Introduction
Gastric cancer (GC) is the second most commonly
diagnosed cancer in men and fourth most commonly diagnosed cancer
in women, and is the second-leading cause of cancer-associated
mortality in China (1). Although the
incidence and mortality rates of GC have significantly decreased, a
large number of newly diagnosed cases have occurred as a result of
population growth and the ageing population (1). Tumor cell metastasis is a major cause of
GC-associated mortality and markedly decreases the efficacy of
treatment (2). An increasing number
of oncogenes and tumor suppressors were observed to be involved in
the progression of GC (2); however,
the mechanism underlying GC metastasis remains largely unclear.
Long non-coding RNAs (lncRNAs), a class of
non-coding RNAs that do not code for proteins, are >200
nucleotides in length (3). The
aberrant expression of lncRNAs is believed to be associated with
the aggressive development of cancer (4–6). It is
well known that lncRNAs participate in cancer-associated pathways
(7). Although an increasing number of
lncRNAs have been reported to functionally regulate the metastatic
behavior of GC cells, a large number of lncRNAs require
identification, as 70–90% of the human genome transcribes RNA
products, whereas protein-coding genes account for only 2%
(8). Therefore, the present study
focused on an lncRNA, AFAP1-antisense RNA 1 (AFAP1-AS1), which was
first observed in esophageal cancer and acted as an oncogene,
mediating effects in cell proliferation, migration and invasion
(9). AFAP1-AS1 was also revealed to
serve a crucial role in various types of cancer, including
colorectal (10), breast (11), hepatocellular (12) and lung cancer (13). However, the expression and function of
AFAP1-AS1 in the development of GC remains largely unknown.
The epithelial-mesenchymal transition (EMT) serves
essential roles in carcinogenesis and tumor progression (14). The determinant step in EMT is local
invasion through the epithelial basement membrane that modifies
cell-cell and cell-matrix interactions (15). A previous study demonstrated that
lncRNA also regulates the progress of EMT in GC (16); AFAP1-AS1 was revealed to participate
in the progression of EMT and to facilitate metastasis in
colorectal cancer (17). However, it
is unclear whether AFAP1-AS1 is associated with the progression of
EMT in GC.
Materials and methods
Patients and samples
All samples used in the present study were collected
from the General Hospital of Chinese People's Liberation Army
(Beijing, China) according to the protocols approved by the Ethics
Review Board and written informed consent was obtained from all
patients. A total of 80 pairs of tumor and matched normal tissues
were included and these samples were registered between April 2011
and September 2013. The characterization of the samples is
presented in Table I. The 36-month
follow-ups of the 80 GC patients (48 male, 32 female; median age,
58 years; age range, 32–76 years) was performed.
 | Table I.The clinical characterization of
patients with GC. |
Table I.
The clinical characterization of
patients with GC.
| Variable | Patients, n (%) |
|---|
| Sex |
|
| Male | 48 (60) |
|
Female | 32 (40) |
| Age, years |
|
| ≥60 | 42 (52.5) |
| ≤60 | 38 (47.5) |
| Tumor location |
|
| Body | 22 (27.5) |
|
Antrum | 31 (38.8) |
|
Cardia | 27 (33.7) |
|
Other | 0 |
| Histology |
|
|
Adenocarcinoma | 52 (65) |
| Mucinous
adenocarcinoma | 28 (35) |
| Signet
cell cancer | 0 |
| TNM stage |
|
| I | 18 (22.5) |
| II | 23 (28.7) |
| III | 26 (32.5) |
| IV | 13 (16.3) |
Cell culture
GC GES-1, AGS, BGC-823, MKN-45 and SGC-7901 cell
lines, which were purchased from the American Type Culture
Collection (Manassas, VA, USA), were used in the present study. AGS
cells were cultured in F-12 medium (Hyclone; GE Healthcare,
Chicago, IL, USA) supplemented with 10% fetal bovine serum (FBS;
Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA), and
GES-1, BGC-823, MKN-45 and SGC-7901 cells were cultured in
High-Glucose Dulbecco's modified Eagle's medium (DMEM),
supplemented with 10% FBS at 37°C in 5% CO2.
Lentivirus infection
Two short hairpin RNAs (shRNAs), shRNA1 and shRNA2,
which are vectors of AFAP1-AS1, were constructed and further
packaged into lentiviruses by Shanghai GenePharma Co., Ltd.,
(Shanghai, China). A total of 108 titer lentiviruses
were obtained. Next, MKN-45 and SGC-7901 cells were infected with
the lentiviruses at a multiplicity of infection at a ratio of 1
cell: 20 lentiviruses. The lentiviruses were added into the cell
culture and softly shaken. A total of 48 h after infection, the
decreased expression of AFAP1-AS1 regulated by the shRNAs were
confirmed using reverse transcription-quantitative polymerase chain
reaction (RT-qPCR), compared with a negative control.
RNA isolation
TRIzol reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) was used to extract the total RNA from frozen
tissues and GC cells, according to the manufacturer's protocol. In
brief, 1 mg tissue or 1×106 cells were mixed with 1 ml
TRIzol reagent and 60 µl nuclease-free H2O was added to
re-suspend the RNA precipitation.
