182. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Ben Q, Xu M, Ning X, Liu J, Hong S, Huang
W, Zhang H and Li Z: Diabetes mellitus and risk of pancreatic
cancer: A meta-analysis of cohort studies. Eur J Cancer.
47:1928–1937. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Huxley R, Ansary-Moghaddam A, de González
Berrington A, Barzi F and Woodward M: Type-II diabetes and
pancreatic cancer: A meta-analysis of 36 studies. Br J Cancer.
92:2076–2083. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Liu Y and Cao X: Organotropic metastasis:
Role of tumor exosomes. Cell Res. 26:149–150. 2016. View Article : Google Scholar : PubMed/NCBI
|