RT-qPCR
RT-qPCR was performed in two steps using two
reaction kits, the PrimeScript RT reagent kit (Takara Biotechnology
Co., Ltd., Dalian, China) and the Premix Ex Taq kit (Takara
Biotechnology Co., Ltd.), according to the manufacturer's
protocols. In the first step, cDNA was synthesized in 20 µl volumes
containing 10 µl total RNA with genomic DNA removed, 4 µl 5X
reverse transcription buffer, 1 µl Prime RT Enzyme, 1 µl RT Primer
mix and 4 µl nuclease-free H2O. In the second step, qPCR
reactions were performed in 20 µl volumes and run on a Bio-Rad IQ5
thermocycler (Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Thermocycling conditions were as follows: 94°C for 2 min, followed
by 40 cycles of 95°C for 15 sec and 60°C for 30 sec. The primer
sequences were as follows: AFAP1-AS1 forward,
5′-TCGCTCAATGGAGTGACGGCA-3′ and reverse,
5′-CGGCTGAGACCGCTGAGAACTT-3′; GAPDH forward,
5′-GGTGGTCTCCTCTGACTTCAACA-3′ and reverse,
5′-TCTCTTCCTCTTGTGCTCTTGCT-3′. GAPDH mRNA was used as an internal
reference. The expression of AFAP1-AS1 was analyzed using the
2−∆∆Cq method (18).
Proliferation assays
As MKN-45 and SGC-7901 cells had a high expression
of AFAP1-AS1, these cells were used to analyze the loss-of-function
of AFAP1-AS1. Then, these cells were seeded into 96-well pates at a
density of 2,000 cells/well, respectively. The cell proliferation
was analyzed using a Cell Counting kit-8 (CCK8; Beyotime Institute
of Biotechnology, Haimen, China) at 0, 24, 48, 72 and 96 h,
according to the manufacturer's protocols. The results were
collected using an ultraviolet spectrophotometer (Thermo Fisher
Scientific, Inc.) at 450 nm.
Migration and invasion assays
AFAP1-AS1-silenced MKN-45 and SGC-7901 cells at
density of 5×104, and the same number of the parallel
control cells, were used to perform migration or invasion assays.
The migration and invasion of the cells were analyzed using a QCM
Laminin Migration Assay kit (cat. no. ECM220; Merck KGaA,
Darmstadt, Germany) and a Cell Invasion Assay kit (cat. no. ECM550;
Merck KGaA), according to the manufacturer's protocols. FBS-free
DMEM was used in the upper chamber and DMEM with 10% FBS was used
in the lower chamber. The number of cells transferred to the lower
chamber were calculated using a light microscope (Olympus
Corporation, Tokyo, Japan). The magnification was ×10.
Western blot analysis
SGC-7901 cells with lentivirus containing negative
control or shRNA-1 of lncRNA AFAP1-AS1, were seeded into 6-well
plates prior to being collected for western blot analysis. The
total proteins were extracted using RIPA buffer reagent (Thermo
Fisher Scientific, Inc.), and the concentration of the lysate was
determined using a BCA kit (Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol. A total of 20 µg of total
proteins were separated using 12% SDS-PAGE gels and then
transferred onto polyvinylidene fluoride (PVDF) membranes. Bovine
serum albumin (3%) solution (Beyotime Institute of Biotechnology)
was used to block the PVDF membranes for 1 h at room temperature.
Primary antibodies, E-cadherin (1:200; cat. no. 14472), N-cadherin
(1:200; cat. no. 14215), GAPDH (1:1,000; cat. no. 5174) and
vimentin (1:500; cat. no. 5741) were purchased from Cell Signaling
Technology, Inc. (Danvers, MA, USA) and were diluted using the
Primary Antibody Dilution Buffer (Beyotime Institute of
Biotechnology) according to the manufacturer's protocols. The PVDF
membranes were incubated with the antibodies overnight at 4°C. The
secondary antibodies, HRP goat anti-mouse (cat. no. TA130003;
1:10,000) and HRP goat anti-rabbit (cat. no. TA140003; 1:10,000)
(OriGene Technologies, Inc., Rockville, MD, USA) were diluted with
the blocking solution (above mentioned) and then incubated with the
PVDF membranes at room temperature for 1 h. Finally, bound proteins
were visualized using a SuperSignal West Dura Extended Duration
Substrate kit (Thermo Fisher Scientific, Inc.) and quantified using
the ChemiDoc™ and ChemiDoc MP Imaging Systems with Image
Lab™ Touch Software (Version 2.0) (Bio-Rad, California,
USA).
Statistical analysis
All data were presented as mean ± standard
deviation. A two-tailed Student's t-test was used to compare the
differences between two groups. One-way analysis of variance,
followed by the Student-Newman-Keuls test, was used to analyze
differences between more than two groups. Receiver operating
characteristic (ROC) curve analysis was used for evaluating the
effect of AFAP1-AS1 on differing GC from the adjacent normal
tissues. The Youden index was used to analyze the optimal cut-off
value. Survival analysis was used to evaluate the role of the
expression of lncRNA AFAP1-AS1 in the survival time of patients
with GC. P<0.05 was considered to indicate a statistically
significant difference. SPSS 17.0 (SPSS, Inc., Chicago, IL, USA)
and GraphPad Prism 6.0 (GraphPad Software, Inc., La Jolla, CA, USA)
software were used to perform all statistical analyses and to
generate the graphs.
Results
AFAP1-AS1 expression is increased in
the primary tumor tissue of GC patients
The expression of AFAP1-AS1 was increased in various
types of cancer (10,12,17);
however, its expression in GC was unclear. The expression of
AFAP1-AS1 in 80 pairs of primary GC tissues and matched normal
tissues was detected. The results revealed that the expression of
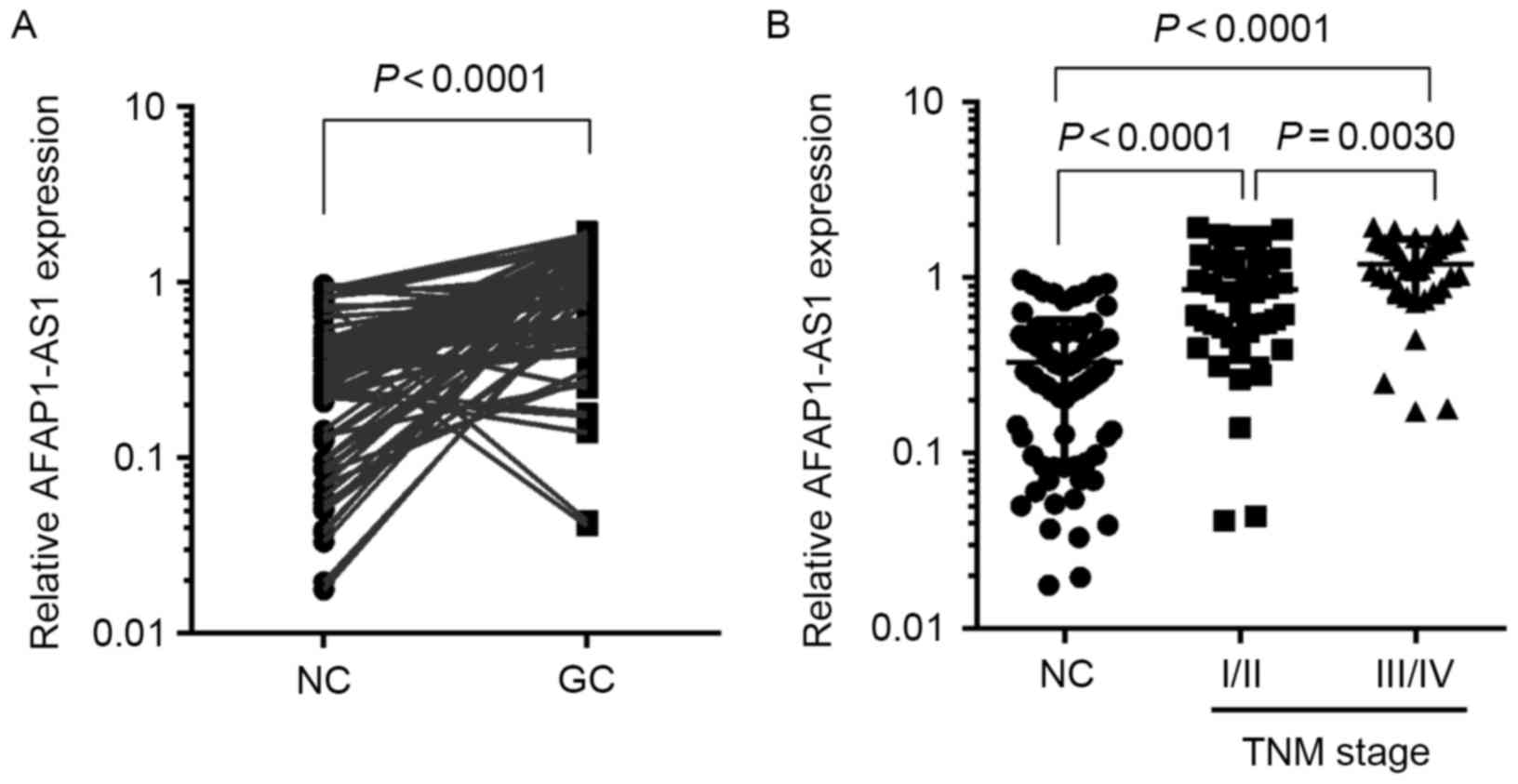
AFAP1-AS1 was significantly higher in GC tissues than in the
matched normal tissues (P<0.01; Fig.
1A). Furthermore, the expression of AFAP1-AS1 was significantly
increased in the tumor tissues of GC patients with
Tumor-Node-Metastasis (TNM) stages (19) III and IV (P<0.01; Fig. 1B), indicating that the overexpression
of AFAP1-AS1 may be associated with the aggressive progression of
GC.
High expression of AFAP1-AS1 is
associated with shorter survival times of patients with GC
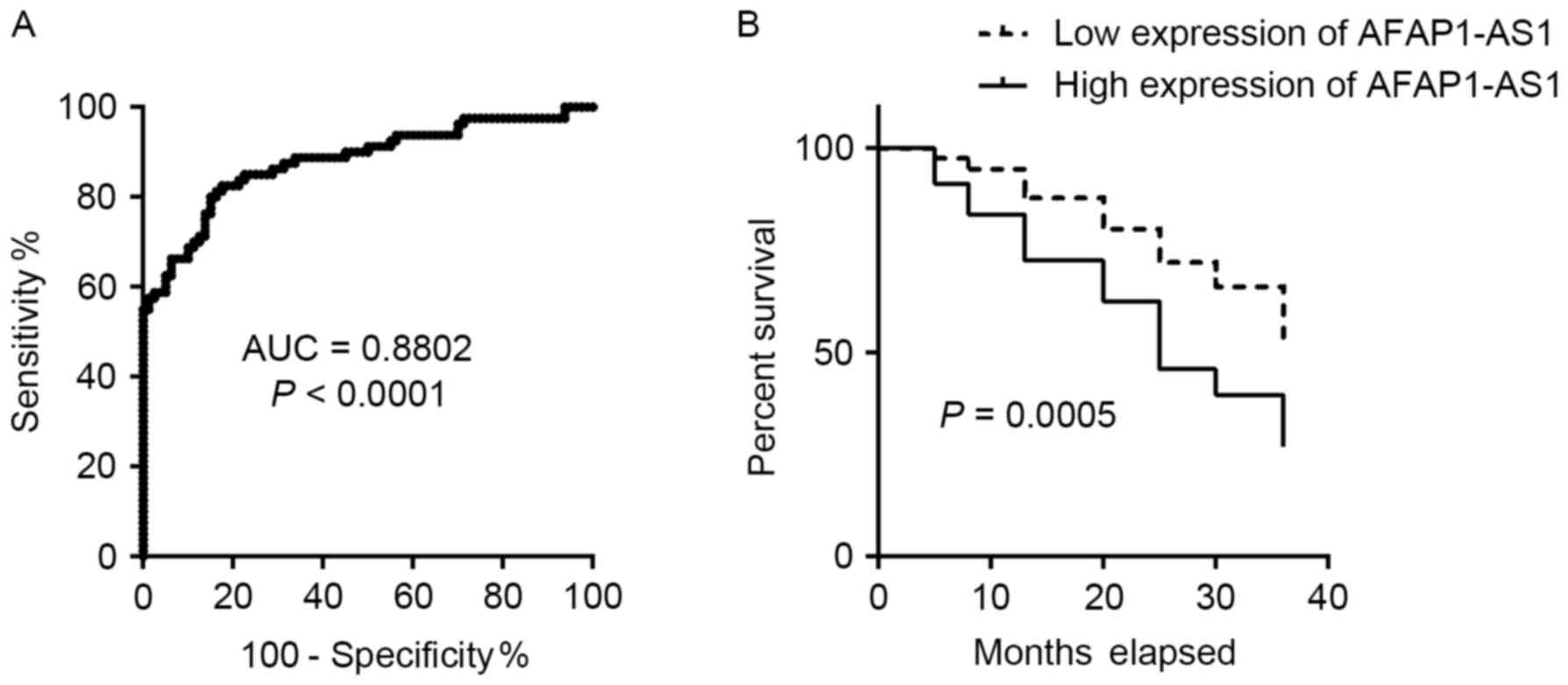
To determine whether AFAP1-AS1 expression signature
could serve as a biomarker for GC detection, ROC curve analysis was
performed. The results revealed that the expression signature of
AFAP1-AS1 had potential as a diagnostic marker of GC when an
optimal cut-off value of 0.5040 was selected (area under the curve,
0.8802; sensitivity, 81.25%; specificity, 83.75%; Fig. 2A). The association between the
expression of AFAP1-AS1 and the survival time of patients with GC
was also analyzed. As depicted in Fig.
2B, patients with GC that expressed high levels of AFAP1-AS1
exhibited a shorter survival time compared with those expressing
low levels of AFAP1-AS1. These results indicated that the
expression signature of AFAP1-AS1 might be a potential biomarker
for the diagnosis and prognosis of GC.
Downregulation of AFAP1-AS1 is induced
by specific shRNAs
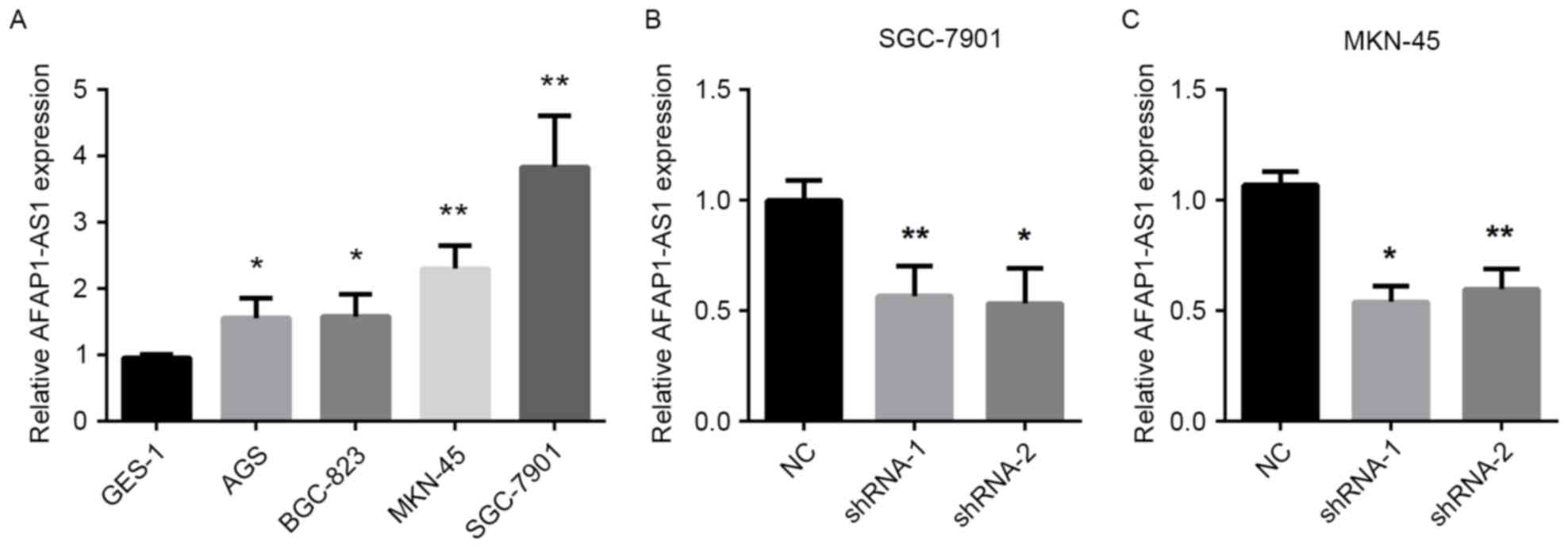
To determine the biological role of AFAP1-AS1 in GC
cells, the expression of AFAP1-AS1 was measured in five GC cells
lines. As demonstrated in Fig. 3A,
the expression of AFAP1-AS1 was significantly increased in SGC-7901
and MKN-45 cells compared with the other GC cells. Therefore,
AFAP1-AS1 downregulation was induced in SGC-7901 and MKN-45 cells
using lentiviruses containing specific shRNA-1 and shRNA-2 which
would target AFAP1-AS1. The results revealed that shRNA-1 and
shRNA-2 significantly decreased the expression of AFAP1-AS1 in the
two cell lines, compared with negative control (NC) (Fig. 3B and C).
Downregulation of AFAP1-AS1 represses
the proliferation, migration and invasion of GC cells in vitro
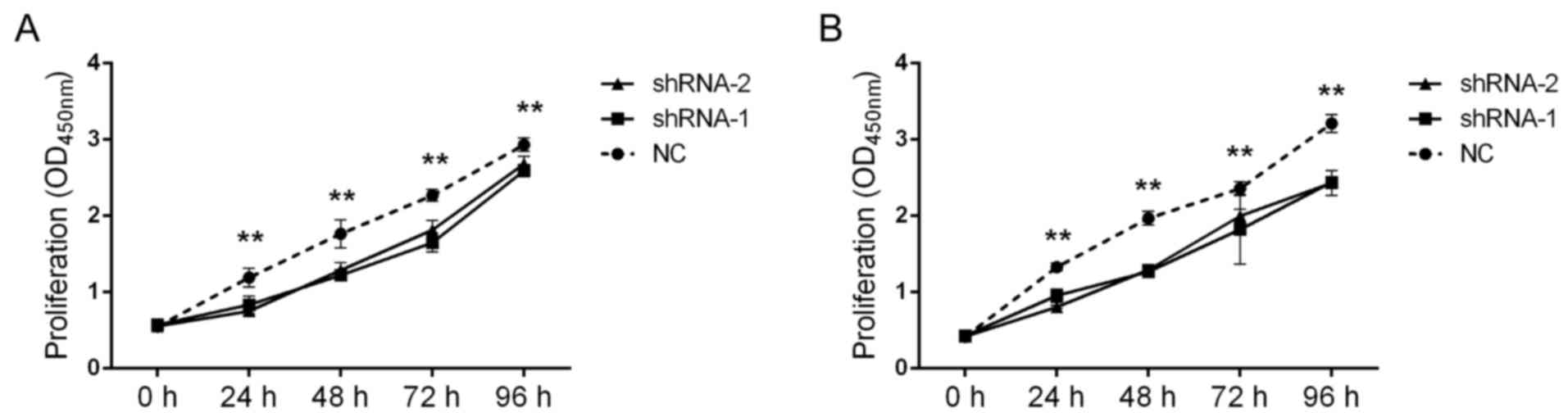
Using the aforementioned constructed cells, the
effect of AFAP1-AS1 on GC cell proliferation was analyzed. The
results revealed that shRNA-1 and shRNA-2 significantly decreased
the proliferation of SGC-7901 and MKN-45 cells at on days 1, 2, 3
and 4 when compared with the negative control (NC) (Fig. 4A and B). Furthermore, the effect of
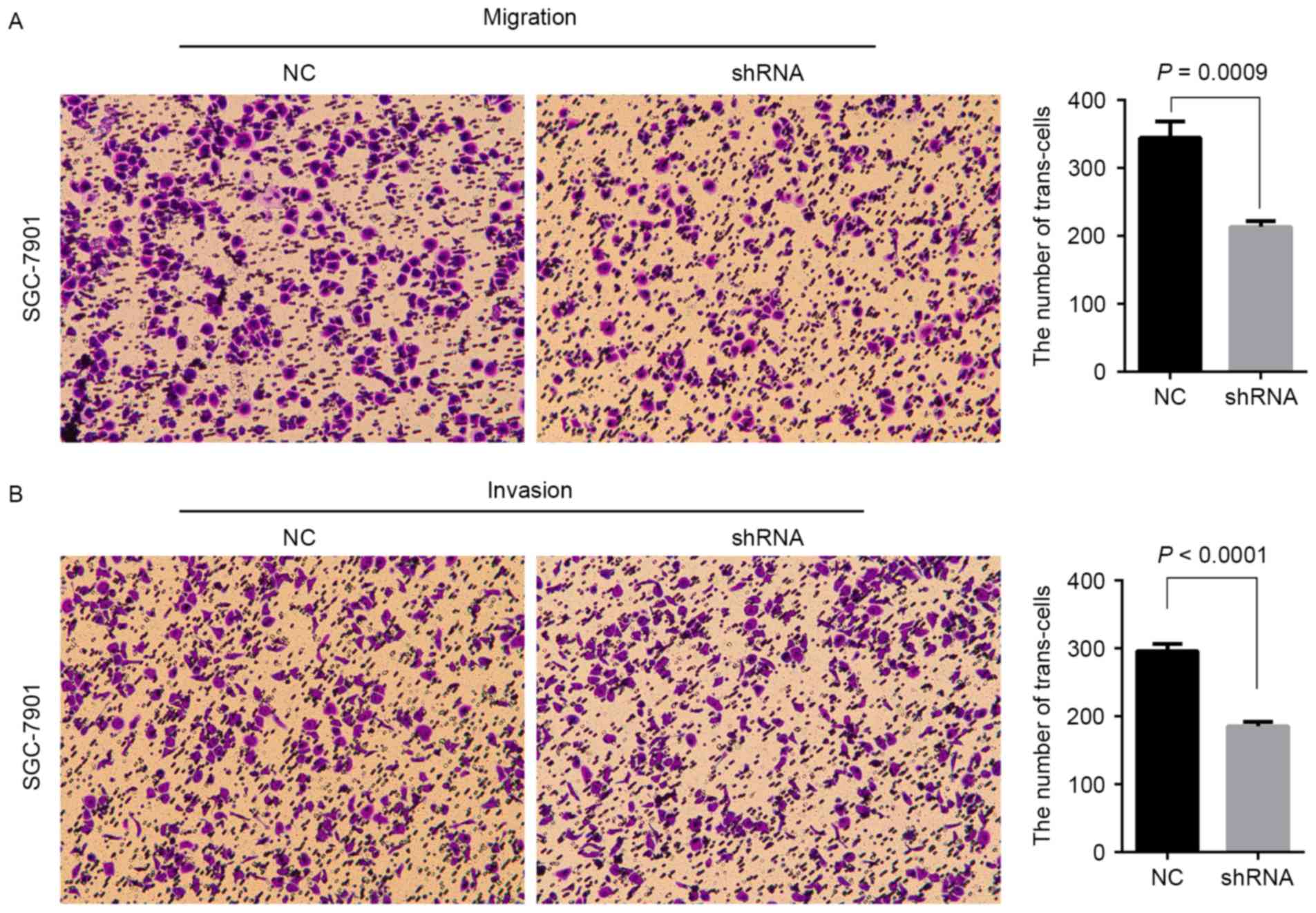
AFAP1-AS1 on the migration and invasion of GC cells was also
analyzed. As demonstrated in Fig. 5A and
B, knockdown of AFAP1-AS1 expression significantly inhibited
the migration of SGC-7901 and MKN-45 cells. These data indicated
that AFAP1-AS1 might promote the proliferation and metastasis of
cells in GC.
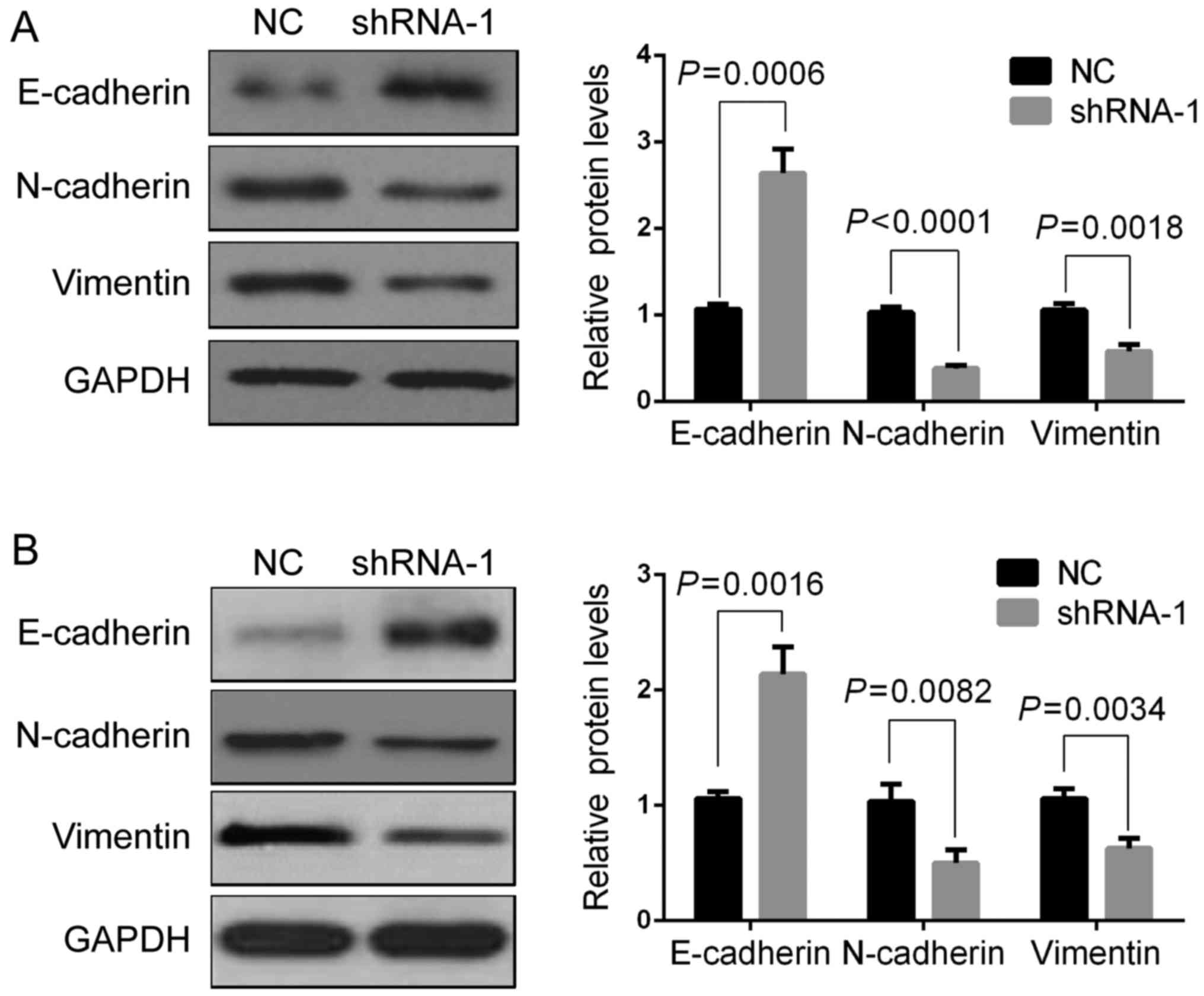
Downregulation of AFAP1-AS1 modulates
the expression of EMT-associated genes
A previous study demonstrated that AFAP1-AS1
functions as an oncogene and promotes GC cell metastasis (20). Therefore, the present study aimed to
determine whether AFAP1-AS1 is involved in regulating
EMT-associated genes, including E-cadherin, N-cadherin and
vimentin. Western blot assays revealed that AFAP1-AS1 knockdown
significantly increased the protein expression of E-cadherin,
whereas the protein expression of N-cadherin and vimentin was
significantly decreased (Fig. 6A and
B). These results indicated that the AFAP1-AS1 induced the
metastasis of GC cells through regulating EMT.
Discussion
The present study revealed that the expression of
AFAP1-AS1 was increased in the GC tissues when compared to the
matched normal tissues, and increased levels were associated with a
shorter survival time in patients with GC. Furthermore, the
expression signature of AFAP-AS1 was revealed to have the ability
to distinguish GC tissues from matched normal tissues. The
biological role of AFAP1-AS1 in GC cells was also determined and
these data revealed that the downregulation of AFAP1-AS1 inhibited
the proliferation and migration of GC cells.
AFAP1-AS1 was first identified in Barrett's
esophagus and esophageal adenocarcinoma; its overexpression
promoted cell proliferation and colony-forming ability, inhibit
apoptosis, and enhance cellular migration and invasion in
esophageal adenocarcinoma (9). The
overexpression of AFAP1-AS1 has also been observed in various types
of cancer, and is involved in the aggressive progression of cancer
(12,21–23).
However, in GC, the expression and function of AFAP1-AS1 remains
largely unclear. The present study detected the expression of
AFAP1-AS1 in GC tissues and matched normal tissues, and observed
that the expression of AFAP1-AS1 was also increased the GC tissues,
a process that is associated with the aggressive progression of GC.
Furthermore, the results of the present study demonstrated that the
overexpression of AFAP1-AS1 was associated with shorter survival
times in patients with GC. These observations indicated that the
expression signature of AFAP1-AS1 may be a potential biomarker for
the prognosis of patients with GC. Evidence has demonstrated that
AFAP1-AS1 serves an oncogenic function in tumor development
(17,24,25). The
results of the present study also indicated that a decrease in
AFAP1-AS1 expression suppressed the proliferation, migration and
invasion of GC cells, indicating that AFAP1-AS1 has a strong
oncogenic function.
E-cadherin, a transmembrane glycoprotein of the type
I cadherin superfamily, is the primary molecule in the adherent
junctions in epithelial cells (14).
The extracellular domain of E-cadherin forms dimers and interacts
with the dimers of E-cadherin on the membrane of a neighboring
cell; its intracellular domain is linked to a protein complex and
is involved in intracellular actin filaments network (14). In contrast to epithelial cells,
mesenchymal cells primarily express N-cadherin, which has a
dominant effect in cell-cell interactions. N-cadherin enhances the
motility of tumor cells via the destabilization of the cell-cell
adhesion complex (26). The present
study revealed that the inhibition of AFAP1-AS1 expression
significantly increased the level of E-cadherin protein expression
and decreased that of N-cadherin in GC cells, indicating that
AFAP1-AS1 participates in the regulation of the EMT progress in GC
cells.
In the present study, the number of available
clinical specimens was relatively small. Although the biological
function of AFAP1-AS1 in GC cells was identified and its mechanism
was investigated, the exact target of AFAP1-AS1 remains largely
unclear. Additionally, the role of AFAP1-AS1 in the progress of GC
requires further clarification in vivo. Additionally, more
clinical samples are required to evaluate the differential ability
of AFAP1-AS1 in the diagnosis of GC, and the exact mechanism of
AFAP1-AS1 in GC cells requires identification.
In conclusion, the present study revealed that the
overexpression of AFAP1-AS1 was associated with the shorter
survival times of patients with GC, and that it promotes the
proliferation, migration and invasion of GC cells, partially
through regulating the EMT process, indicating that AFAP1-AS1 may
serve as a novel biomarker and therapeutic target for the diagnosis
and treatment of GC.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the
National Nature Science Foundation of China (grant nos. 81272698,
81672319 and 81602507).
Availability of data and materials
The datasets generated and analyzed in the present
study are included in the published article.
Authors' contributions
HZ, KZ, and TW performed the experiments. JC, HX,
YW, and YS collected the patient samples. XZ analyzed the data. BW
and LC designed the present study and wrote this paper.
Ethics approval and consent to
participate
All samples used in the present study were collected
from the General Hospital of Chinese People's Liberation Army
(Beijing, China) according to the protocols approved by the Ethics
Review Board and written informed consent was obtained from all
patients.
Consent for publication
Study participants provided consent for the data to
be published.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ushijima T and Sasako M: Focus on gastric
cancer. Cancer Cell. 5:121–125. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
ENCODE Project Consortium: An integrated
encyclopedia of DNA elements in the human genome. Nature.
489:57–74. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Glover AR, Zhao JT, Ip JC, Lee JC,
Robinson BG, Gill AJ, Soon PS and Sidhu SB: Long noncoding RNA
profiles of adrenocortical cancer can be used to predict
recurrence. Endocr Relat Cancer. 22:99–109. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yuan JH, Yang F, Wang F, Ma JZ, Guo YJ,
Tao QF, Liu F, Pan W, Wang TT, Zhou CC, et al: A Long Noncoding RNA
activated by TGF-β promotes the invasion-metastasis cascade in
hepatocellular carcinoma. Cancer Cell. 25:666–681. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Liu B, Sun L, Liu Q, Gong C, Yao Y, Lv X,
Lin L, Yao H, Su F, Li D, et al: A cytoplasmic NF-κB interacting
long noncoding RNA Blocks IκB phosphorylation and suppresses breast
cancer metastasis. Cancer Cell. 27:370–381. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Schmitt AM and Chang HY: Long noncoding
RNAs in cancer pathways. Cancer Cell. 29:452–463. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zeng S, Xiao YF, Tang B, Hu CJ, Xie R,
Yang SM and Li BS: Long noncoding RNA in digestive tract cancers:
function, mechanism and potential biomarker. Oncologist.
20:898–906. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wu W, Bhagat TD, Yang X, Song JH, Cheng Y,
Agarwal R, Abraham JM, Ibrahim S, Bartenstein M, Hussain Z, et al:
Hypomethylation of noncoding DNA regions and overexpression of the
long noncoding RNA, AFAP1-AS1, in Barrett's esophagus and
esophageal adenocarcinoma. Gastroenterology. 144:956–966.e4. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang F, Ni H, Sun F, Li M and Chen L:
Overexpression of lncRNA AFAP1-AS1 correlates with poor prognosis
and promotes tumorigenesis in colorectal cancer. Biomed
Pharmacother. 81:152–159. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yang F, Lyu S, Dong S, Liu Y, Zhang X and
Wang O: Expression profile analysis of long noncoding RNA in
HER-2-enriched subtype breast cancer by next-generation sequencing
and bioinformatics. Onco Targets Ther. 9:761–772. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang JY, Weng MZ, Song FB, Xu YG, Liu Q,
Wu JY, Qin J, Jin T and Xu JM: Long noncoding RNA AFAP1-AS1
indicates a poor prognosis of hepatocellular carcinoma and promotes
cell proliferation and invasion via upregulation of the RhoA/Rac2
signaling. Int J Oncol. 48:1590–1598. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zeng Z, Bo H, Gong Z, Lian Y, Li X, Li X,
Zhang W, Deng H, Zhou M, Peng S, et al: AFAP1-AS1, a long noncoding
RNA upregulated in lung cancer and promotes invasion and
metastasis. Tumour Biol. 37:729–737. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Voulgari A and Pintzas A:
Epithelial-mesenchymal transition in cancer metastasis: mechanisms,
markers and strategies to overcome drug resistance in the clinic.
Biochim Biophys Acta. 1796:75–90. 2009.PubMed/NCBI
|
|
15
|
Mathias RA and Simpson RJ: Towards
understanding epithelial-mesenchymal transition: a proteomics
perspective. Biochim Biophys Acta. 1794:1325–1331. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhou H, Wang F, Chen H, Tan Q, Qiu S, Chen
S, Jing W, Yu M, Liang C, Ye S and Tu J: Increased expression of
long-noncoding RNA ZFAS1 is associated with epithelial mesenchymal
transition of gastric cancer. Aging (Albany NY). 8:2023–2038. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Han X, Wang L, Ning Y, Li S and Wang Z:
Long non-coding RNA AFAP1-AS1 facilitates tumor growth and promotes
metastasis in colorectal cancer. Biol Res. 49:362016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C (T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sano T, Coit DG, Kim HH, Roviello F,
Kassab P, Wittekind C, Yamamoto Y and Ohashi Y: Proposal of a new
stage grouping of gastric cancer for TNM classification:
International gastric cancer association staging project. Gastric
Cancer. 20:217–225. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bo H, Gong Z, Zhang W, Li X, Zeng Y, Liao
Q, Chen P, Shi L, Lian Y, Jing Y, et al: Upregulated long
non-coding RNA AFAP1-AS1 expression is associated with progression
and poor prognosis of nasopharyngeal carcinoma. Oncotarget.
6:20404–20418. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li Q, Dai Y, Wang F and Hou S:
Differentially expressed long non-coding RNAs and the prognostic
potential in colorectal cancer. Neoplasma. 63:977–983. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Bo H, Gong Z, Zhang W, Li X, Zeng Y, Liao
Q, Chen P, Shi L, Lian Y, Jing Y, et al: Upregulated long
non-coding RNA AFAP1-AS1 expression is associated with progression
and poor prognosis of nasopharyngeal carcinoma. Oncotarget.
6:20404–20418. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ye Y, Chen J, Zhou Y, Fu Z, Zhou Q, Wang
Y, Gao W, Zheng S, Zhao X, Chen T and Chen R: High expression of
AFAP1-AS1 is associated with poor survival and short-term
recurrence in pancreatic ductal adenocarcinoma. J Transl Med.
13:1372015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Luo HL, Huang MD, Guo JN, Fan RH, Xia XT,
He JD and Chen XF: AFAP1-AS1 is upregulated and promotes esophageal
squamous cell carcinoma cell proliferation and inhibits cell
apoptosis. Cancer Med. 5:2879–2885. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhou XL, Wang WW, Zhu WG, Yu CH, Tao GZ,
Wu QQ, Song YQ, Pan P and Tong YS: High expression of long
non-coding RNA AFAP1-AS1 predicts chemoradioresistance and poor
prognosis in patients with esophageal squamous cell carcinoma
treated with definitive chemoradiotherapy. Mol Carcinog.
55:2095–2105. 2016. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Nieman MT, Prudoff RS, Johnson KR and
Wheelock MJ: N-cadherin promotes motility in human breast cancer
cells regardless of their E-cadherin expression. J Cell Biol.
147:631–644. 1999. View Article : Google Scholar : PubMed/NCBI
|